
Norsesquiterpenes from Lolium perenne and Their Replacement Control of an Invasive Plant, Ageratina adenophora, Through Allelopathy

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
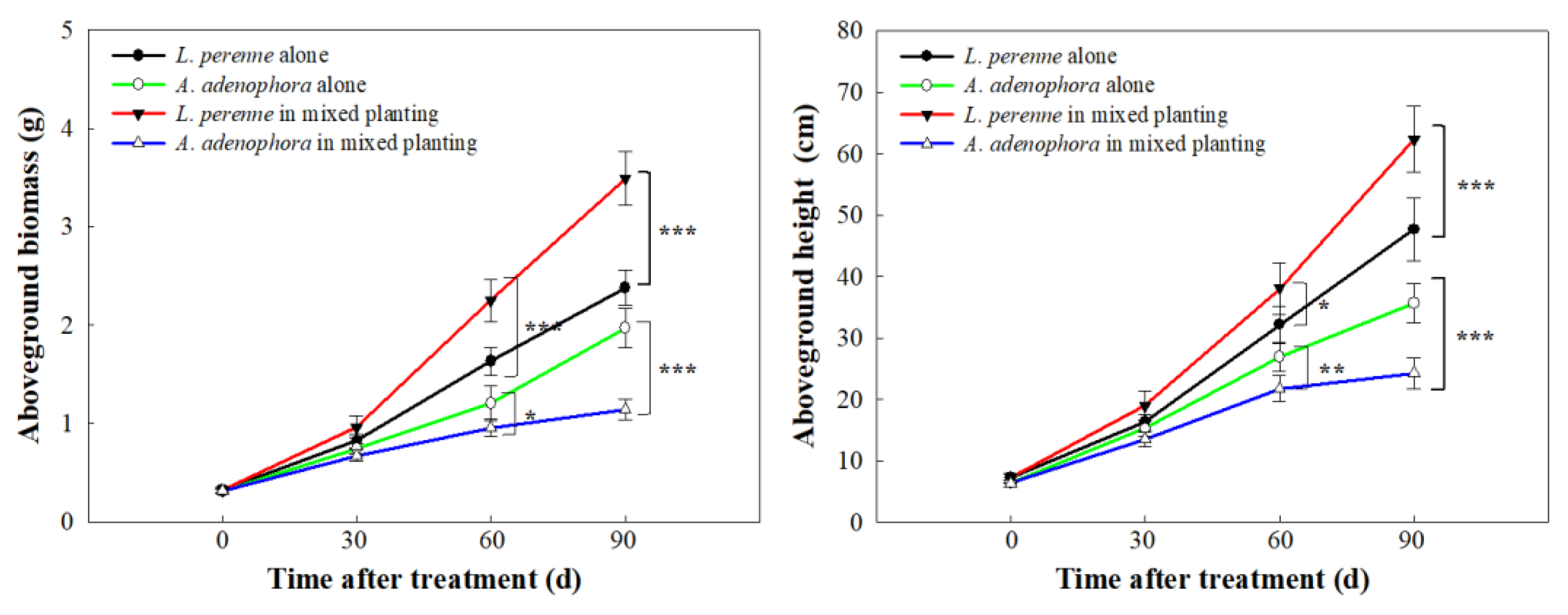
2.1. Growth Competition Assay
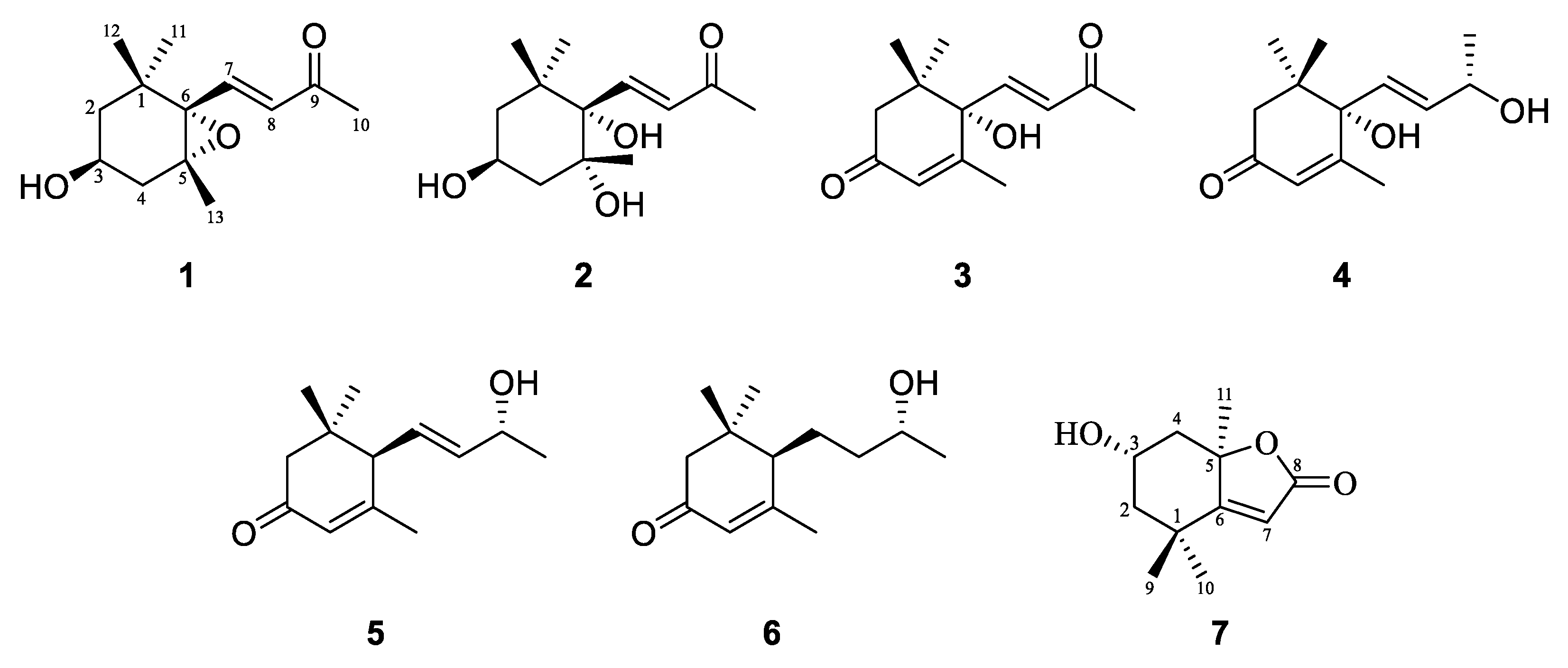
2.2. Structural Identification
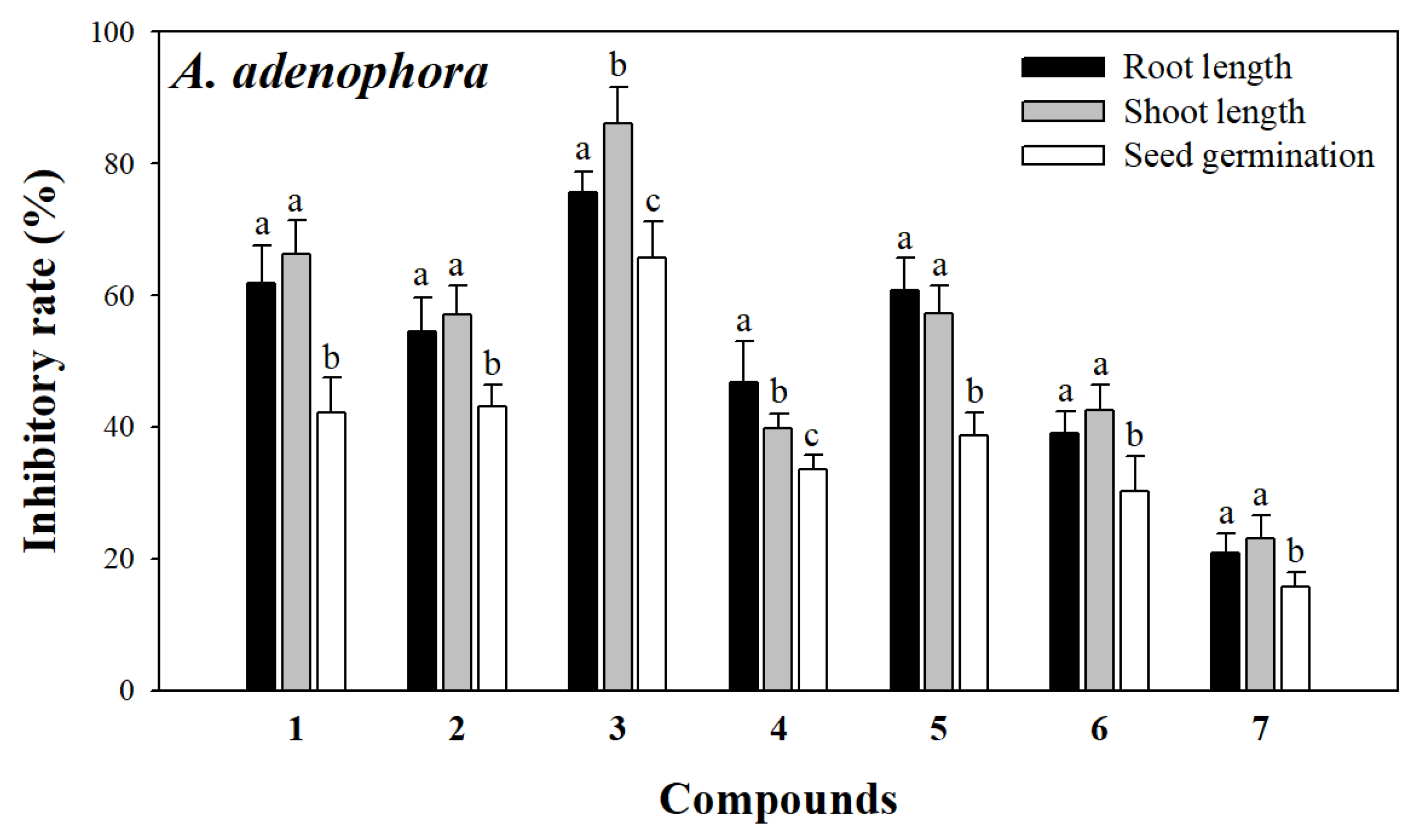
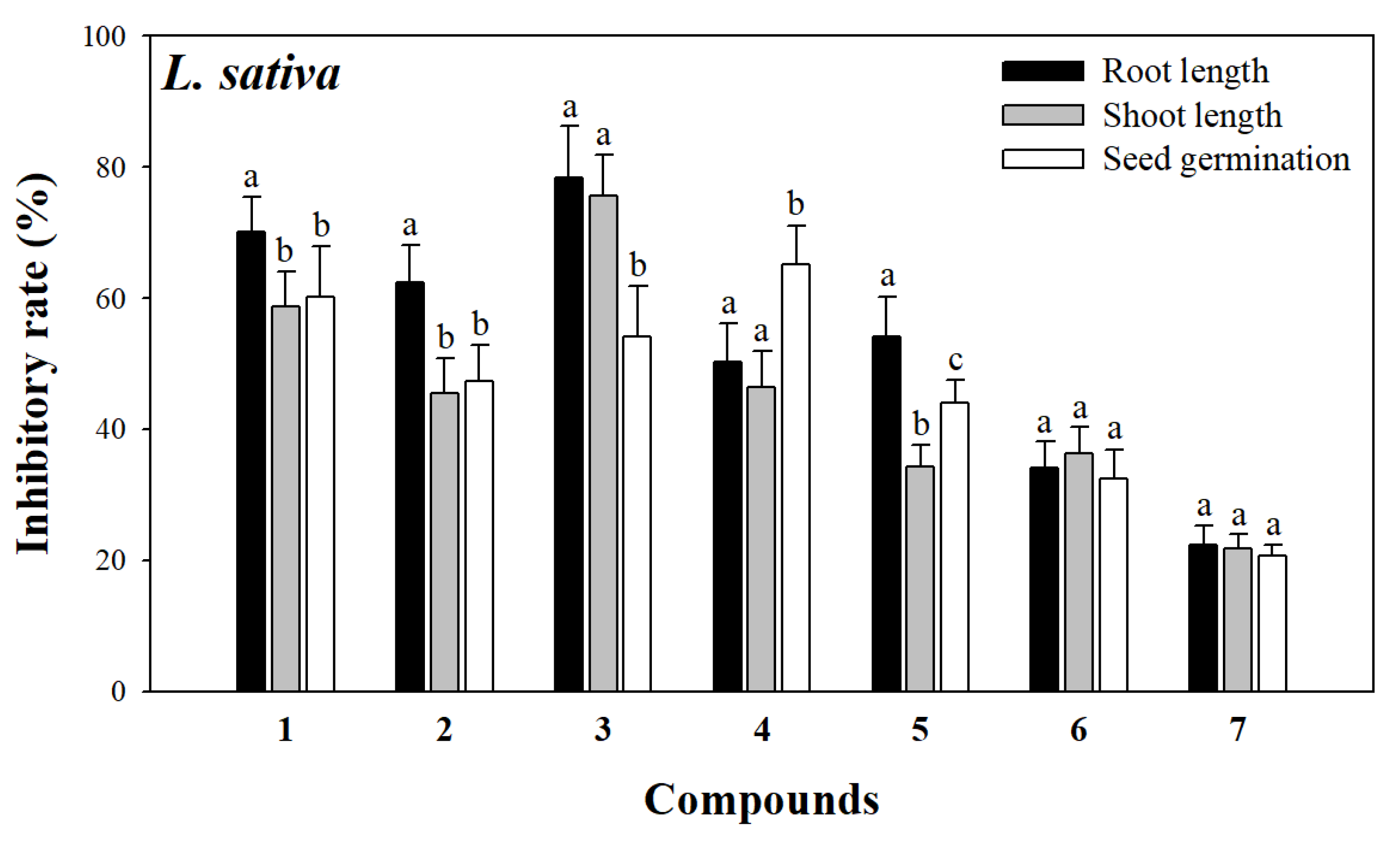
2.3. Allelopathic Assay of Compounds
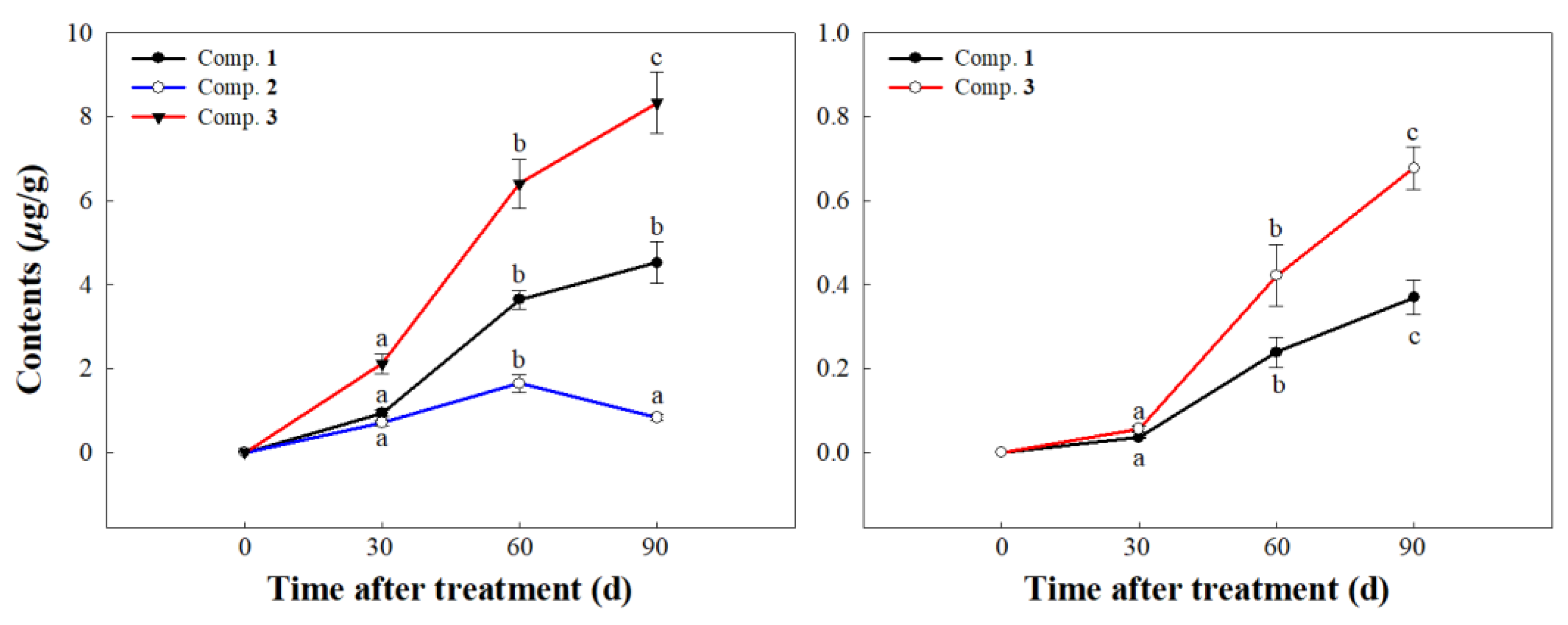
2.4. UPLC-MS/MS Analyses
3. Materials and Methods
3.1. Plant Material
3.2. Growth Competition Assay
3.3. Extraction and Isolation
3.4. Structural Identification
3.5. Allelopathic Assay
3.6. Collection of Root Secretion and Rainwater Leaching
3.7. UPLC-MS/MS Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonnamour, A.; Gippet, J.M.W.; Bertelsmeier, C.; Gurevitch, J. Insect and plant invasions follow two waves of globalisation. Ecol. Lett. 2021, 24, 2418–2426. [Google Scholar] [CrossRef] [PubMed]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pysek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- Sobuj, N.; Byun, C. A synthesis of plant invasion control: Important factors to consider when choosing a control method. Écoscience 2023, 30, 234–246. [Google Scholar] [CrossRef]
- Weidlich, E.W.A.; Flórido, F.G.; Sorrini, T.B.; Brancalion, P.H.S. Controlling invasive plant species in ecological restoration: A global review. J. Appl. Ecol. 2020, 57, 1806–1817. [Google Scholar] [CrossRef]
- Piemeisel, R.L.; Carsner, E. Replacement control and biological control. Science 1951, 113, 14–15. [Google Scholar] [CrossRef]
- Li, W.; Luo, J.; Tian, X.; Chow, W.S.; Sun, Z.; Zhang, T.; Peng, S.; Peng, C. A new strategy for controlling invasive weeds: Selecting valuable native plants to defeat them. Sci. Rep. 2015, 5, 11004. [Google Scholar] [CrossRef]
- Jia, P.; Wang, J.; Liang, H.; Wu, Z.H.; Li, F.; Li, W. Replacement control of Mikania micrantha in orchards and its eco-physiological mechanism. Front. Ecol. Evol. 2022, 10, 1095946. [Google Scholar] [CrossRef]
- Chen, B.M.; Liao, H.X.; Chen, W.B.; Wei, H.J.; Peng, S.L. Role of allelopathy in plant invasion and control of invasive plants. Allelopathy J. 2017, 41, 155–166. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. The ecological importance of allelopathy. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 25–45. [Google Scholar] [CrossRef]
- Khamare, Y.; Chen, J.; Marble, S.C. Allelopathy and its application as a weed management tool: A review. Front. Plant Sci. 2022, 13, 1034649. [Google Scholar] [CrossRef]
- Putnam, A.R. Allelochemicals from plants as herbicides. Weed Technol. 1988, 2, 510–518. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.J.; Fu, H.Y.; He, W.; Wang, Y.X.; Sampietro, D.A.; Yang, S.X.; Kuang, Y. Herbicide potential of new phytotoxins structurally based on plant allelochemicals. Allelopathy J. 2022, 57, 191–204. [Google Scholar] [CrossRef]
- Jayasinghe, C.; Badenhorst, P.; Jacobs, J.; Spangenberg, G.; Smith, K. Image-based high-throughput phenotyping for the estimation of persistence of perennial ryegrass (Lolium perenne L.)—A review. Grass Forage Sci. 2021, 76, 321–339. [Google Scholar] [CrossRef]
- Wims, C.M.; Delaby, L.; Boland, T.M.; O’Donovan, M. Effect of pre-grazing herbage mass on dairy cow performance, grass dry matter production and output from perennial ryegrass (Lolium perenne L.) pastures. Animal 2014, 8, 141–151. [Google Scholar] [CrossRef]
- Dineen, M.; McCarthy, B.; Ross, D.; Ortega, A.; Dillon, P.; van Amburgh, M.E. Characterization of the nutritive value of perennial ryegrass (Lolium perenne L.) dominated pastures using updated chemical methods with application for the Cornell Net Carbohydrate and Protein System. Anim. Feed Sci. Technol. 2021, 272, 114752. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Jing, Q.; Wang, J. Light pollution on the growth, physiology and chlorophyll fluorescence response of landscape plant perennial ryegrass (Lolium perenne L.). Ecol. Indic. 2020, 115, 106448. [Google Scholar] [CrossRef]
- Poudel, A.S.; Jha, P.K.; Shrestha, B.B.; Muniappan, R. Biology and management of the invasive weed Ageratina adenophora (Asteraceae): Current state of knowledge and future research needs. Weed Res. 2019, 59, 79–92. [Google Scholar] [CrossRef]
- Kluge, R.L. Biological control of crofton weed, Ageratina adenophora (Asteraceae), in South Africa. Agr. Ecosyst. Environ. 1991, 37, 187–191. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, Q.; Chen, Y.; Zhang, L.D.; Tan, L.; Fu, R.H.; Yang, J.T.; Li, Y.; Liu, M.; Compton, S.G. Impacts of a biocontrol agent on invasive Ageratina adenophora in Southwest China: Friend or foe? Biol. Control 2021, 152, 104471. [Google Scholar] [CrossRef]
- Ren, Z.; Okyere, S.K.; Wen, J.; Xie, L.; Cui, Y.; Wang, S.; Wang, J.; Cao, S.; Shen, L.; Ma, X.; et al. An overview: The toxicity of Ageratina adenophora on animals and its possible interventions. Int. J. Mol. Sci. 2021, 22, 11581. [Google Scholar] [CrossRef]
- Zhao, L.; Li, B.P.; Meng, L.; Zhu, H.W. The relative competitive ability of perennial Lolium perenne and the invasive alien weed, Eupatorium adenophorum (Compositae) at different nitrogen and phosphorus levels in the seedling stage. Acta Prataculturae Sin. 2008, 17, 151–155. [Google Scholar]
- Zhu, H.; Meng, L.; Li, B. Relative competitive ability of Lolium perenne and the invasive alien weed, Eupatorium adenophorum (Compositae) at seedling stage. Chin. J. Appl. Environ. Biol. 2007, 13, 29–32. [Google Scholar]
- Matsuo, T.; Bongers, F.; Martínez-Ramos, M.; van der Sande, M.T.; Poorter, L. Height growth and biomass partitioning during secondary succession differ among forest light strata and successional guilds in a tropical rainforest. Oikos 2024, 2024, e10486. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, K.H.; Choi, S.U.; Kim, Y.H.; Lee, K.R. Terpene and phenolic constituents of Lactuca indica L. Arch. Pharm. Res. 2008, 31, 983–988. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, D.G.; Yeon, S.W.; Kwon, H.S.; Ko, J.H.; Shin, D.J.; Park, H.S.; Kim, Y.S.; Bang, M.H.; Baek, N.I. Isolation of megastigmane sesquiterpenes from the Silkworm (Bombyx mori L.) droppings and their promotion activity on HO-1 and SIRT1. Arch. Pharm. Res. 2011, 34, 533–542. [Google Scholar] [CrossRef]
- Yan, Z.H.; Han, Z.Z.; Hu, X.Q.; Liu, Q.X.; Zhang, W.D.; Liu, R.H.; Li, H.L. Chemical constituents of Euonymus alatus. Chem. Nat. Compd. 2013, 49, 340–342. [Google Scholar] [CrossRef]
- Le, H.L.; Nguyen, T.M.H.; Vu, T.T.; Nguyen, T.T.O.; Ly, D.T.; Le, N.T.; Nguyen, V.H.; Nguyen, T.V.A. Potent antiplatelet aggregation, anticoagulant and antioxidant activity of aerial Canna x generalis L.H Bailey & E.Z Bailey and its phytoconstituents. S. Afr. J. Bot. 2022, 147, 882–893. [Google Scholar]
- Yuan, Z.; Zheng, X.; Zhao, Y.; Liu, Y.; Zhou, S.; Wei, C.; Hu, Y.; Shao, H. Phytotoxic compounds isolated from leaves of the invasive weed Xanthium spinosum. Molecules 2018, 23, 2840. [Google Scholar] [CrossRef]
- Morais, A.M.M.B.; Kumla, D.; Martins, V.F.R.; Alves, A.; Gales, L.; Silva, A.M.S.; Costa, P.M.; Mistry, S.; Kijjoa, A.; Morais, R.M.S.C. Monoterpene hydroxy lactones isolated from Thalassiosira sp. microalga and their antibacterial and antioxidant activities. Molecules 2024, 29, 5175. [Google Scholar] [CrossRef]
- Finch, S.C.; Prinsep, M.R.; Popay, A.J.; Wilkins, A.L.; Webb, N.G.; Bhattarai, S.; Jensen, J.G.; Hawkes, A.D.; Babu, J.V.; Tapper, B.A.; et al. Identification and structure elucidation of epoxyjanthitrems from Lolium perenne infected with the endophytic fungus Epichloë festucae var. lolii and determination of the tremorgenic and anti-insect activity of Epoxyjanthitrem I. Toxins 2020, 12, 526. [Google Scholar] [CrossRef]
- Reddy, P.; Deseo, M.A.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.; Rochfort, S. Toxic indole diterpenes from endophyte-infected perennial ryegrass Lolium perenne L.: Isolation and stability. Toxins 2019, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Hossen, K.; Asato, Y.; Teruya, T.; Kato-Noguchi, H. Identification of four allelopathic compounds including a novel compound from Elaeocarpus foribundus Blume and determination of their allelopathic activity. J. Environ. Manag. 2023, 326, 116728. [Google Scholar] [CrossRef] [PubMed]
- Macias, F.A.; Varela, R.M.; Torres, A.; Oliva, R.M.; Molinillo, J.M.G. Bioactive norsesquiterpenes from Helianthus annuus with potential allelopathic activity. Phytochemistry 1998, 48, 631–636. [Google Scholar] [CrossRef]
- Kyaw, E.H.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Allelopathy of the medicinal plant Dregea volubilis (L.f.) Benth.ex Hook.f. and its phytotoxic substances with allelopathic activity. Agronomy 2022, 12, 303. [Google Scholar] [CrossRef]
- Ma, W.; Tang, S.; Deng, Z.; Zhang, D.; Zhang, T.; Ma, X. Root exudates contribute to belowground ecosystem hotspots: A review. Front. Microbiol. 2022, 13, 937940. [Google Scholar] [CrossRef]
- Wang, A.K.; Huang, K.W.; Ning, Y.L.; Bi, Y.F. Allelochemicals from Moso bamboo: Identification and their effects on neighbor species. Forests 2024, 15, 2040. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.; An, T.; Yang, X.; Li, Y.; Yimingniyazi, A.; Liu, Z.; Feng, Y. Norsesquiterpenes from Lolium perenne and Their Replacement Control of an Invasive Plant, Ageratina adenophora, Through Allelopathy. Molecules 2025, 30, 2384. https://doi.org/10.3390/molecules30112384
Shi W, An T, Yang X, Li Y, Yimingniyazi A, Liu Z, Feng Y. Norsesquiterpenes from Lolium perenne and Their Replacement Control of an Invasive Plant, Ageratina adenophora, Through Allelopathy. Molecules. 2025; 30(11):2384. https://doi.org/10.3390/molecules30112384
Chicago/Turabian StyleShi, Wenbo, Tong An, Xiaomin Yang, Youlin Li, Amanula Yimingniyazi, Zhixiang Liu, and Yulong Feng. 2025. "Norsesquiterpenes from Lolium perenne and Their Replacement Control of an Invasive Plant, Ageratina adenophora, Through Allelopathy" Molecules 30, no. 11: 2384. https://doi.org/10.3390/molecules30112384
APA StyleShi, W., An, T., Yang, X., Li, Y., Yimingniyazi, A., Liu, Z., & Feng, Y. (2025). Norsesquiterpenes from Lolium perenne and Their Replacement Control of an Invasive Plant, Ageratina adenophora, Through Allelopathy. Molecules, 30(11), 2384. https://doi.org/10.3390/molecules30112384