


Gypenosides Attenuates CORT-Induced Ferroptosis via Inhibiting TNF-α/NF-κB Signaling Pathway in PC12 Cells

Abstract
1. Introduction
2. Results
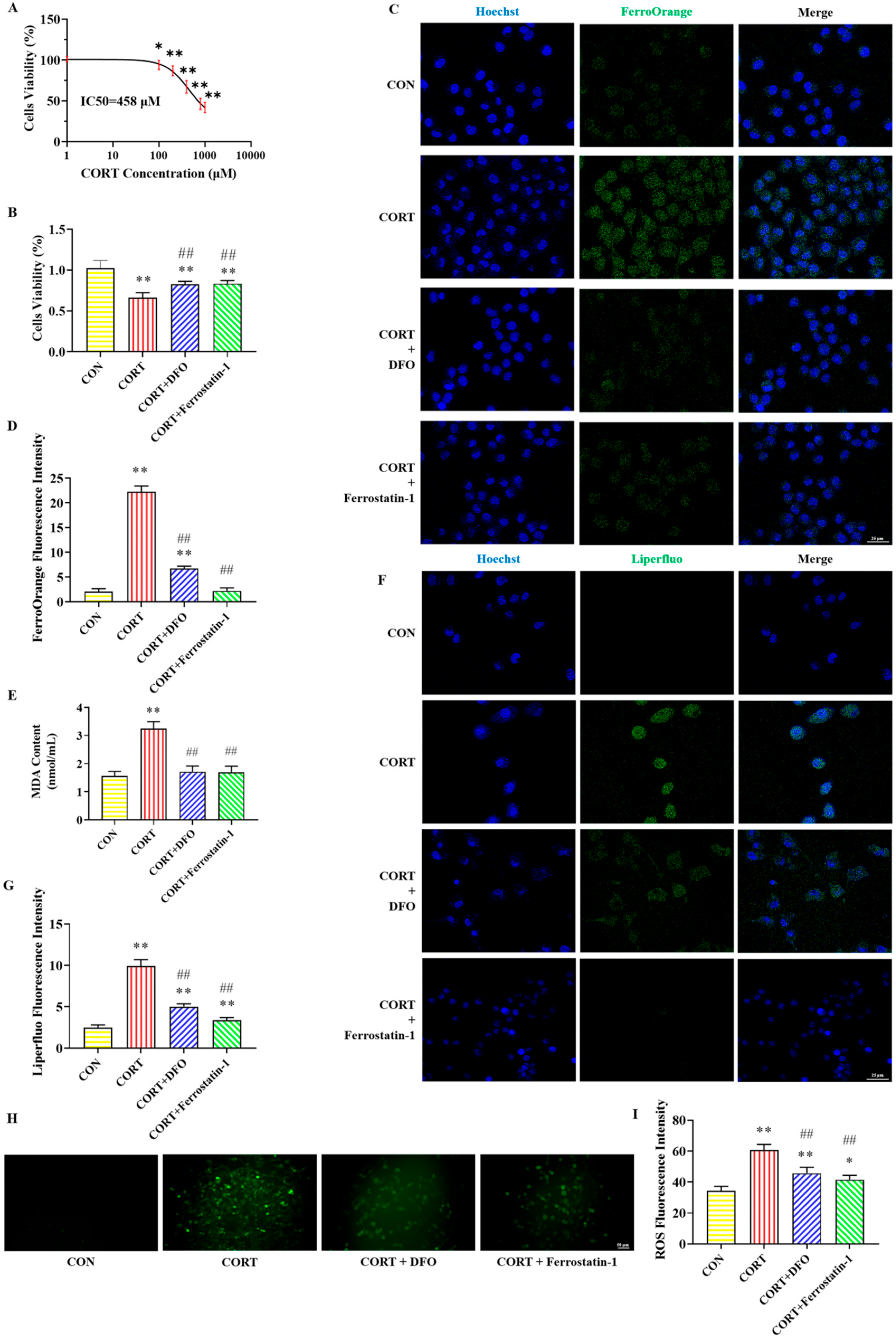
2.1. Effects of DFO and Ferrostatin-1 on CORT-Induced PC12 Cells Viability
2.2. Effect of DFO and Ferrostatin-1 on CORT-Induced PC12 Cells Iron Accumulation
2.3. Effect of DFO and Ferrostatin-1 on CORT-Induced Lipid Peroxidation in PC12 Cells
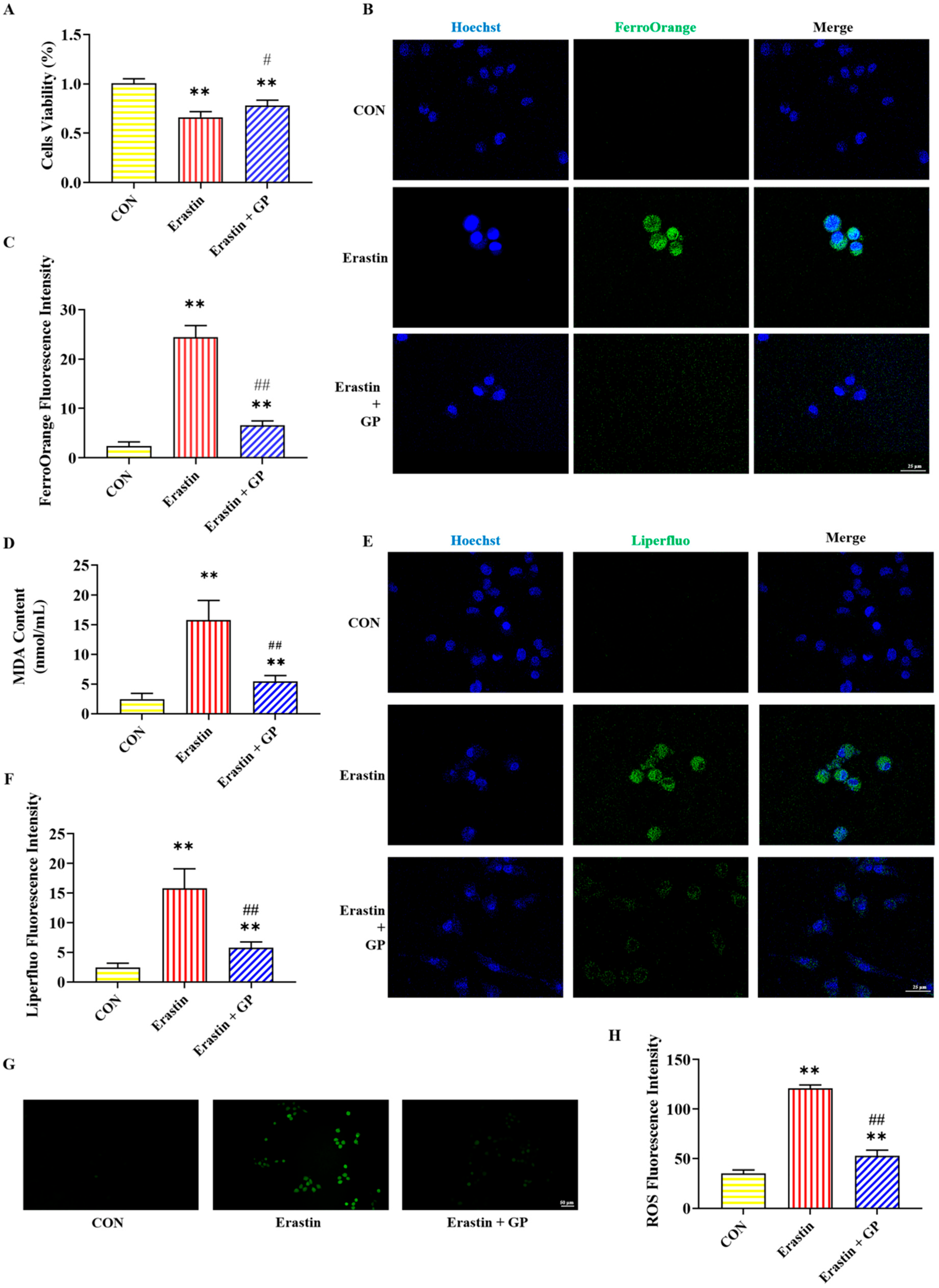
2.4. Effect of GPs on Erastin-Induced Viability of PC12 Cells
2.5. Effect of GPs on Erastin-Induced Iron Accumulation in PC12 Cells
2.6. Effect of GPs on Erastin-Induced Lipid Peroxidation in PC12 Cells
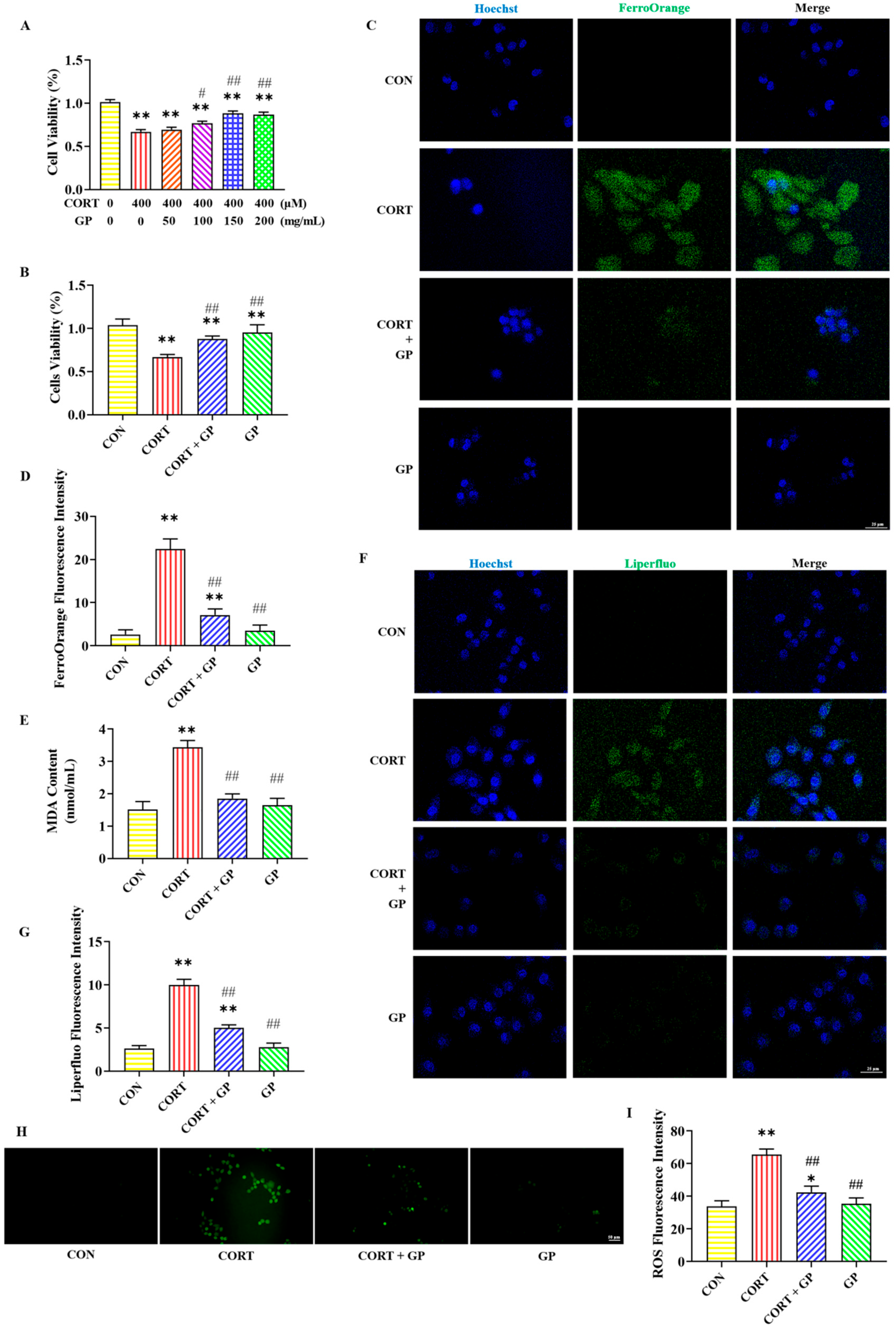
2.7. Effect of GPs on CORT-Induced PC12 Cells Viability
2.8. Effect of GPs on CORT-Induced PC12 Cells Iron Accumulation
2.9. Effect of GPs on CORT-Induced PC12 Cells Lipid Peroxidation
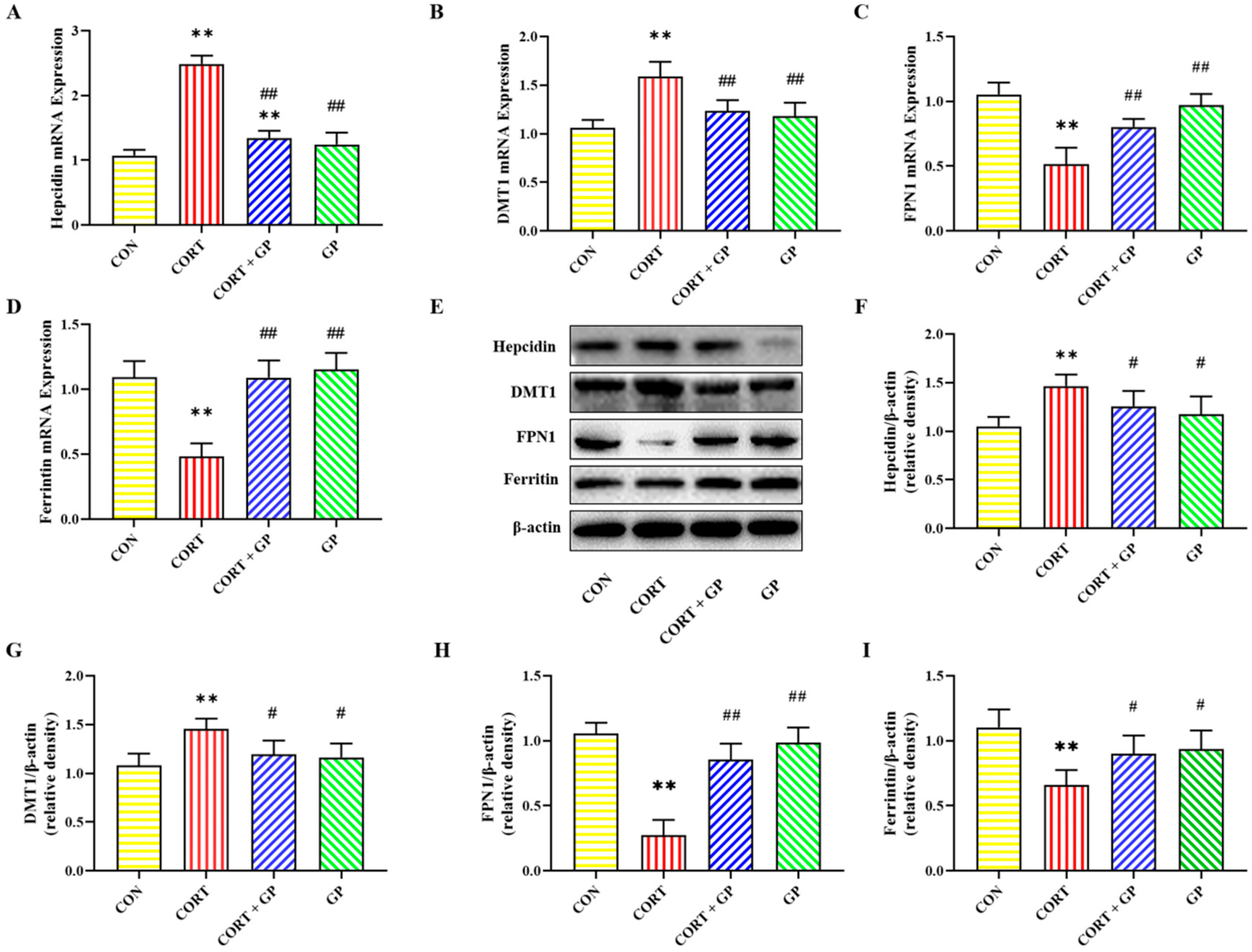
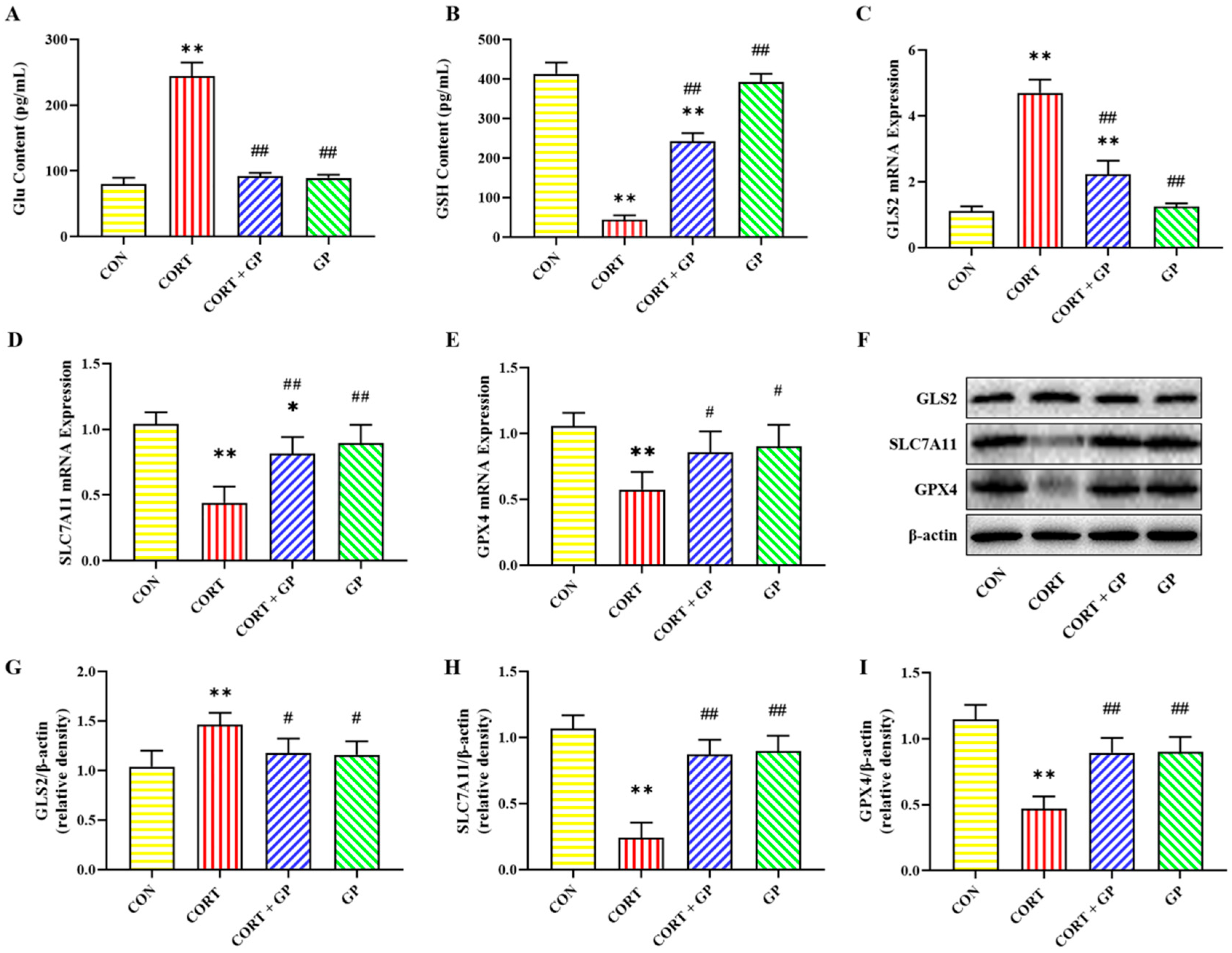
2.10. Effect of GPs on CORT-Induced PC12 Cells Iron Homeostasis
2.11. Effect of GPs on CORT-Induced PC12 Cells Glu Metabolism
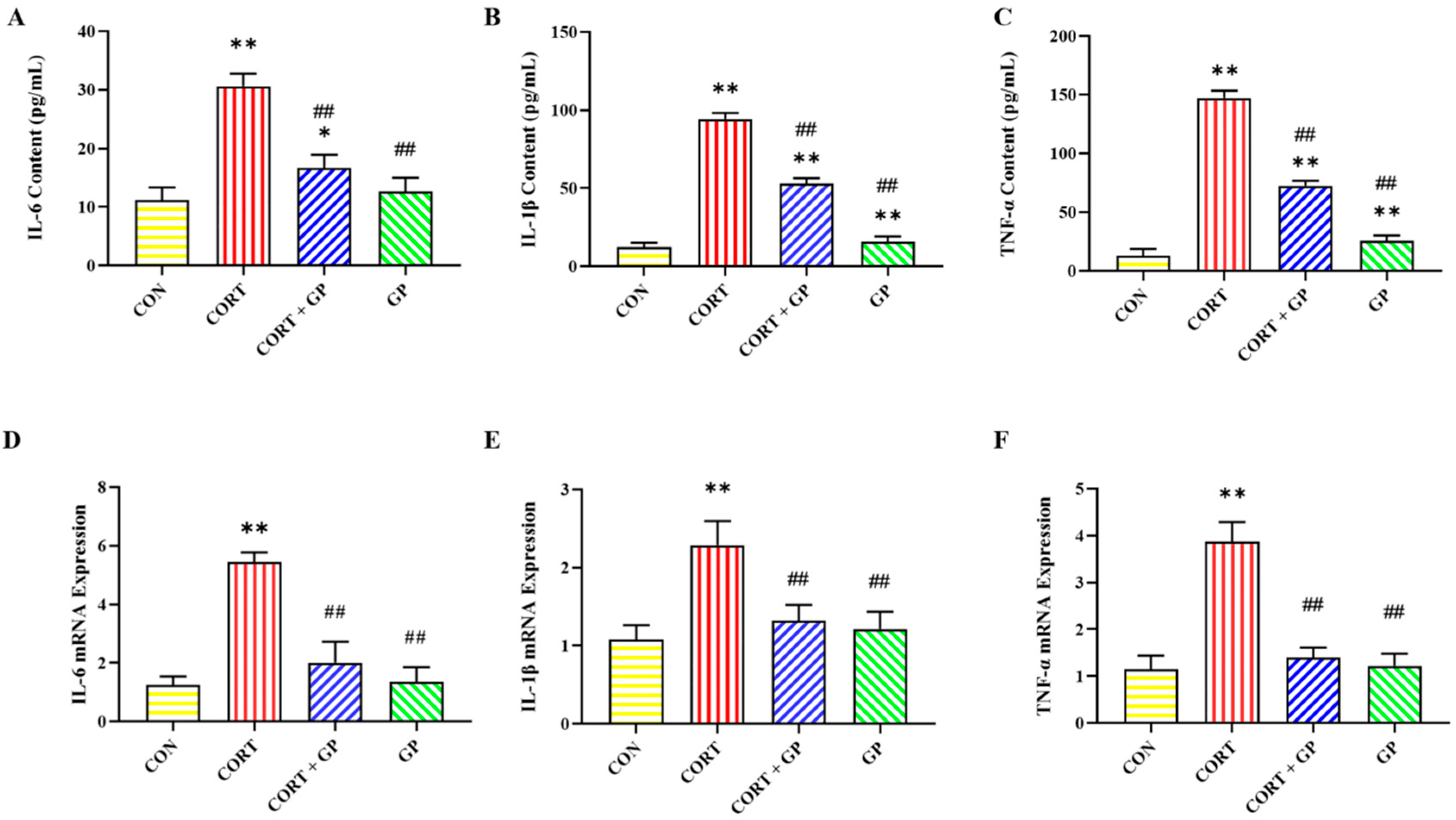
2.12. Effect of GPs on CORT-Induced Release of Inflammatory Cytokines in PC12 Cells
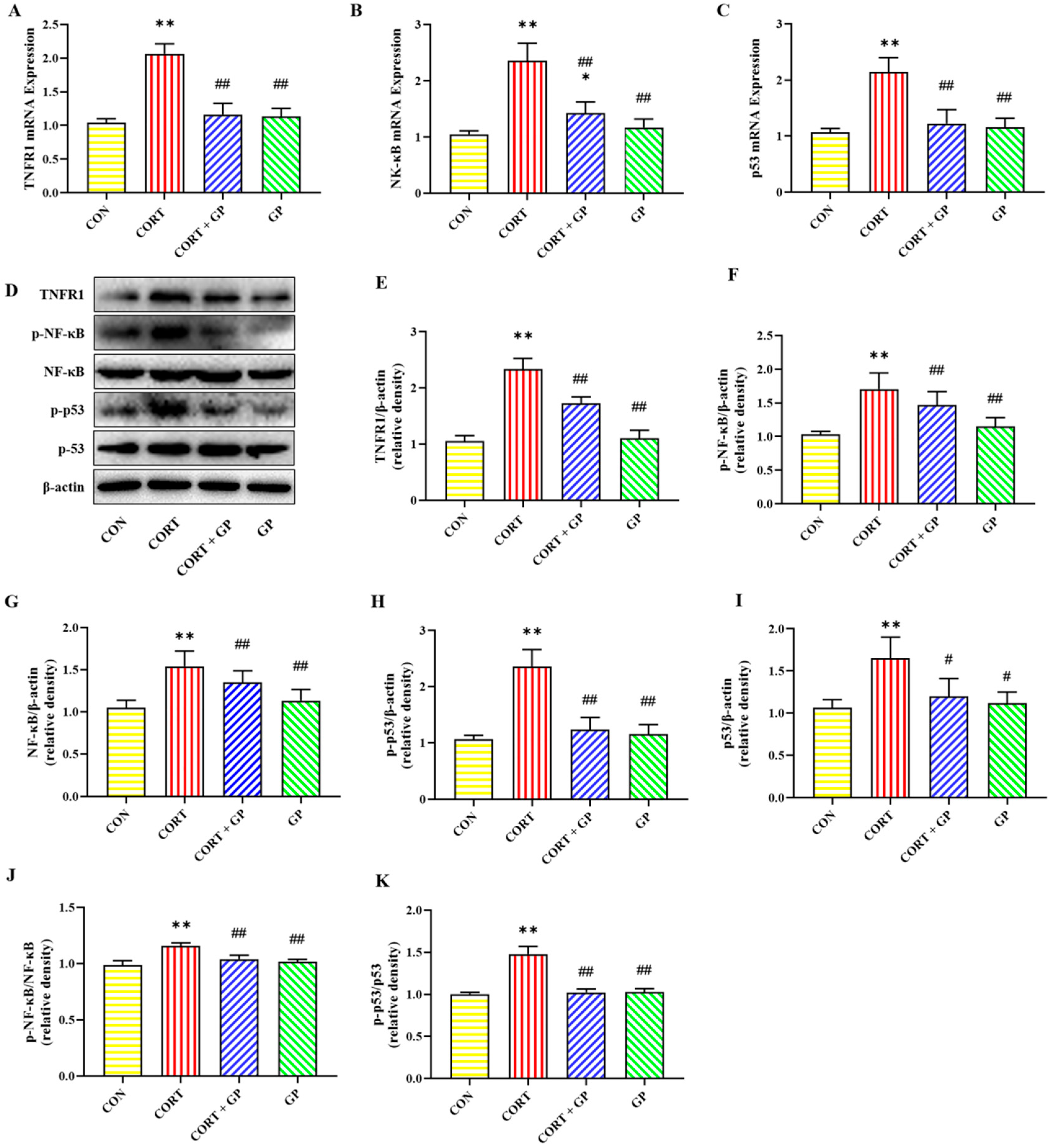
2.13. Effect of GPs on CORT-Induced TNF-α/NF-κB Signaling Pathway in PC12 Cells
3. Materials and Methods
3.1. Cell Culture and Drug Treatments
3.2. Cell Viability Assay
3.3. ELISA Kits Assay
3.4. Iron Accumulation Assay
3.5. LPO Fluorescence Intensity Assa0079
3.6. Western Blot Analysis
3.7. Quantitative Real-Time PCR Analysis
3.8. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Menneson, S.; Ménicot, S.; Ferret-Bernard, S.; Guérin, S.; Romé, V.; Le Normand, L.; Randuineau, G.; Gambarota, G.; Noirot, V.; Etienne, P.; et al. Validation of a Psychosocial Chronic Stress Model in the Pig Using a Multidisciplinary Approach at the Gut-Brain and Behavior Levels. Front. Behav. Neurosci. 2019, 13, 161. [Google Scholar] [CrossRef] [PubMed]
- Prims, S.; Vanden Hole, C.; Van Cruchten, S.; Van Ginneken, C.; Van Ostade, X.; Casteleyn, C. Hair or salivary cortisol analysis to identify chronic stress in piglets? Vet. J. 2019, 252, 105357. [Google Scholar] [CrossRef] [PubMed]
- Noushad, S.; Ahmed, S.; Ansari, B.; Mustafa, U.H.; Saleem, Y.; Hazrat, H. Physiological biomarkers of chronic stress: A systematic review. Int. J. Health Sci. 2021, 15, 46–59. [Google Scholar]
- Milligan Armstrong, A.; Porter, T. Chronic stress and Alzheimer’s disease: The interplay between the hypothalamic-pituitary-adrenal axis, genetics and microglia. Biol. Rev. 2021, 96, 2209–2228. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascon, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, Q.; Peng, J.; Zhao, Y.; Wei, M.; Fan, H. Lycopene alleviates chronic stress-induced hippocampal microglia ferroptosis by inhibiting the ASK1/JNK signaling pathway. Food Biosci. 2025, 64, 105859. [Google Scholar] [CrossRef]
- Hambright, W.S.; Fonseca, R.S.; Chen, L.; Na, R.; Ran, Q. Ablation of ferroptosis regulator glutathione peroxidase 4 in forebrain neurons promotes cognitive impairment and neurodegeneration. Redox Biol. 2017, 12, 8–17. [Google Scholar] [CrossRef]
- Masaldan, S.; Bush, A.I.; Devos, D.; Rolland, A.S.; Moreau, C. Striking while the iron is hot: Iron metabolism and ferroptosis in neurodegeneration. Free Radic. Biol. Med. 2019, 133, 221–233. [Google Scholar] [CrossRef]
- Mehrpouya, S.; Nahavandi, A.; Khojasteh, F.; Soleimani, M.; Ahmadi, M.; Barati, M. Iron administration prevents BDNF decrease and depressive-like behavior following chronic stress. Brain Res. 2015, 1596, 79–87. [Google Scholar] [CrossRef]
- Zhang, H.; Jiao, W.; Cui, H.; Sun, Q.; Fan, H. Combined exposure of alumina nanoparticles and chronic stress exacerbates hippocampal neuronal ferroptosis via activating IFN-γ/ASK1/JNK signaling pathway in rats. J. Hazard. Mater. 2021, 411, 125179. [Google Scholar] [CrossRef] [PubMed]
- Farajdokht, F.; Soleimani, M.; Mehrpouya, S.; Barati, M.; Nahavandi, A. The role of hepcidin in chronic mild stress-induced depression. Neurosci. Lett. 2015, 588, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Joffe, M.E.; Santiago, C.I.; Oliver, K.H.; Maksymetz, J.; Harris, N.A.; Engers, J.L.; Lindsley, C.W.; Winder, D.G.; Conn, P.J. mGlu and mGlu Negative Allosteric Modulators Divergently Enhance Thalamocortical Transmission and Exert Rapid Antidepressant-like Effects. Neuron 2020, 105, 46–59. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, H.; Wang, H.; Li, Q.; Zhang, T. Neuroprotective effect of amantadine on corticosterone-induced abnormal glutamatergic synaptic transmission of CA3-CA1 pathway in rat’s hippocampal slices. Synapse 2017, 71, 12. [Google Scholar] [CrossRef]
- Wei, S.; Xu, H.; Xia, D.; Zhao, R. Curcumin attenuates the effects of transport stress on serum cortisol concentration, hippocampal NO production, and BDNF expression in the pig. Domest. Anim. Endocrinol. 2010, 39, 231–239. [Google Scholar] [CrossRef]
- Chen, Q.; Qiu, F.S.; Xie, W.; Yu, W.Y.; Su, Z.A.; Qin, G.M.; Kang, Y.K.; Jiang, S.L.; Yu, C.H. Gypenoside A-loaded mPEG-PLGA nanoparticles ameliorate high-glucose-induced retinal microvasculopathy by inhibiting ferroptosis. Int. J. Pharm. 2024, 666, 124758. [Google Scholar] [CrossRef]
- Xie, P.; Luo, H.T.; Pei, W.J.; Xiao, M.Y.; Li, F.F.; Gu, Y.L.; Piao, X.L. Saponins derived from Gynostemma pentaphyllum regulate triglyceride and cholesterol metabolism and the mechanisms: A review. J. Ethnopharmacol. 2024, 319, 117186. [Google Scholar] [CrossRef]
- Dong, S.Q.; Zhang, Q.P.; Zhu, J.X.; Chen, M.; Li, C.F.; Liu, Q.; Geng, D.; Yi, L.T. Gypenosides reverses depressive behavior via inhibiting hippocampal neuroinflammation. Biomed. Pharmacother. 2018, 106, 1153–1160. [Google Scholar] [CrossRef]
- Zhang, G.L.; Deng, J.P.; Wang, B.H.; Zhao, Z.W.; Li, J.; Gao, L.; Liu, B.L.; Xong, J.R.; Guo, X.D.; Yan, Z.Q.; et al. Gypenosides improve cognitive impairment induced by chronic cerebral hypoperfusion in rats by suppressing oxidative stress and astrocytic activation. Behav. Pharmacol. 2011, 22, 633–644. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, M.; Sun, N.; Wang, H.; Fan, H. Melatonin attenuates chronic stress-induced hippocampal inflammatory response and apoptosis by inhibiting ADAM17/TNF-alpha axis. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2022, 169, 113441. [Google Scholar] [CrossRef]
- Xiao, J.; Yao, R.; Xu, B.; Wen, H.; Zhong, J.; Li, D.; Zhou, Z.; Xu, J.; Wang, H. Inhibition of PDE4 Attenuates TNF-alpha-Triggered Cell Death Through Suppressing NF-kappaB and JNK Activation in HT-22 Neuronal Cells. Cell. Mol. Neurobiol. 2020, 40, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, J.; Mukherjee, A.; He, T.; Wang, X.Y.; Ma, Y.; Fang, X. Lysophosphatidic acid induces tumor necrosis factor-alpha to regulate a pro-inflammatory cytokine network in ovarian cancer. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 13935–13948. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, P.; Aguirre, P.; Esparza, A.; Tapia, V.; Mena, N.P.; Arredondo, M.; Gonzalez-Billault, C.; Nunez, M.T. Inflammation alters the expression of DMT1, FPN1 and hepcidin, and it causes iron accumulation in central nervous system cells. J. Neurochem. 2013, 126, 541–549. [Google Scholar] [CrossRef]
- Wang, J.; Deng, B.; Liu, Q.; Huang, Y.; Chen, W.; Li, J.; Zhou, Z.; Zhang, L.; Liang, B.; He, J.; et al. Pyroptosis and ferroptosis induced by mixed lineage kinase 3 (MLK3) signaling in cardiomyocytes are essential for myocardial fibrosis in response to pressure overload. Cell Death Dis. 2020, 11, 574. [Google Scholar] [CrossRef]
- Jiang, L.; Kon, N.; Li, T.; Wang, S.-J.; Su, T.; Hibshoosh, H.; Baer, R.; Gu, W. Ferroptosis as a p53-mediated activity during tumour suppression. Nature 2015, 520, 57–62. [Google Scholar] [CrossRef]
- Wang, L.; Wang, H.; Li, L.; Li, W.; Dong, X.; Li, M.; Lv, L. Corticosterone induces dysregulation of iron metabolism in hippocampal neurons in vitro. Biol. Trace Elem. Res. 2010, 137, 88–95. [Google Scholar] [CrossRef]
- Zhang, Z.; Bai, H.; Ma, X.; Shen, M.; Li, R.; Qiu, D.; Li, S.; Gao, L. Blockade of the NLRP3/caspase-1 axis attenuates ketamine-induced hippocampus pyroptosis and cognitive impairment in neonatal rats. J. Neuroinflammation 2021, 18, 239. [Google Scholar] [CrossRef]
- Duan, L.; Zhang, Y.; Yang, Y.; Su, S.; Zhou, L.; Lo, P.C.; Cai, J.; Qiao, Y.; Li, M.; Huang, S.; et al. Baicalin Inhibits Ferroptosis in Intracerebral Hemorrhage. Front. Pharmacol. 2021, 12, 629379. [Google Scholar] [CrossRef]
- Bai, L.; Yan, F.; Deng, R.; Gu, R.; Zhang, X.; Bai, J. Thioredoxin-1 Rescues MPP(+)/MPTP-Induced Ferroptosis by Increasing Glutathione Peroxidase 4. Mol. Neurobiol. 2021, 58, 3187–3197. [Google Scholar] [CrossRef]
- Roth, J.A.; Feng, L.; Dolan, K.G.; Lis, A.; Garrick, M.D. Effect of the iron chelator desferrioxamine on manganese-induced toxicity of rat pheochromocytoma (PC12) cells. J. Neurosci. Res. 2002, 68, 76–83. [Google Scholar] [CrossRef]
- Zhu, Q.; Gao, Z.; Peng, J.; Liu, C.; Wang, X.; Li, S.; Zhang, H. Lycopene Alleviates Chronic Stress-Induced Hippocampal Microglial Pyroptosis by Inhibiting the Cathepsin B/NLRP3 Signaling Pathway. J. Agric. Food Chem. 2023, 71, 20034–20046. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Liu, J.; Zhu, Q.; Zhao, L.; Feng, Y.; Wang, X.; Cao, W.; Xin, H. Gypenosides protect primary cultures of rat cortical cells against oxidative neurotoxicity. Brain Res. 2006, 1102, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Wang, B.-Y.; Yang, Y.-F.; Kuai, L.-Y.; Wan, J.-j.; Zhang, M.; Xia, H.-Y.; Wang, Y.; Zheng, Z.; Meng, X.-W.; et al. Ciprofol Ameliorates Myocardial Ischemia/Reperfusion Injury by Inhibiting Ferroptosis Through Upregulating HIF-1α. Drug Des. Dev. Ther. 2024, 18, 6115–6132. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Khan, S.; Nabi, G.; Gamallat, Y.; Su, P.; Jamalat, Y.; Duan, P.; Yao, L. Natural gypenosides: Targeting cancer through different molecular pathways. Cancer Manag. Res. 2019, 11, 2287–2297. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Number | Upstream and Downstream Primer Sequence | Product Length (bp) |
|---|---|---|---|
| Hepcidin | NM_053469.2 | F: CTATCTCCGGCAACAGACGA R: TGTCTCGCTTCCTTCGCTTC | 110 |
| DMT1 | NM_013173.2 | F: TGGTTAGCGTGGCTTATCTGG R: AGTATTGCCACCGCTGGTATC | 143 |
| FPN1 | NM_133315.2 | F: TGGGAGCATCAGCAATAAC R: CAGACCAGTCCGAACAAGG | 86 |
| Ferritin | NM_022500.5 | F: GGAACTTCACAAACTGGCTAC R: TGGATTTCACCTGCTCATT | 89 |
| GLS2 | NM_001270786.1 | F: GGGTGTCCGGTACTACTTCG R: GTTCGAGGCATCATGGTCCG | 94 |
| GPX4 | NM_001039849.3 | F: GACCTTCCCCAGACCAGCAAC R: CGCAACCCCTGTACTTATCCAG | 145 |
| SLC7A11 | NM_001107673.3 | F: TCAAATCCTTGGCCATCTGC R: ACCAATTCCTTTAGCCCATCATC | 92 |
| IL-6 | NM_012589.2 | F: CTTCTTGGGACTGATGTTG R: TACTGGTCTGTTGTGGGTG | 97 |
| IL-1β | NM_031512.2 | F: CTCGTGGGATGATGACGACC R: AGCTTTCAGCTCACATGGGT | 118 |
| TNF-α | NM_012675.3 | F: GCCACCACGCTCTTCTGTC R: GCTACGGGCTTGTCACTCG | 149 |
| TNFR1 | NM_013091.2 | F: CCAAGTGCCACAAAGGAACC R: GTGCCTTTATCACACACCTCG | 85 |
| NF-κB | NM_199267.2 | F: ACTGCCGGGATGGCTTCTAT R: CTTGCTCCAGGTCTCGCTTC | 105 |
| p53 | NM_030989.3 | F: AGCGACTACAGTTAGGGGGT R: ACAGTTATCCAGTCTTCAGGGG | 89 |
| GAPDH | NM_017008.4 | F: GGCAAGTTCAACGGCACAG R: CGCCAGTAGACTCCACGAC | 142 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, L.; Peng, J.; Zhang, M.; Hu, Y.; Gao, Z.; Wang, J.; Zhang, H.; Li, S. Gypenosides Attenuates CORT-Induced Ferroptosis via Inhibiting TNF-α/NF-κB Signaling Pathway in PC12 Cells. Molecules 2025, 30, 2103. https://doi.org/10.3390/molecules30102103
Dai L, Peng J, Zhang M, Hu Y, Gao Z, Wang J, Zhang H, Li S. Gypenosides Attenuates CORT-Induced Ferroptosis via Inhibiting TNF-α/NF-κB Signaling Pathway in PC12 Cells. Molecules. 2025; 30(10):2103. https://doi.org/10.3390/molecules30102103
Chicago/Turabian StyleDai, Lingling, Jinghui Peng, Manyu Zhang, Yulin Hu, Zhicheng Gao, Jibin Wang, Haiyang Zhang, and Shoujun Li. 2025. "Gypenosides Attenuates CORT-Induced Ferroptosis via Inhibiting TNF-α/NF-κB Signaling Pathway in PC12 Cells" Molecules 30, no. 10: 2103. https://doi.org/10.3390/molecules30102103
APA StyleDai, L., Peng, J., Zhang, M., Hu, Y., Gao, Z., Wang, J., Zhang, H., & Li, S. (2025). Gypenosides Attenuates CORT-Induced Ferroptosis via Inhibiting TNF-α/NF-κB Signaling Pathway in PC12 Cells. Molecules, 30(10), 2103. https://doi.org/10.3390/molecules30102103