
Characterization of Terpene Synthases Reveals the Diversity of Terpenoids in Andrographis paniculata


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
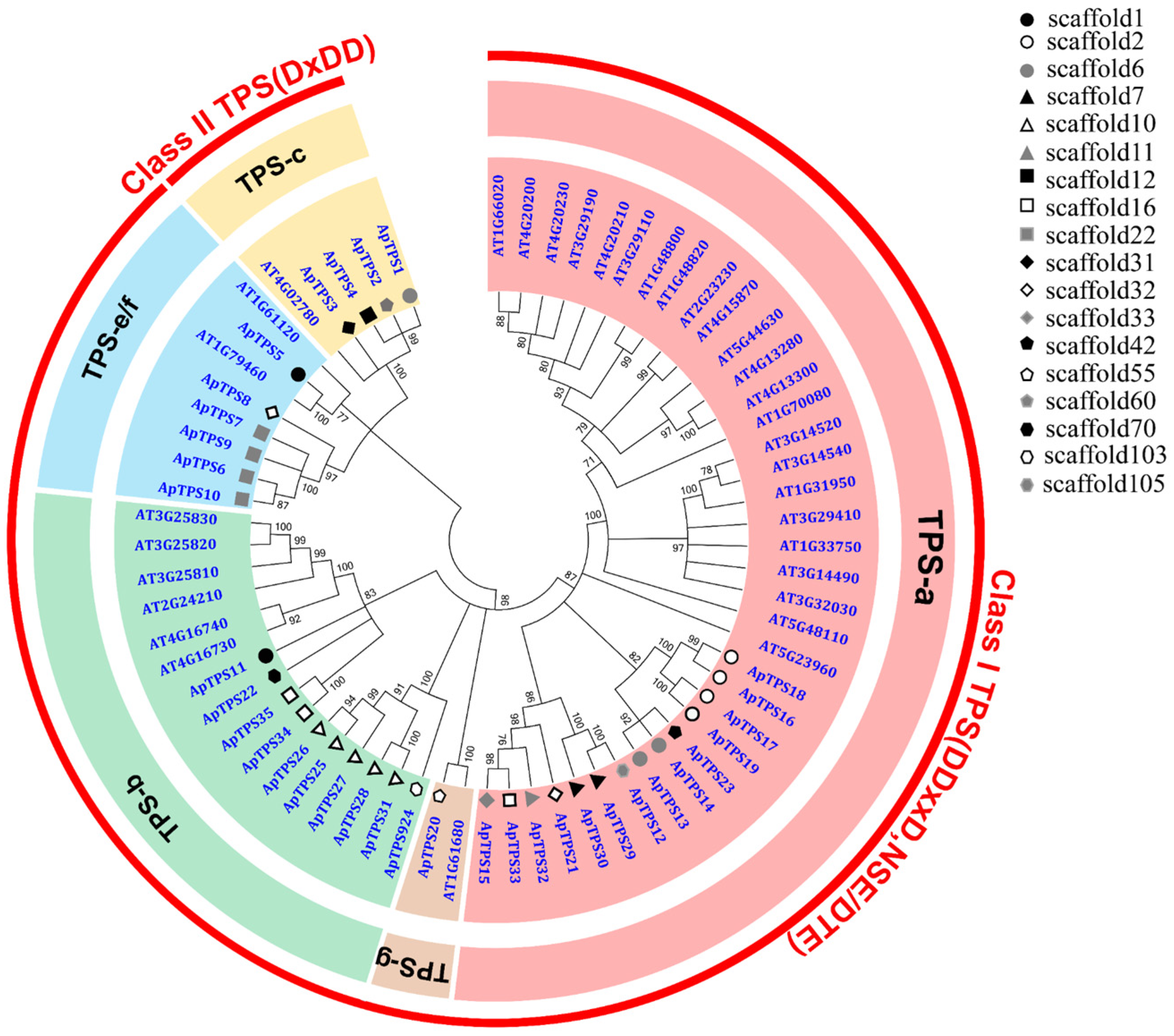
2.1. Phylogenetic Analysis of ApTPSs
2.2. Molecular Cloning of the Full-Length cDNA Encoding ApTPSs
2.3. Functional Identification of Recombinant Class II diTPS (TPS-c Subfamily)
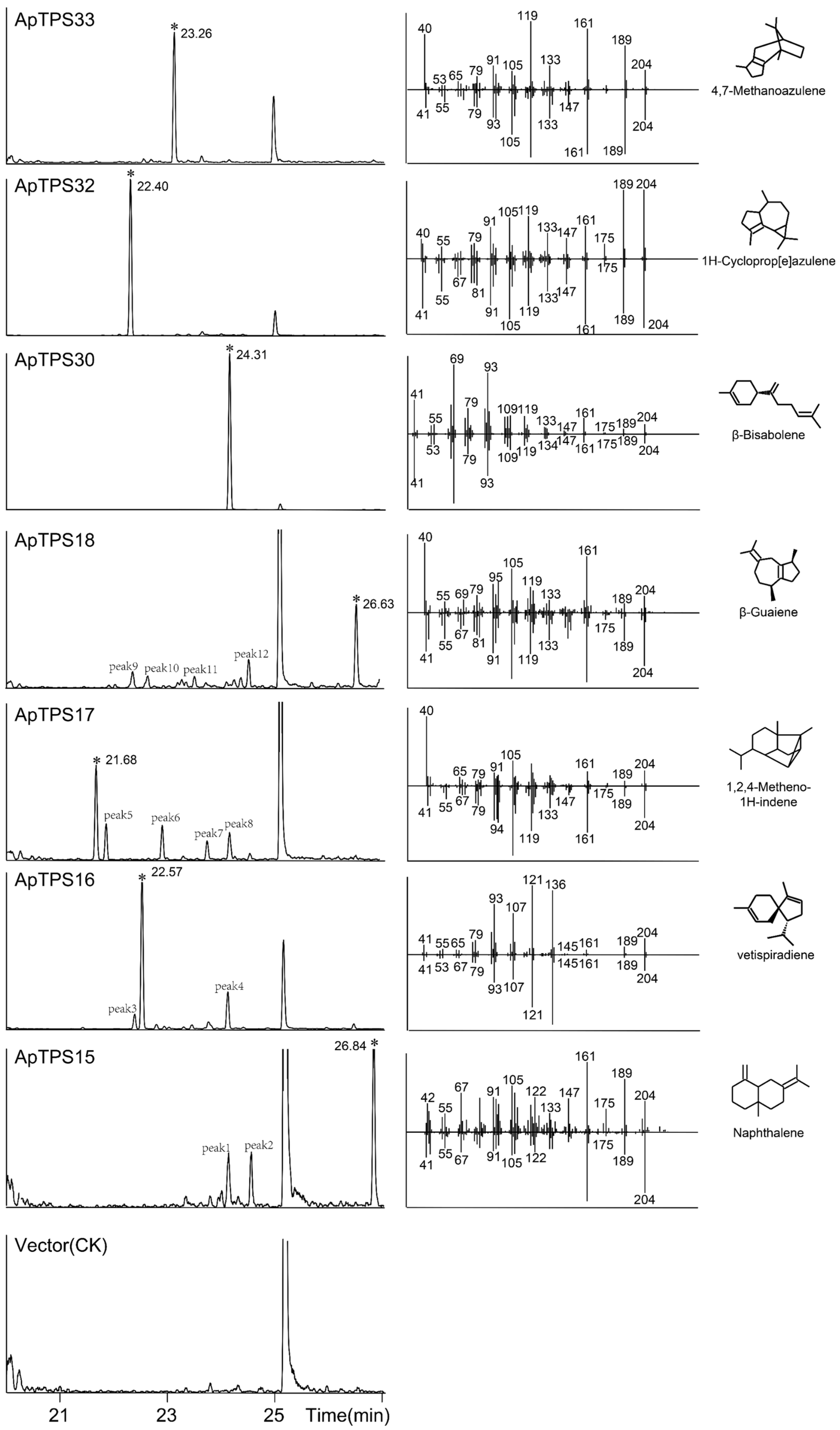
2.4. Identification of Recombinant Class I diTPS (TPS-e/f Subfamily)
2.5. Identification of Other Recombinant Class I TPSs (TPS-a/b/g Subfamily)
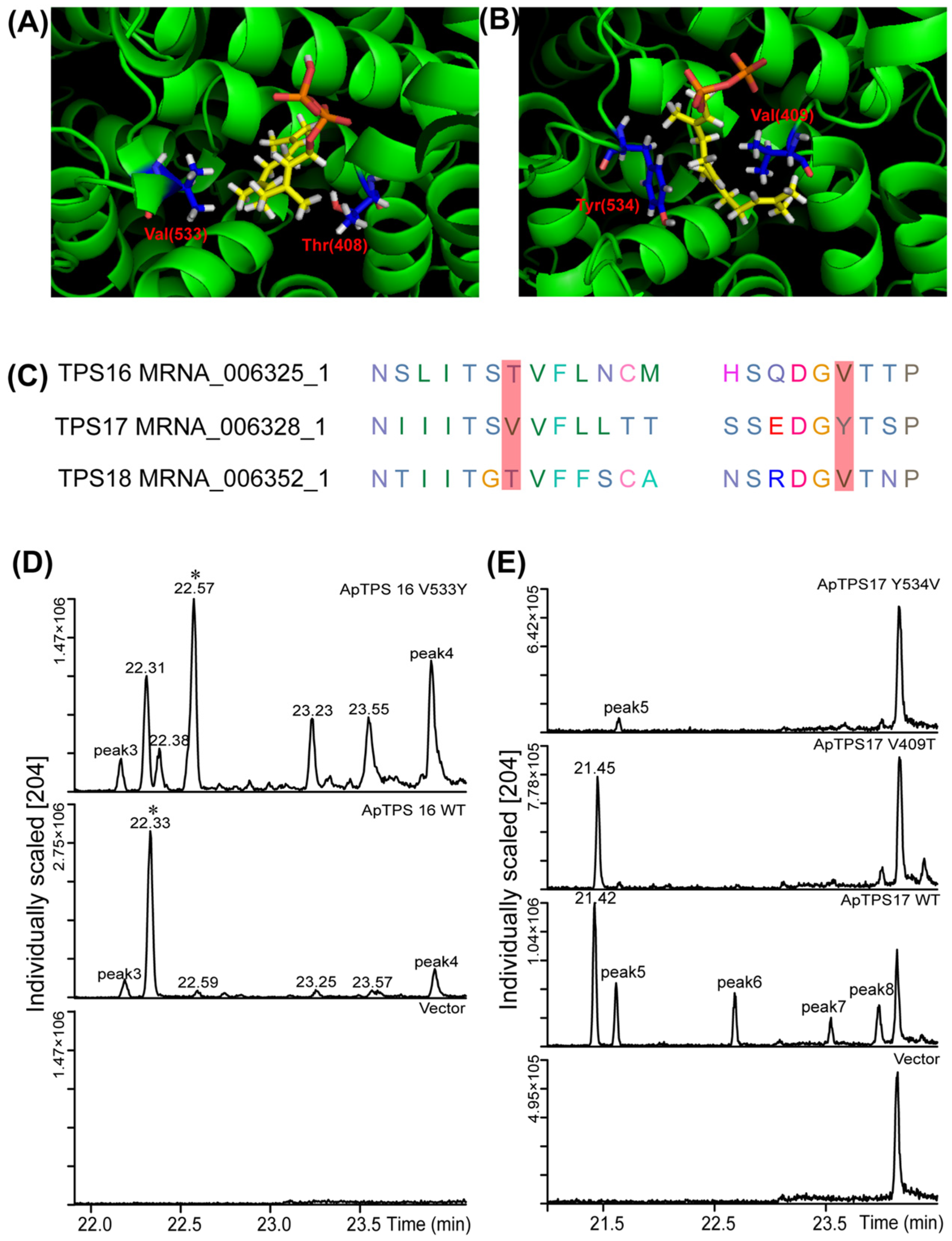
2.6. Key Residues and Site-Directed Mutagenesis of ApTPS
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Total RNA Isolation and cDNA Synthesis
4.3. ApTPS Screening in A. Paniculata and Bioinformatic Analysis
4.4. Cloning of ApTPS Coding Sequences
4.5. Functional Characterization of Class II ApTPSs
4.6. Functional Characterization of Class I ApTPSs
4.7. Homology Modelling and Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TPS | terpene synthase |
| diTPS | diterpene synthase |
| GGPP | geranylgeranyl pyrophosphate |
| GPP | geranyl pyrophosphate |
| FPP | farnesyl pyrophosphate |
| CDS | the full-length coding sequences |
| CPS | copalyl diphosphate synthase |
References
- Subramanian, R.; Zaini Asmawi, M.; Sadikun, A. A bitter plant with a sweet future? A comprehensive review of an oriental medicinal plant: Andrographis paniculata. Phytochem. Rev. 2012, 11, 39–75. [Google Scholar] [CrossRef]
- Hossain, M.S.; Urbi, Z.; Sule, A.; Rahman, K.H. Andrographis paniculata (burm. F.) wall. Ex nees: A review of ethnobotany, phytochemistry, and pharmacology. Sci. World J. 2014, 2014, 274905. [Google Scholar] [CrossRef] [PubMed]
- Allison, A.; Butcher, D.; Connolly, J.; Overton, K. Paniculides a, b, and c, bisabolenoid lactones from tissue cultures of andrographis paniculata. Chem. Commun. 1968, 23, 1493. [Google Scholar] [CrossRef]
- Mir, H.; Kapur, N.; Singh, R.; Sonpavde, G.; Lillard, J.W., Jr.; Singh, S. Andrographolide inhibits prostate cancer by targeting cell cycle regulators, cxcr3 and cxcr7 chemokine receptors. Cell Cycle 2016, 15, 819–826. [Google Scholar] [CrossRef]
- Wintachai, P.; Kaur, P.; Lee, R.C.H.; Ramphan, S.; Kuadkitkan, A.; Wikan, N.; Ubol, S.; Roytrakul, S.; Chu, J.J.H.; Smith, D.R. Activity of andrographolide against chikungunya virus infection. Sci. Rep. 2015, 5, 14179. [Google Scholar] [CrossRef]
- Chua, L.S. Review on liver inflammation and antiinflammatory activity of andrographis paniculata for hepatoprotection. Phytother. Res. 2014, 28, 1589–1598. [Google Scholar] [CrossRef]
- Wang, J.; Ma, Y.; Tang, J.; Lin, H.; Cui, G.; Tang, J.; Liu, J.; Su, P.; Zhao, Y.; Guo, J. Two cyp72 enzymes function as ent-labdane hydroxylases in the biosynthesis of andrographolide in andrographis paniculata. Plant Biotechnol. J. 2025, 23, 1136. [Google Scholar] [CrossRef]
- Kumar, A.; Hoskins, P.; Tinker, A. Dose-dense paclitaxel in advanced ovarian cancer. Clin. Oncol. 2015, 27, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiang, P.; Ye, M.; Kim, S.-H.; Jiang, C.; Lü, J. Tanshinones: Sources, pharmacokinetics and anti-cancer activities. Int. J. Mol. Sci. 2012, 13, 13621–13666. [Google Scholar] [CrossRef]
- Efferth, T. From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. In Seminars in Cancer Biology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 65–83. [Google Scholar] [CrossRef]
- Zhai, B.; Zeng, Y.; Zeng, Z.; Zhang, N.; Li, C.; Zeng, Y.; You, Y.; Wang, S.; Chen, X.; Sui, X. Drug delivery systems for elemene, its main active ingredient β-elemene, and its derivatives in cancer therapy. Int. J. Nanomed. 2018, 6279–6296. [Google Scholar] [CrossRef]
- Zhao, D.-D.; Jiang, L.-L.; Li, H.-Y.; Yan, P.-F.; Zhang, Y.-L. Chemical components and pharmacological activities of terpene natural products from the genus paeonia. Molecules 2016, 21, 1362. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Meyer-Gauen, G.; Croteau, R. Plant terpenoid synthases: Molecular biology and phylogenetic analysis. Proc. Natl. Acad. Sci. USA 1998, 95, 4126–4133. [Google Scholar] [CrossRef]
- Tholl, D.; Lee, S. Terpene specialized metabolism in arabidopsis thaliana. Arab. Book/Am. Soc. Plant Biol. 2011, 9, e0143. [Google Scholar]
- Hall, D.E.; Zerbe, P.; Jancsik, S.; Quesada, A.L.; Dullat, H.; Madilao, L.L.; Yuen, M.; Bohlmann, J. Evolution of conifer diterpene synthases: Diterpene resin acid biosynthesis in lodgepole pine and jack pine involves monofunctional and bifunctional diterpene synthases. Plant Physiol. 2013, 161, 600–616. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Zerbe, P.; Hamberger, B.; Yuen, M.M.; Chiang, A.; Sandhu, H.K.; Madilao, L.L.; Nguyen, A.; Hamberger, B.; Bach, S.S.; Bohlmann, J. Gene discovery of modular diterpene metabolism in nonmodel systems. Plant Physiol. 2013, 162, 1073–1091. [Google Scholar] [CrossRef]
- Garg, A.; Agrawal, L.; Misra, R.C.; Sharma, S.; Ghosh, S. Andrographis paniculata transcriptome provides molecular insights into tissue-specific accumulation of medicinal diterpenes. BMC Genom. 2015, 16, 659. [Google Scholar] [CrossRef]
- Shen, Q.; Li, L.; Jiang, Y.; Wang, Q. Functional characterization of ent-copalyl diphosphate synthase from andrographis paniculata with putative involvement in andrographolides biosynthesis. Biotechnol. Lett. 2016, 38, 131–137. [Google Scholar] [CrossRef]
- Sun, W.; Leng, L.; Yin, Q.; Xu, M.; Huang, M.; Xu, Z.; Zhang, Y.; Yao, H.; Wang, C.; Xiong, C. The genome of the medicinal plant andrographis paniculata provides insight into the biosynthesis of the bioactive diterpenoid neoandrographolide. Plant J. 2019, 97, 841–857. [Google Scholar] [CrossRef]
- Wendt, K.U.; Schulz, G.E. Isoprenoid biosynthesis: Manifold chemistry catalyzed by similar enzymes. Structure 1998, 6, 127–133. [Google Scholar] [CrossRef]
- Christianson, D.W. Unearthing the roots of the terpenome. Curr. Opin. Chem. Biol. 2008, 12, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Wendt, K.U.; Schulz, G.E.; Corey, E.J.; Liu, D.R. Enzyme mechanisms for polycyclic triterpene formation. Angew. Chem. Int. Ed. 2000, 39, 2812–2833. [Google Scholar] [CrossRef]
- Jia, Q.; Köllner, T.G.; Gershenzon, J.; Chen, F. Mtpsls: New terpene synthases in nonseed plants. Trends Plant Sci. 2018, 23, 121–128. [Google Scholar] [CrossRef]
- Misra, R.C.; Garg, A.; Roy, S.; Chanotiya, C.S.; Vasudev, P.G.; Ghosh, S. Involvement of an ent-copalyl diphosphate synthase in tissue-specific accumulation of specialized diterpenes in Andrographis paniculata. Plant Sci. 2015, 240, 50–64. [Google Scholar] [CrossRef]
- Sun, T.; Kamiya, Y. The arabidopsis ga1 locus encodes the cyclase ent-kaurene synthetase a of gibberellin biosynthesis. Plant Cell 1994, 6, 1509–1518. [Google Scholar]
- Gao, W.; Hillwig, M.L.; Huang, L.; Cui, G.; Wang, X.; Kong, J.; Yang, B.; Peters, R.J. A functional genomics approach to tanshinone biosynthesis provides stereochemical insights. Org. Lett. 2009, 11, 5170–5173. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Sun, T.-P.; Kawaide, H.; Kamiya, Y. The ga2 locus of arabidopsis thaliana encodes ent-kaurene synthase of gibberellin biosynthesis. Plant Physiol. 1998, 116, 1271–1278. [Google Scholar] [CrossRef]
- Fischer, M.J.; Meyer, S.; Claudel, P.; Bergdoll, M.; Karst, F. Metabolic engineering of monoterpene synthesis in yeast. Biotechnol. Bioeng. 2011, 108, 1883–1892. [Google Scholar] [CrossRef]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A. The b73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef]
- Lim, J.C.W.; Chan, T.K.; Ng, D.S.; Sagineedu, S.R.; Stanslas, J.; Wong, W.F. Andrographolide and its analogues: Versatile bioactive molecules for combating inflammation and cancer. Clin. Exp. Pharmacol. Physiol. 2012, 39, 300–310. [Google Scholar] [CrossRef]
- Sareer, O.; Ahmad, S.; Umar, S. Andrographis paniculata: A critical appraisal of extraction, isolation and quantification of andrographolide and other active constituents. Nat. Prod. Res. 2014, 28, 2081–2101. [Google Scholar] [CrossRef] [PubMed]
- Whittington, D.A.; Wise, M.L.; Urbansky, M.; Coates, R.M.; Croteau, R.B.; Christianson, D.W. Bornyl diphosphate synthase: Structure and strategy for carbocation manipulation by a terpenoid cyclase. Proc. Natl. Acad. Sci. USA 2002, 99, 15375–15380. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.C.; Youn, B.; Zhao, Y.; Santhamma, B.; Coates, R.M.; Croteau, R.B.; Kang, C. Structure of limonene synthase, a simple model for terpenoid cyclase catalysis. Proc. Natl. Acad. Sci. USA 2007, 104, 5360–5365. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, P.R.; Xu, M.; Jin, Y.; Coates, R.M.; Peters, R.J. Identification of syn-pimara-7, 15-diene synthase reveals functional clustering of terpene synthases involved in rice phytoalexin/allelochemical biosynthesis. Plant Physiol. 2004, 135, 2098–2105. [Google Scholar] [CrossRef]
- Trapp, S.C.; Croteau, R.B. Genomic organization of plant terpene synthases and molecular evolutionary implications. Genetics 2001, 158, 811–832. [Google Scholar] [CrossRef]
- Lin, H.; Wang, J.; Qi, M.; Guo, J.; Rong, Q.; Tang, J.; Wu, Y.; Ma, X.; Huang, L. Molecular cloning and functional characterization of multiple nadph-cytochrome p450 reductases from Andrographis paniculata. Int. J. Biol. Macromol. 2017, 102, 208–217. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. Meme suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37 (Suppl. S2), W202–W208. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Mega, K.T. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, Y.; Huang, L.; Zhang, X. Production of miltiradiene by metabolically engineered saccharomyces cerevisiae. Biotechnol. Bioeng. 2012, 109, 2845–2853. [Google Scholar] [CrossRef]
- Guo, J.; Ma, X.; Cai, Y.; Ma, Y.; Zhan, Z.; Zhou, Y.J.; Liu, W.; Guan, M.; Yang, J.; Cui, G. Cytochrome p450 promiscuity leads to a bifurcating biosynthetic pathway for tanshinones. New Phytol. 2016, 210, 525–534. [Google Scholar] [CrossRef]
- Song, X.; Chen, Y.; Cui, G.; Jin, B.; Wang, J.; Ma, Y.; Xia, M.; Zhang, Y.; Guo, J.; Yuan, S. Functional identification of the diterpene synthases exploring the landscape of diterpene structural diversity in Isodon. Plant Physiol. Biochem. 2025, 222, 109677. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, Z.; Shen, S.; Lin, L.; Li, T.; Tian, B.; Hua, Y. Identification and characterization of the geranylgeranyl diphosphate synthase in deinococcus radiodurans. Lett. Appl. Microbiol. 2014, 58, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N.L.; Heskes, A.M.; Hamberger, B.; Olsen, C.E.; Hallström, B.M.; Andersen-Ranberg, J.; Hamberger, B. The terpene synthase gene family in tripterygium wilfordii harbors a labdane-type diterpene synthase among the monoterpene synthase tps-b subfamily. Plant J. 2017, 89, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Guan, H.; Zhang, Y.; Wang, X.; Gao, L.; Zhao, Y.; Hu, T.; Zhou, J.; Ma, B.; Tu, L. Probing the single key amino acid responsible for the novel catalytic function of ent-kaurene oxidase supported by nadph-cytochrome p450 reductases in tripterygium wilfordii. Front. Plant Sci. 2017, 8, 1756. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.; Ma, Y.; Zhao, Y.; Ma, X.; Wang, J. Characterization of Terpene Synthases Reveals the Diversity of Terpenoids in Andrographis paniculata. Molecules 2025, 30, 2208. https://doi.org/10.3390/molecules30102208
Tang J, Ma Y, Zhao Y, Ma X, Wang J. Characterization of Terpene Synthases Reveals the Diversity of Terpenoids in Andrographis paniculata. Molecules. 2025; 30(10):2208. https://doi.org/10.3390/molecules30102208
Chicago/Turabian StyleTang, Junhao, Ying Ma, Yujun Zhao, Xiaohui Ma, and Jian Wang. 2025. "Characterization of Terpene Synthases Reveals the Diversity of Terpenoids in Andrographis paniculata" Molecules 30, no. 10: 2208. https://doi.org/10.3390/molecules30102208
APA StyleTang, J., Ma, Y., Zhao, Y., Ma, X., & Wang, J. (2025). Characterization of Terpene Synthases Reveals the Diversity of Terpenoids in Andrographis paniculata. Molecules, 30(10), 2208. https://doi.org/10.3390/molecules30102208