
Nootkatone Alleviates Type 2 Diabetes in db/db Mice Through AMPK Activation and ERK Inhibition: An Integrated In Vitro and In Vivo Study

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
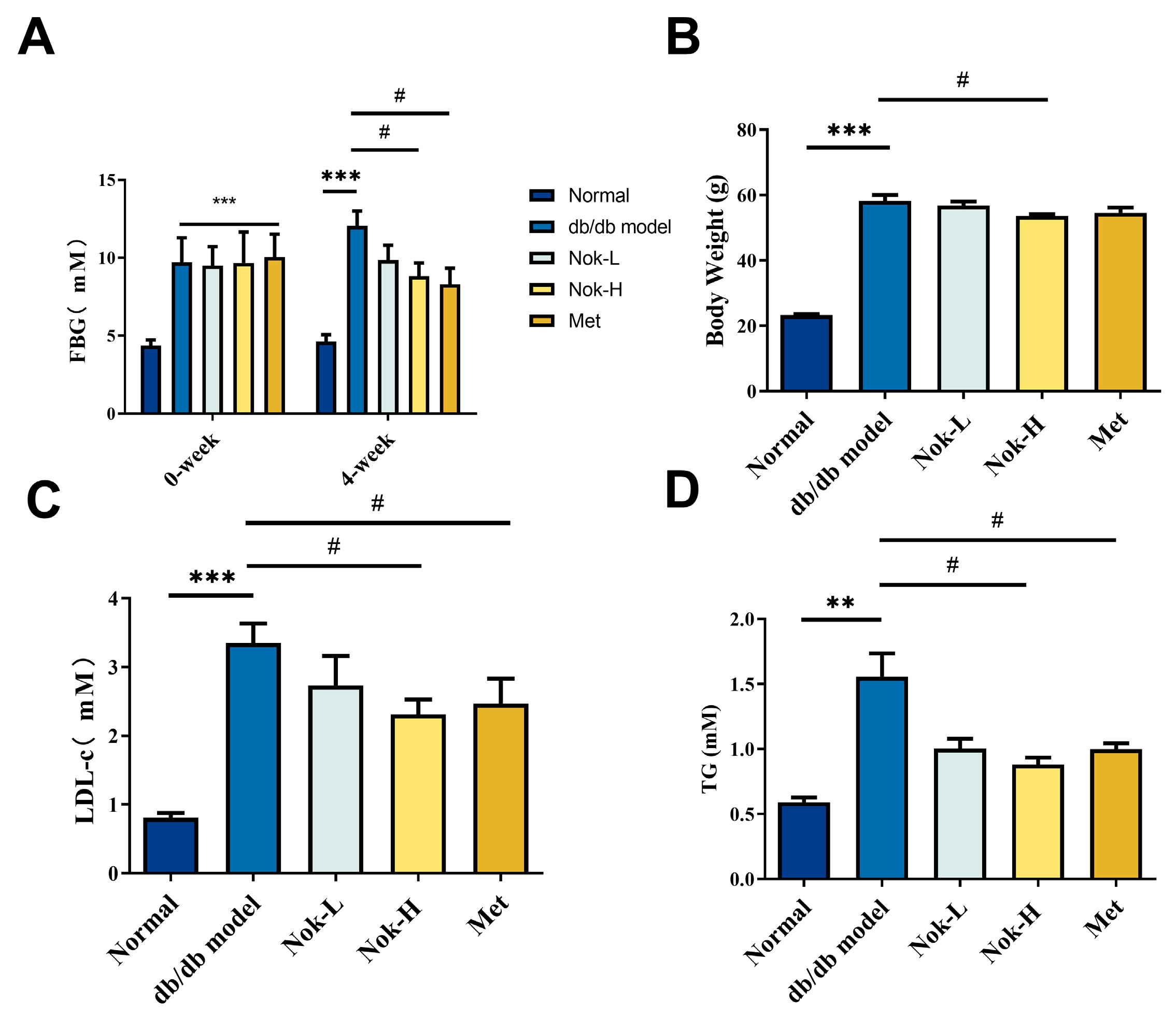
2.1. Nok Improves Glucose and Lipid Metabolism and Reduces Body Weight in db/db Mice
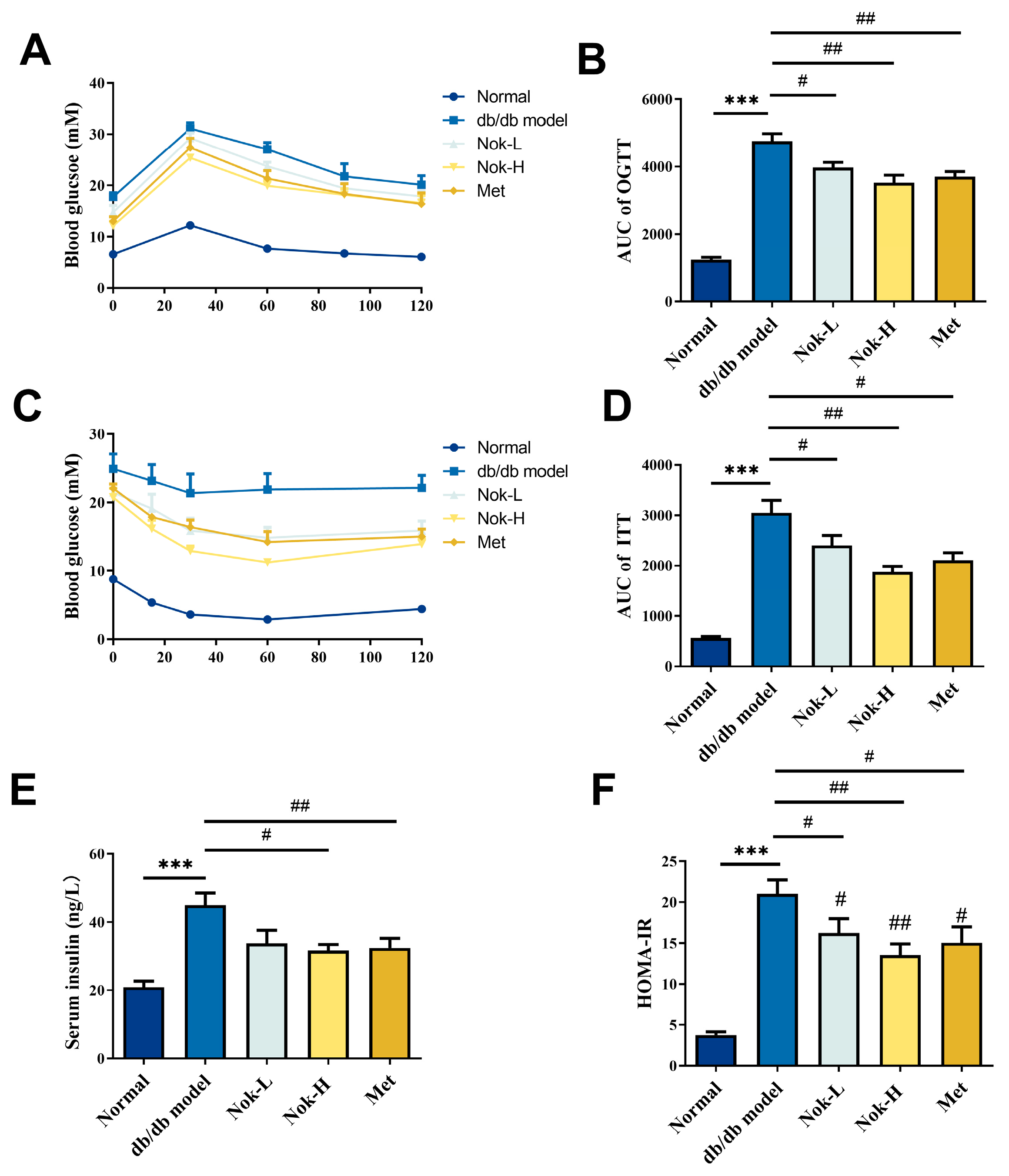
2.2. Nok Improves Glucose Tolerance and Insulin Resistance in db/db Diabetic Mice
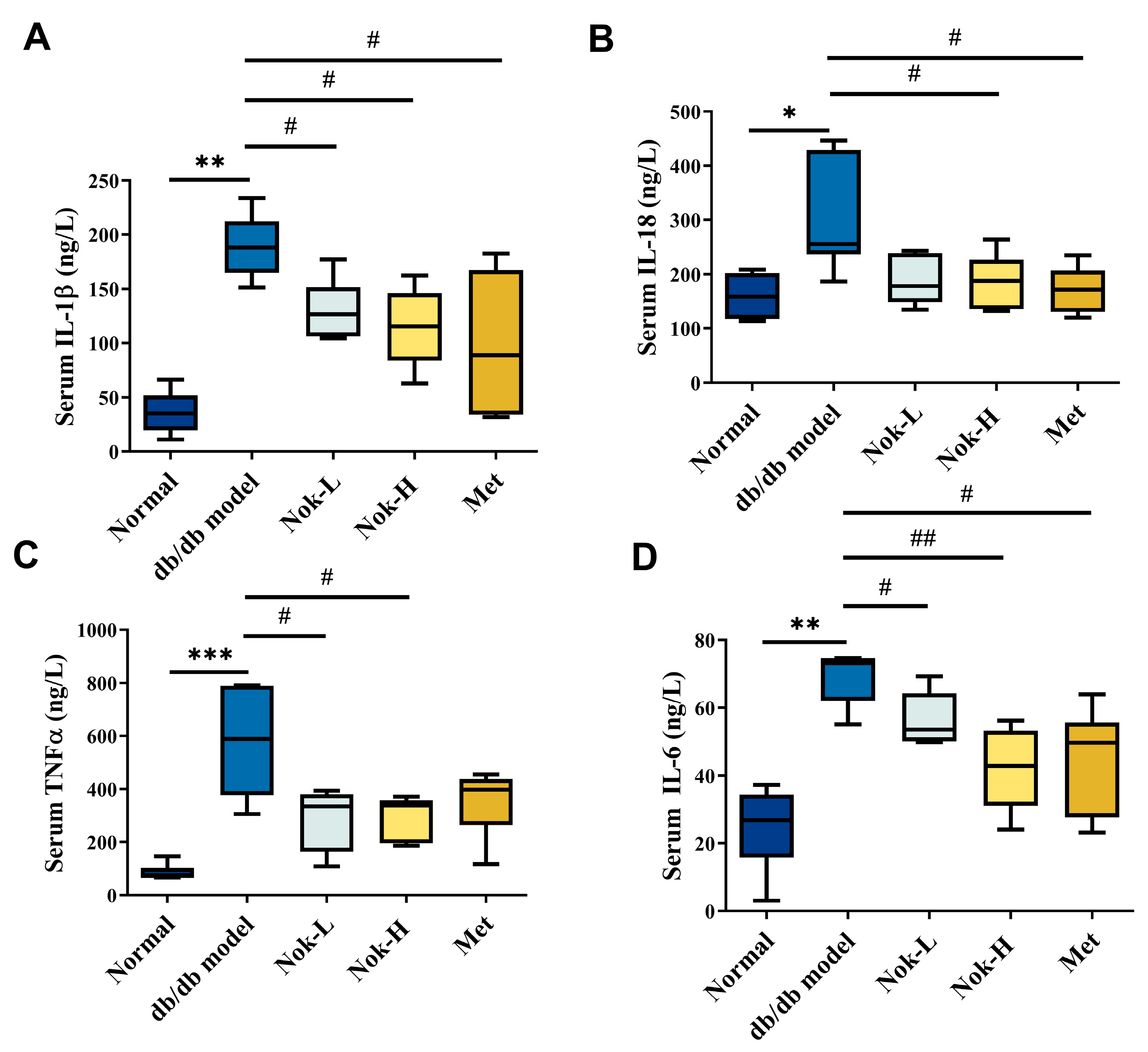
2.3. Nok Significantly Reduces Inflammation in db/db Diabetic Mice
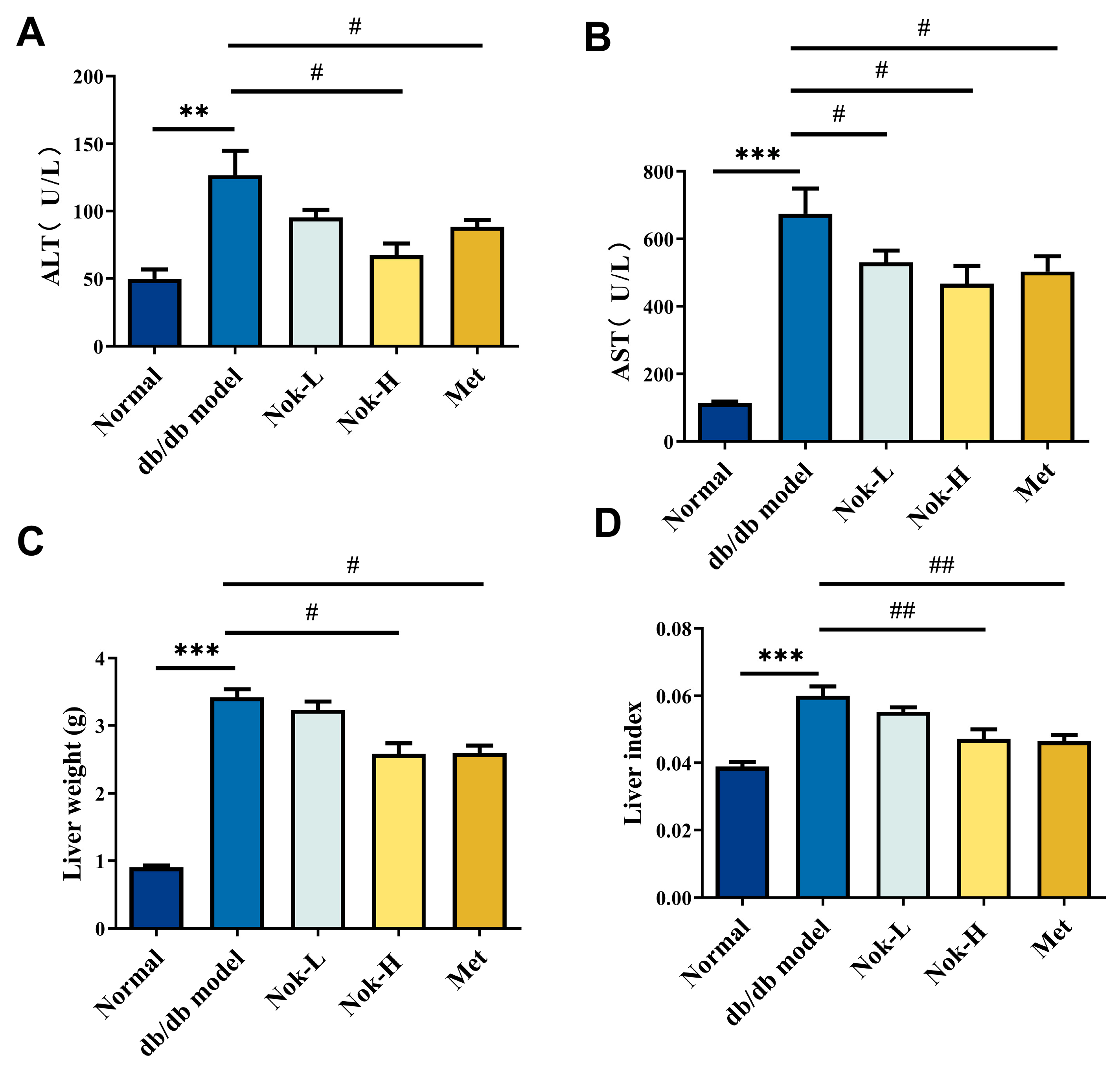
2.4. Nok Improves Liver Function in db/db Diabetic Mice
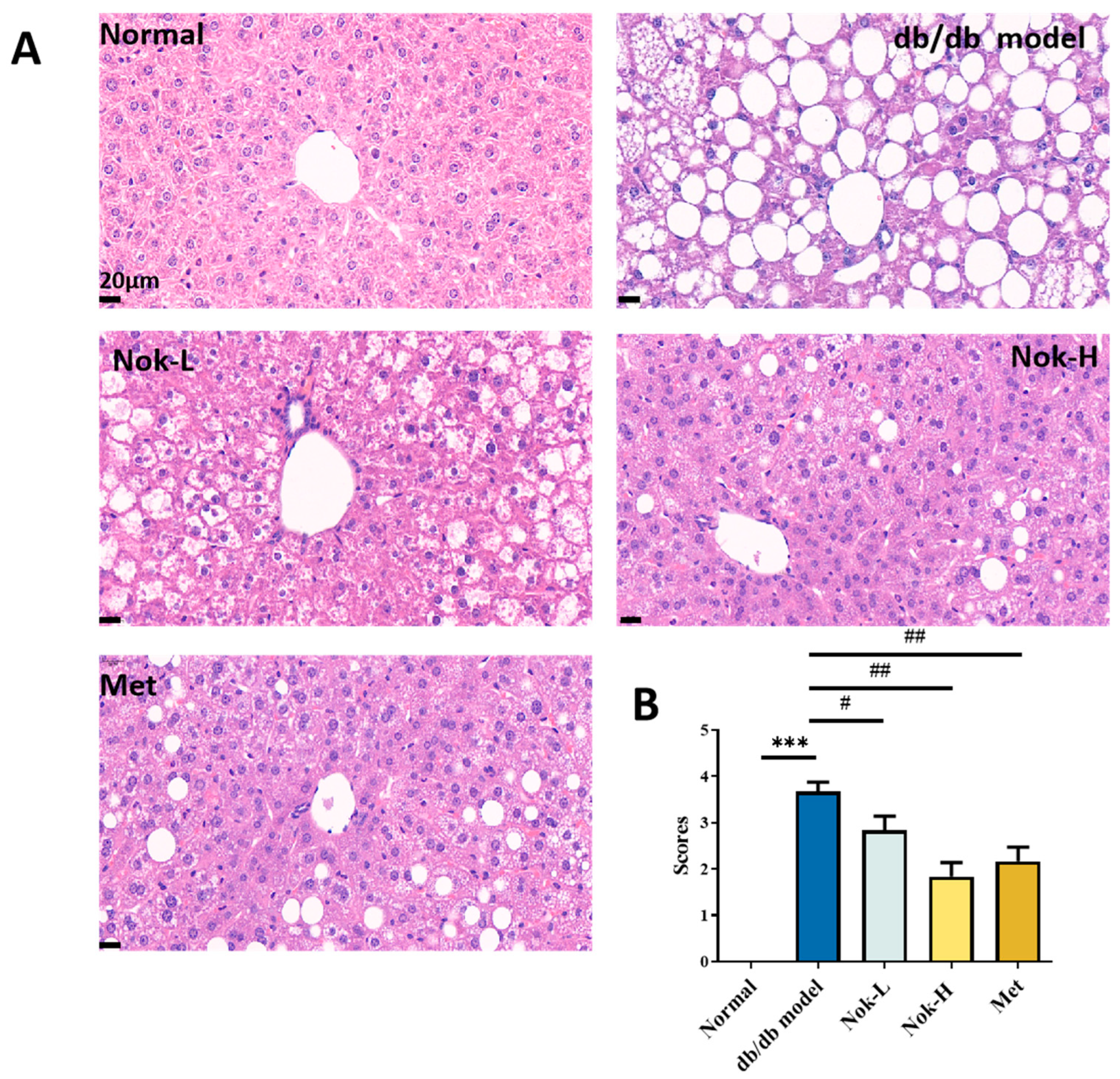
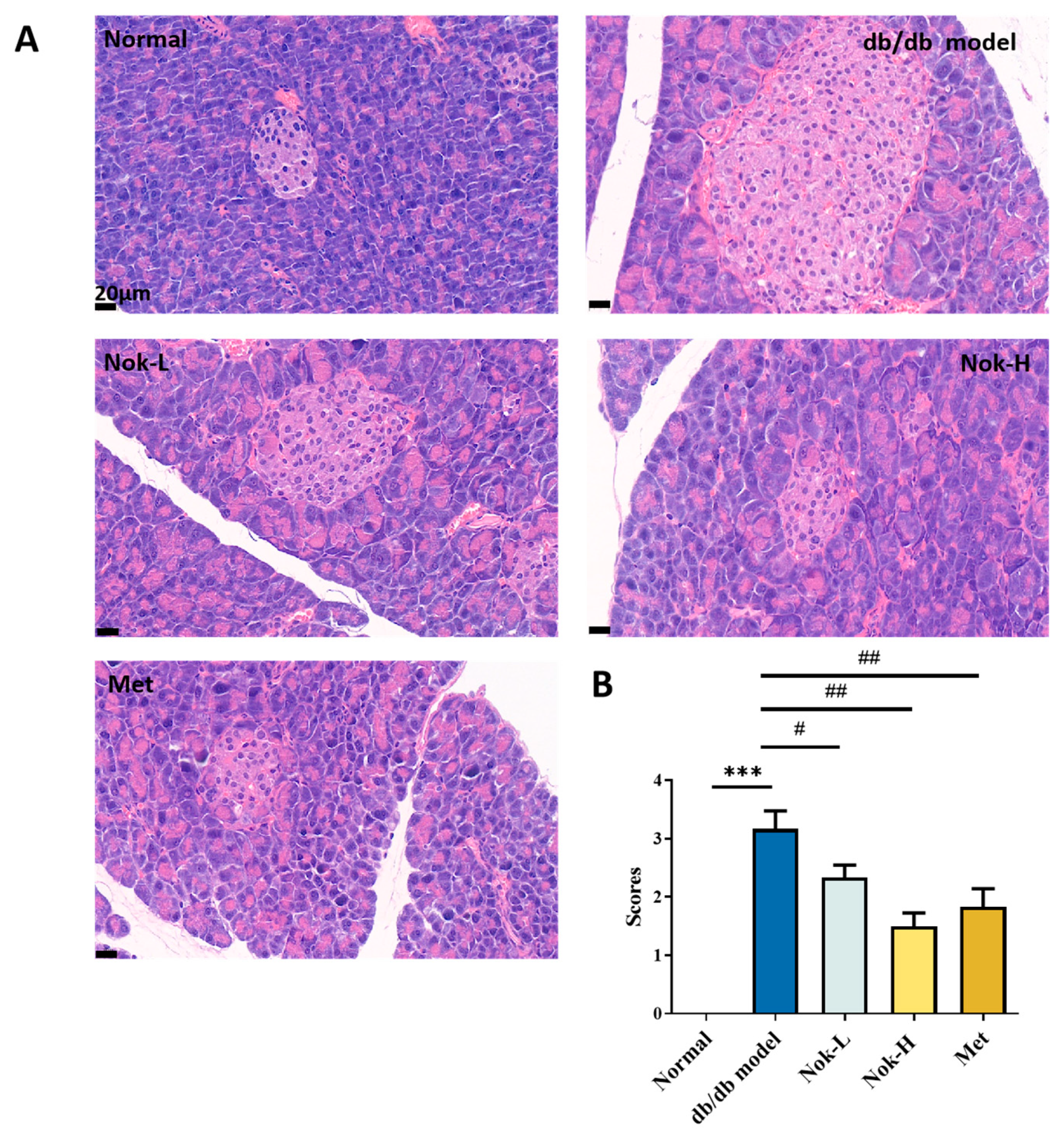
2.5. Nok Improves Pathological Changes in the Liver and Pancreas of db/db Mice
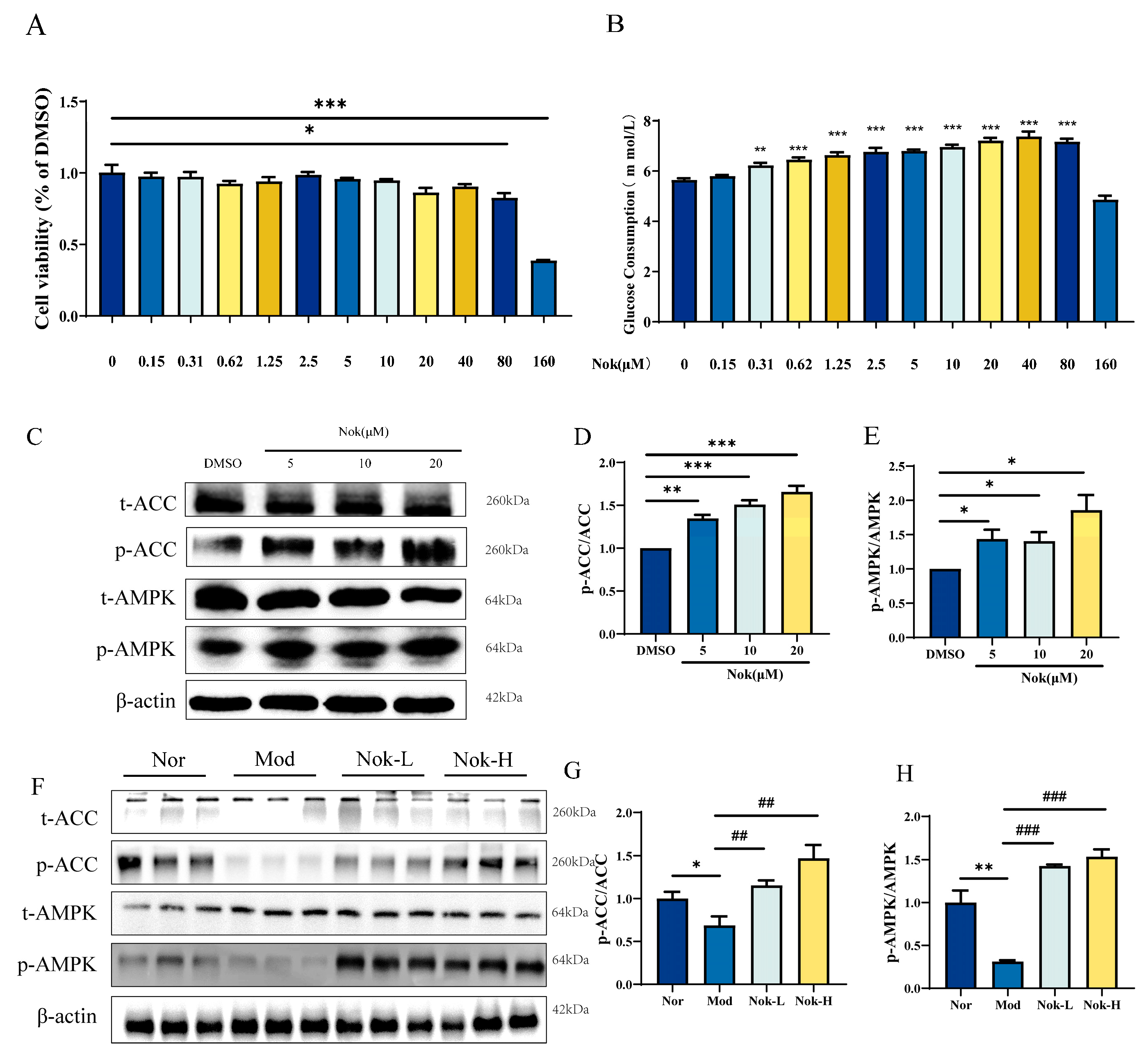
2.6. The Effects of NOK on Glucose Metabolism and AMPK Activation
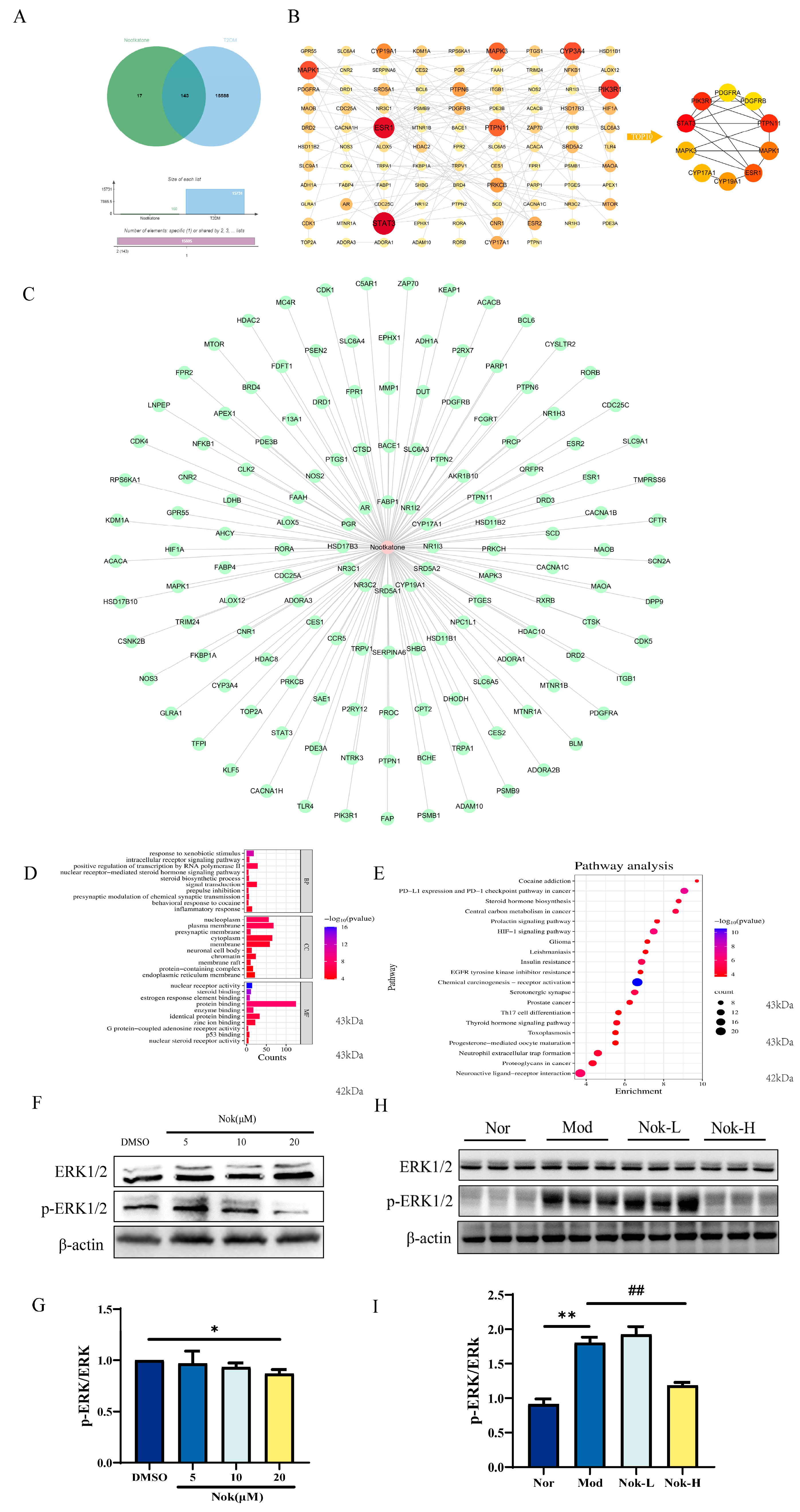
2.7. Network Pharmacology Prediction and Experimental Validation of Nok’s Targets
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Experimental Animals
4.3. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT)
4.4. HE Staining and Histopathological Examination
4.5. Drug Database and Related Target Prediction
4.6. Prediction of T2DM Related Targets from Online Database
4.7. Cell Culture and Treatment
4.8. Cytotoxicity and Glucose Consumption Assay
4.9. Western Blot
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Werfalli, M.; Raubenheimer, P.J.; Engel, M.; Musekiwa, A.; Bobrow, K.; Peer, N.; Hoegfeldt, C.; Kalula, S.; Kengne, A.P.; Levitt, N.S. The effectiveness of peer and community health worker-led self-management support programs for improving diabetes health-related outcomes in adults in low- and-middle-income countries: A systematic review. Syst. Rev. 2020, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Jia, W. Diabetes in China: Epidemiology and Genetic Risk Factors and Their Clinical Utility in Personalized Medication. Diabetes 2018, 67, 3–11. [Google Scholar] [CrossRef]
- Sanchez-Rangel, E.; Inzucchi, S.E. Metformin: Clinical use in type 2 diabetes. Diabetologia 2017, 60, 1586–1593. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Ma, T.; Xie, Y. Research progress on Alpinia oxyphylla in the treatment of diabetic nephropathy. Front. Pharmacol. 2024, 15, 1390672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zheng, Y.; Hu, X.; Hu, X.; Lv, W.; Lv, D.; Chen, J.; Wu, M.; Song, Q.; Shentu, J. Ethnopharmacological uses, phytochemistry, biological activities, and therapeutic applications of Alpinia oxyphylla Miquel: A review. J. Ethnopharmacol. 2018, 224, 149–168. [Google Scholar] [CrossRef]
- Deng, X.; Ye, Z.; Duan, J.; Chen, F.; Zhi, Y.; Huang, M.; Huang, M.; Cheng, W.; Dou, Y.; Kuang, Z.; et al. Complete pathway elucidation and heterologous reconstitution of (+)-nootkatone biosynthesis from Alpinia oxyphylla. New Phytol. 2024, 241, 779–792. [Google Scholar] [CrossRef]
- Hung, L.V.M.; Moon, J.Y.; Ryu, J.Y.; Cho, S.K. Nootkatone, an AMPK activator derived from grapefruit, inhibits KRAS downstream pathway and sensitizes non-small-cell lung cancer A549 cells to adriamycin. Phytomedicine 2019, 63, 153000. [Google Scholar] [CrossRef]
- Yan, T.; Li, F.; Xiong, W.; Wu, B.; Xiao, F.; He, B.; Jia, Y. Nootkatone improves anxiety- and depression-like behavior by targeting hyperammonemia-induced oxidative stress in D-galactosamine model of liver injury. Environ. Toxicol. 2021, 36, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabTolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar]
- Carling, D. AMPK signalling in health and disease. Curr. Opin. Cell Biol. 2017, 45, 31–37. [Google Scholar] [CrossRef]
- Trefts, E.; Shaw, R.J. AMPK: Restoring metabolic homeostasis over space and time. Mol. Cell. 2021, 81, 3677–3690. [Google Scholar] [CrossRef]
- Yonamine, C.Y.; Pinheiro-Machado, E.; Michalani, M.L.; Alves-Wagner, A.B.; Esteves, J.V.; Freitas, H.S.; Machado, U.F. Resveratrol Improves Glycemic Control in Type 2 Diabetic Obese Mice by Regulating Glucose Transporter Expression in Skeletal Muscle and Liver. Molecules 2017, 22, 1180. [Google Scholar] [CrossRef] [PubMed]
- Yong, Z.; Yang, Y.; Zhou, Z.; Tan, Y.; Dong, L.; Li, Y.; Lu, W.; Wu, C.; Zhang, X. Laurolitsine ameliorates type 2 diabetes by regulating the hepatic LKB1-AMPK pathway and gut microbiota. Phytomedicine 2022, 106, 154423. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Hardie, D.G. AMPK: Sensing Glucose as well as Cellular Energy Status. Cell Metab. 2014, 27, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, M.D.; Galic, S.; Marcinko, K.; Sikkema, S.; Pulinilkunnil, T.; Chen, Z.-P.; O’Neill, H.M.; Ford, R.J.; Palanivel, R.; O’Brien, M.; et al. Single phosphorylation sites in Acc1 and Acc2 regulate lipid homeostasis and the insulin-sensitizing effects of metformin. Nat. Med. 2013, 19, 1649–1654. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, X.; Chen, H.; You, L.; Zhang, T.; Cheng, M.; Yao, Y.; Pan, X.; Yang, X. Mulberry extract ameliorates T2DM-related symptoms via AMPK pathway in STZ-HFD-induced C57BL/6J mice. J. Ethnopharmacol. 2023, 313, 116475. [Google Scholar] [CrossRef]
- Xiang, H.C.; Lin, L.X.; Hu, X.F.; Zhu, H.; Li, H.P.; Zhang, R.Y.; Hu, L.; Liu, W.T.; Zhao, Y.L.; Shu, Y.; et al. AMPK activation attenuates inflammatory pain through inhibiting NF-κB activation and IL-1β expression. J. Neuroinflammat. 2019, 16, 34. [Google Scholar] [CrossRef]
- Vahidi Ferdowsi, P.; Ahuja, K.D.K.; Beckett, J.M.; Myers, S. TRPV1 Activation by Capsaicin Mediates Glucose Oxidation and ATP Production Independent of Insulin Signalling in Mouse Skeletal Muscle Cells. Cells 2021, 10, 1560. [Google Scholar] [CrossRef]
- Lontchi-Yimagou, E.; Sobngwi, E.; Matsha, T.E.; Kengne, A.P. Diabetes mellitus and inflammation. Curr. Diab. Rep. 2013, 13, 435–444. [Google Scholar] [CrossRef]
- Yu, L.; Xu, M.; Yan, Y.; Huang, S.; Yuan, M.; Cui, B.; Lv, C.; Zhang, Y.; Wang, H.; Jin, X.; et al. ZFYVE28 mediates insulin resistance by promoting phosphorylated insulin receptor degradation via increasing late endosomes production. Nat. Commun. 2023, 14, 6833. [Google Scholar] [CrossRef]
- Ahmadzadeh, A.M.; Aliabadi, M.M.; Mirheidari, S.B.; Hamedi-Asil, M.; Garousi, S.; Mottahedi, M.; Sahebkar, A. Beneficial effects of resveratrol on diabetes mellitus and its complications: Focus on mechanisms of action. Naunyn Schmiedebergs Arch. Pharmacol. 2024, 398, 2407–2442. [Google Scholar] [CrossRef] [PubMed]
- Yong, Z.; Huang, Z.; Zhou, Z.; Ma, N.; Wang, R.; Chen, M.; He, X. Nootkatone, a Sesquiterpene Ketone From Alpiniae oxyphyllae Fructus, Ameliorates Metabolic-Associated Fatty Liver by Regulating AMPK and MAPK Signaling. Front. Pharmacol. 2022, 13, 909280. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zheng, L.; Chen, M.; Li, R.; Yu, Y.; Qiao, L.; Liu, J.; Zhang, X.; Zhang, Y.; Zhang, Y.; et al. Nootkatone Alleviates Type 2 Diabetes in db/db Mice Through AMPK Activation and ERK Inhibition: An Integrated In Vitro and In Vivo Study. Molecules 2025, 30, 2111. https://doi.org/10.3390/molecules30102111
Li Y, Zheng L, Chen M, Li R, Yu Y, Qiao L, Liu J, Zhang X, Zhang Y, Zhang Y, et al. Nootkatone Alleviates Type 2 Diabetes in db/db Mice Through AMPK Activation and ERK Inhibition: An Integrated In Vitro and In Vivo Study. Molecules. 2025; 30(10):2111. https://doi.org/10.3390/molecules30102111
Chicago/Turabian StyleLi, Yingjie, Linlin Zheng, Mimi Chen, Ruodi Li, Yansu Yu, Lu Qiao, Jialu Liu, Xiaopo Zhang, Yong Zhang, Yuxin Zhang, and et al. 2025. "Nootkatone Alleviates Type 2 Diabetes in db/db Mice Through AMPK Activation and ERK Inhibition: An Integrated In Vitro and In Vivo Study" Molecules 30, no. 10: 2111. https://doi.org/10.3390/molecules30102111
APA StyleLi, Y., Zheng, L., Chen, M., Li, R., Yu, Y., Qiao, L., Liu, J., Zhang, X., Zhang, Y., Zhang, Y., & Zheng, W. (2025). Nootkatone Alleviates Type 2 Diabetes in db/db Mice Through AMPK Activation and ERK Inhibition: An Integrated In Vitro and In Vivo Study. Molecules, 30(10), 2111. https://doi.org/10.3390/molecules30102111