Mechanism of Mulberry Leaves and Black Sesame in Alleviating Slow Transit Constipation Revealed by Multi-Omics Analysis

 ,
,
Abstract
1. Introduction
2. Results
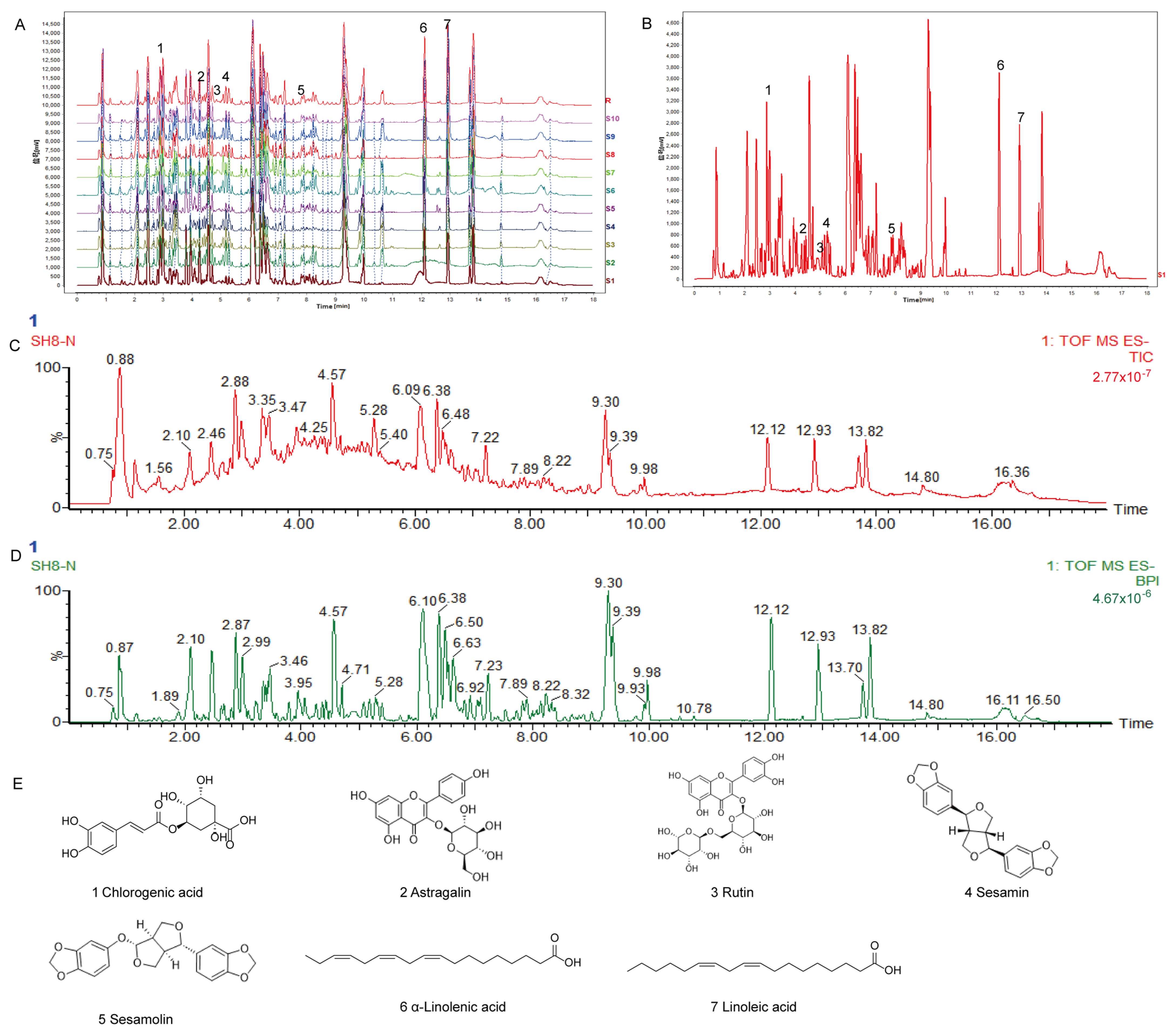
2.1. SH Fingerprint and Quantitative Analysis
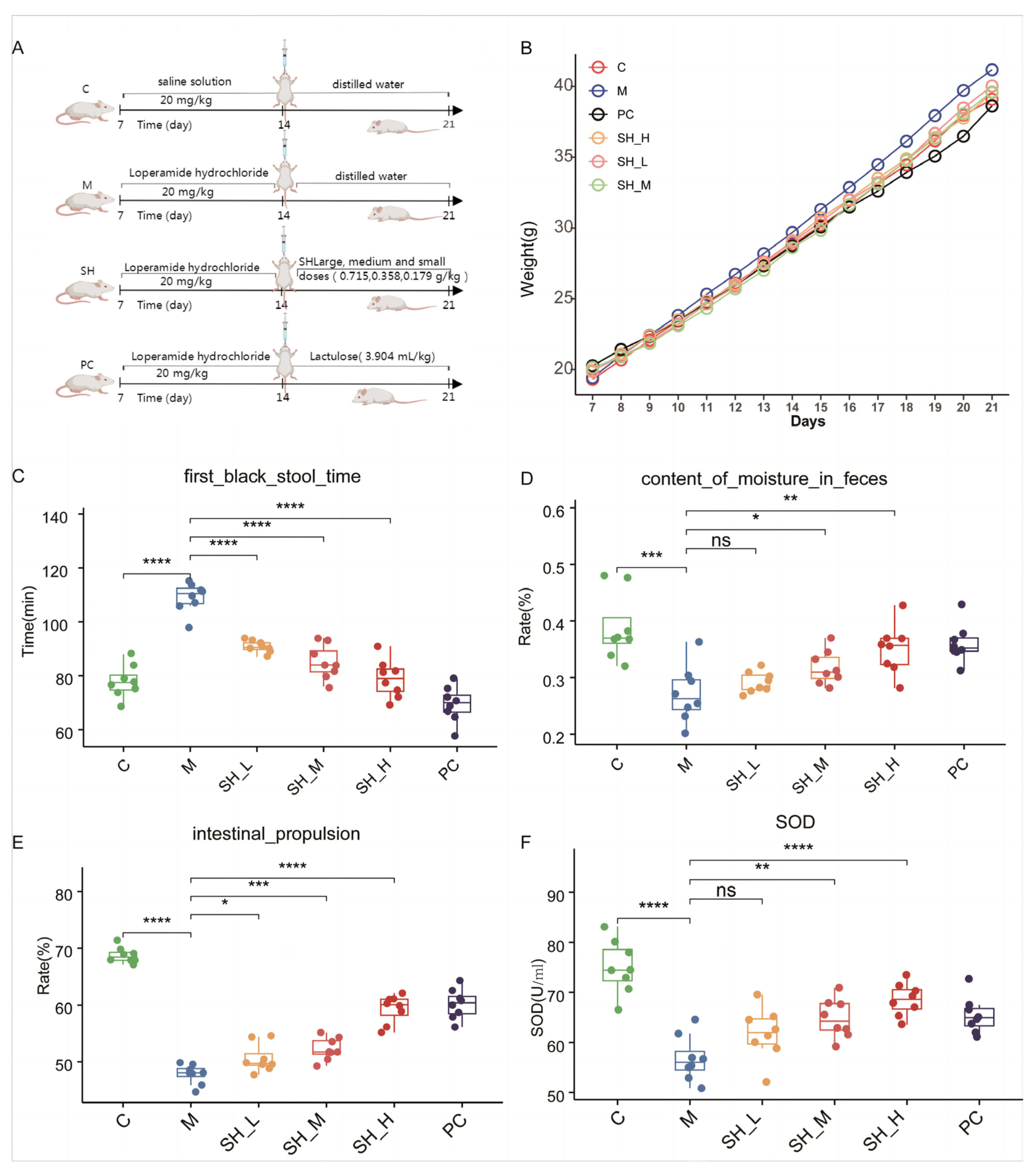
2.2. SH Relieves STC Symptoms in Mice
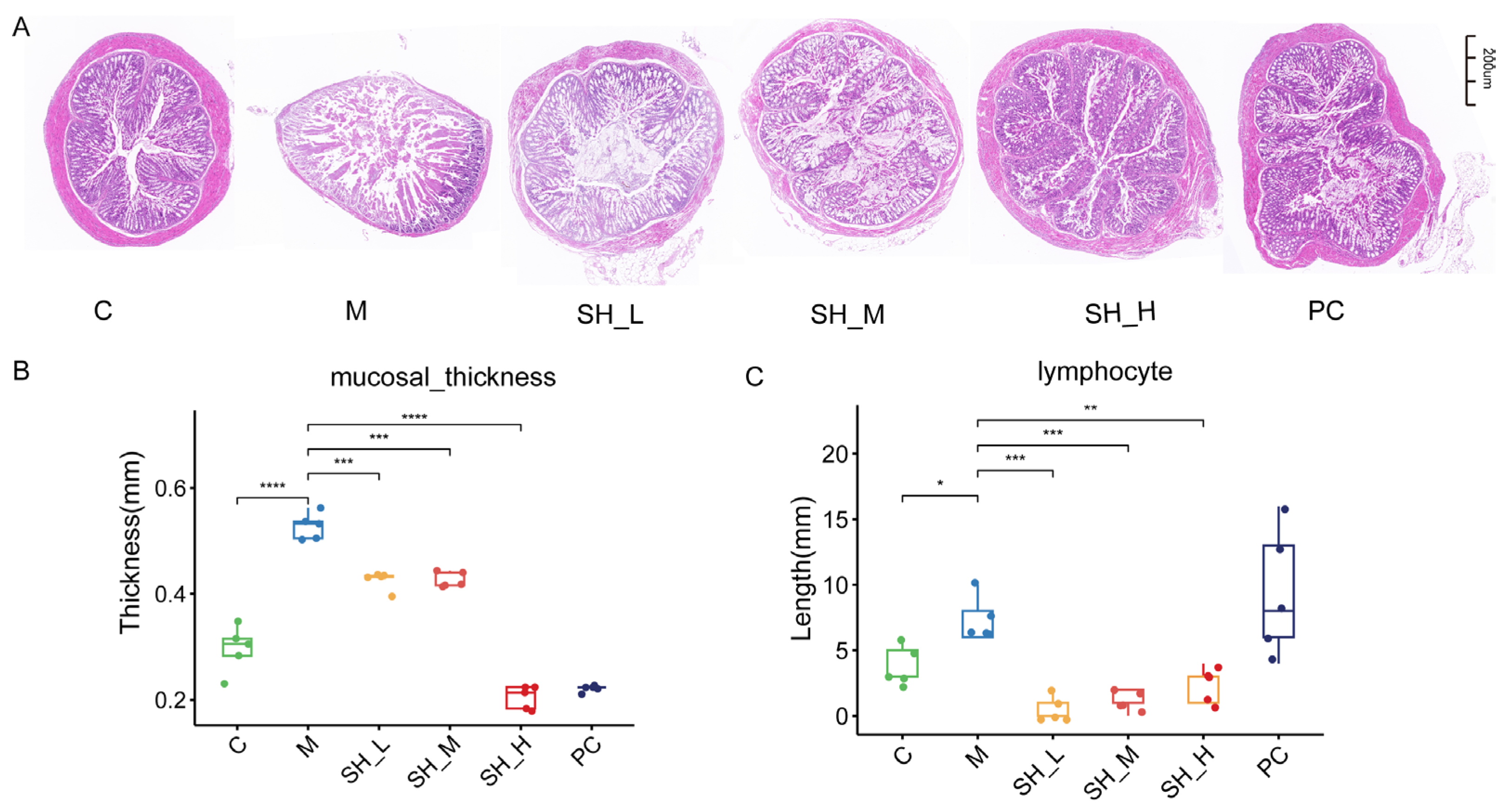
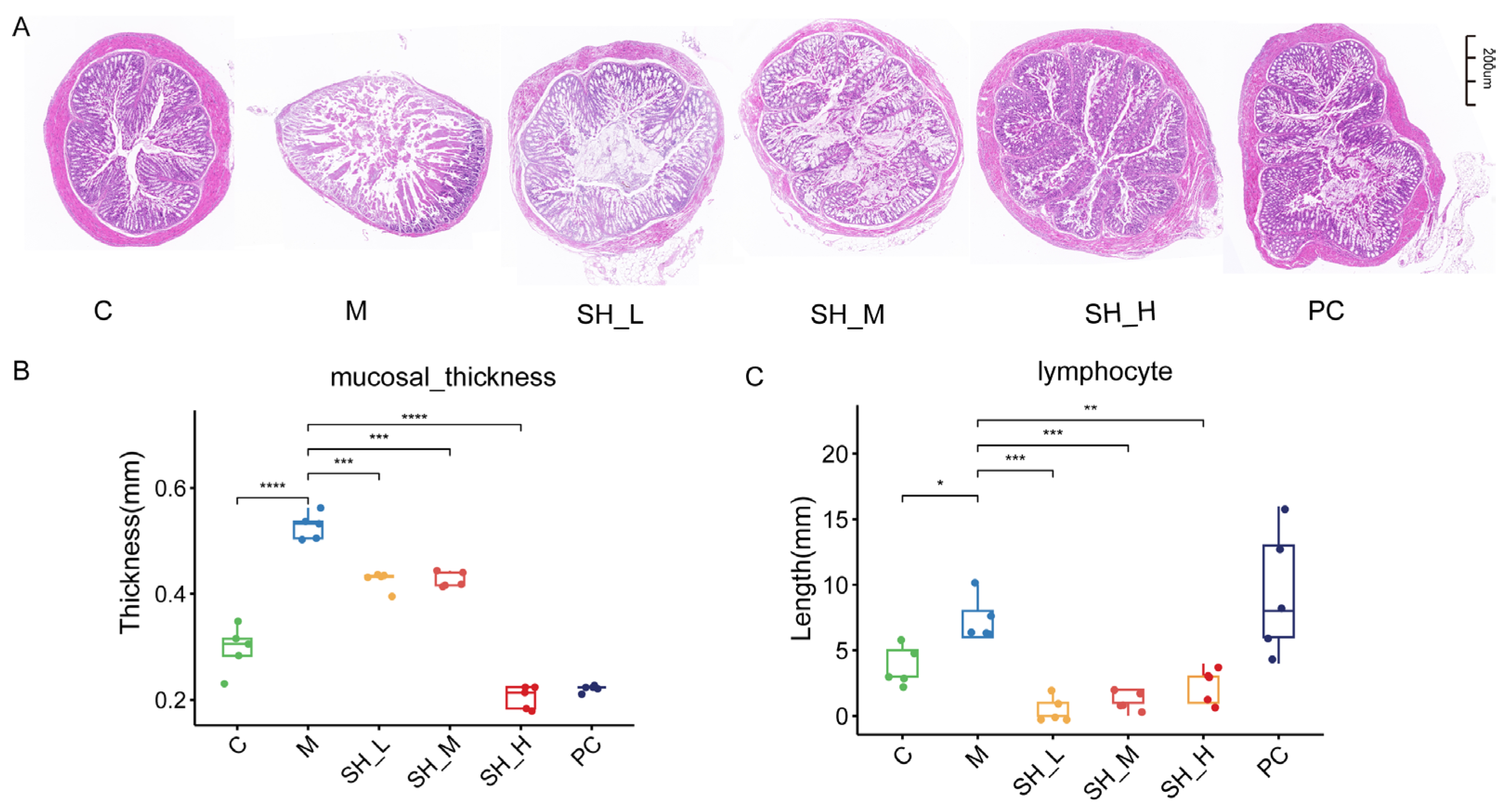
2.3. SH Can Alleviate Colonic Injury Caused by STC
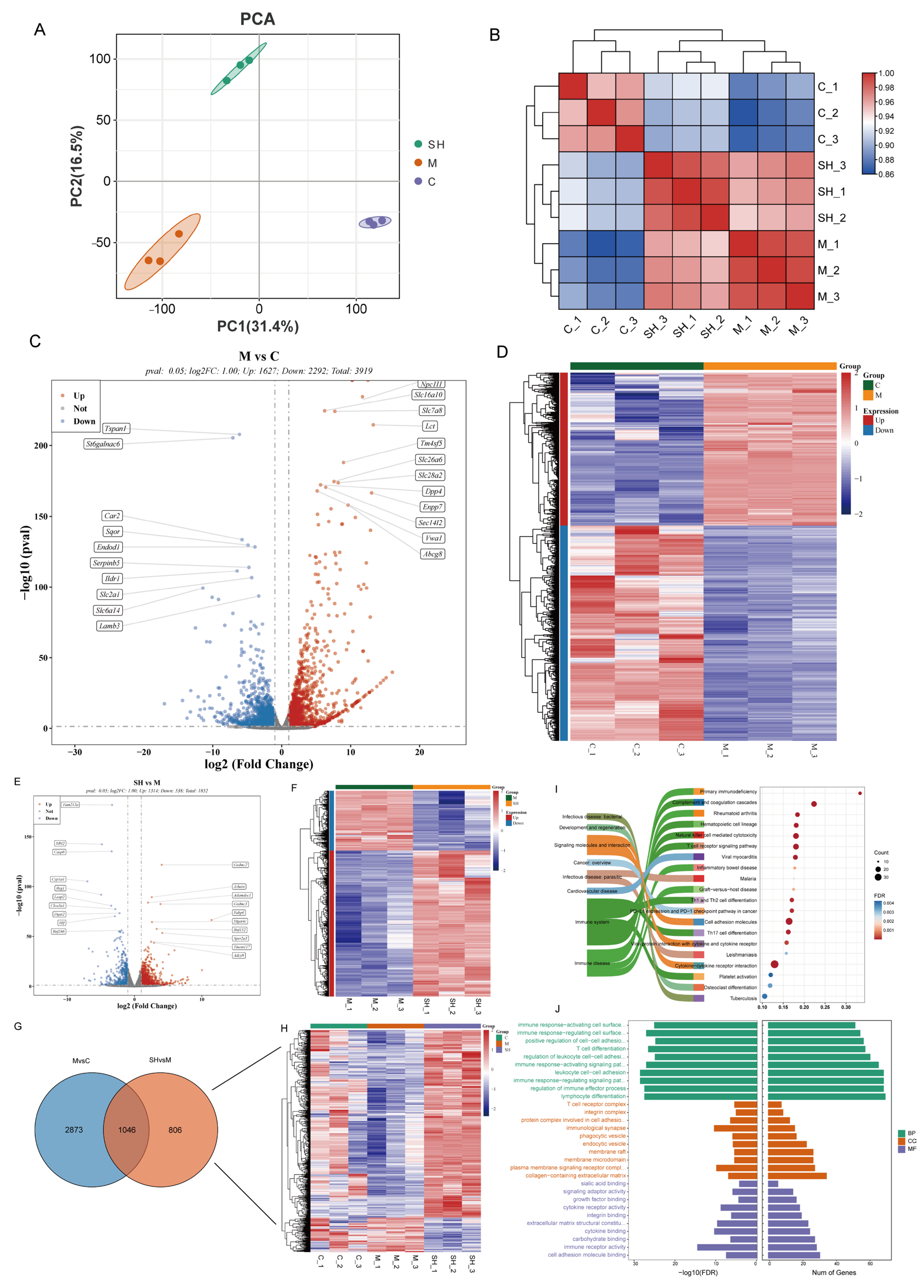
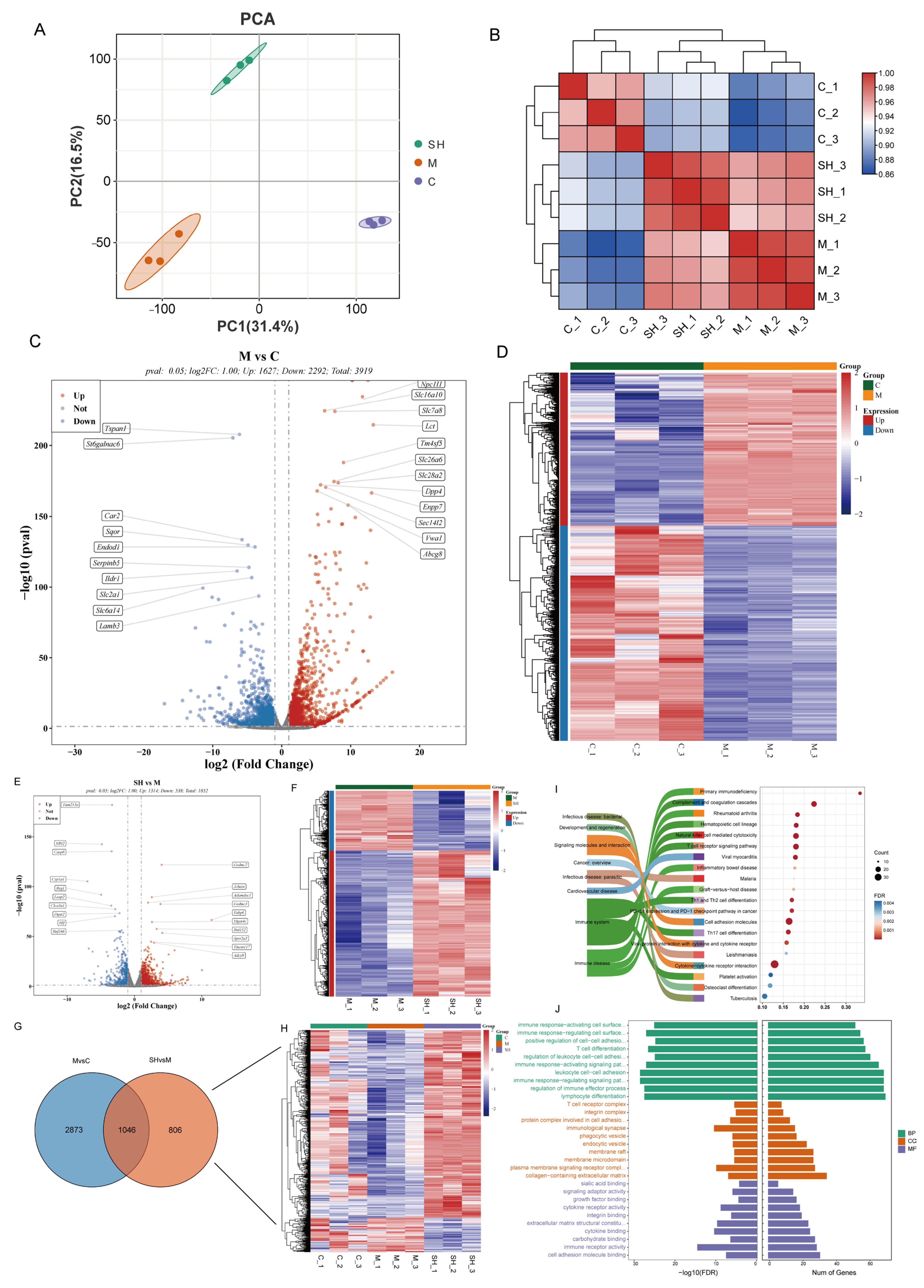
2.4. Transcriptomic Analysis of SH on STC Mice
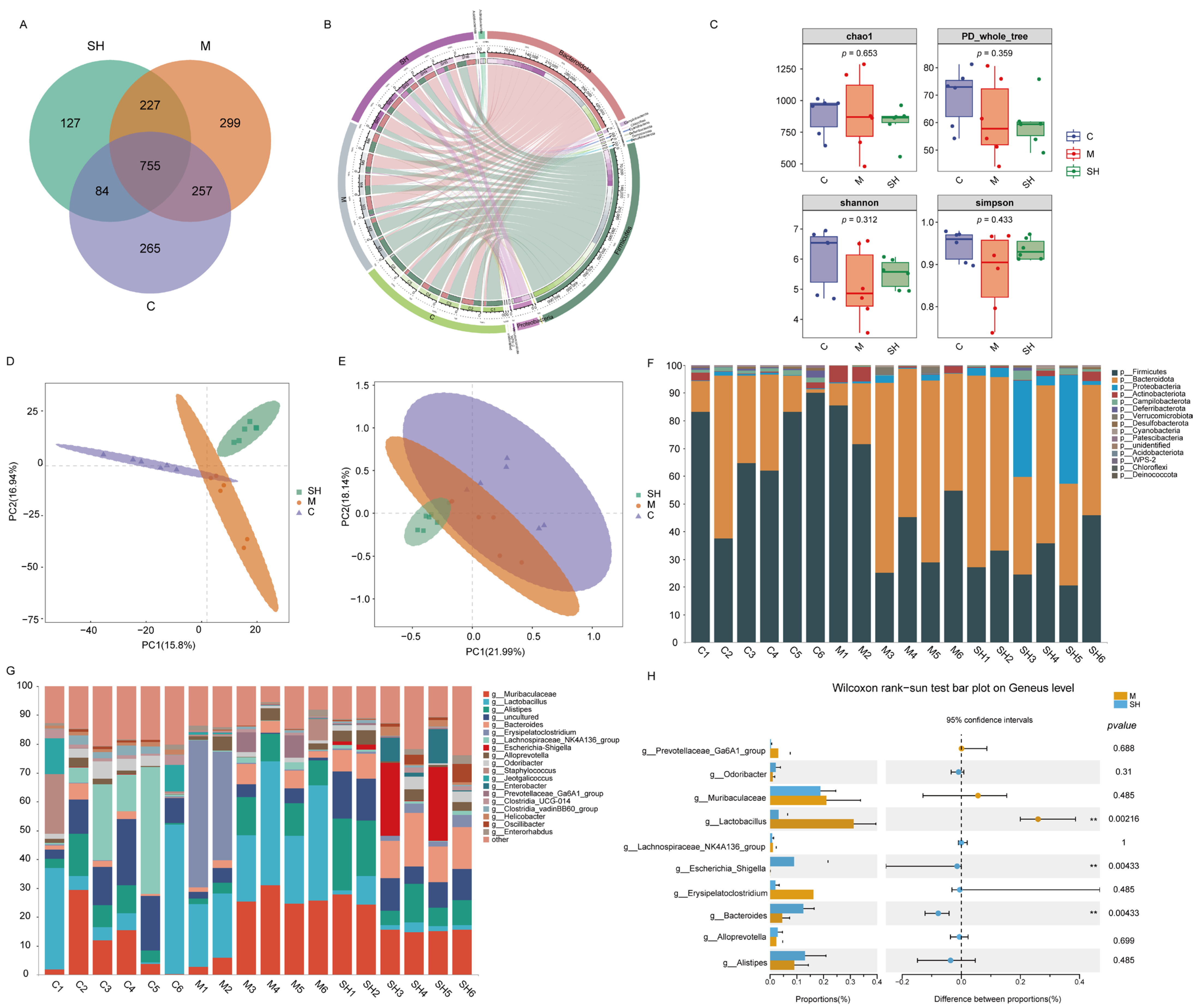
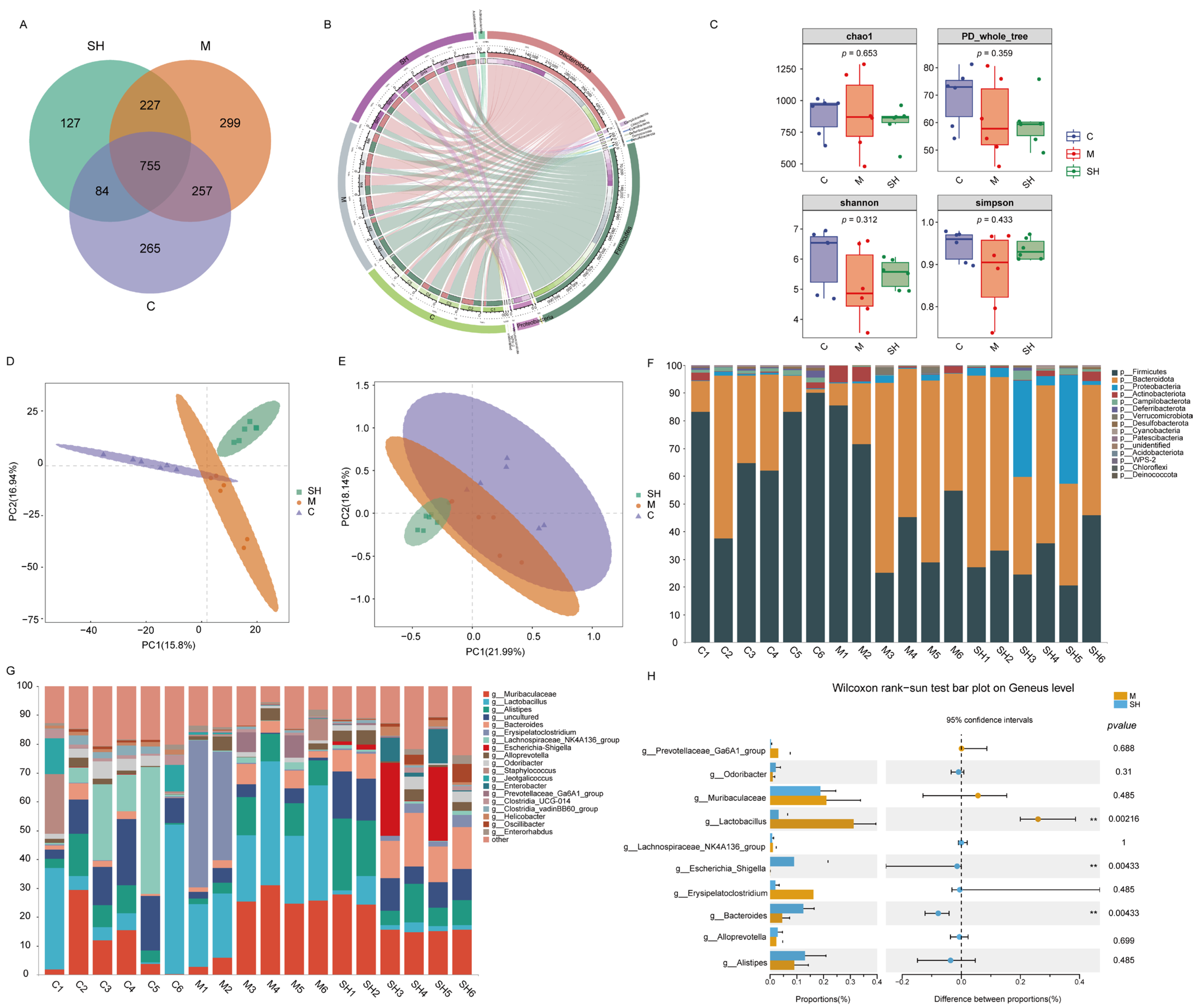
2.5. 16S rRNA Sequencing Analysis of SH on STC Mice
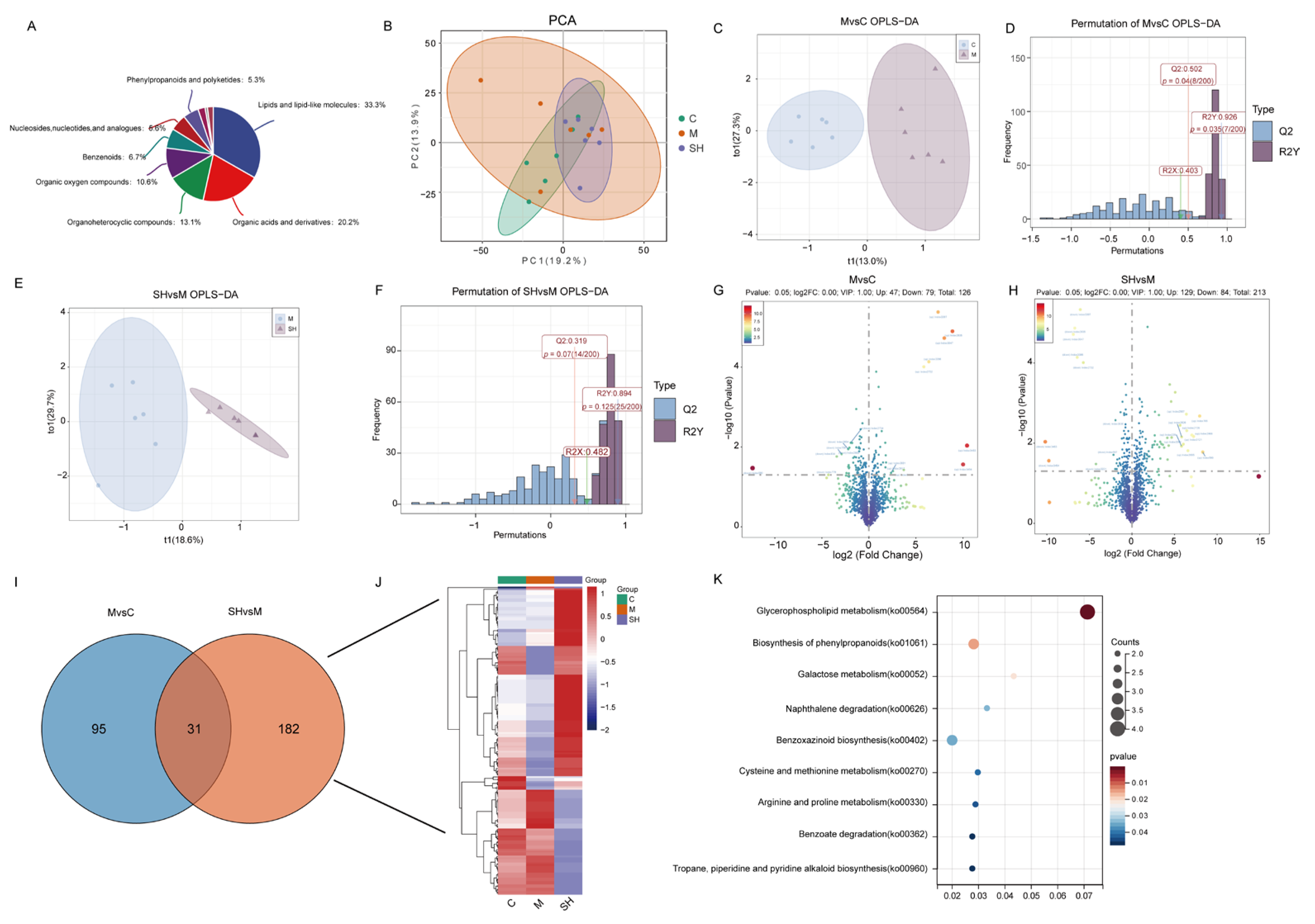
2.6. Metabolomics Analysis of SH on STC Mice

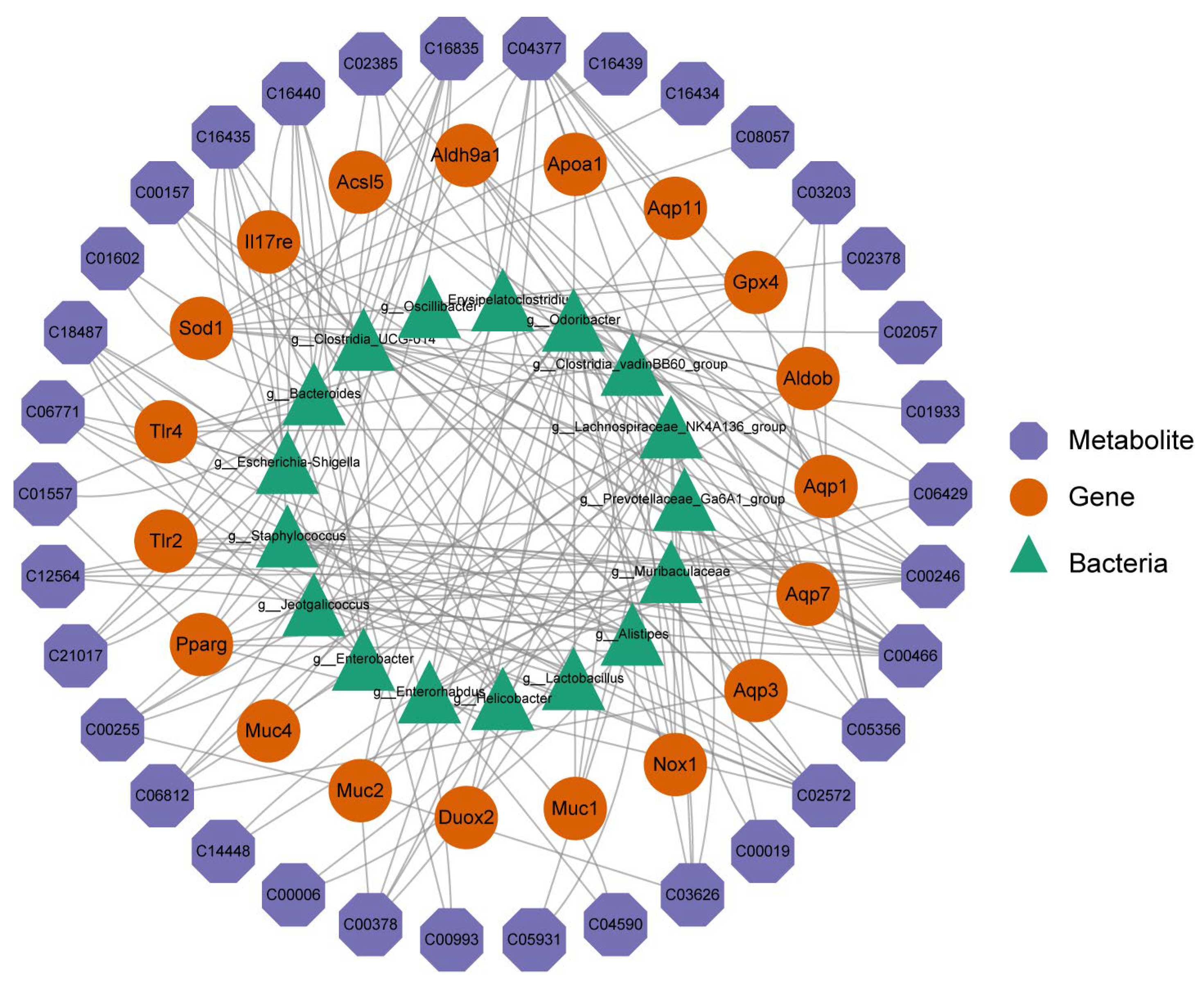
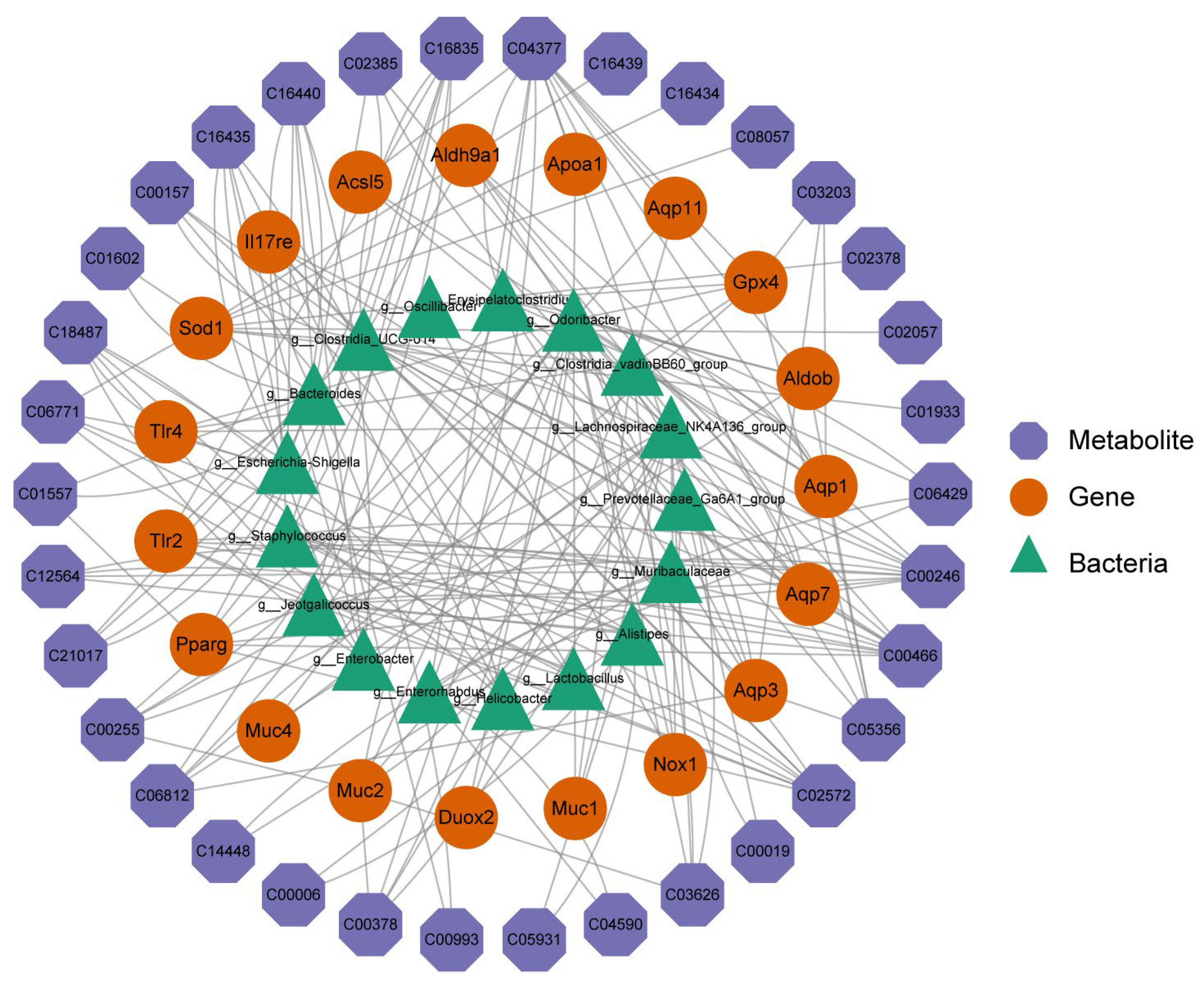
2.7. Combined Transcriptomics, 16S rRNA Sequencing, and Metabolomics Analysis of SH on STC Mice
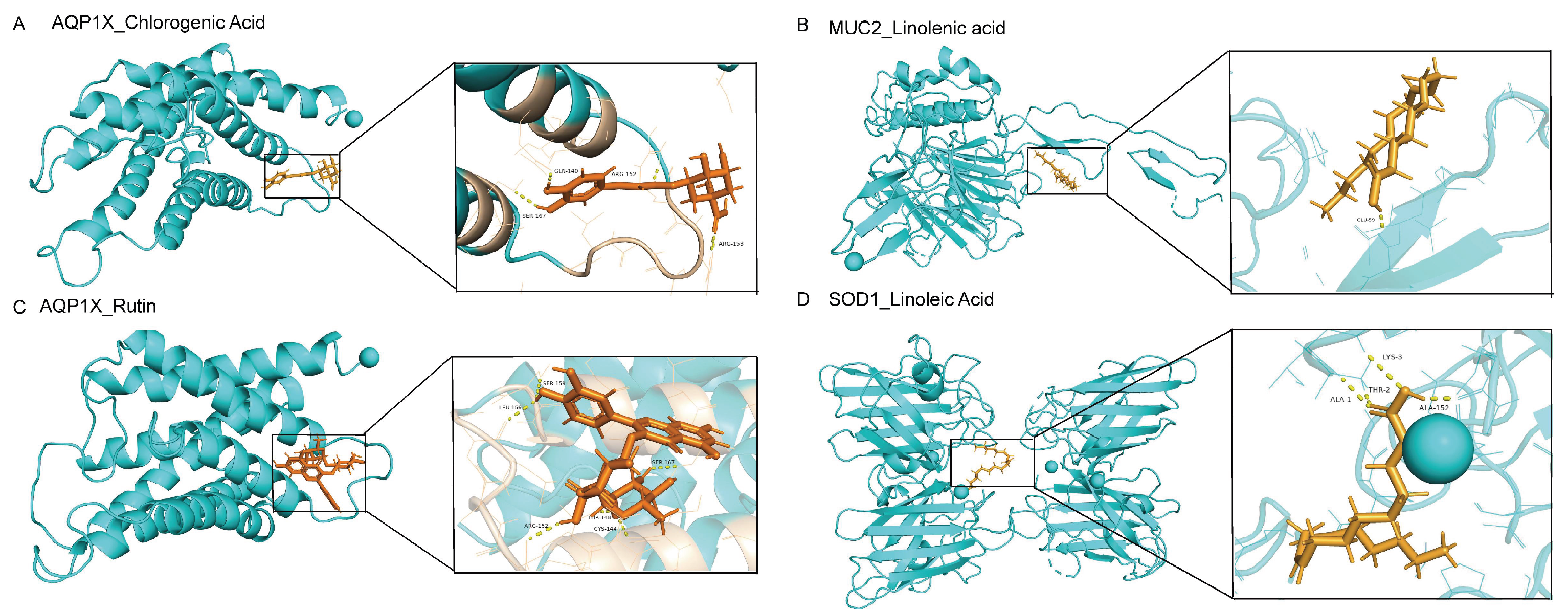
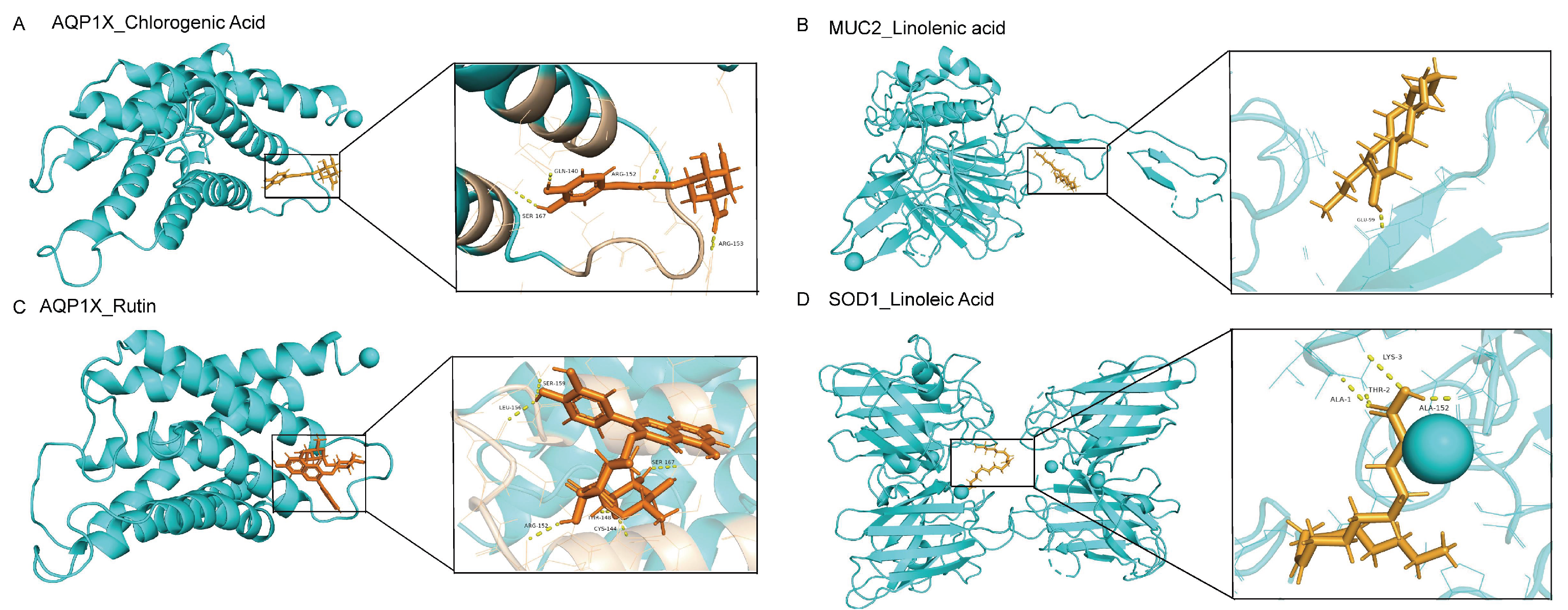
2.8. Verification of SH Core Compounds and Core Protein Targets by Molecular Docking
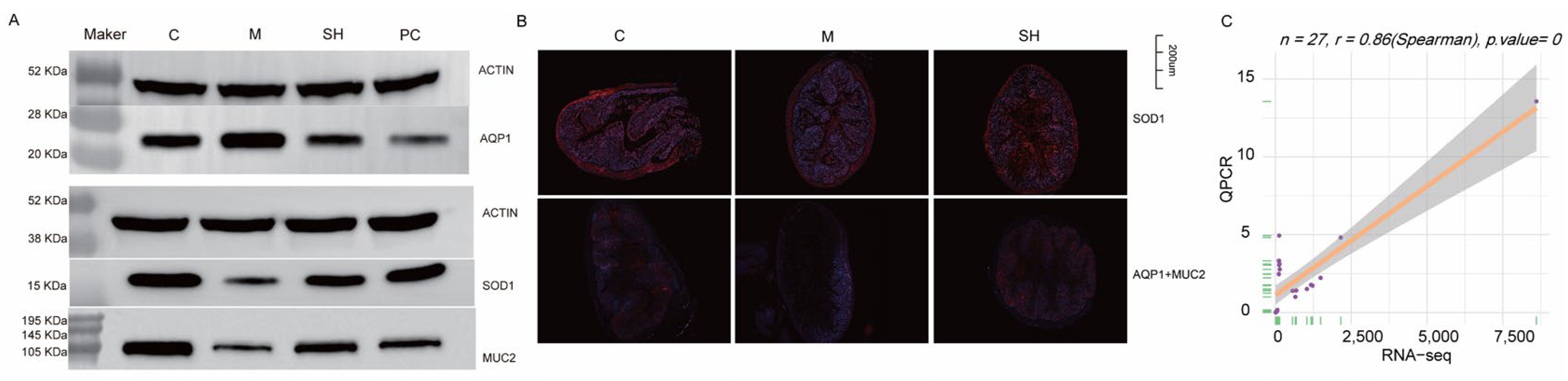
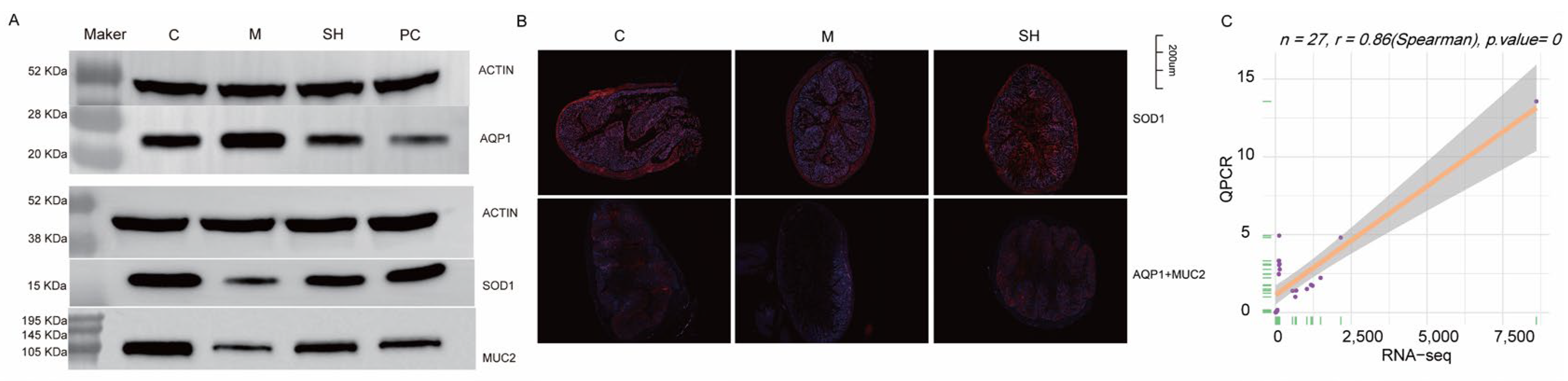
2.9. SH Inhibited AQP1 Expression and Promoted SOD1 and MUC2 Expression
3. Discussion
4. Materials and Methods
4.1. Reagents and Materials
4.2. Animal Experiments
4.3. Studies on the Fingerprint of SH
4.4. Quantitative Analysis and Detection of SH Using High-Performance Liquid Chromatography (HPLC)
4.5. Hematoxylin–Eosin (HE) Staining
4.6. Transcriptomics Analysis of SH Intervention in STC Mouse Colon
4.7. 16S rRNA Sequencing and Data Analysis
4.8. Metabonomics Analysis
4.9. Immunofluorescence Double-Label Staining Analysis
4.10. Western Blotting
4.11. Superoxide Dismutase (SOD) Levels
4.12. Quantitative Real-Time PCR
4.13. Molecular Docking
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.Z.; Jian, S.N.; Li, Y.C. Intervention effect of benzoylmesaconine on slow transit constipation in rats. J. Biol. Regul. Homeost. Agents 2023, 10, 5641–5651. [Google Scholar]
- Yao, Z.W.; Fu, S.Q.; Ren, B.B.; Ma, L.S.; Sun, D.Q. Based on Network Pharmacology and Gut Microbiota Analysis to Investigate the Mechanism of the Laxative Effect of Pterostilbene on Loperamide-Induced Slow Transit Constipation in Mice. Front. Pharmacol. 2022, 13, 913420. [Google Scholar] [CrossRef]
- Chang, X.-Y.; Liu, Y.-Y.; Hu, M.-M.; Liu, Y.-Q.; Jiang, C.-H.; Wang, Q.; Jin, Q.-M.; Zhang, D.-J.; Yin, Z.-Q.; Zhang, J. Comparative effects of different enzymatic hydrolysates of konjac glucomannan on gut flora and constipation in rats. Food Funct. 2022, 13, 8717–8729. [Google Scholar] [CrossRef]
- Lin, C.H.; He, H.Q.; Kim, J.J.; Zheng, X.; Huang, Z.H.; Dai, N. Osmotic pressure induces translocation of aquaporin-8 by P38 and JNK MAPK signaling pathways in patients with functional constipation. Dig. Liver Dis. 2023, 55, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Mu, K.W.; Kitts, D.D. Intestinal polyphenol antioxidant activity involves redox signaling mechanisms facilitated by aquaporin activity. Redox Biol. 2023, 68, 102948. [Google Scholar] [CrossRef]
- Yao, Y.K.; Kim, G.; Shafer, S.; Chen, Z.J.; Kubo, S.; Ji, Y.L.; Luo, J.L.; Yang, W.M.; Perner, S.P.; Kanellopoulou, C.; et al. Mucus sialylation determines intestinal host-commensal homeostasis. Cell 2022, 185, 1172–1188. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, N.; Chen, Q.Y.; Qin, H.L. Fecal Microbiota Transplantation Modulates the Gut Flora Favoring Patients with Functional Constipation. Front. Microbiol. 2021, 12, 700718. [Google Scholar] [CrossRef]
- Kim, J.E.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Lee, Y.J.; Sung, J.E.; Jung, Y.S.; Lee, H.S.; Hong, J.T.; Hwang, D.Y. Antioxidant activity and laxative effects of tannin-enriched extract of Ecklonia cava in loperamide-induced constipation of SD rats. PLoS ONE 2021, 16, e0246363. [Google Scholar] [CrossRef] [PubMed]
- Forget, P.; Vermeersch, M. To what extent are we confident that tapentadol induces less constipation and other side effects than the other opioids in chronic pain patients? a confidence evaluation in network meta-analysis. Br. J. Pain 2021, 15, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Wu, F.; Hong, Y.L.; Shen, L.; Zhao, L.J.; Lin, X. Research progress in the treatment of slow transit constipation by traditional Chinese medicine. J. Ethnopharmacol. 2022, 290, 115075. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.T.; Fu, Z.T.; Yan, W.; Nie, K.C.; Ding, L.L.; Ma, D.H.; Huang, H.D.; Li, T.; Xie, J.X.; Fu, L.C. The different effects of Chinese Herb Solid Drink and lactulose on gut microbiota in rats with slow transit constipation induced by compound diphenoxylate. Food Res. Int. 2021, 143, 110273. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Deng, M.; Tian, H.; Liu, D.; Li, Y.; Liu, G.; Sun, B.; Guo, Y. Effects of Mulberry Leaves and Pennisetum Hybrid Mix-Silage on Fermentation Parameters and Bacterial Community. Fermentation 2022, 8, 197. [Google Scholar] [CrossRef]
- Akl, E.M. Oilseeds Dietary Fiber and Their Health Benefits. Egypt. J. Chem. 2023, 66, 459–471. [Google Scholar] [CrossRef]
- Hu, T.-G.; Wen, P.; Fu, H.-Z.; Lin, G.-Y.; Liao, S.-T.; Zou, Y.-X. Protective effect of mulberry (Morus atropurpurea) fruit against diphenoxylate-induced constipation in mice through the modulation of gut microbiota. Food Funct. 2019, 10, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.K.; Liu, X.; Hou, T.; Zhan, F.C.; Geng, F.; Zhang, Z.Y.; Li, B. Evaluation of the effect of prebiotic sesame candies on loperamide-induced constipation in mice. Food Funct. 2022, 13, 5690–5700. [Google Scholar] [CrossRef] [PubMed]
- Comini, E.; Rubiales, D.; Reveglia, P. Variability of Fatty Acid Composition and Lignan Content in Sesame Germplasm, and Effect of Roasting. ACS Food Sci. Technol. 2023, 3, 1747–1758. [Google Scholar] [CrossRef]
- Chen, C.; Mokhtar, R.A.M.; Sani, M.S.A.; Noor, N.Q.I.M. The Effect of Maturity and Extraction Solvents on Bioactive Compounds and Antioxidant Activity of Mulberry (Morus alba) Fruits and Leaves. Molecules 2022, 27, 2406. [Google Scholar] [CrossRef]
- Song, L.; Wu, T.; Zhang, L.; Wan, J.; Ruan, Z. Chlorogenic acid improves the intestinal barrier by relieving endoplasmic reticulum stress and inhibiting ROCK/MLCK signaling pathways. Food Funct. 2022, 13, 4562–4575. [Google Scholar] [CrossRef]
- Chen, J.-H.; Zhao, C.-L.; Li, Y.-S.; Yang, Y.-B.; Luo, J.-G.; Zhang, C.; Wang, L. Moutai distiller’s grains polyphenol extracts and rutin alleviate dss-induced colitis in mice: Modulation of gut microbiota and intestinal barrier function (r2). Heliyon 2023, 9, e22186. [Google Scholar] [CrossRef]
- Xu, Q.; Zhao, J.; Jian, H.; Ye, J.; Gong, M.; Zou, X.; Dong, X. Linoleic acid ameliorates intestinal mucosal barrier injury in early weaned pigeon squabs (Columba livia). J. Anim. Sci. 2023, 101, skad125. [Google Scholar] [CrossRef] [PubMed]
- Sayhan, M.B.; Oguz, S.; Salt, O.; Can, N.; Ozgurtas, T.; Yalta, T.D. Sesamin ameliorates mucosal tissue injury of mesenteric ischemia and reperfusion in an experimental rat model. Arch. Med. Sci. 2019, 15, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhan, Y.; Tang, S.Y.; Liu, F.; Wu, R.; Kong, P.F.; Li, Q.; Tang, X.G. Zhizhu decoction alleviates slow transit constipation by regulating aryl hydrocarbon receptor through gut microbiota. Pharm. Biol. 2023, 61, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, E.U.; Ozel, A.; Erol, M.; Tenekecigil, A.; Gayret, O.B.; Buke, O.; Tosun, V. Comparison of effects of quercetin and ascorbic acid on inflammatory cytokines and antioxidant biomarkers in infant rats using an experimental sepsis model. Bratisl. Med. J. 2023, 124, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-W.; Choi, J.; Ryu, K.-Y. Recent progress and future directions of the research on nanoplastic-induced neurotoxicity. Neural Regen. Res. 2024, 19, 331–335. [Google Scholar] [CrossRef]
- Lin, R.T.; Wu, W.; Chen, H.M.; Gao, H.; Wu, X.H.; Li, G.F.; He, Q.; Lu, H.Y.; Sun, M.M.; Liu, Z.J. GPR65 promotes intestinal mucosal Th1 and Th17 cell differentiation and gut inflammation through downregulating NUAK2. Clin. Transl. Med. 2022, 12, e771. [Google Scholar] [CrossRef]
- Li, X.Y.; Wang, F.; Gao, Z.F.; Huang, W.J.; Zhang, X.M.; Liu, F.; Yi, H.L.; Guan, J.; Wu, X.L.; Xu, H.J.; et al. Melatonin attenuates chronic intermittent hypoxia-induced intestinal barrier dysfunction in mice. Microbiol. Res. 2023, 276, 127480. [Google Scholar] [CrossRef] [PubMed]
- Varma, S.R.; Sivaprakasam, T.O.; Arumugam, I.; Dilip, N.; Raghuraman, M.; Pavan, K.B.; Rafiq, M.; Paramesh, R. In vitro anti-inflammatory and skin protective properties of Virgin coconut oil. J. Tradit. Complement. Med. 2019, 9, 5–14. [Google Scholar] [CrossRef]
- Gandhi, J.; Sushma, M.V.; Rengan, A.K.; Naik, M.N.; Mishra, D.K.; Boyinpally, S.R.; Joseph, J. Proteomic profiling of exosomes in a mouse model of Candida albicans endophthalmitis. Exp. Cell Res. 2022, 417, 113222. [Google Scholar] [CrossRef]
- Jia, Z.-F.; Wang, J.-L.; Pan, W.; Hu, J. Croton tiglium L. seeds ameliorate loperamide-induced constipation via regulating gastrointestinal hormones and gut microbiota before and after processing. J. Ethnopharmacol. 2024, 319, 117378. [Google Scholar] [CrossRef]
- Wang, L.L.; Yang, S.R.; Mei, C.X.; Tang, N.; Wang, J.L.; Yu, Q.Q.; Wang, G.; Wu, G.J.; Zhao, J.X.; Chen, W. Lactobacillus paracasei Relieves Constipation by Acting on the Acetic Acid-5-HT-Intestinal Motility Pathway. Foods 2023, 12, 4176. [Google Scholar] [CrossRef] [PubMed]
- Ghiasi, F.; Hashemi, S.M.B.; Abedi, E. Effective enhancement of food oxidative stability induced by Lactobacillus strains: In vitro activity. Food Control 2023, 153, 109912. [Google Scholar] [CrossRef]
- Baltazar-Diaz, T.A.; Gonzalez-Hernandez, L.A.; Aldana-Ledesma, J.M.; Pena-Rodriguez, M.; Vega-Magana, A.N.; Zepeda-Morales, A.S.M.; Lopez-Roa, R.I.; del Toro-Arreola, S.; Martinez-Lopez, E.; Salazar-Montes, A.M.; et al. Escherichia/Shigella, SCFAs, and metabolic pathways-the triad that orchestrates intestinal dysbiosis in patients with decompensated alcoholic cirrhosis from Western Mexico. Microorganisms 2022, 10, 1231. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.S.; Kong, C.; Gao, R.Y.; Yan, X.B.; Yu, H.J.; Wen, B.; Zhu, Q.; Shen, T.Y.; Sun, Z.L.; Qin, H.L. Analysis of fecal microbiota in patients with functional constipation undergoing treatment with synbiotics. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 555–563. [Google Scholar] [CrossRef]
- Costa, L.M.; Mendes, M.M.; Oliveira, A.C.; Magalhães, K.G.; Shivappa, N.; Hebert, J.R.; da Costa, T.H.M.; Botelho, P.B. Dietary inflammatory index and its relationship with gut microbiota in individuals with intestinal constipation: A cross-sectional study. Eur. J. Nutr. 2022, 61, 341–355. [Google Scholar] [CrossRef]
- Engevik, M.A.; Engevik, A.C.; Engevik, K.A.; Auchtung, J.M.; Chang-Graham, A.L.; Ruan, W.; Luna, R.A.; Hyser, J.M.; Spinler, J.K.; Versalovic, J. Mucin-Degrading Microbes Release Monosaccharides That Chemoattract Clostridioides difficile and Facilitate Colonization of the Human Intestinal Mucus Layer. ACS Infect. Dis. 2021, 7, 1126–1142. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Ren, P.P.; Wang, M.L.; Wang, J.J.; He, X.Y.; Gu, J.Y.; Lu, Y.Y.; Wu, Y.N.; Liu, J.H.; Wang, L.D.; et al. Changes in intestinal barrier protein expression and intestinal flora in a rat model of visceral hypersensitivity. Neurogastroenterol. Motil. 2022, 34, e14299. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.Q.; Fan, L.L.; Zheng, N.; Blecker, C.; Delcenserie, V.; Li, H.Y.; Wang, J.Q. 2′-Fucosyllactose Ameliorates Inflammatory Bowel Disease by Modulating Gut Microbiota and Promoting MUC2 Expression. Front. Nutr. 2022, 9, 822020. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Li, X.; Sun-Waterhouse, D.X.; Zhu, B.Y.; You, L.J.; Hileuskaya, K. Polysaccharides from Sargassum fusiforme after UV/H2O2 degradation effectively ameliorate dextran sulfate sodium-induced colitis. Food Funct. 2021, 12, 11747–11759. [Google Scholar]
- Zeng, N.Y.; Wu, F.; Lu, J.Q.; Li, X.; Lin, S.M.; Zhou, L.; Wang, Z.W.; Wu, G.Y.; Huang, Q.F.; Zheng, D.W.; et al. High-fat diet impairs gut barrier through intestinal microbiota-derived reactive oxygen species. Sci. China Life Sci. 2023, 66, 1–15. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Yu, H.T.; Zhang, H.Y.; Zhao, Q.Y.; Si, W.; Qin, Y.C.; Zhang, J.M. Dietary Epimedium extract supplementation improves intestinal functions and alters gut microbiota in broilers. J. Anim. Sci. Biotechnol. 2023, 14, 14. [Google Scholar] [CrossRef]
- Liu, S.; Yang, C.; Li, H.; Bai, X.; Hu, T.; Xue, X.; An, J.; Zhang, Y.; Dong, X. Alteration of Serum Metabolites in Women of Reproductive Age with Chronic Constipation. Med. Sci. Monit. 2022, 28, e934117. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wei, Y.; Zhu, Y.; Guo, J.; Zhang, J.; He, Y.; Li, X.; Liu, J.; Zhou, W. The Interaction between Gut Microbiota and Host Amino Acids Metabolism in Multiple Myeloma. Cancers 2023, 15, 1942. [Google Scholar] [CrossRef]
- Meng, Y.; Li, X.; Wang, X.; Zhang, L.; Guan, J. Network pharmacological prediction and molecular docking analysis of the combination of Atractylodes macrocephala Koidz. and Paeonia lactiflora Pall. in the treatment of functional constipation and its verification. Anim. Models Exp. Med. 2022, 5, 120–132. [Google Scholar]
- Qi, B.; Zhang, Y.; Ren, D.; Qin, X.; Wang, N.; Yang, X. Fu Brick Tea Alleviates Constipation via Regulating the Aquaporins-Mediated Water Transport System in Association with Gut Microbiota. J. Agric. Food Chem. 2023, 71, 3862–3875. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Zhou, K.; Jiang, P.; Deng, N.; Peng, X.; Tan, Z. Brain-bacteria-gut axis and oxidative stress mediated by intestinal mucosal microbiota might be an important mechanism for constipation in mice. 3 Biotech 2023, 13, 192. [Google Scholar] [CrossRef]
- Rodrigues, C.; Bujak, I.T.; Mihaljević, B.; Soveral, G.; Gasparovic, A.C. Yeast aquaporin regulation by 4-hydroxynonenal is implicated in oxidative stress response. IUBMB Life 2017, 44, 355–362. [Google Scholar] [CrossRef]
- Gao, H.; Gao, C.-C.; Wang, T.-T.; Gao, L.; Li, G.-W.; Jin, L.-Y.; He, C.-W.; Wang, B.-Y.; Zhang, L.; Guo, Y.-X.; et al. An Unexpected Alteration Colonic Mucus Appearance in the Constipation Model via an Intestinal Microenvironment. Microsc. Microanal. 2022, 28, 1720–1733. [Google Scholar] [CrossRef]
- Takami, M.; Aoi, W.; Matsumoto, K.; Kato, Y.; Kobayashi, Y.; Kuwahata, M. High-intensity exercise impairs intestinal barrier function by generating oxidative stress. J. Clin. Biochem. Nutr. 2024, 74, 136–140. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Retention Time (min) | Linear Regression Equation | R2 | Content (mg/g) |
|---|---|---|---|---|
| Chlorogenic acid | 1.756 | y = 96.398x + 0.1338 | 0.999 | 1.028 |
| Astragalin | 3.931 | y = 69.121x − 0.1113 | 0.999 | 0.120 |
| Rutin | 3.573 | y = 42.638x − 0.0621 | 0.999 | 0.240 |
| Sesamin | 6.842 | y = 46.262x − 0.0258 | 1.000 | 0.837 |
| Sesamolin | 7.117 | y = 43.919x + 0.0459 | 0.999 | 0.266 |
| α-linolenic acid | 9.027 | y = 52.997x + 0.0994 | 0.999 | 0.397 |
| Linoleic acid | 9.569 | y = 53.942x + 0.361 | 0.999 | 2.164 |
| Gene | PDB ID | Compound | PubChem CID | Docking Score (kcal/mol) |
|---|---|---|---|---|
| AQP1 | 1h6i | Chlorogenic acid | 1,794,427 | −6.952 |
| AQP1 | 1h6i | Rutin | 5,280,805 | −8.183 |
| MUC2 | 7qcn | Linolenic acid | 5,282,457 | −7.277 |
| SOD1 | 3re0 | Linoleic acid | 5,280,450 | −7.047 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Wang, Z.; Tan, Y.; Li, L.; Zhou, F.; Hu, S.-A.; Yan, Q.-W.; Li, L.-H.; Pei, G. Mechanism of Mulberry Leaves and Black Sesame in Alleviating Slow Transit Constipation Revealed by Multi-Omics Analysis. Molecules 2024, 29, 1713. https://doi.org/10.3390/molecules29081713
Sun C, Wang Z, Tan Y, Li L, Zhou F, Hu S-A, Yan Q-W, Li L-H, Pei G. Mechanism of Mulberry Leaves and Black Sesame in Alleviating Slow Transit Constipation Revealed by Multi-Omics Analysis. Molecules. 2024; 29(8):1713. https://doi.org/10.3390/molecules29081713
Chicago/Turabian StyleSun, Chen, Zheng Wang, Yang Tan, Ling Li, Feng Zhou, Shi-An Hu, Qin-Wen Yan, Lin-Hui Li, and Gang Pei. 2024. "Mechanism of Mulberry Leaves and Black Sesame in Alleviating Slow Transit Constipation Revealed by Multi-Omics Analysis" Molecules 29, no. 8: 1713. https://doi.org/10.3390/molecules29081713
APA StyleSun, C., Wang, Z., Tan, Y., Li, L., Zhou, F., Hu, S.-A., Yan, Q.-W., Li, L.-H., & Pei, G. (2024). Mechanism of Mulberry Leaves and Black Sesame in Alleviating Slow Transit Constipation Revealed by Multi-Omics Analysis. Molecules, 29(8), 1713. https://doi.org/10.3390/molecules29081713