
Study on the Synchronous Removal of Nitrogen and Phosphorus by Autotrophic/Heterotrophic Denitrification in the Presence of Pyrite

Abstract

1. Introduction
2. Materials and Methods
2.1. Experimental Reagents and Devices
2.2. Reactor Start-Up and Parameter Optimization
2.3. High Throughput Sequencing and Microbial Diversity Analysis
3. Results and Discussion
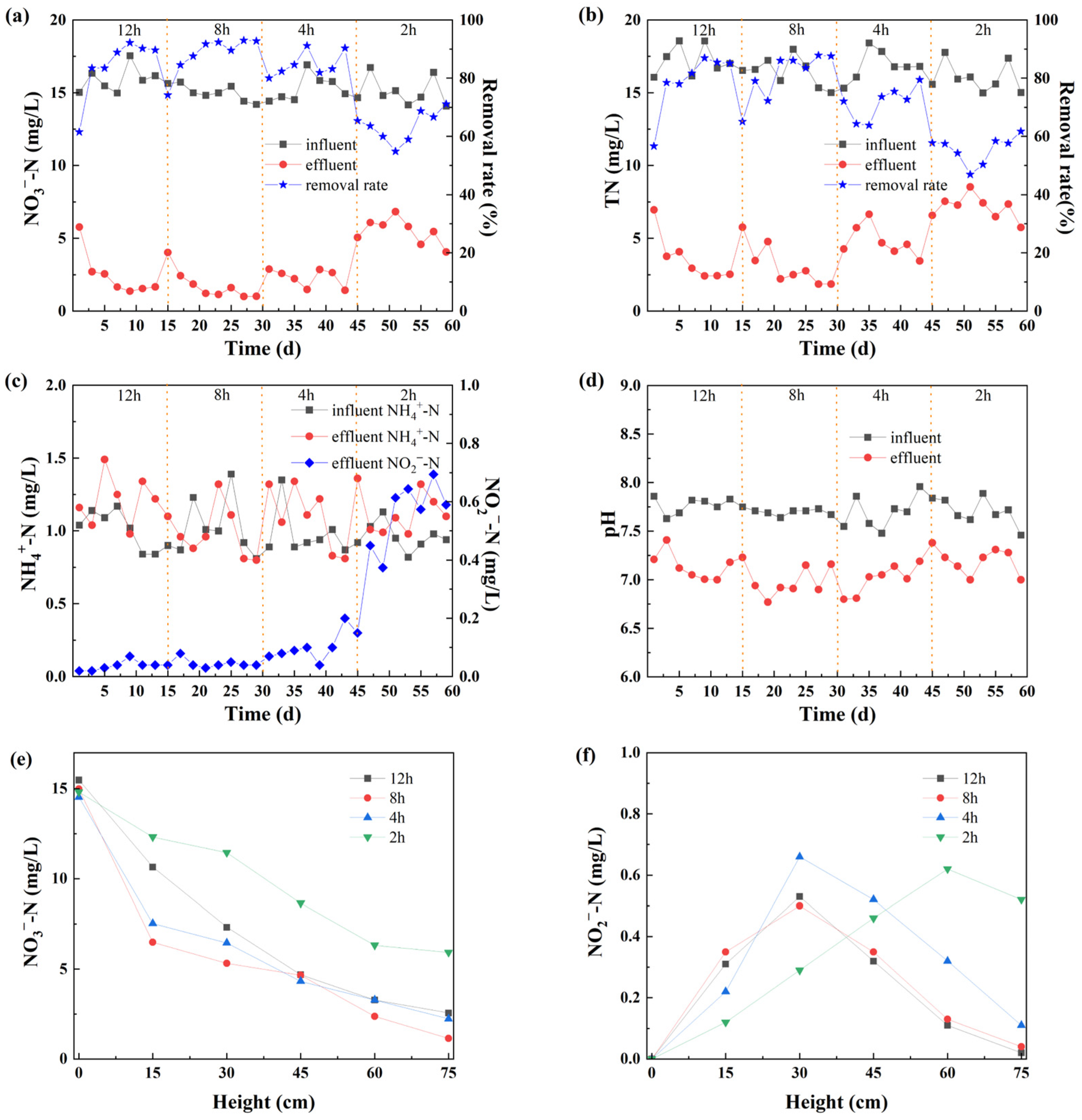
3.1. Impact of EBCT on N Removal
3.1.1. N Removal Rates
3.1.2. Changes of pH Values
3.1.3. Influence of the Height of Medium Level
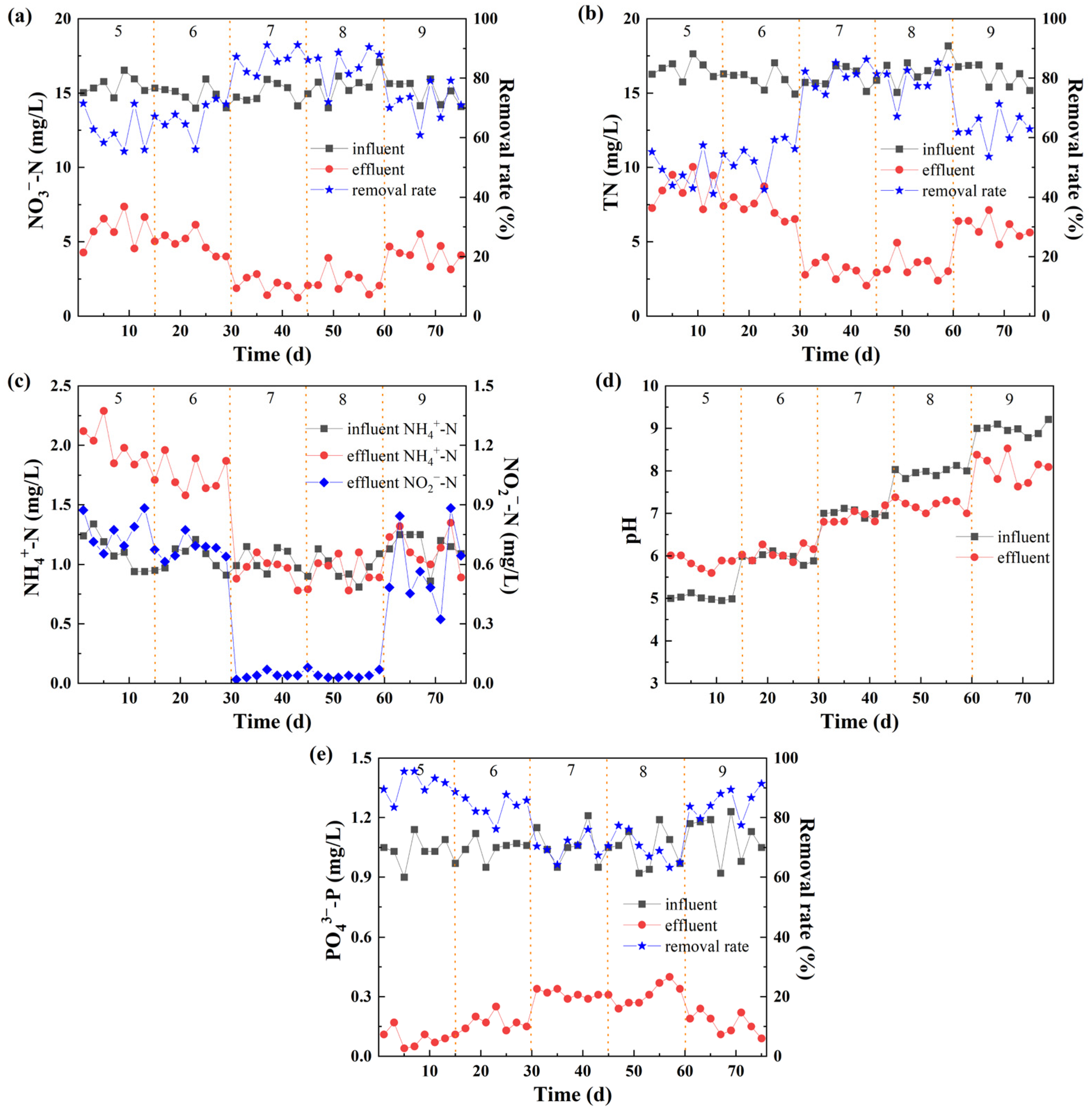
3.2. Impact of pH Value on N and P Removal
3.2.1. N Removal
3.2.2. P Removal
3.3. Impact of C/N on N and P Removal
3.3.1. NO3−-N Removal
3.3.2. PO43−-P Removal
3.4. Comparison of Different Processes
3.5. Analysis of Microbial Characteristics
3.5.1. Microbial Abundance and Diversity
3.5.2. Microbial Community Composition
3.6. Prediction of Microbial Community Functions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brozinčević, A.; Grgas, D.; Štefanac, T.; Habuda-Stanić, M.; Zelić, B.; Landeka Dragičević, T. Cost Reduction in the Process of Biological Denitrification by Choosing Traditional or Alternative Carbon Sources. Energies 2024, 17, 3660. [Google Scholar] [CrossRef]
- Chen, D.; Chen, P.; Zheng, X.; Cheng, W.; Wang, Q.; Wei, X. Enhanced Denitrification of Integrated Sewage Treatment System by Supplementing Denitrifying Carbon Source. Int. J. Environ. Res. Public Health 2021, 18, 9569. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.C.; Lampe, D.G. Sulfur: Limestone autotrophic denitrification processes for treatment of nitrate-contaminated water: Batch experiments. Water Res. 1999, 33, 599–608. [Google Scholar] [CrossRef]
- Hongbo, L.; Zihua, C.; Yongnian, G.; Suyun, X. Role and application of iron in water treatment for nitrogen removal: A review. Chemosphere 2018, 204, 51–62. [Google Scholar] [CrossRef]
- Sahinkaya, E.; Gunes, F.M.; Ucar, D.; Kaksonen, A.H. Sulfidogenic fluidized bed treatment of real acid mine drainage water. Bioresour. Technol. 2011, 102, 683–689. [Google Scholar] [CrossRef]
- Soares, M.I.M. Denitrification of groundwater with elemental sulfur. Water Res. 2002, 36, 1392–1395. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, T.; Morrison, L.; Gerrity, S.; Collins, G.; Porca, E.; Li, R.; Zhan, X. Nanostructured pyrrhotite supports autotrophic denitrification for simultaneous nitrogen and phosphorus removal from secondary effluents. Chem. Eng. J. 2017, 328, 511–518. [Google Scholar] [CrossRef]
- Zhang, M.; Zheng, P.; Li, W.; Wang, R.; Ding, S.; Abbas, G. Performance of nitrate-dependent anaerobic ferrous oxidizing (NAFO) process: A novel prospective technology for autotrophic denitrification. Bioresour. Technol. 2015, 179, 543–548. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, H.; Yang, K.; Ji, B.; Chen, D.; Zhang, H.; Sun, Y.; Tian, J. Autotrophic denitrification by nitrate-dependent Fe(II) oxidation in a continuous up-flow biofilter. Bioprocess Biosyst. Eng. 2016, 39, 277–284. [Google Scholar] [CrossRef]
- Zhou, Q.; Sun, H.; Jia, L.; Wu, W.; Wang, J. Simultaneous biological removal of nitrogen and phosphorus from secondary effluent of wastewater treatment plants by advanced treatment: A review. Chemosphere 2022, 296, 134054. [Google Scholar] [CrossRef]
- Peng, Y.; He, S.; Wu, F. Biochemical processes mediated by iron-based materials in water treatement: Enhancing nitrogen and phosphorus removal in low C/N ratio wastewater. Sci. Total Environ. 2021, 775, 145137. [Google Scholar] [CrossRef]
- Dai, H.; Sun, Y.; Wan, D.; Abbasi, H.N.; Guo, Z.; Geng, H.; Wang, X.; Chen, Y. Simultaneous denitrification and phosphorus removal: A review on the functional strains and activatedsludge processes. Sci. Total Environ. 2022, 835, 155409. [Google Scholar] [CrossRef] [PubMed]
- Kuba, T.; Loosdrecht, M.; Heijnen, J. Phosphorus and nitrogen removal with minimal COD requirement by integration of denitrifying dephosphatation and nitrification in a two-sludge system. Water Res. 1996, 30, 1702–1710. [Google Scholar] [CrossRef]
- HJ 636-2012; Water quality-Determination of total nitrogen-Alkaline potassium persulfate digestion UV spectrophotometric method. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China.
- HJ 828-2017; Water quality-Determination of the chemical oxygen demand-Dichromate method. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China.
- HJ 670-2013; Water quality-Determination of orthophosphate and total phosphorus-Continuous flow analysis(CFA) and Ammonium molybdate spectrophotometry. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China.
- GB 7480-87; Water quality-Determination of nitrate-Spectrophotometric method with phenol disulfonic acid. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China.
- HJ 535-2009; Water quality―Determination of ammonia nitrogen―Nessler’s reagent spectrophotometry. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China.
- GB 7493-87; Water quality-Determination of nitrogen (nitrite)-Spectrophotometric method. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China.
- Tong, S.; Rodriguez-Gonzalez, L.C.; Feng, C.; Ergas, S.J. Comparison of particulate pyrite autotrophic denitrification (PPAD) and sulfur oxidizing denitrification (SOD) for treatment of nitrified wastewater. Water Sci. Technol. 2017, 75, 239–246. [Google Scholar] [CrossRef]
- Tong, S.; Stocks, J.L.; Rodriguez-Gonzalez, L.C.; Feng, C.; Ergas, S.J. Effect of oyster shell medium and organic substrate on the performance of a particulate pyrite autotrophic denitrification (PPAD) process. Bioresour. Technol. 2017, 244, 296–303. [Google Scholar] [CrossRef]
- Francesco, C.D.; Giovanni, E. Pyrite-assisted denitrification in recirculated biofilter tolerates pH lower than 5. Water Environ. Res. 2022, 94, 10721. [Google Scholar] [CrossRef]
- Jin, S.; Feng, C.; Tong, S.; Chen, N.; Liu, H.; Zhao, J. Effect of sawdust dosage and hydraulic retention time (HRT) on nitrate removal in sawdust/pyrite mixotrophic denitrification (SPMD) systems. Environ. Sci. Water Res. Technol. 2019, 5, 346–357. [Google Scholar] [CrossRef]
- Liu, N.; Skauge, T.; Landa-Marbán, D.; Hovland, B.; Thorbjørnsen, B.; Radu, F.A.; Vik, B.F.; Baumann, T.; Bødtker, G. Microfluidic study of effects of flow velocity and nutrient concentration on biofilm accumulation and adhesive strength in the flowing and no-flowing microchannels. J. Ind. Microbiol. Biotechnol. 2019, 46, 855–868. [Google Scholar] [CrossRef]
- Jin, R.; Wang, B.; Liu, G.; Wang, Y.; Zhou, J.; Wang, J. Bioreduction of Cr(VI) by Acinetobacter sp. WB-1 during simultaneous nitrification/denitrification process. J. Chem. Technol. Biotechnol. 2017, 92, 649–656. [Google Scholar] [CrossRef]
- Matsubayashi, M.; Harada, H.; Okubo, T.; Uemura, S. Recovery of partial nitrification in a down-flow hanging sponge reactor by salt shock loading. J. Environ. Sci. Health A 2016, 51, 262–266. [Google Scholar] [CrossRef]
- Li, R.; Pan, J.; Qin, W.; Yang, J.; He, Y. Effect of pH on characterization and coagulation performance of poly-silicic-cation coagulant. Desalination 2014, 351, 37–42. [Google Scholar] [CrossRef]
- Huang, Z.; Gao, J.; Liu, L.; Mao, Y.; Kang, H.; Song, Z.; Cai, M.; Guo, P.; Chen, K. Performance and by-product generation in sulfur-siderite/limestone autotrophic denitrification systems: Enhancing nitrogen removal efficiency and operational insights. J. Environ. Manag. 2024, 370, 123042. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xue, L.; Wang, L.; Liu, C.; Li, Y. Simultaneous heterotrophic and FeS2-based ferrous autotrophic denitrification process for low-C/N ratio wastewater treatment: Nitrate removal performance and microbial community analysis. Sci. Total Environ. 2022, 829, 154682. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Sun, R.; Hu, S.; Qiao, Z.; Wang, S.; Zhang, Z. Nitrogen species control the interaction between NO3−-N reduction and aniline degradation and microbial communitystructure in the oxic-anoxic transition zone. Environ. Sci. Pollut. Res. 2021, 28, 29650–29664. [Google Scholar] [CrossRef]
- Almeida, J.S.; Reis, M.A.M.; Carrondo, M.J.T. Competition between nitrate and nitrite reduction in denitrification by Pseudomonas fluorescens. Biotechnol. Bioeng. 1995, 46, 476–484. [Google Scholar] [CrossRef]
- Zhao, J.; Li, N.; Yu, R.; Zhao, Z.; Nan, J. Magnetic field enhanced denitrification in nitrate and ammonia contaminated water under 3D/2D Mn2O3/g-C3N4 photocatalysis. Chem. Eng. J. 2018, 349, 530–538. [Google Scholar] [CrossRef]
- Qian, J.; Lu, H.; Cui, Y.; Wei, L.; Liu, R.; Chen, G.H. Investigation on thiosulfate-involved organics and nitrogen removal by a sulfur cycle-based biological wastewater treatment process. Water Res. 2015, 69, 295–306. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, J.J.; Choi, G.C.; Byun, I.G.; Park, T.J.; Lee, T.H. Field-scale application of spent sulfidic caustic as a source of alternative electron donor for autotrophic denitrification. Water Sci. Technol. 2013, 68, 479–485. [Google Scholar] [CrossRef]
- Mezenner, N.Y.; Bensmaili, A. Kinetics and thermodynamic study of phosphate adsorption on iron hydroxide-eggshell waste. Chem. Eng. J. 2009, 147, 87–96. [Google Scholar] [CrossRef]
- Verbaendert, I.; Boon, N.; De Vos, P.; Heylen, K. Denitrification is a common feature among members of the genus Bacillus. Syst. Appl. Microbiol. 2011, 34, 385–391. [Google Scholar] [CrossRef]
- Paul, D.; Banerjee, A. Technologies for Biological and Bioelectrochemical Removal of Inorganic Nitrogen from Wastewater: A Review. Nitrogen 2022, 3, 298–313. [Google Scholar] [CrossRef]
- Klauson, D.; Budarnaja, O.; Beltran, I.C.; Krichevskaya, M.; Preis, S. Photocatalytic decomposition of humic acids in anoxic aqueous solutions producing hydrogen, oxygen and light hydrocarbons. Env. Technol. 2014, 35, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, L.; Su, J.; Ali, A.; Huang, T.; Wang, Y.; Zhang, P. Microbially driven Fe-N cycle: Intrinsic mechanisms, enhancement, and perspectives. Sci. Total Environ. 2024, 908, 168084. [Google Scholar] [CrossRef] [PubMed]
- Ntagia, E.; Lens, P. Pyrite-based denitrification combined with electrochemical disinfection to remove nitrate and microbial contamination from groundwater. npj Clean Water 2023, 6, 59. [Google Scholar] [CrossRef]
- Guo, H.; Zhai, X.; Hu, M.; Chang, J.; Lee, D. Atypical removals of nitrogen and phosphorus with biochar-pyrite vertical flow constructed wetlands treating wastewater at low C/N ratio. Bioresour. Technol. 2025, 422, 132219. [Google Scholar] [CrossRef]
- Li, J.; Zeng, W.; Liu, H.; Zhan, M.; Miao, H. Achieving deep autotrophic nitrogen removal in aerated biofilter driven by sponge iron: Performance and mechanism. Environ. Res. 2022, 213, 113653. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, H.; Chu, Z.; Hang, Q.; Liu, K.; Li, C. Influence of C/N ratio on MBBR denitrification for advanced nitrogen removal of wastewater treatment plant effluent. Desalination Water Treat. 2017, 66, 158–165. [Google Scholar] [CrossRef]
- Zhou, Q.; Jia, L.; Wu, W.; Wu, W. Introducing PHBV and controlling the pyrite sizes achieved the pyrite-based mixotrophic denitrification under natural aerobic conditions: Low sulfate production and functional microbe interaction. J. Clean. Prod. 2022, 366, 132986. [Google Scholar] [CrossRef]
- Shen, J.; Huang, G.; An, C.; Song, P.; Xin, X.; Yao, Y.; Zheng, R. Biophysiological and factorial analyses in the treatment of rural domestic wastewater using multi-soil-layering systems. J. Environ. Manag. 2018, 226, 83–94. [Google Scholar] [CrossRef]
- Liu, C.; Xie, J.; Song, M.; Gao, Z.; Zheng, D.; Liu, X.; Ning, G.; Cheng, X.; Bruning, H. Nitrogen removal performance and microbial community changes in subsurface wastewater infiltration systems (SWISs) at low temperature with different bioaugmentation strategies. Bioresour. Technol. 2018, 250, 603–610. [Google Scholar] [CrossRef]
- Yang, S.; Zheng, Y.; Mao, Y.; Xu, L.; Jin, Z.; Zhao, M.; Kong, H.; Huang, X.; Zheng, X. Domestic wastewater treatment for single household via novel subsurface wastewater infiltration systems (SWISs) with NiiMi process: Performance and microbial community. J. Clean. Prod. 2021, 279, 123434. [Google Scholar] [CrossRef]
- Arshad, A.; Dalcin Martins, P.; Frank, J.; Jetten, M.S.M.; Op den Camp, H.J.M.; Welte, C.U. Mimicking microbial interactions under nitrate-reducing conditions in an anoxic bioreactor: Enrichment of novel Nitrospirae bacteria distantly related to Thermodesulfovibrio. Environ. Microbiol. 2017, 19, 4965–4977. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, X.; Chi, L.; Wang, J. The design and operation of subsurface wastewater infiltration systems for domestic wastewater. Water Environ. Res. 2019, 91, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Srinandan, C.S.; Shah, M.; Patel, B.; Nerurkar, A.S. Assessment of denitrifying bacterial composition in activated sludge. Bioresour Technol. 2011, 102, 9481–9489. [Google Scholar] [CrossRef]
- Haig, S.J.; Quince, C.; Davies, R.L.; Dorea, C.C.; Collins, G. Replicating the microbial community and water quality performance of full-scale slow sand filters in laboratory-scale filters. Water Res. 2014, 61, 141–151. [Google Scholar] [CrossRef]
- Zhu, Y.; Xu, J.; Cao, X.; Cheng, Y. Characterization of functional microbial communities involved in different transformation stages in a full-scale printing and dyeing wastewater treatment plant. Biochem. Eng. J. 2018, 137, 162–171. [Google Scholar] [CrossRef]
- Song, B.; Palleroni, N.J.; Kerkhof, L.J.; Haggblom, M.M. Characterization of halobenzoate-degrading, denitrifying Azoarcus and Thauera isolates and description of Thauera chlorobenzoica sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 589–602. [Google Scholar] [CrossRef]
- Oprime, M.E.A.G.; Jr, O.G.; Cardoso, A.A. Oxidation of H2S in acid solution by Thiobacillus ferrooxidans and Thiobacillus thiooxidans. Process Biochem. 2001, 37, 111–114. [Google Scholar] [CrossRef]
- Coates, J.D.; Bhupathiraju, V.K.; Achenbach, L.A.; McInerney, M.J.; Lovley, D.R. Geobacter hydrogenophilus, Geobacter chapellei and Geobacter grbiciae, three new, strictly anaerobic, dissimilatory Fe(III)-reducers. Int. J. Syst. Evol. Microbiol. 2001, 51, 581–588. [Google Scholar] [CrossRef]
- Semrau, J.D. Bioremediation via Methanotrophy: Overview of Recent Findings and Suggestions for Future Research. Front. Microbiol. 2011, 2, 209. [Google Scholar] [CrossRef]
- Ochsner, A.M.; Sonntag, F.; Buchhaupt, M.; Schrader, J.; Vorholt, J.A. Methylobacterium extorquens: Methylotrophy and biotechnological applications. Appl. Microbiol. Biotechnol. 2015, 99, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Keller, K.L.; Wall, J.D. Genetics and molecular biology of the electron flow for sulfate respiration in desulfovibrio. Front. Microbiol. 2011, 2, 135. [Google Scholar] [CrossRef] [PubMed]
- Schwedt, A.; Kreutzmann, A.C.; Polerecky, L.; Schulz-Vogt, H.N. Sulfur respiration in a marine chemolithoautotrophic beggiatoa strain. Front. Microbiol. 2011, 2, 276. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Tao, Y.; Song, L.; Wang, H.; Ren, B. A unique microbiome in a highly polluted and alkalic lake in a seasonally frozen area. Environ. Res. 2022, 204, 112056. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Results | Comparisons | Reference |
|---|---|---|---|
| Pyrite-based denitrification combined with electrochemical disinfection | At a hydraulic retention time of 18 h, a nitrate removal efficiency of 79% was achieved with an initial nitrate concentration of 178 mg/L NO3−. | This study achieved a higher denitrification efficiency while exhibiting significantly lower energy consumption compared to study of Ntagia et al. | [40] |
| Biochar-pyrite vertical flow constructed wetlands | At a biochar-to-pyrite volume ratio of 1:1, Highest TN removal reached at 86.0 ± 2.5%. No optimal biochar-to-pyrite ratio exists to peak TP removal. | In this study the P removal was higher and needed lower carbon source. | [41] |
| Aerated biofilter driven by sponge iron | The results showed ammonia removal efficiency reached 94.1% and total inorganic nitrogen removal efficiency was up to 70.6% at HRT of 19 h and gas–water ratio of 18. | The N removal was lower than this study, and it might need more energy for aeration. | [42] |
| Moving bed biofilm reactor | The optimum choice of C/N ratio for nitrogen removal by denitrification MBBR was 4.6. | The optimal C/N ratio of this study was 4, which was lower while the removal of N was higher. | [43] |
| Pyrite/PHBV mixotrophic denitrification system | The removal efficiency of nitrogen and phosphorus was 96% and 25%, with low sulfate production. | Compared with study of Zhou et al., this study showed high phosphorus removal, strong resistance to shock loads, and low operational costs. | [44] |
| Sample | OTUs | Chao | Ace | Simpson | Shannon | Coverage |
|---|---|---|---|---|---|---|
| A1 | 2038 | 2418.3208 | 2383.0271 | 0.0075 | 5.9269 | 0.9908 |
| A2 | 2073 | 2477.6946 | 2437.3617 | 0.0086 | 5.9390 | 0.9909 |
| A3 | 2026 | 2383.2749 | 2330.4918 | 0.0084 | 5.8709 | 0.9919 |
| B1 | 1753 | 2044.6811 | 2003.0342 | 0.0307 | 5.3508 | 0.9943 |
| B2 | 1840 | 2167.6146 | 2110.4507 | 0.0113 | 5.7408 | 0.9939 |
| B3 | 1523 | 1818.5789 | 1812.5949 | 0.0135 | 5.3331 | 0.9942 |
| C1 | 1569 | 1896.4264 | 1843.5595 | 0.0204 | 5.0978 | 0.9933 |
| C2 | 1576 | 1852.0476 | 1827.5043 | 0.0294 | 4.9609 | 0.9939 |
| C3 | 1629 | 1933.4663 | 1919.5652 | 0.0257 | 5.0537 | 0.9940 |
| D1 | 1439 | 1765.9856 | 1681.0433 | 0.0117 | 5.5351 | 0.9946 |
| D2 | 1461 | 1826.9272 | 1776.9442 | 0.0220 | 5.1596 | 0.9941 |
| D3 | 1510 | 1806.4188 | 1789.8089 | 0.0427 | 4.9013 | 0.9948 |
| E | 1476 | 1845.8095 | 1781.8664 | 0.0540 | 4.6825 | 0.9948 |
| Group | A1 | A2 | A3 | B1 | B2 | B3 | C1 | C2 | C3 | D1 | D2 | D3 | E |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| methanotrophy | 754 | 632 | 598 | 568 | 513 | 470 | 87 | 134 | 138 | 110 | 101 | 313 | 222 |
| methylotrophy | 754 | 633 | 598 | 570 | 517 | 956 | 95 | 136 | 139 | 110 | 105 | 317 | 338 |
| aerobic_ammonia_oxidation | 11 | 14 | 10 | 5 | 83 | 30 | 41 | 41 | 35 | 86 | 188 | 42 | 142 |
| aerobic_nitrite_oxidation | 772 | 689 | 467 | 369 | 1191 | 996 | 36 | 45 | 42 | 69 | 24 | 52 | 1764 |
| nitrification | 783 | 703 | 477 | 374 | 1274 | 1026 | 77 | 86 | 77 | 155 | 212 | 94 | 1906 |
| sulfate_respiration | 246 | 253 | 247 | 277 | 343 | 71 | 82 | 100 | 85 | 375 | 1201 | 892 | 131 |
| sulfur_respiration | 2 | 9 | 8 | 27 | 9 | 8 | 7 | 7 | 8 | 282 | 13 | 9 | 10 |
| respiration_of_sulfur_compounds | 249 | 262 | 256 | 304 | 354 | 80 | 91 | 107 | 94 | 658 | 1216 | 901 | 142 |
| anammox | 855 | 908 | 649 | 139 | 378 | 57 | 77 | 20 | 197 | 2 | 4 | 3 | 0 |
| nitrate_denitrification | 0 | 1 | 2 | 95 | 311 | 0 | 4 | 2 | 2 | 1 | 0 | 0 | 2 |
| nitrite_denitrification | 0 | 1 | 2 | 95 | 311 | 0 | 4 | 2 | 2 | 1 | 0 | 0 | 2 |
| nitrous_oxide_denitrification | 0 | 1 | 2 | 95 | 311 | 0 | 4 | 2 | 2 | 1 | 0 | 0 | 2 |
| denitrification | 0 | 1 | 2 | 95 | 311 | 0 | 4 | 2 | 2 | 1 | 0 | 0 | 2 |
| nitrogen_fixation | 39 | 53 | 57 | 302 | 62 | 64 | 170 | 189 | 236 | 87 | 183 | 294 | 225 |
| nitrite_respiration | 1325 | 1468 | 1097 | 10,723 | 5566 | 706 | 2347 | 236 | 643 | 71 | 34 | 46 | 123 |
| dark_sulfide_oxidation | 42 | 41 | 64 | 94 | 198 | 51 | 40 | 85 | 115 | 624 | 89 | 98 | 10 |
| dark_oxidation_of_sulfur_compounds | 44 | 43 | 65 | 107 | 202 | 55 | 83 | 97 | 148 | 642 | 93 | 101 | 19 |
| aerobic_chemoheterotrophy | 1952 | 1973 | 2208 | 12,725 | 6481 | 10,502 | 5985 | 2936 | 3822 | 3947 | 2112 | 2048 | 4458 |
| hydrocarbon_degradation | 766 | 638 | 606 | 583 | 518 | 476 | 105 | 138 | 140 | 116 | 102 | 314 | 223 |
| nitrate_respiration | 564 | 679 | 531 | 10,945 | 5314 | 774 | 2438 | 363 | 707 | 181 | 174 | 104 | 1006 |
| nitrate_reduction | 612 | 742 | 591 | 11,047 | 5661 | 873 | 2502 | 418 | 762 | 244 | 247 | 193 | 1089 |
| nitrogen_respiration | 1419 | 1587 | 1180 | 11,084 | 5692 | 831 | 2515 | 383 | 904 | 183 | 178 | 107 | 1006 |
| chemoheterotrophy | 3106 | 2966 | 3172 | 13,795 | 7770 | 11,800 | 6506 | 3544 | 4427 | 6098 | 2930 | 3074 | 5377 |
| sulfite_respiration | 5 | 2 | 2 | 7 | 19 | 4 | 7 | 6 | 8 | 16 | 39 | 68 | 32 |
| thiosulfate_respiration | 3 | 9 | 9 | 27 | 11 | 9 | 9 | 7 | 9 | 285 | 15 | 9 | 16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Ma, M.; Chen, S.; Yuan, R.; Wang, S. Study on the Synchronous Removal of Nitrogen and Phosphorus by Autotrophic/Heterotrophic Denitrification in the Presence of Pyrite. Molecules 2025, 30, 2412. https://doi.org/10.3390/molecules30112412
Zhu M, Ma M, Chen S, Yuan R, Wang S. Study on the Synchronous Removal of Nitrogen and Phosphorus by Autotrophic/Heterotrophic Denitrification in the Presence of Pyrite. Molecules. 2025; 30(11):2412. https://doi.org/10.3390/molecules30112412
Chicago/Turabian StyleZhu, Minyi, Minhui Ma, Shuo Chen, Rongfang Yuan, and Shaona Wang. 2025. "Study on the Synchronous Removal of Nitrogen and Phosphorus by Autotrophic/Heterotrophic Denitrification in the Presence of Pyrite" Molecules 30, no. 11: 2412. https://doi.org/10.3390/molecules30112412
APA StyleZhu, M., Ma, M., Chen, S., Yuan, R., & Wang, S. (2025). Study on the Synchronous Removal of Nitrogen and Phosphorus by Autotrophic/Heterotrophic Denitrification in the Presence of Pyrite. Molecules, 30(11), 2412. https://doi.org/10.3390/molecules30112412