
Artemisinin Confers Cytoprotection toward Hydrogen Peroxide-Induced Cell Apoptosis in Retinal Pigment Epithelial Cells in Correlation with the Increased Acetylation of Histone H4 at Lysine 8



Abstract
1. Introduction
2. Results
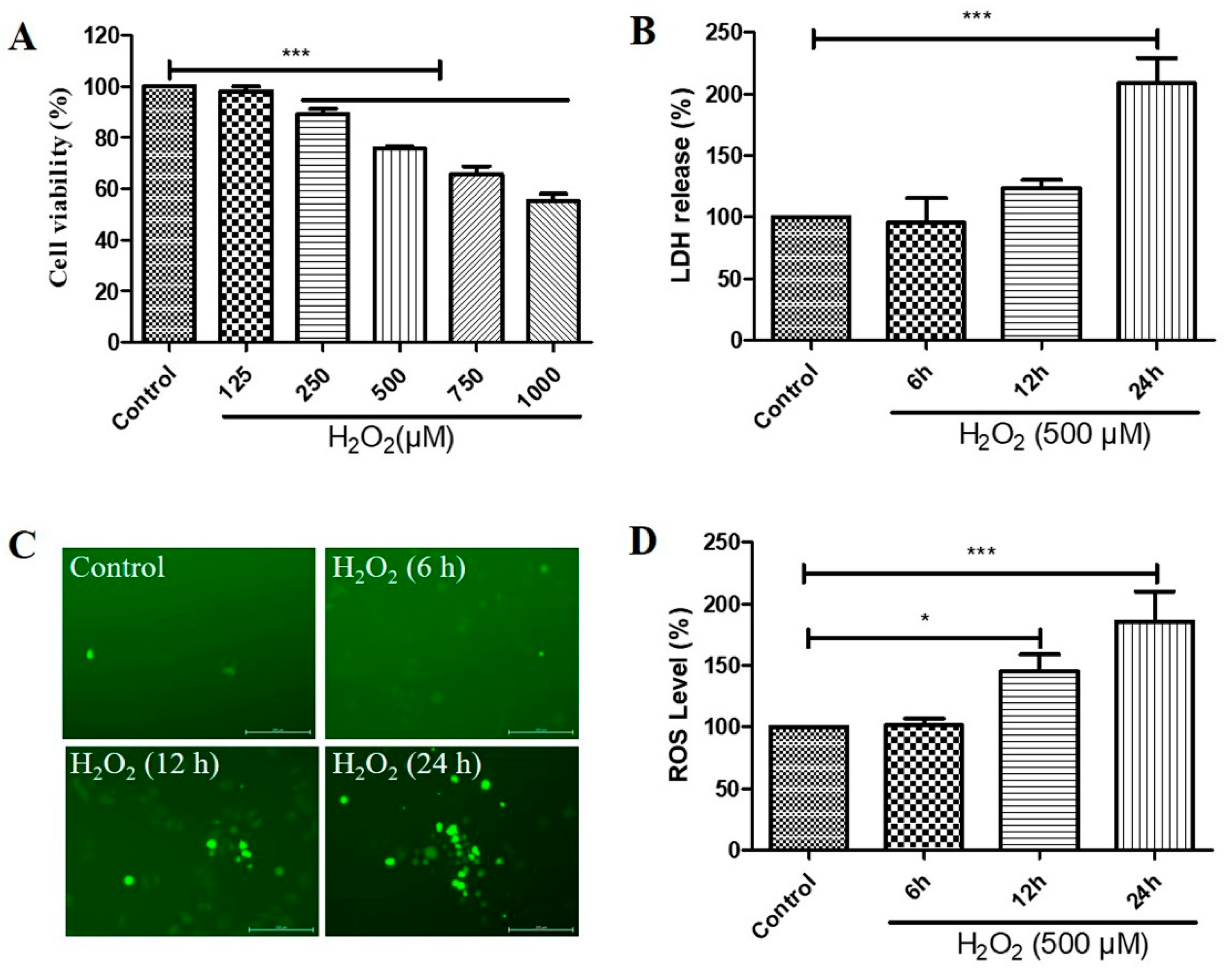
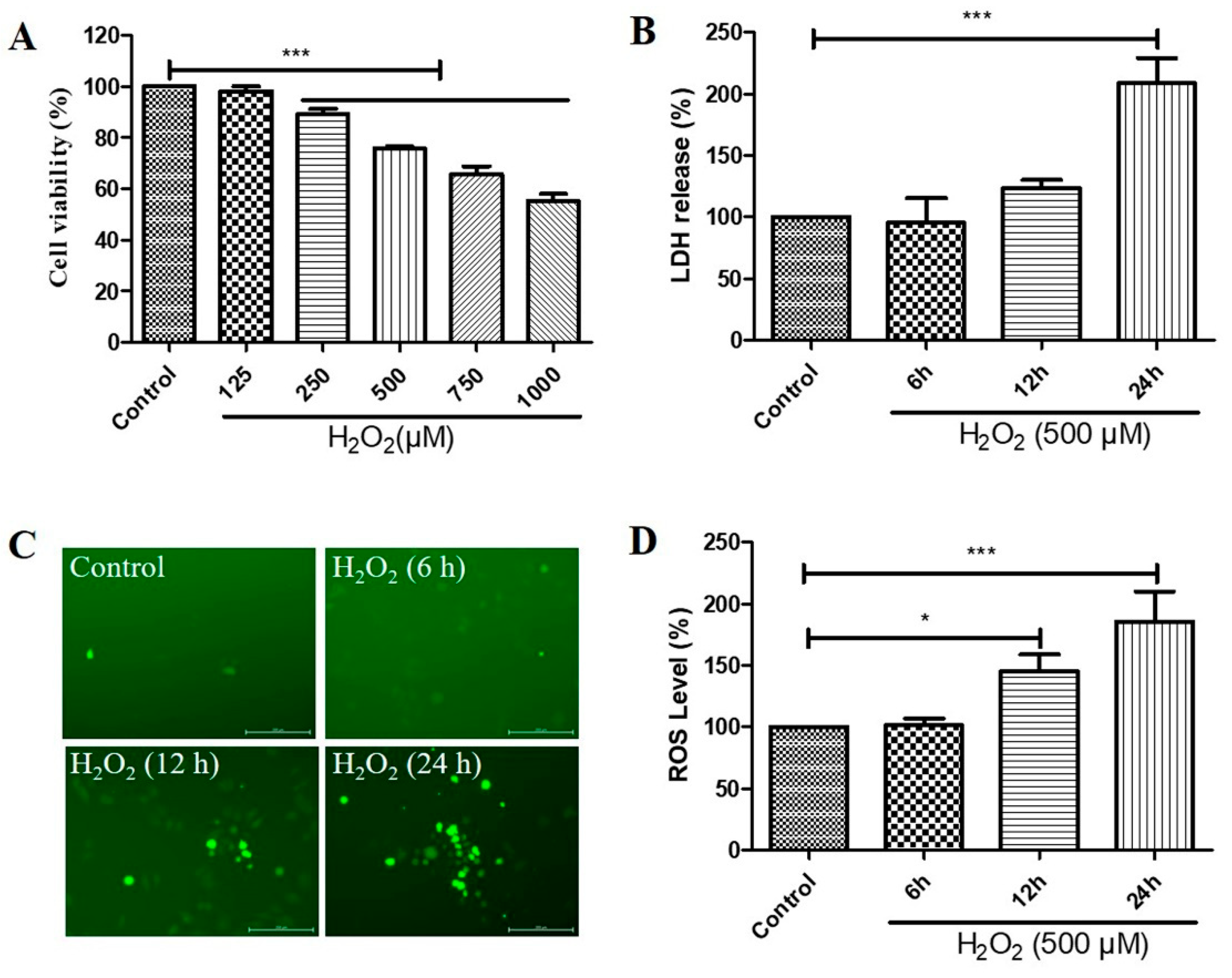
2.1. H2O2 Decreased Cell Viability, Increased Cytotoxicity, and Caused Reactive Oxygen Species (ROS) Generation in D407 Cell Cultures
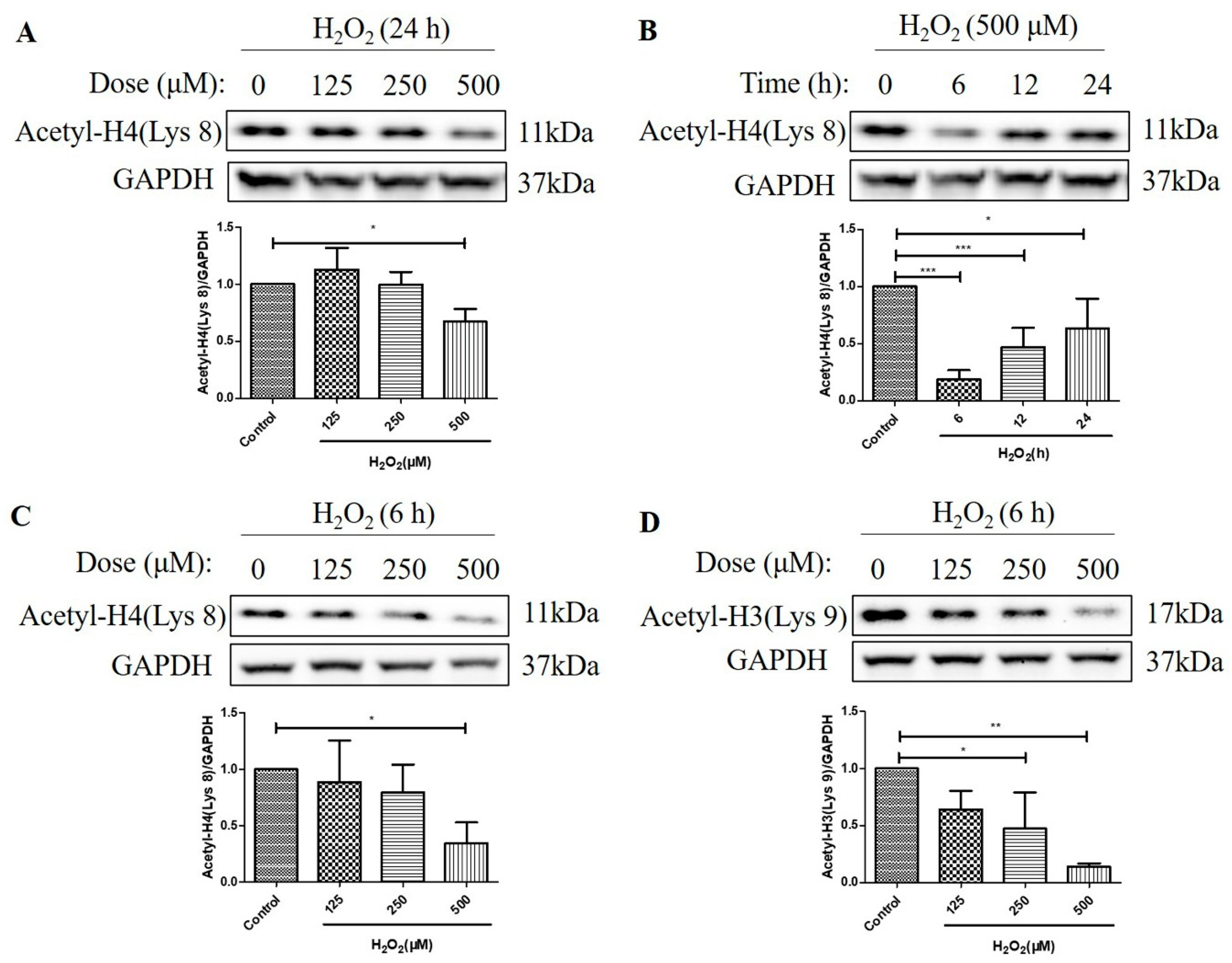
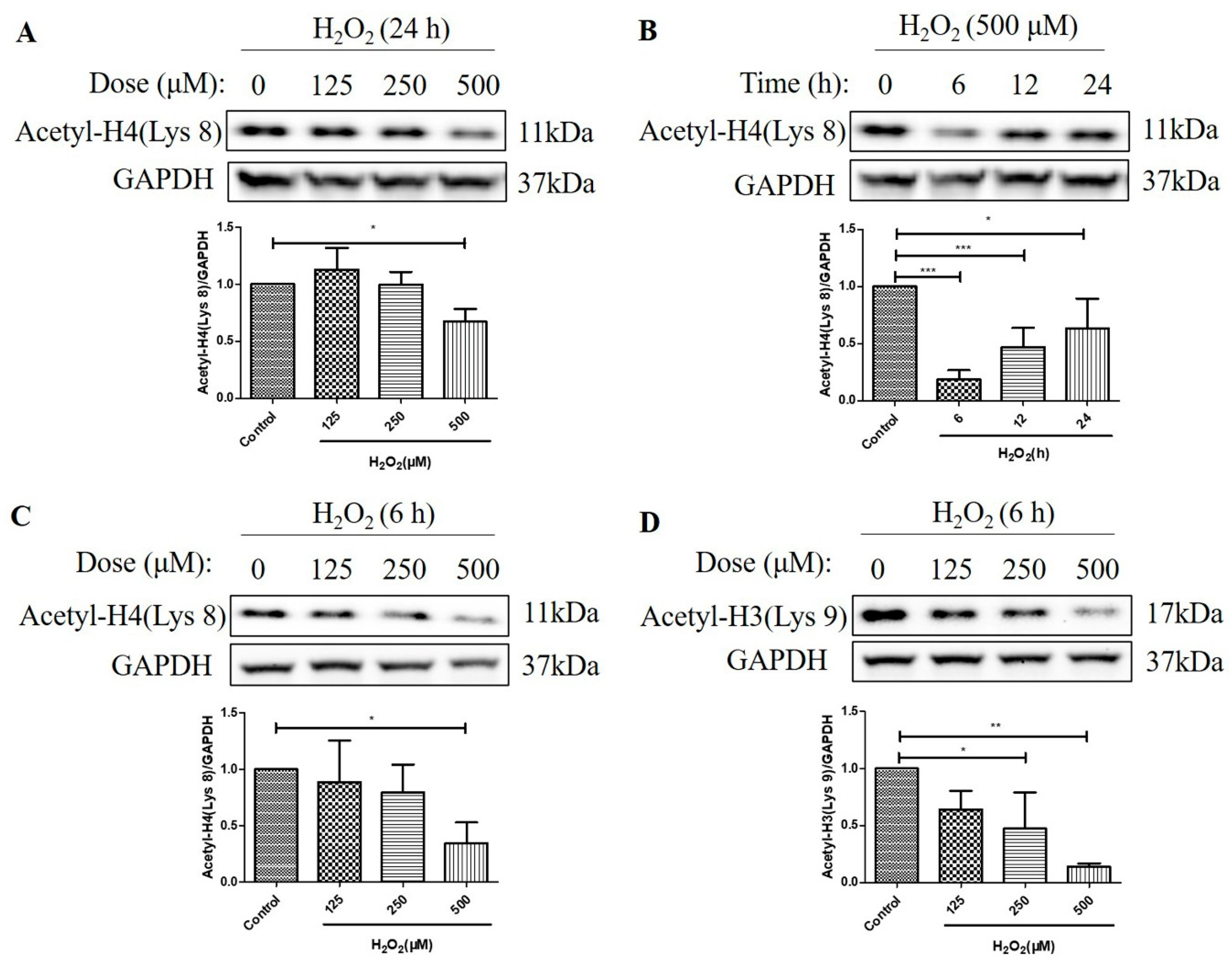
2.2. H2O2 Decreased Histone H4 (Lys 8) and Histone H3 (Lys 9) Acetylation Levels in D407 Cell Cultures
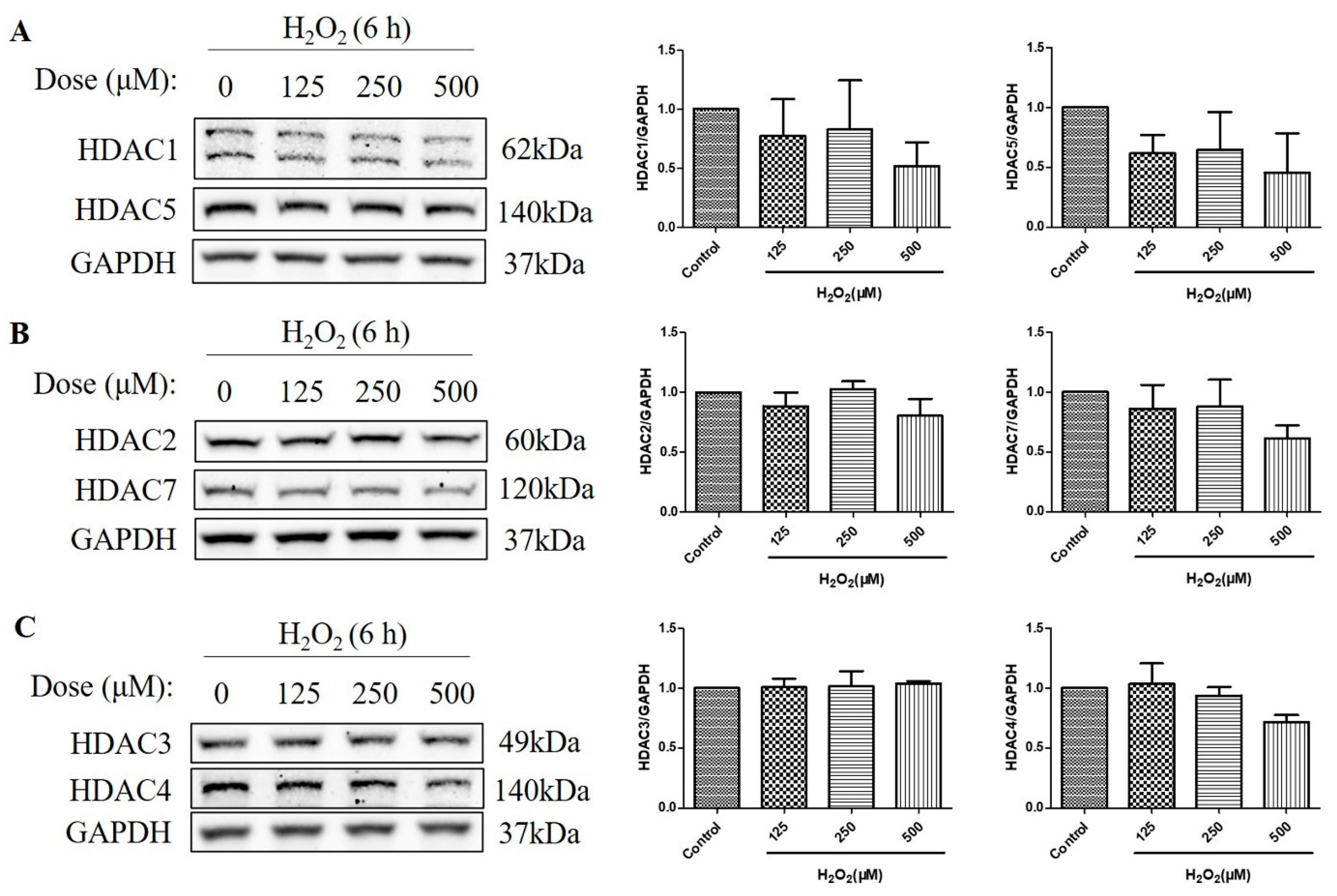
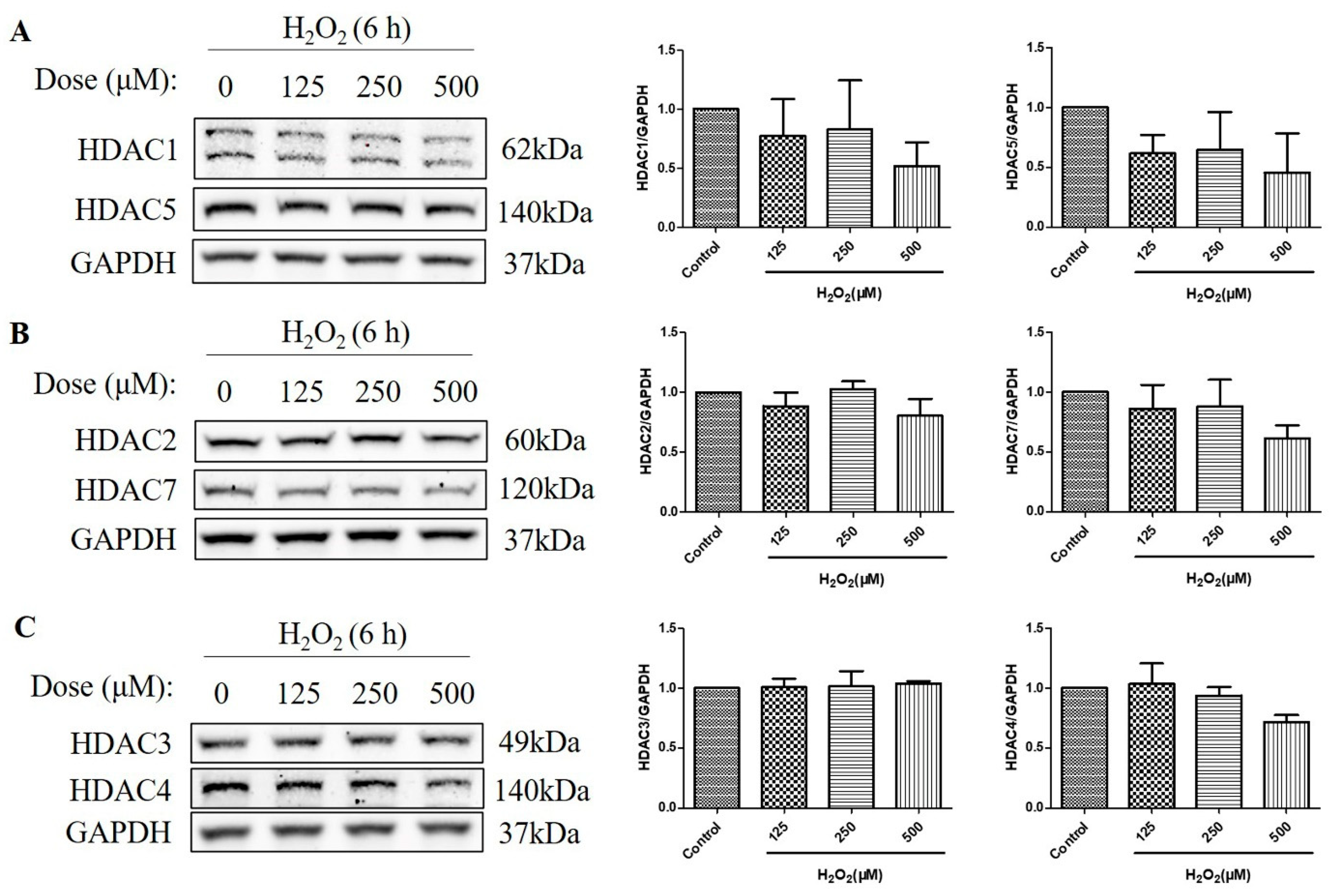
2.3. HDAC Protein Levels Were Not Upregulated after the Treatment with H2O2 for 6 h in D407 Cell Cultures
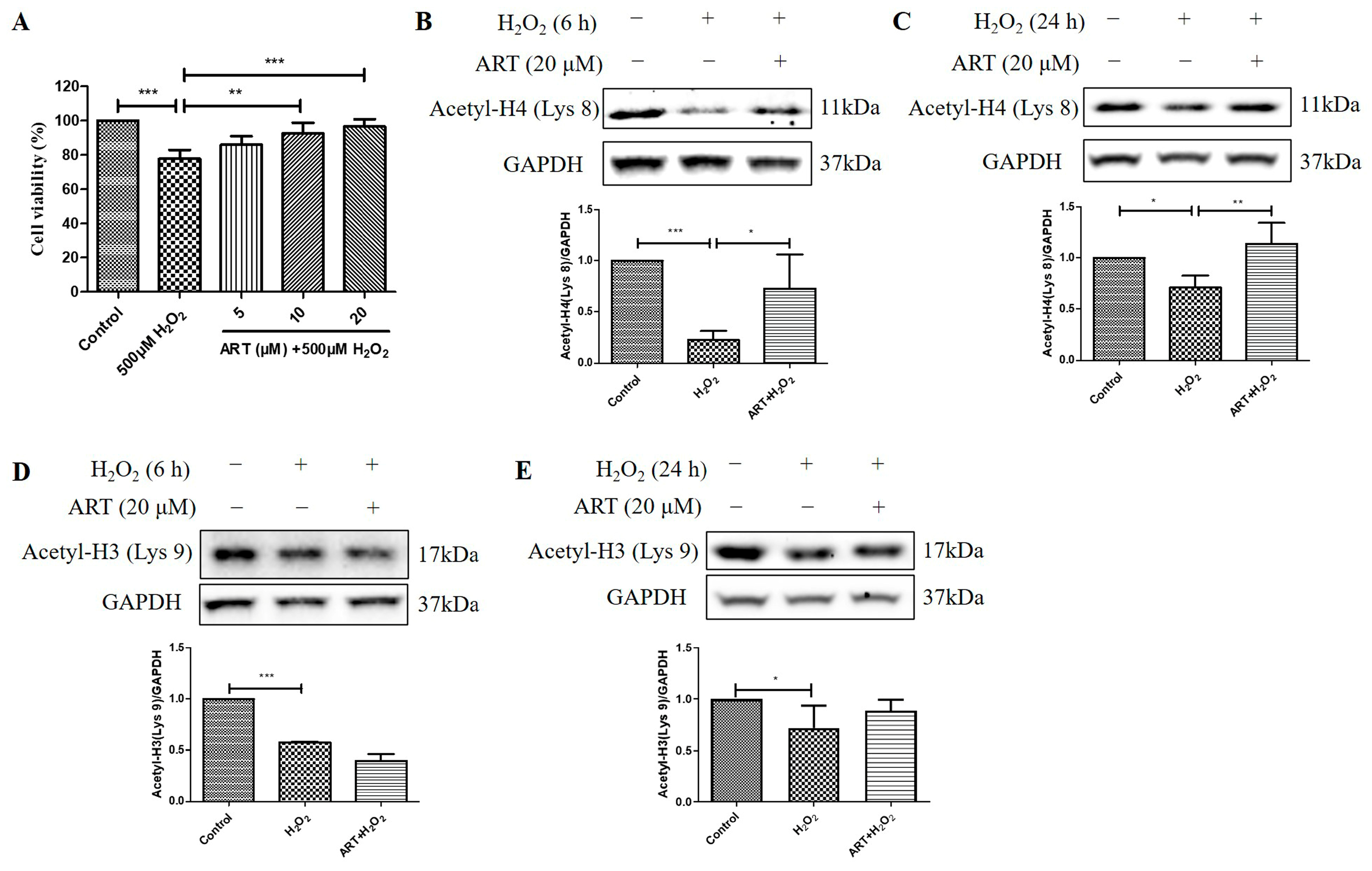
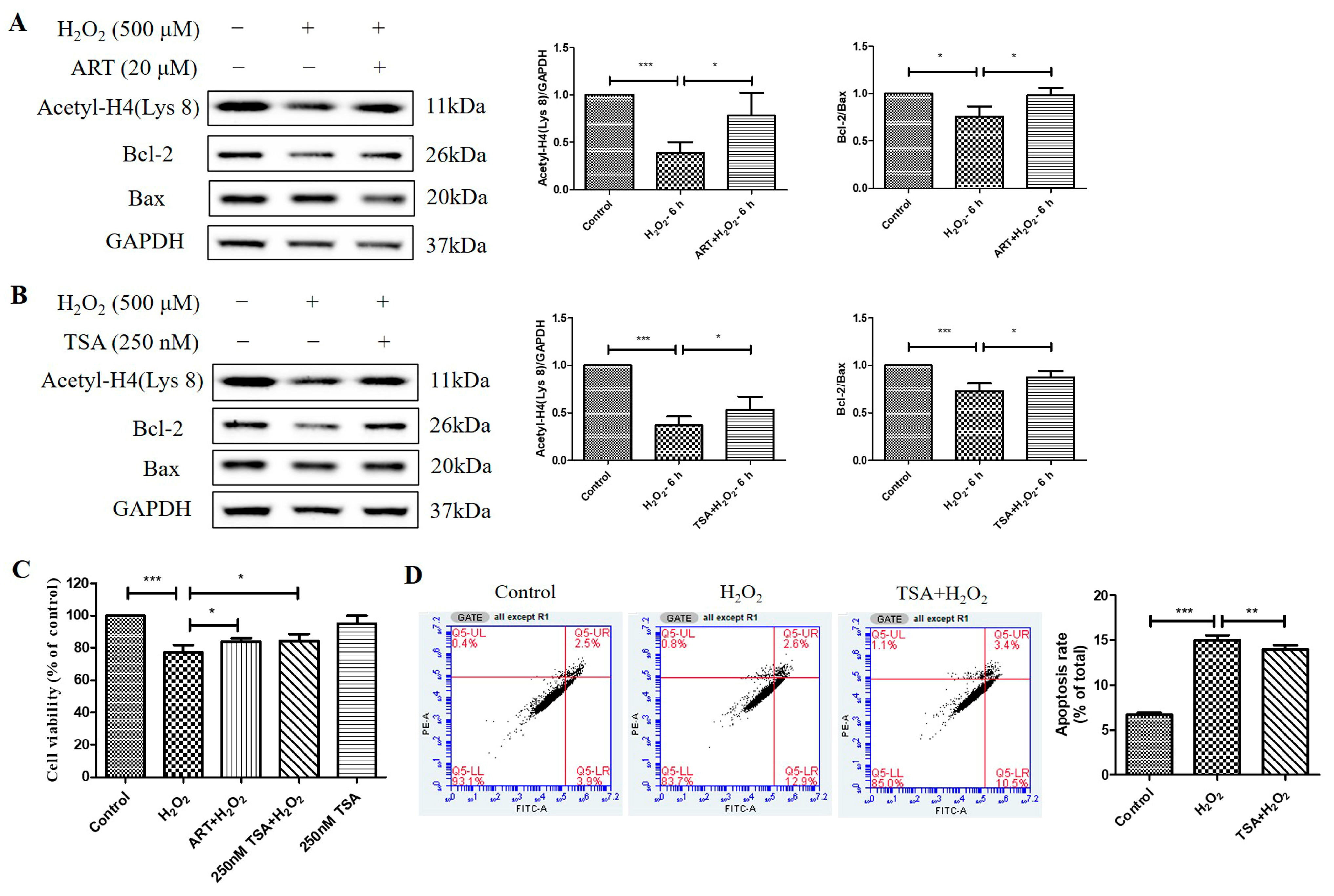
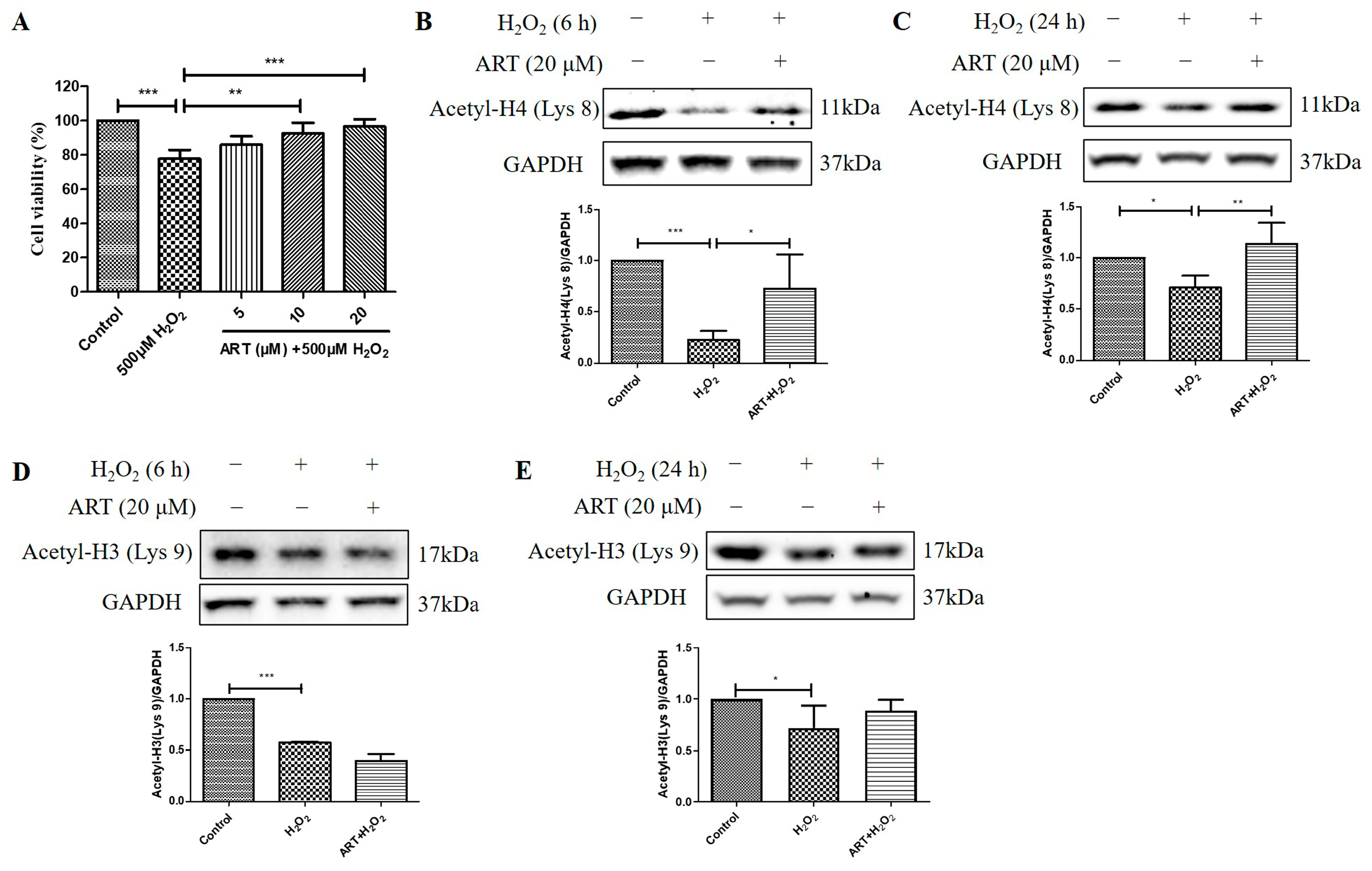
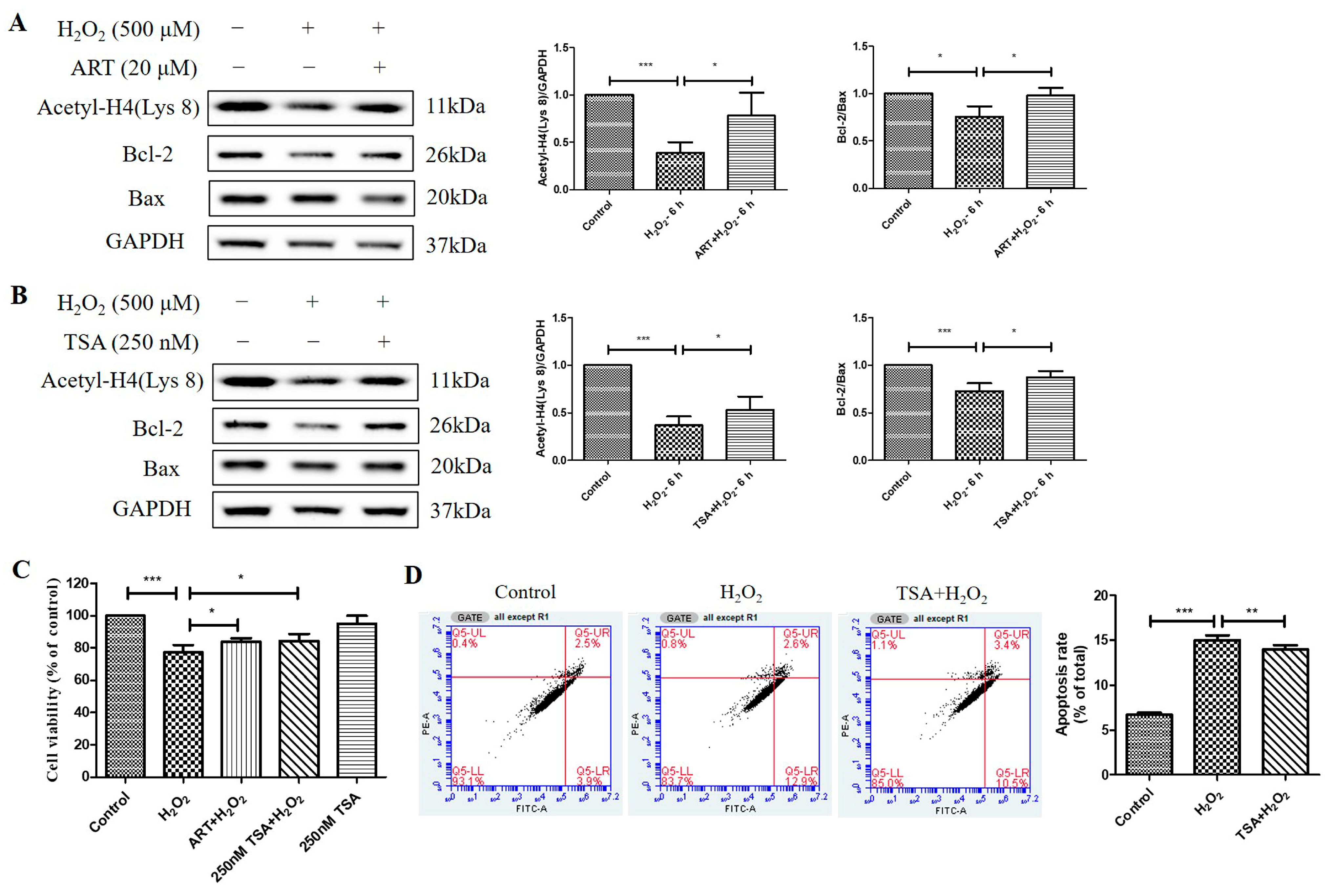
2.4. Artemisinin Corrected H2O2-Induced Cell Viability Decline, and Decrease in Histone H4 (Lys 8) Acetylation Level in D407 Cell Cultures
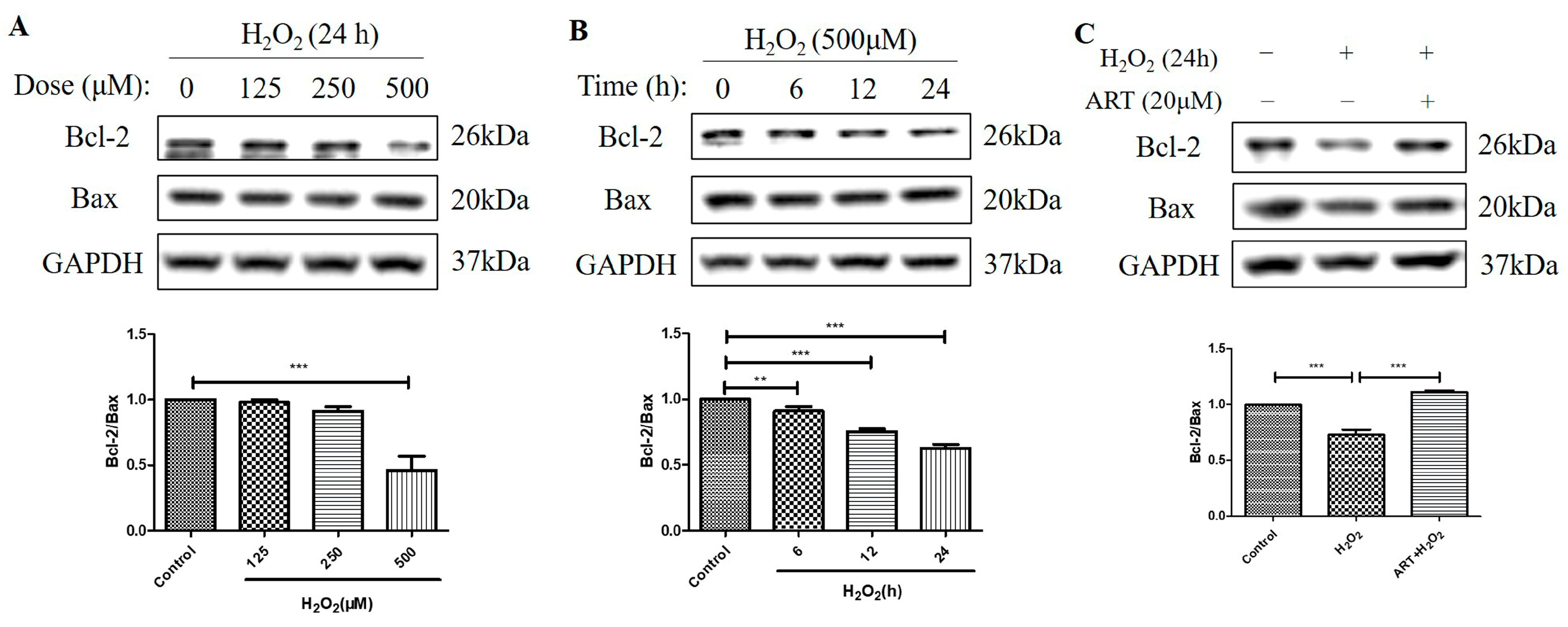
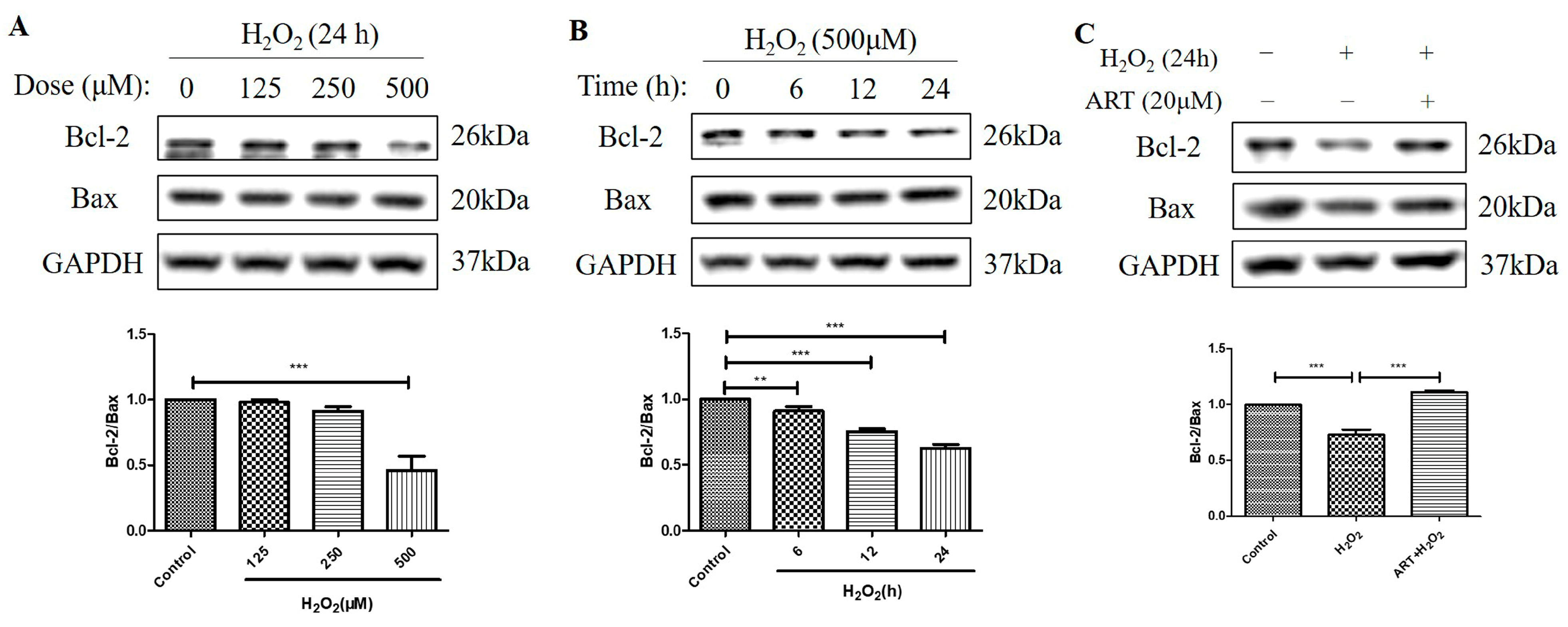
2.5. Artemisinin Reversed H2O2-Induced Downregulation of B-Cell Lymphoma 2 (Bcl-2) to Bcl-2 Associated X-Protein (Bax) Protein Ratio in D407 Cell Cultures
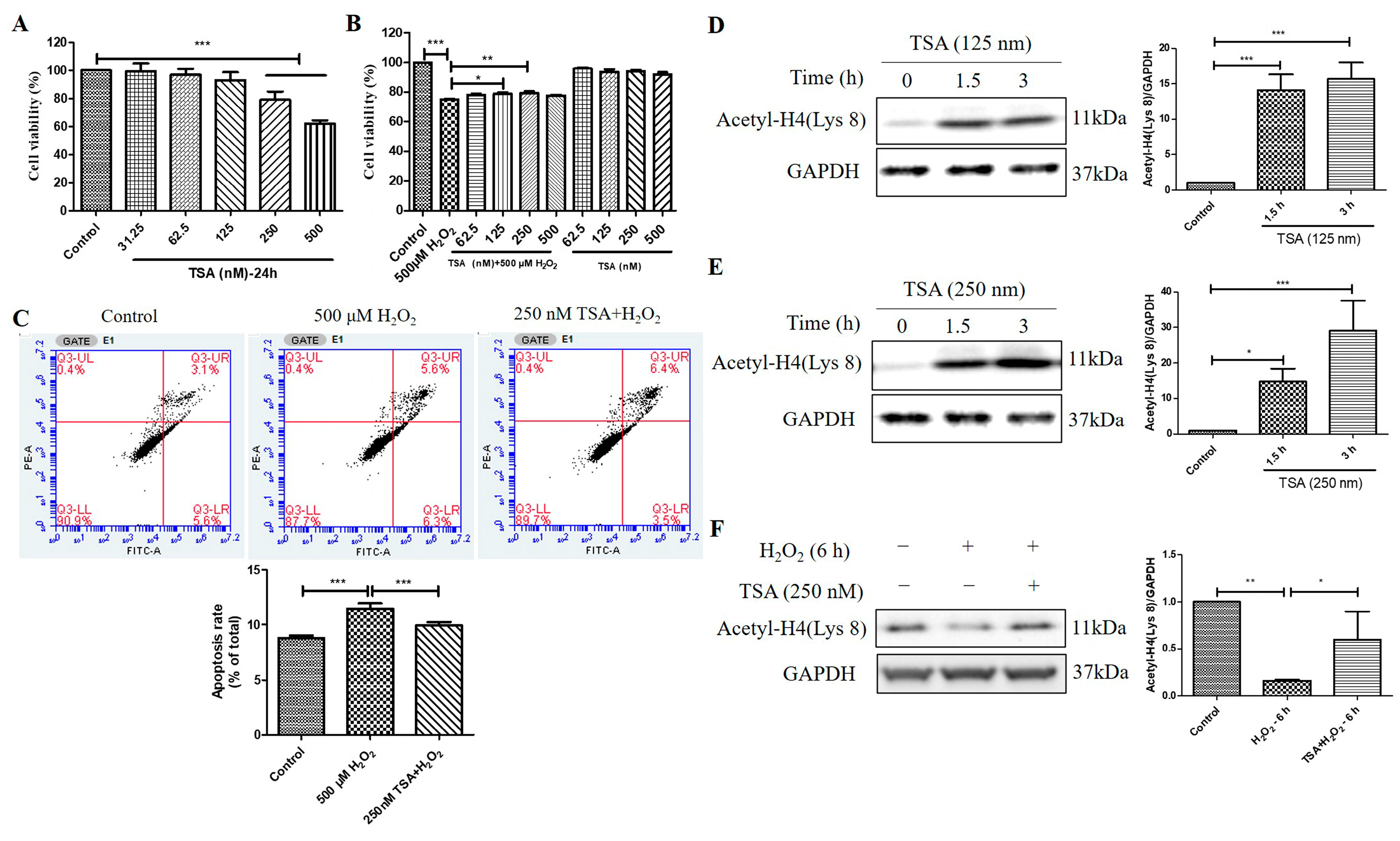
2.6. Trichostatin A (TSA) Reversed H2O2-Induced Cell Viability Decline, Apoptosis, and Decrease in Histone H4 (Lys 8) Acetylation Levels in D407 Cell Cultures
2.7. The Cytoprotective Effect of Artemisinin toward H2O2-Induced Apoptosis in ARPE19 Cell Cultures Is Correlated to Reversal of the Histone H4 (Lys 8) Acetylation Level Decline
3. Discussion
4. Materials and Methods
4.1. Drugs, Regents, and Kits
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. LDH Release Assay
4.5. Flow Cytometry
4.6. ROS Assay
4.7. Western Blotting
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Al Gwairi, O.; Thach, L.; Zheng, W.; Osman, N.; Little, P.J. Cellular and molecular pathology of age-related macular degeneration: Potential role for proteoglycans. J. Ophthalmol. 2016, 2016, 291361. [Google Scholar] [CrossRef] [PubMed]
- Somasundaran, S.; Constable, I.J.; Mellough, C.B.; Carvalho, L.S. Retinal pigment epithelium and age-related macular degeneration: A review of major disease mechanisms. Clin. Exp. Ophthalmol. 2020, 48, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Kushwah, N.; Bora, K.; Maurya, M.; Pavlovich, M.C.; Chen, J. Oxidative Stress and Antioxidants in Age-Related Macular Degeneration. Antioxidants 2023, 12, 1379. [Google Scholar] [CrossRef] [PubMed]
- Maurya, M.; Bora, K.; Blomfield, A.K.; Pavlovich, M.C.; Huang, S.; Liu, C.H.; Chen, J. Oxidative stress in retinal pigment epithelium degeneration: From pathogenesis to therapeutic targets in dry age-related macular degeneration. Neural Regen. Res. 2023, 18, 2173–2181. [Google Scholar]
- Corso-Díaz, X.; Jaeger, C.; Chaitankar, V.; Swaroop, A. Epigenetic control of gene regulation during development and disease: A view from the retina. Progress Retin. Eye Res. 2018, 65, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Nashine, S.; Nesburn, A.B.; Kuppermann, B.D.; Kenney, M.C. Age-related macular degeneration (AMD) mitochondria modulate epigenetic mechanisms in retinal pigment epithelial cells. Exp. Eye Res. 2019, 189, 107701. [Google Scholar] [CrossRef]
- Campos, E.I.; Reinberg, D. Histones: Annotating Chromatin. Annu. Rev. Genet. 2009, 43, 559–599. [Google Scholar] [CrossRef]
- Bowman, G.D.; Poirier, M.G. Post-Translational Modifications of Histones That Influence Nucleosome Dynamics. Chem. Rev. 2015, 115, 2274–2295. [Google Scholar] [CrossRef]
- Chen, H.P.; Zhao, Y.T.; Zhao, T.C. Histone deacetylases and mechanisms of regulation of gene expression. Crit. Rev. Oncog. 2015, 20, 35–47. [Google Scholar] [CrossRef]
- Eberhart, A.; Feodorova, Y.; Song, C.; Wanner, G.; Kiseleva, E.; Furukawa, T.; Kimura, H.; Schotta, G.; Leonhardt, H.; Joffe, B.; et al. Epigenetics of eu- and heterochromatin in inverted and conventional nuclei from mouse retina. Chromosome Res. 2013, 21, 535–554. [Google Scholar] [CrossRef]
- Kizilyaprak, C.; Spehner, D.; Devys, D.; Schultz, P. In Vivo Chromatin Organization of Mouse Rod Photoreceptors Correlates with Histone Modifications. PLoS ONE 2010, 5, e11039. [Google Scholar] [CrossRef]
- Pelzel, H.R.; Schlamp, C.L.; Nickells, R.W. Histone H4 deacetylation plays a critical role in early gene silencing during neuronal apoptosis. BMC Neurosci. 2010, 11, 62. [Google Scholar] [CrossRef]
- Wang, D.; Chen, Y.; Li, J.; Wu, E.; Tang, T.; Singla, R.K.; Shen, B.; Zhang, M. Natural Products for the Treatment of Age-Related Macular Degeneration. Phytomedicine, 2024, in press.
- Chong, C.-M.; Zheng, W. Artemisinin protects human retinal pigment epithelial cells from hydrogen peroxide-induced oxidative damage through activation of ERK/CREB signaling. Redox Biol. 2016, 9, 50–56. [Google Scholar] [CrossRef]
- Li, S.; Zhao, X.; Gaur, U.; Fang, J.; Yan, F.; Zheng, W. Artemisinin Protects Human Retinal Pigmented Epithelial Cells against Hydrogen Peroxide-induced Oxidative Damage by Enhancing the Activation of AMP-active Protein Kinase. Int. J. Biol. Sci. 2019, 15, 2016–2028. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Wang, H.; Gao, Y.; Xu, J.; Zheng, W. Artemisinin protects retinal neuronal cells against oxidative stress and restores rat retinal physiological function from light exposed damage. ACS Chem. Neurosci. 2017, 8, 1713–1723. [Google Scholar] [CrossRef]
- Xu, G.; Huang, Y.L.; Li, P.L.; Guo, H.M.; Han, X.P. Neuroprotective effects of artemisinin against isoflurane-induced cognitive impairments and neuronal cell death involve JNK/ERK1/2 signalling and improved hippocampal histone acetylation in neonatal rats. J. Pharm. Pharmacol. 2017, 69, 684–697. [Google Scholar] [CrossRef]
- Ransy, C.; Vaz, C.; Lombès, A.; Bouillaud, F. Use of H2O2 to Cause Oxidative Stress, the Catalase Issue. Int. J. Mol. Sci. 2020, 21, 9149. [Google Scholar] [CrossRef] [PubMed]
- Dokmanovic, M.; Clarke, C.; Marks, P.A. Histone deacetylase inhibitors: Overview and perspectives. Mol. Cancer Res. 2007, 5, 981–989. [Google Scholar] [CrossRef]
- Seto, E.; Yoshida, M. Erasers of Histone Acetylation: The Histone Deacetylase Enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6, a018713. [Google Scholar] [CrossRef]
- Samardzija, M.; Corna, A.; Gomez-Sintes, R.; Jarboui, M.A.; Armento, A.; Roger, J.E.; Petridou, E.; Haq, W.; Paquet-Durand, F.; Zrenner, E.; et al. HDAC inhibition ameliorates cone survival in retinitis pigmentosa mice. Cell Death Differ. 2021, 28, 1317–1332. [Google Scholar] [CrossRef]
- Sancho-Pelluz, J.; Alavi, M.V.; Sahaboglu, A.; Kustermann, S.; Farinelli, P.; Azadi, S.; van Veen, T.; Romero, F.J.; Paquet-Durand, F.; Ekström, P. Excessive HDAC activation is critical for neurodegeneration in the rd1 mouse. Cell Death Dis. 2010, 1, e24. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Wei, Z.; Yang, W.; Huang, J.; Yang, Y.; Wang, J. The role of BCL-2 family proteins in regulating apoptosis and cancer therapy. Front. Oncol. 2022, 12, 985363. [Google Scholar] [CrossRef] [PubMed]
- Ayna, A. Caffeic acid prevents hydrogen peroxide-induced oxidative damage in SH-SY5Y cell line through mitigation of oxidative stress and apoptosis. Bratisl. Med. J. 2021, 122, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, J.; Liu, M.; Chen, G. Fluvastatin protects neuronal cells from hydrogen peroxide-induced toxicity with decreasing oxidative damage and increasing PI3K/Akt/mTOR signalling. J. Pharm. Pharmacol. 2021, 73, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Fang, J.; Li, S.; Gaur, U.; Xing, X.; Wang, H.; Zheng, W. Artemisinin attenuated hydrogen peroxide (H2O2)-induced oxidative injury in SH-SY5Y and hippocampal neurons via the activation of AMPK pathway. Int. J. Mol. Sci. 2019, 20, 2680. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, X.; Yang, C.; Jiang, Y.; Zhou, W.; Zheng, W. Artemisinin Attenuates Amyloid-Induced Brain Inflammation and Memory Impairments by Modulating TLR4/NF-κB Signaling. Int. J. Mol. Sci. 2022, 23, 6354. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, S.; Gaur, U.; Zheng, W. Artemisinin Improved Neuronal Functions in Alzheimer’s Disease Animal Model 3xtg Mice and Neuronal Cells via Stimulating the ERK/CREB Signaling Pathway. Aging Dis. 2020, 11, 801–819. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dai, X.; Qi, Y.; He, Y.; Du, W.; Pang, J.J. Histone Deacetylases Inhibitors in the Treatment of Retinal Degenerative Diseases: Overview and Perspectives. J. Ophthalmol. 2015, 2015, 250812. [Google Scholar] [CrossRef]
- Niu, Y.; DesMarais, T.L.; Tong, Z.; Yao, Y.; Costa, M. Oxidative stress alters global histone modification and DNA methylation. Free Radic. Biol. Med. 2015, 82, 22–28. [Google Scholar] [CrossRef]
- Chen, S.; Wang, M.; Jian, R.; Li, H.; Liu, G.; Zhou, C.; Xiong, Y.; Wang, W. Circ_HIPK3 Inhibits H2O2-Induced Lens Epithelial Cell Injury in Age-Related Cataract Depending on the Regulation of miR-495-3p/HDAC4 Pathway. Biochem. Genet. 2023, 61, 565–577. [Google Scholar] [CrossRef]
- Li, T.; Zhang, X.; Jiang, K.; Liu, J.; Liu, Z. Dural effects of oxidative stress on cardiomyogenesis via Gata4 transcription and protein ubiquitination. Cell Death Dis. 2018, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Song, M.J.; Park, J.H.; Shin, M.H.; Kim, M.K.; Hwang, D.; Lee, D.H.; Chung, J.H. Histone deacetylase 4 reverses cellular senescence via DDIT4 in dermal fibroblasts. Aging 2022, 14, 4653–4672. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yin, Y. Inhibition of histone deacetylase protects the damaged cataract via regulating the NF-kappa B pathway in cultured lens epithelial cells. Eur. J. Inflamm. 2019, 17, 2058739219870093. [Google Scholar] [CrossRef]
- Koriyama, Y.; Sugitani, K.; Ogai, K.; Kato, S. Neuritogenic activity of trichostatin A in adult rat retinal ganglion cells through acetylation of histone H3 lysine 9 and RARbeta induction. J. Pharmacol. Sci. 2014, 124, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Sancho-Pelluz, J.; Paquet-Durand, F. HDAC inhibition prevents Rd1 mouse photoreceptor degeneration. Adv. Exp. Med. Biol. 2012, 723, 107–113. [Google Scholar] [PubMed]
- Trifunović, D.; Petridou, E.; Comitato, A.; Marigo, V.; Ueffing, M.; Paquet-Durand, F. Primary Rod and Cone Degeneration Is Prevented by HDAC Inhibition. Retin. Degener. Dis. Mech. Exp. Ther. 2018, 1074, 367–373. [Google Scholar]
- Crosson, C.E.; Mani, S.K.; Husain, S.; Alsarraf, O.; Menick, D.R. Inhibition of histone deacetylase protects the retina from ischemic injury. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3639–3645. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Name | Clonality | Source | Dilution | Company (Catalog) |
|---|---|---|---|---|
| GAPDH | Monoclonal | Mouse | 1:1000 | SAB (40,493) |
| Acetyl-Histone 4 (Lys 8) | Polyclonal | Rabbit | 1:1000 | CST (2594) |
| Acetyl-Histone 3 (Lys 9) | Polyclonal | Rabbit | 1:1000 | CST (9671) |
| Bcl-2 | Monoclonal | Rabbit | 1:1000 | CST (3498) |
| Bax | Polyclonal | Rabbit | 1:1000 | CST (2772) |
| HDAC1 | Polyclonal | Rabbit | 1:1000 | CST (2062) |
| HDAC2 | Polyclonal | Rabbit | 1:1000 | CST (2540) |
| HDAC3 | Polyclonal | Rabbit | 1:1000 | CST (2632) |
| HDAC4 | Polyclonal | Rabbit | 1:1000 | CST (2072) |
| HDAC5 | Polyclonal | Rabbit | 1:1000 | CST (2082) |
| HDAC7 | Polyclonal | Rabbit | 1:1000 | CST (2882) |
| Anti-mouse IgG HRP-conjugated, secondary antibody | Polyclonal | Horse | 1:2000 | CST (7076) |
| Anti-rabbit IgG HRP-conjugated, secondary antibody | Polyclonal | Goat | 1:2000 | SAB (L3012) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Ge, L.; Yu, X.; Lazarovici, P.; Zheng, W. Artemisinin Confers Cytoprotection toward Hydrogen Peroxide-Induced Cell Apoptosis in Retinal Pigment Epithelial Cells in Correlation with the Increased Acetylation of Histone H4 at Lysine 8. Molecules 2024, 29, 1789. https://doi.org/10.3390/molecules29081789
Yang C, Ge L, Yu X, Lazarovici P, Zheng W. Artemisinin Confers Cytoprotection toward Hydrogen Peroxide-Induced Cell Apoptosis in Retinal Pigment Epithelial Cells in Correlation with the Increased Acetylation of Histone H4 at Lysine 8. Molecules. 2024; 29(8):1789. https://doi.org/10.3390/molecules29081789
Chicago/Turabian StyleYang, Chao, Lijun Ge, Xiyong Yu, Philip Lazarovici, and Wenhua Zheng. 2024. "Artemisinin Confers Cytoprotection toward Hydrogen Peroxide-Induced Cell Apoptosis in Retinal Pigment Epithelial Cells in Correlation with the Increased Acetylation of Histone H4 at Lysine 8" Molecules 29, no. 8: 1789. https://doi.org/10.3390/molecules29081789
APA StyleYang, C., Ge, L., Yu, X., Lazarovici, P., & Zheng, W. (2024). Artemisinin Confers Cytoprotection toward Hydrogen Peroxide-Induced Cell Apoptosis in Retinal Pigment Epithelial Cells in Correlation with the Increased Acetylation of Histone H4 at Lysine 8. Molecules, 29(8), 1789. https://doi.org/10.3390/molecules29081789