
Channel Expansion in the Ligand-Binding Domain of the Glucocorticoid Receptor Contributes to the Activity of Highly Potent Glucocorticoid Analogues

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Results
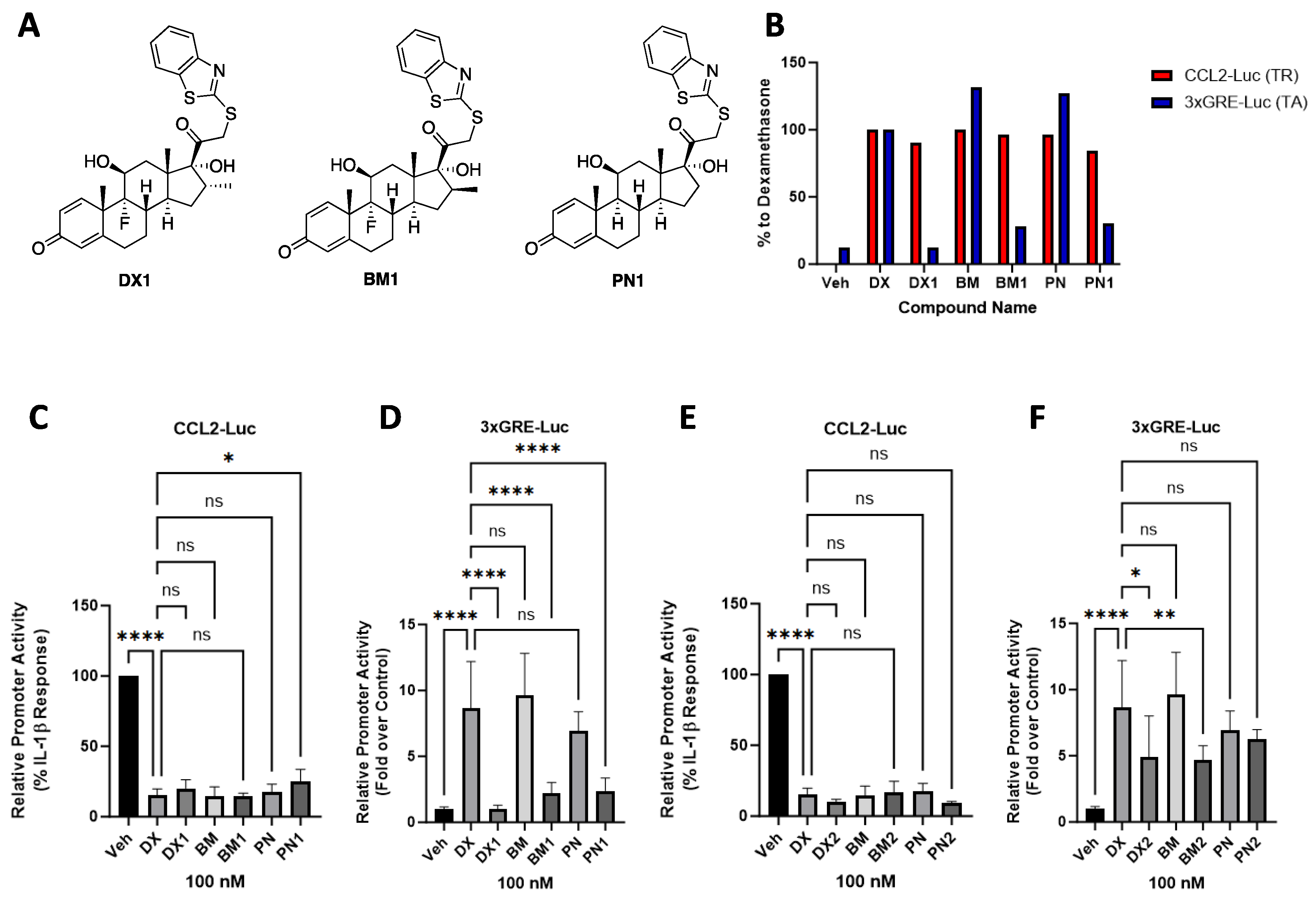
2.1. Series 1 GC Analogues Selectively Repress Ccl2 Promoter Activity without Inducing Transactivation
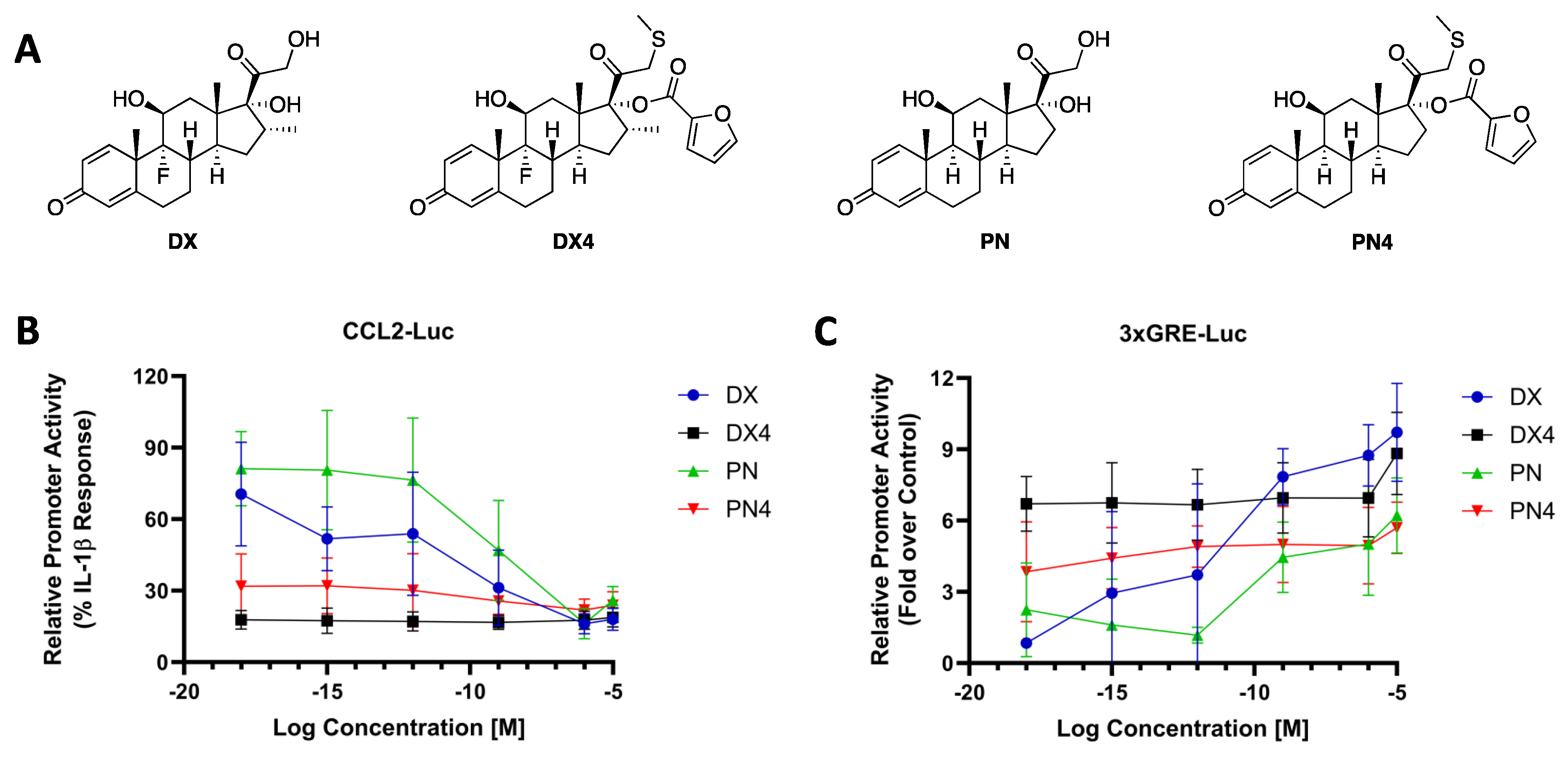
2.2. DX4 and PN4 Display Attomolar Potency versus Nanomolar Potency for Parent Scaffolds
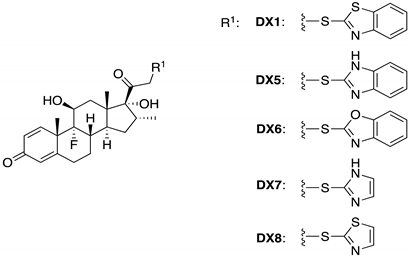
2.3. DX1 and DX4 Exhibit Reduced Toxicity When Compared with DX
2.4. Sgk1 and Rgs2 Gene Expression Induced at Low Concentrations in Response to DX4 and PN4, but Decreased upon MBT Addition
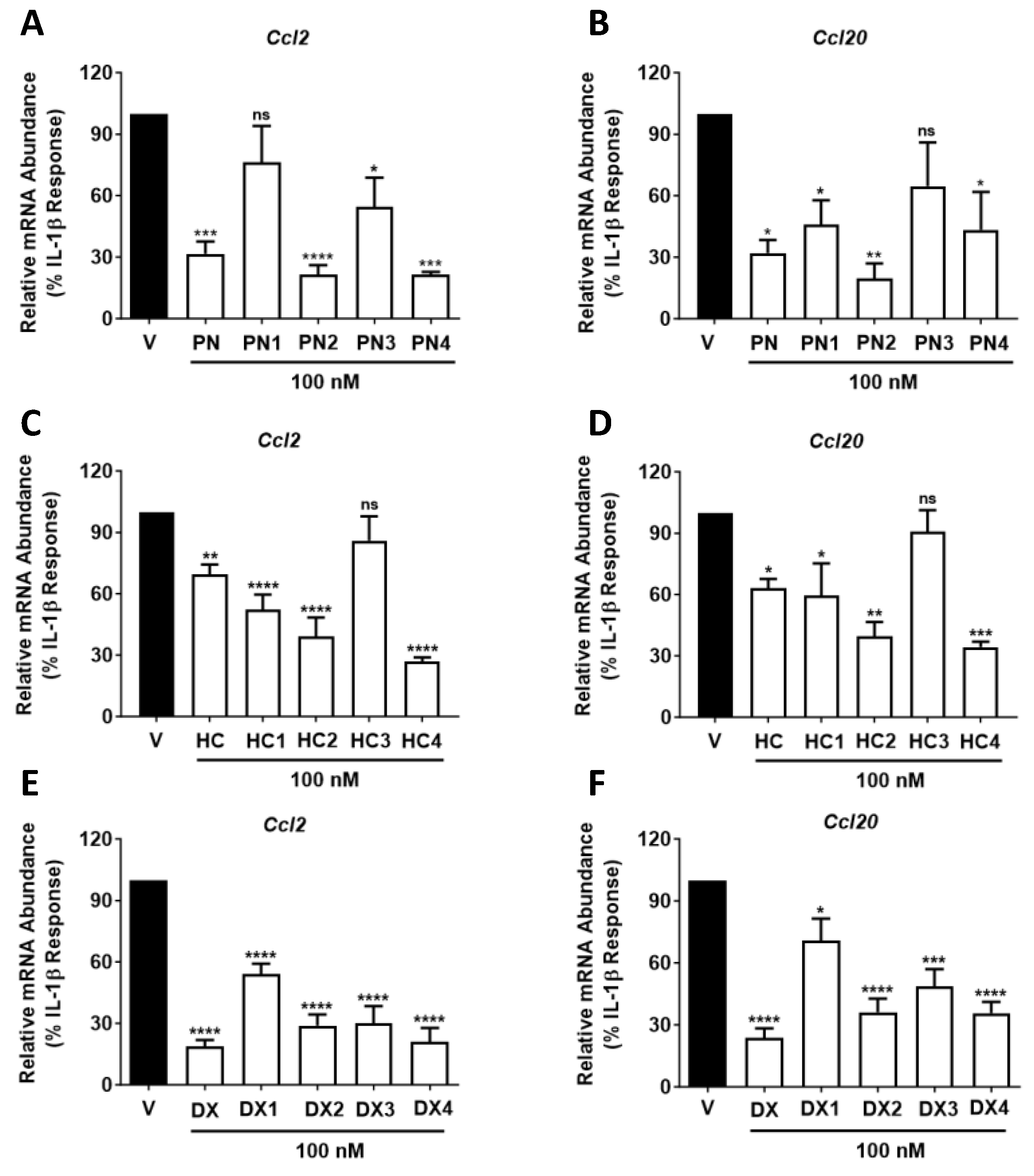
2.5. All GC Analogues, Except HC3, Reduce Ccl2 or Ccl20 Transcript and Protein Abundance
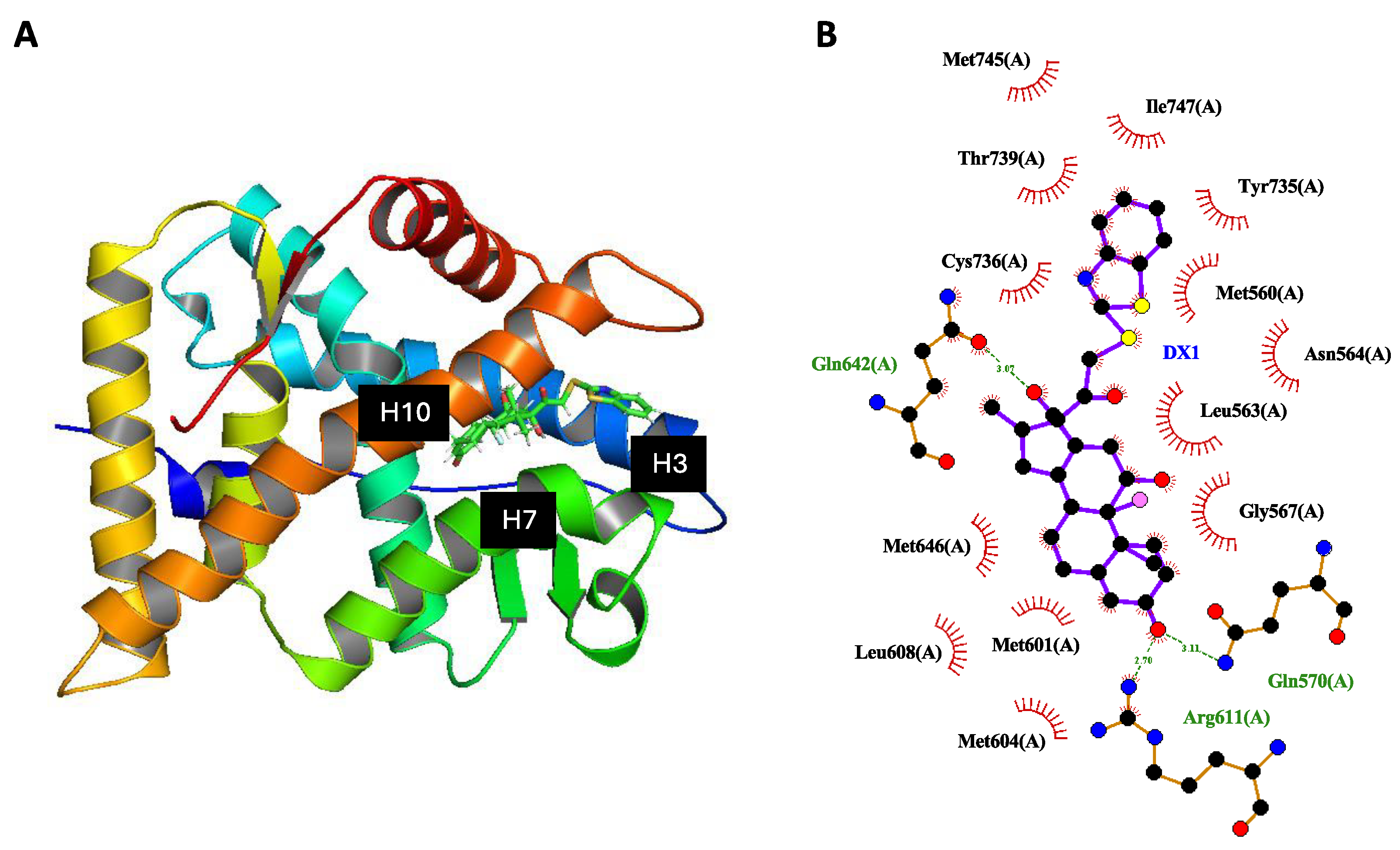
2.6. Weakened Interaction between DX1 and AF-2 Domain of GR
2.7. MBT Structure Selectively Modulates the GR
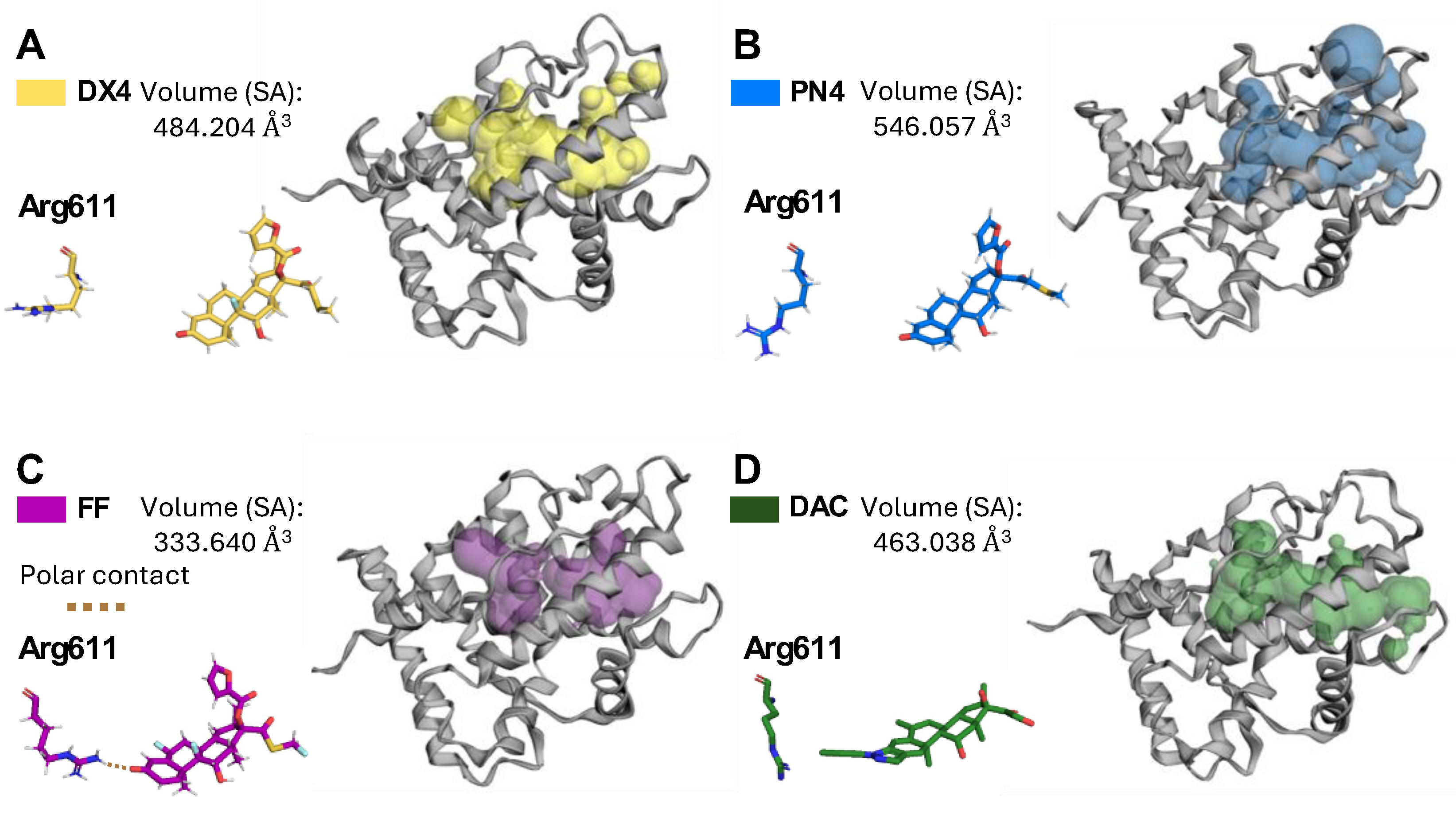
2.8. DX4 and PN4 Expand the Glucocorticoid Receptor Ligand-Binding Pocket
3. Discussion
4. Materials and Methods
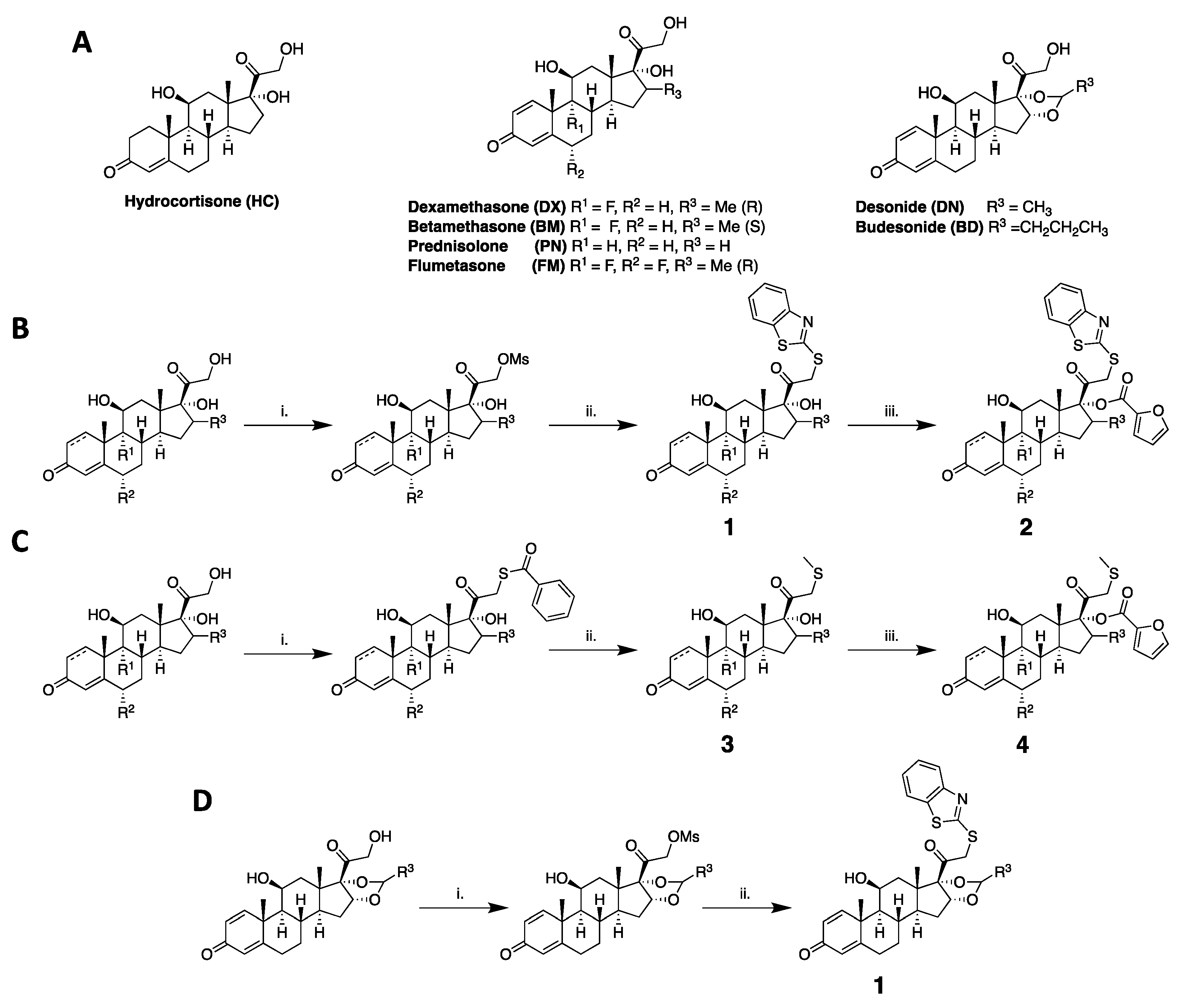
4.1. Synthesis
4.2. Biological Assays
4.3. Molecular Docking and PDB Rational
4.4. Molecular Dynamic Simulations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hwang, J.L.; Weiss, R.E. Steroid-induced diabetes: A clinical and molecular approach to understanding and treatment. Diabetes/Metab. Res. Rev. 2014, 30, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Van Staa, T.P.; Leufkens, H.G.M.; Abenhaim, L.; Begaud, B.; Zhang, B.; Cooper, C. Use of Oral Corticosteroids in the United Kingdom. QJM Mon. J. Assoc. Physicians 2000, 93, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, S.; Cidlowski, J.A. Corticosteroids: Mechanisms of action in health and disease. Rheum. Dis. Clin. 2016, 42, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Schäcke, H.; Döcke, W.-D.; Asadullah, K. Mechanisms involved in the side effects of glucocorticoids. Pharmacol. Ther. 2002, 96, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Overman, R.A.; Yeh, J.Y.; Deal, C.L. Prevalence of oral glucocorticoid usage in the United States: A general population perspective. Arthritis Care Res. 2013, 65, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Onwubalili, J.; Obineche, E. High incidence of post-transplant diabetes mellitus in a single-centre study. Nephrol. Dial. Transplant. 1992, 7, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.; McQueen, A.; Chen, T.-C.; Wang, J.-C. Regulation of glucose homeostasis by glucocorticoids. Glucocorticoid Signal. 2015, 872, 99–126. [Google Scholar]
- Steiner, R.W.; Awdishu, L. Steroids in kidney transplant patients. In Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 157–167. [Google Scholar]
- Lim, H.-W.; Uhlenhaut, N.H.; Rauch, A.; Weiner, J.; Hübner, S.; Hübner, N.; Won, K.-J.; Lazar, M.A.; Tuckermann, J.; Steger, D.J. Genomic redistribution of GR monomers and dimers mediates transcriptional response to exogenous glucocorticoid in vivo. Genome Res. 2015, 25, 836–844. [Google Scholar] [CrossRef]
- Reichardt, H.M.; Tuckermann, J.P.; Göttlicher, M.; Vujic, M.; Weih, F.; Angel, P.; Herrlich, P.; Schütz, G. Repression of inflammatory responses in the absence of DNA binding by the glucocorticoid receptor. EMBO J. 2001, 20, 7168–7173. [Google Scholar] [CrossRef]
- Holden, N.S.; Bell, M.J.; Rider, C.F.; King, E.M.; Gaunt, D.D.; Leigh, R.; Johnson, M.; Siderovski, D.P.; Heximer, S.P.; Giembycz, M.A. β2-Adrenoceptor agonist-induced RGS2 expression is a genomic mechanism of bronchoprotection that is enhanced by glucocorticoids. Proc. Natl. Acad. Sci. USA 2011, 108, 19713–19718. [Google Scholar] [CrossRef]
- Flamini, S.; Sergeev, P.; Viana de Barros, Z.; Mello, T.; Biagioli, M.; Paglialunga, M.; Fiorucci, C.; Prikazchikova, T.; Pagano, S.; Gagliardi, A. Glucocorticoid-induced leucine zipper regulates liver fibrosis by suppressing CCL2-mediated leukocyte recruitment. Cell Death Dis. 2021, 12, 421. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; Goff, M.R.; Updegraff, B.L.; Lu, D.; Brown, P.L.; Minkin Jr, S.C.; Biggerstaff, J.P.; Zhao, L.; Karlstad, M.D.; Collier, J.J. Regulation of the CCL2 gene in pancreatic β-cells by IL-1β and glucocorticoids: Role of MKP-1. PLoS ONE 2012, 7, e46986. [Google Scholar] [CrossRef] [PubMed]
- Burke, S.J.; May, A.L.; Noland, R.C.; Lu, D.; Brissova, M.; Powers, A.C.; Sherrill, E.M.; Karlstad, M.D.; Campagna, S.R.; Stephens, J.M.; et al. Thiobenzothiazole-modified Hydrocortisones Display Anti-inflammatory Activity with Reduced Impact on Islet beta-Cell Function. J. Biol. Chem. 2015, 290, 13401–13416. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Y.; Haroon, M.; Akhtar, T.; Ashfaq, M.; Tahir, M.N.; Rasheed, L.; Yousuf, M.; Zia, M.A. Synthesis, spectroscopic characterization, single crystal XRD, Hirshfeld Surface analysis and theoretical studies (DFT) of 4-adamantyl-(2-(substitutedbenzylidene) hydrazinyl) thiazoles. J. Mol. Struct. 2022, 1267, 133620. [Google Scholar] [CrossRef]
- He, Y.; Shi, J.; Yi, W.; Ren, X.; Gao, X.; Li, J.; Wu, N.; Weaver, K.; Xie, Q.; Khoo, S.K. Discovery of a highly potent glucocorticoid for asthma treatment. Cell Discov. 2015, 1, 15035. [Google Scholar] [CrossRef] [PubMed]
- Roma, L.P.; Oliveira, C.A.; Carneiro, E.M.; Albuquerque, G.G.; Boschero, A.C.; Souza, K.L. N-acetylcysteine protects pancreatic islet against glucocorticoid toxicity. Redox Rep. 2011, 16, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Smoak, P.; Burke, S.J.; Martin, T.M.; Batdorf, H.M.; Floyd, Z.E.; Collier, J.J. Artemisia dracunculus L. Ethanolic Extract and an Isolated Component, DMC2, Ameliorate Inflammatory Signaling in Pancreatic β-Cells via Inhibition of p38 MAPK. Biomolecules 2022, 12, 708. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.; Berchtold, S.; Ranta, F.; Seebohm, G.; Henke, G.; Lupescu, A.; Mack, A.F.; Chao, C.-M.; Su, J.; Nitschke, R. Serum-and glucocorticoid-inducible kinase 1 (SGK1) mediates glucocorticoid-induced inhibition of insulin secretion. Diabetes 2005, 54, 1090–1099. [Google Scholar] [CrossRef]
- Burke, S.J.; Karlstad, M.D.; Regal, K.M.; Sparer, T.E.; Lu, D.; Elks, C.M.; Grant, R.W.; Stephens, J.M.; Burk, D.H.; Collier, J.J. CCL20 is elevated during obesity and differentially regulated by NF-κB subunits in pancreatic β-cells. Biochim. Et Biophys. Acta (BBA)-Gene Regul. Mech. 2015, 1849, 637–652. [Google Scholar] [CrossRef]
- Mishra, V.; Yadav, N.; Saraogi, G.K.; Tambuwala, M.M.; Giri, N. Dendrimer based nanoarchitectures in diabetes management: An overview. Curr. Pharm. Des. 2019, 25, 2569–2583. [Google Scholar] [CrossRef]
- Cardozo, A.K.; Proost, P.; Gysemans, C.; Chen, M.-C.; Mathieu, C.; Eizirik, D.L. IL-1β and IFN-γ induce the expression of diverse chemokines and IL-15 in human and rat pancreatic islet cells, and in islets from pre-diabetic NOD mice. Diabetologia 2003, 46, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Stojceski, F.; Buetti-Dinh, A.; Stoddart, M.J.; Danani, A.; Della Bella, E.; Grasso, G. Influence of dexamethasone on the interaction between glucocorticoid receptor and SOX9: A molecular dynamics study. J. Mol. Graph. Model. 2023, 125, 108587. [Google Scholar] [CrossRef]
- Bridgham, J.T.; Ortlund, E.A.; Thornton, J.W. An epistatic ratchet constrains the direction of glucocorticoid receptor evolution. Nature 2009, 461, 515–519. [Google Scholar] [CrossRef]
- Veleiro, A.S.; Alvarez, L.D.; Eduardo, S.L.; Burton, G. Structure of the glucocorticoid receptor, a flexible protein that can adapt to different ligands. ChemMedChem 2010, 5, 649–659. [Google Scholar] [CrossRef]
- Frank, F.; Ortlund, E.A.; Liu, X. Structural insights into glucocorticoid receptor function. Biochem. Soc. Trans. 2021, 49, 2333–2343. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Gutierrez, J.S.; Damsker, J.M.; Nagaraju, K.; Hoffman, E.P.; Ortlund, E.A. Disruption of a key ligand-H-bond network drives dissociative properties in vamorolone for Duchenne muscular dystrophy treatment. Proc. Natl. Acad. Sci. USA 2020, 117, 24285–24293. [Google Scholar] [CrossRef]
- Suino-Powell, K.; Xu, Y.; Zhang, C.; Tao, Y.-g.; Tolbert, W.D.; Simons, S.S., Jr.; Xu, H.E. Doubling the size of the glucocorticoid receptor ligand binding pocket by deacylcortivazol. Mol. Cell. Biol. 2008, 28, 1915–1923. [Google Scholar] [CrossRef]
- Vandevyver, S.; Dejager, L.; Tuckermann, J.; Libert, C. New insights into the anti-inflammatory mechanisms of glucocorticoids: An emerging role for glucocorticoid-receptor-mediated transactivation. Endocrinology 2013, 154, 993–1007. [Google Scholar] [CrossRef]
- Biju, P.; McCormick, K.; Aslanian, R.; Berlin, M.; Solomon, D.; Chapman, R.; McLeod, R.; Prelusky, D.; Eckel, S.; Kelly, G. Steroidal C-21 mercapto derivatives as dissociated steroids: Discovery of an inhaled dissociated steroid. Bioorganic Med. Chem. Lett. 2011, 21, 6343–6347. [Google Scholar] [CrossRef]
- Wuest, F.; Carlson, K.E.; Katzenellenbogen, J.A. Expeditious synthesis of steroids containing a 2-methylsulfanyl-acetyl side chain as potential glucocorticoid receptor imaging agents. Steroids 2008, 73, 69–76. [Google Scholar] [CrossRef]
- Hohmeier, H.E.; Mulder, H.; Chen, G.; Henkel-Rieger, R.; Prentki, M.; Newgard, C.B. Isolation of INS-1-derived cell lines with robust ATP-sensitive K+ channel-dependent and-independent glucose-stimulated insulin secretion. Diabetes 2000, 49, 424–430. [Google Scholar] [CrossRef]
- Collier, J.J.; Batdorf, H.M.; Merrifield, K.L.; Martin, T.M.; White, U.; Ravussin, E.; Burk, D.H.; Cooley, C.R.; Karlstad, M.D.; Burke, S.J. Pioglitazone Reverses Markers of Islet Beta-Cell De-Differentiation in db/db Mice While Modulating Expression of Genes Controlling Inflammation and Browning in White Adipose Tissue from Insulin-Resistant Mice and Humans. Biomedicines 2021, 9, 1189. [Google Scholar] [CrossRef] [PubMed]
- Postel, S.; Wissler, L.; Johansson, C.A.; Gunnarsson, A.; Gordon, E.; Collins, B.; Castaldo, M.; Köhler, C.; Öling, D.; Johansson, P. Quaternary glucocorticoid receptor structure highlights allosteric interdomain communication. Nat. Struct. Mol. Biol. 2023, 30, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, L. The PyMOL Molecular Graphics System, Version 2.0; Schrödinger LLC.: New York, NY, USA, 2017. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand–protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Yu, Y.; Kramer, A.; Venable, R.M.; Simmonett, A.C.; MacKerell Jr, A.D.; Klauda, J.B.; Pastor, R.W.; Brooks, B.R. Semi-automated optimization of the CHARMM36 lipid force field to include explicit treatment of long-range dispersion. J. Chem. Theory Comput. 2021, 17, 1562–1580. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Abraham, M.; Alekseenko, A.; Bergh, C.; Blau, C.; Briand, E.; Doijade, M.; Fleischmann, S.; Gapsys, V.; Garg, G.; Gorelov, S. Gromacs 2023.2 Manual; Zenodo, CERN European Organization for Nuclear Research: Geneve, Switzerland, 2023. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Biggadike, K.; Bledsoe, R.K.; Hassell, A.M.; Kirk, B.E.; McLay, I.M.; Shewchuk, L.M.; Stewart, E.L. X-ray crystal structure of the novel enhanced-affinity glucocorticoid agonist fluticasone furoate in the glucocorticoid receptor−ligand binding domain. J. Med. Chem. 2008, 51, 3349–3352. [Google Scholar] [CrossRef]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed atlas of surface topography of proteins. Nucleic Acids Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compd | CCL2 | 3xGRE | ||||
|---|---|---|---|---|---|---|
| pIC50 | Emax | b %DX | pEC50 | Emax | b %DX | |
| DX | 8.3 | 88.6 | 100.0 | 6.8 | 7.9 | 100.0 |
| a HC1 | 6.9 | 77.0 | 94.0 | 5.3 | 1.8 | 24.6 |
| a HC2 | 6.2 | 72.8 | 82.2 | 7.4 | 8.2 | 104.8 |
| HC3 | NDR | 37.1 | 41.8 | NDR | 1.4 | 17.2 |
| HC4 | 7.1 | 78.6 | 88.7 | AAC | 3.6 | 45.9 |
| DX1 | 7.2 | 80.0 | 90.3 | NDR | 1.0 | 12.6 |
| DX2 | 8.3 | 89.7 | 101.3 | 6.7 | 4.9 | 62.2 |
| DX3 | AAC | 75.1 | 84.7 | NDR | 7.4 | 94.4 |
| DX4 | AAC | 84.7 | 95.6 | AAC | 8.9 | 113.0 |
| BM | 8.3 | 88.7 | 100.1 | 9.8 | 10.3 | 131.3 |
| BM1 | 7.2 | 85.7 | 96.7 | 7.2 | 2.2 | 28.0 |
| BM2 | 7.8 | 83.3 | 94.0 | 7.8 | 4.7 | 59.9 |
| BM3 | 8.9 | 83.5 | 94.3 | 7.3 | 4.8 | 60.6 |
| BM4 | 7.2 | 81.6 | 92.1 | 7.5 | 4.9 | 62.0 |
| PN | 8.4 | 85.1 | 96.1 | 9.4 | 10.0 | 127.2 |
| PN1 | 5.9 | 75.0 | 84.7 | 6.1 | 2.4 | 30.5 |
| PN2 | AAC | 90.9 | 102.6 | 8.8 | 7.5 | 95.9 |
| PN3 | 7.1 | 49.6 | 56.0 | 6.6 | 3.4 | 43.3 |
| PN4 | AAC | 80.8 | 91.2 | AAC | 5.4 | 69.2 |
| FM | AAC | 90.4 | 102.1 | AAC | 10.2 | 129.5 |
| FM1 | 7.5 | 85.9 | 97.0 | 8.8 | 4.8 | 60.4 |
| FM2 | AAC | 88.3 | 99.6 | 7.9 | 11.7 | 149.1 |
| FM3 | AAC | 82.2 | 92.8 | AAC | 5.6 | 71.6 |
| FM4 | AAC | 75.9 | 85.7 | AAC | 4.9 | 62.6 |
| DN | 8.8 | 83.2 | 93.9 | AAC | 9.6 | 121.7 |
| DN1 | 7.9 | 65.6 | 74.1 | 7.0 | 2.5 | 31.8 |
| BD | AAC | 83.4 | 94.1 | AAC | 7.0 | 88.7 |
| BD1 | 6.6 | 57.6 | 65.0 | 6.0 | 2.0 | 25.3 |
 | ||||||
|---|---|---|---|---|---|---|
| Compd | CCL2 | 3xGRE | ||||
| pIC50 | Emax | a %DX | pEC50 | Emax | a %DX | |
| DX | 8.3 | 88.6 | 100.0 | 6.8 | 7.9 | 100.0 |
| DX1 | 7.2 | 80.0 | 90.3 | NDR | 1.0 | 12.6 |
| DX5 | 6.2 | 67.2 | 75.8 | 5.8 | 3.2 | 40.5 |
| DX6 | 7.1 | 65.0 | 73.4 | 7.5 | 4.9 | 61.7 |
| DX7 | 7.1 | 66.7 | 75.3 | 7.7 | 6.5 | 82.4 |
| DX8 | 6.3 | 54.0 | 60.9 | 6.9 | 2.3 | 29.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seaton, W.B.; Burke, S.J.; Fisch, A.R.; Schilletter, W.A.; Beck, M.G.A.; Cassagne, G.A.; Harvey, I.; Fontenot, M.S.; Collier, J.J.; Campagna, S.R. Channel Expansion in the Ligand-Binding Domain of the Glucocorticoid Receptor Contributes to the Activity of Highly Potent Glucocorticoid Analogues. Molecules 2024, 29, 1546. https://doi.org/10.3390/molecules29071546
Seaton WB, Burke SJ, Fisch AR, Schilletter WA, Beck MGA, Cassagne GA, Harvey I, Fontenot MS, Collier JJ, Campagna SR. Channel Expansion in the Ligand-Binding Domain of the Glucocorticoid Receptor Contributes to the Activity of Highly Potent Glucocorticoid Analogues. Molecules. 2024; 29(7):1546. https://doi.org/10.3390/molecules29071546
Chicago/Turabian StyleSeaton, Wesley B., Susan J. Burke, Alexander R. Fisch, William A. Schilletter, Mary Grace A. Beck, Gabrielle A. Cassagne, Innocence Harvey, Molly S. Fontenot, J. Jason Collier, and Shawn R. Campagna. 2024. "Channel Expansion in the Ligand-Binding Domain of the Glucocorticoid Receptor Contributes to the Activity of Highly Potent Glucocorticoid Analogues" Molecules 29, no. 7: 1546. https://doi.org/10.3390/molecules29071546
APA StyleSeaton, W. B., Burke, S. J., Fisch, A. R., Schilletter, W. A., Beck, M. G. A., Cassagne, G. A., Harvey, I., Fontenot, M. S., Collier, J. J., & Campagna, S. R. (2024). Channel Expansion in the Ligand-Binding Domain of the Glucocorticoid Receptor Contributes to the Activity of Highly Potent Glucocorticoid Analogues. Molecules, 29(7), 1546. https://doi.org/10.3390/molecules29071546