
Isolation, Purification and Tyrosinase Inhibitory Activity of Anthocyanins and Their Novel Degradation Compounds from Solanum tuberosum L.


Abstract
1. Introduction
2. Results and Discussion
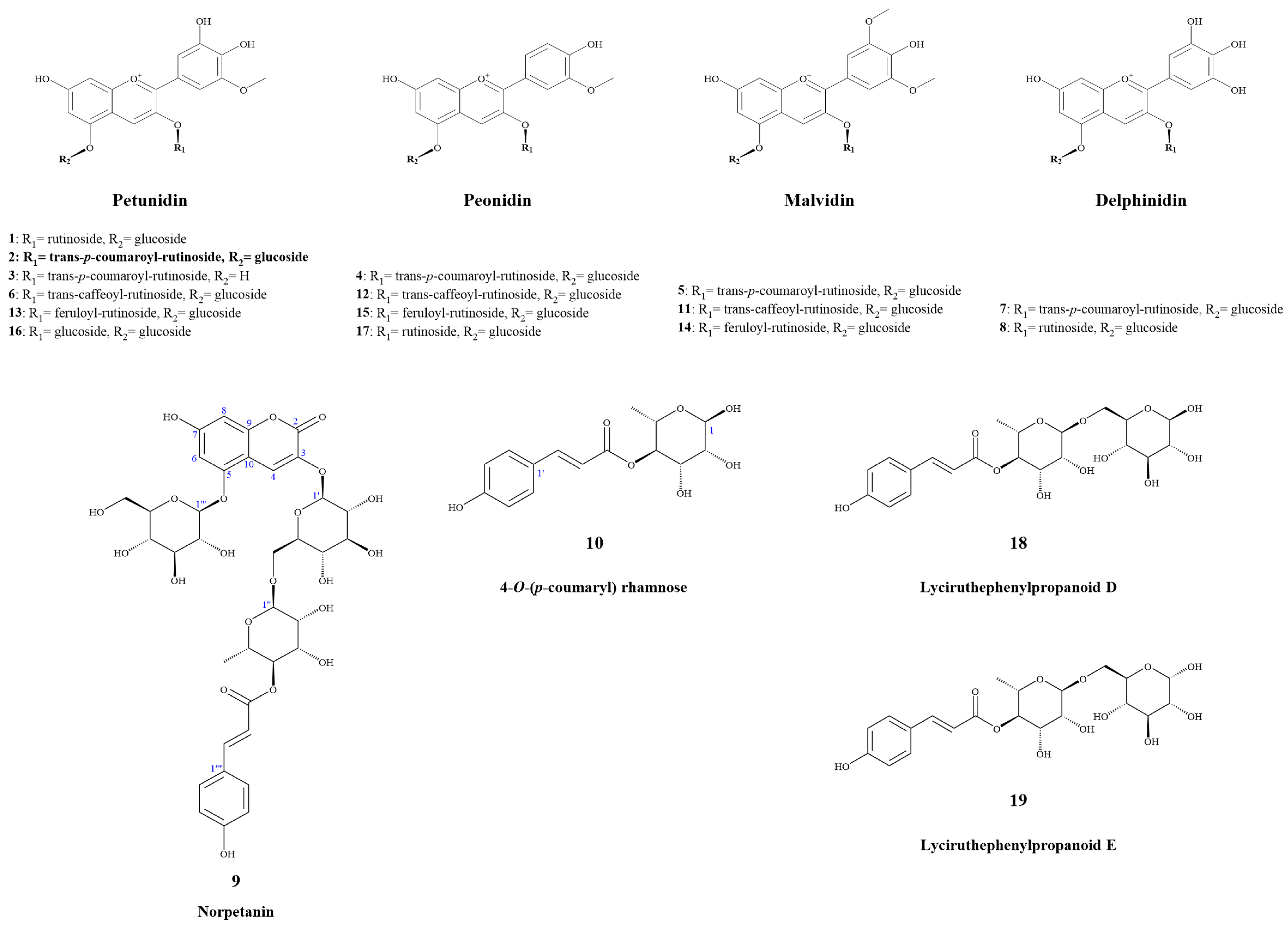
2.1. Compounds Structure Identification
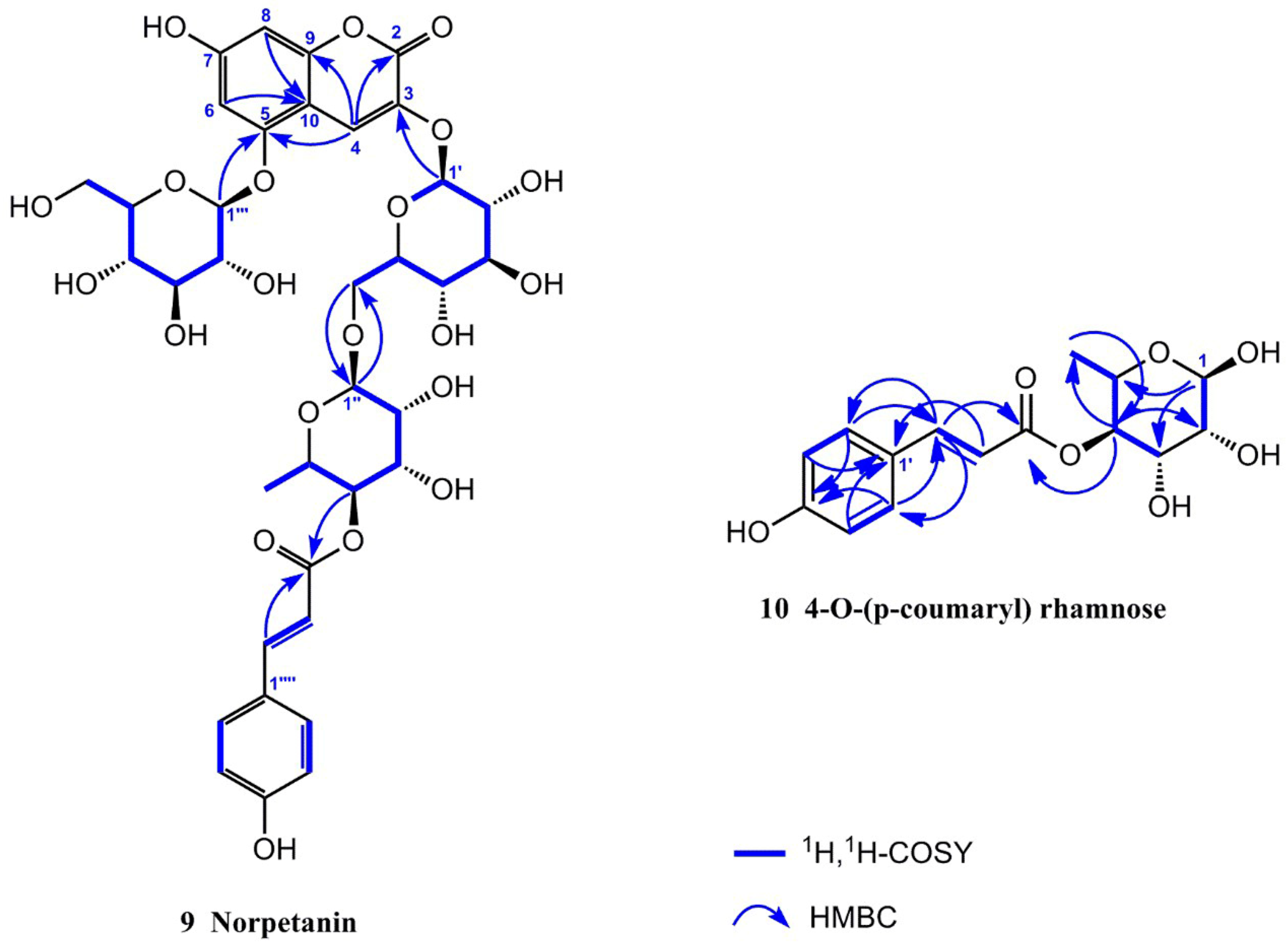
2.1.1. Resolution and Identification of New Compounds
2.1.2. First Report of NMR Data for Known Compounds
2.1.3. Identification of Known Compounds
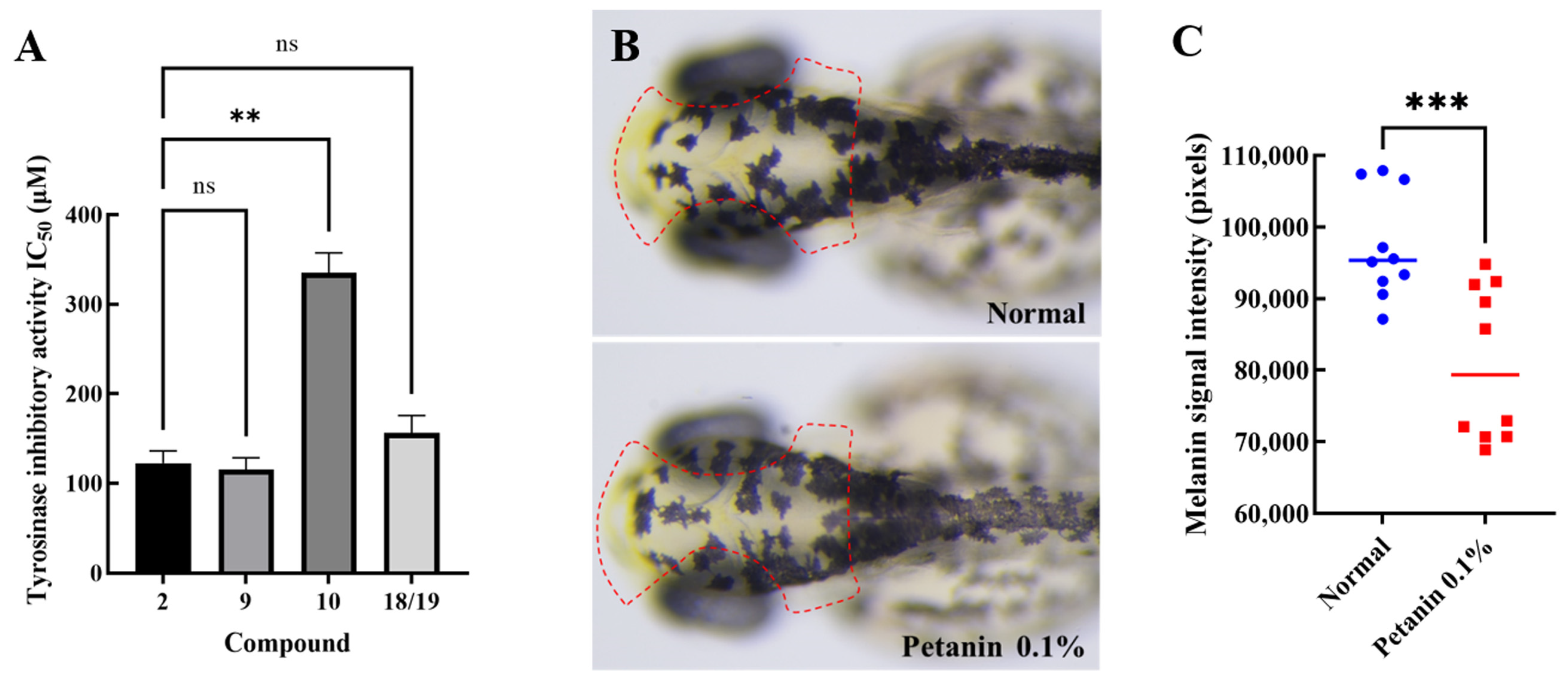
2.2. Screening and Antimelanogenesis of Tyrosinase Inhibitors
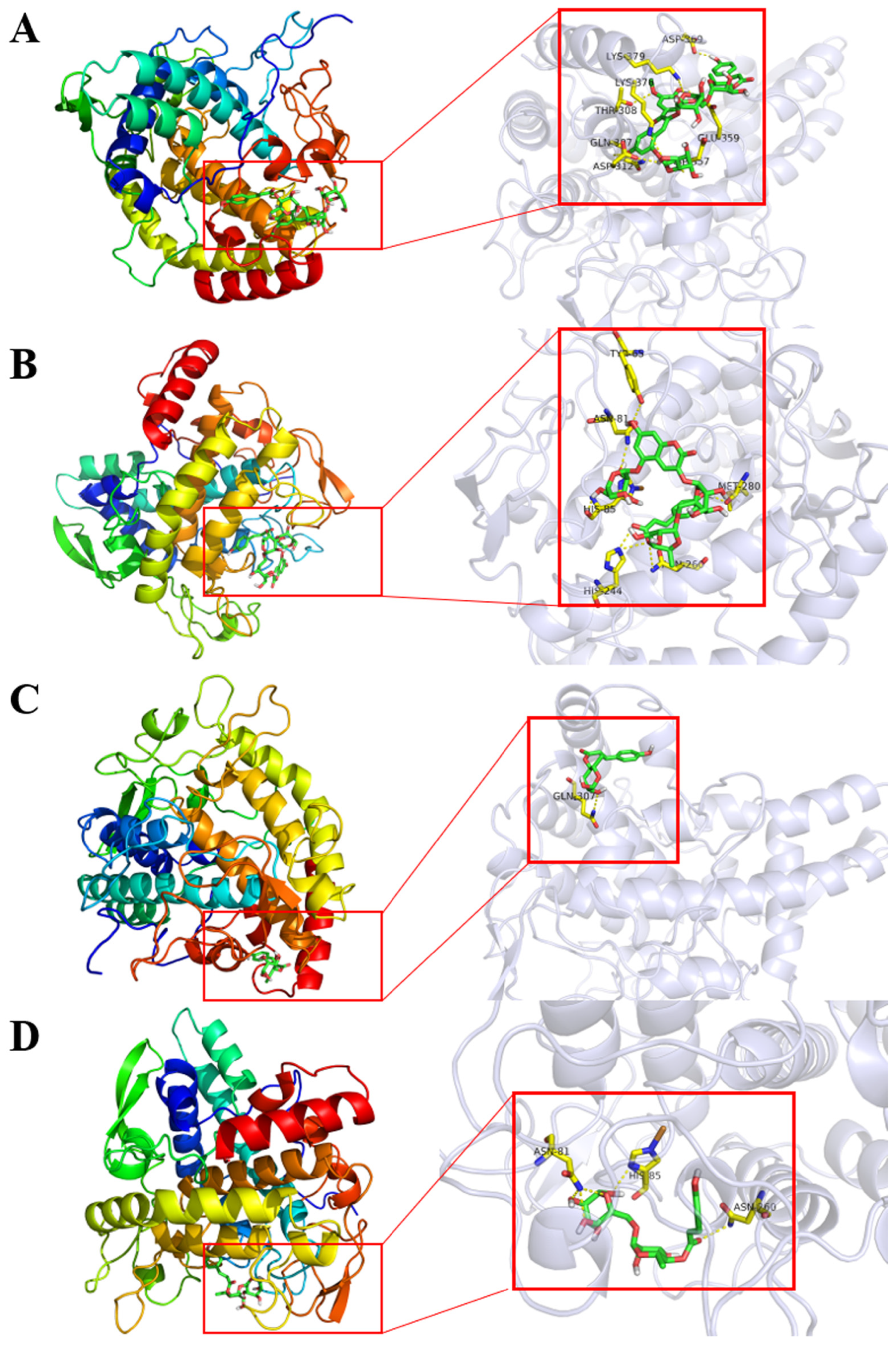
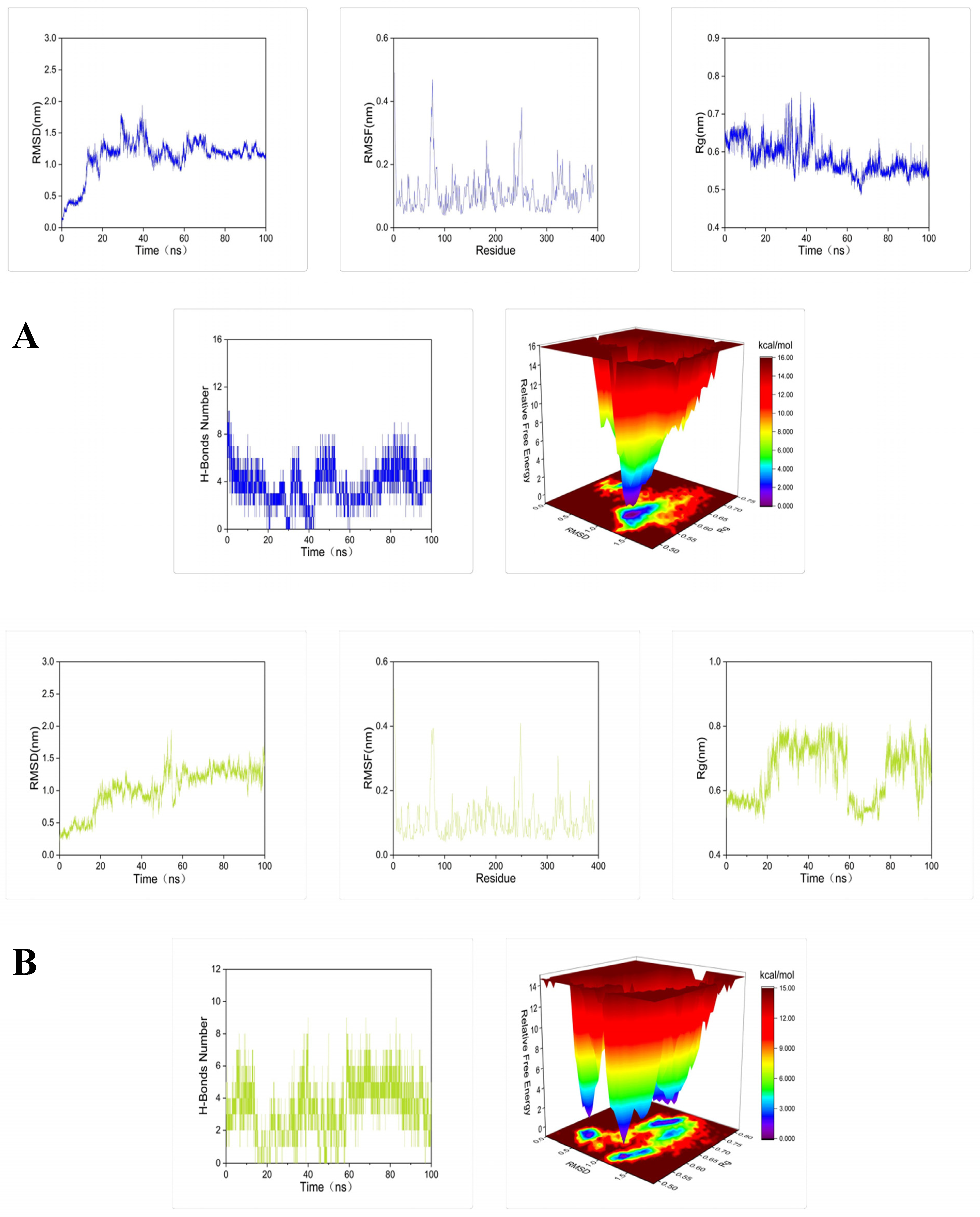
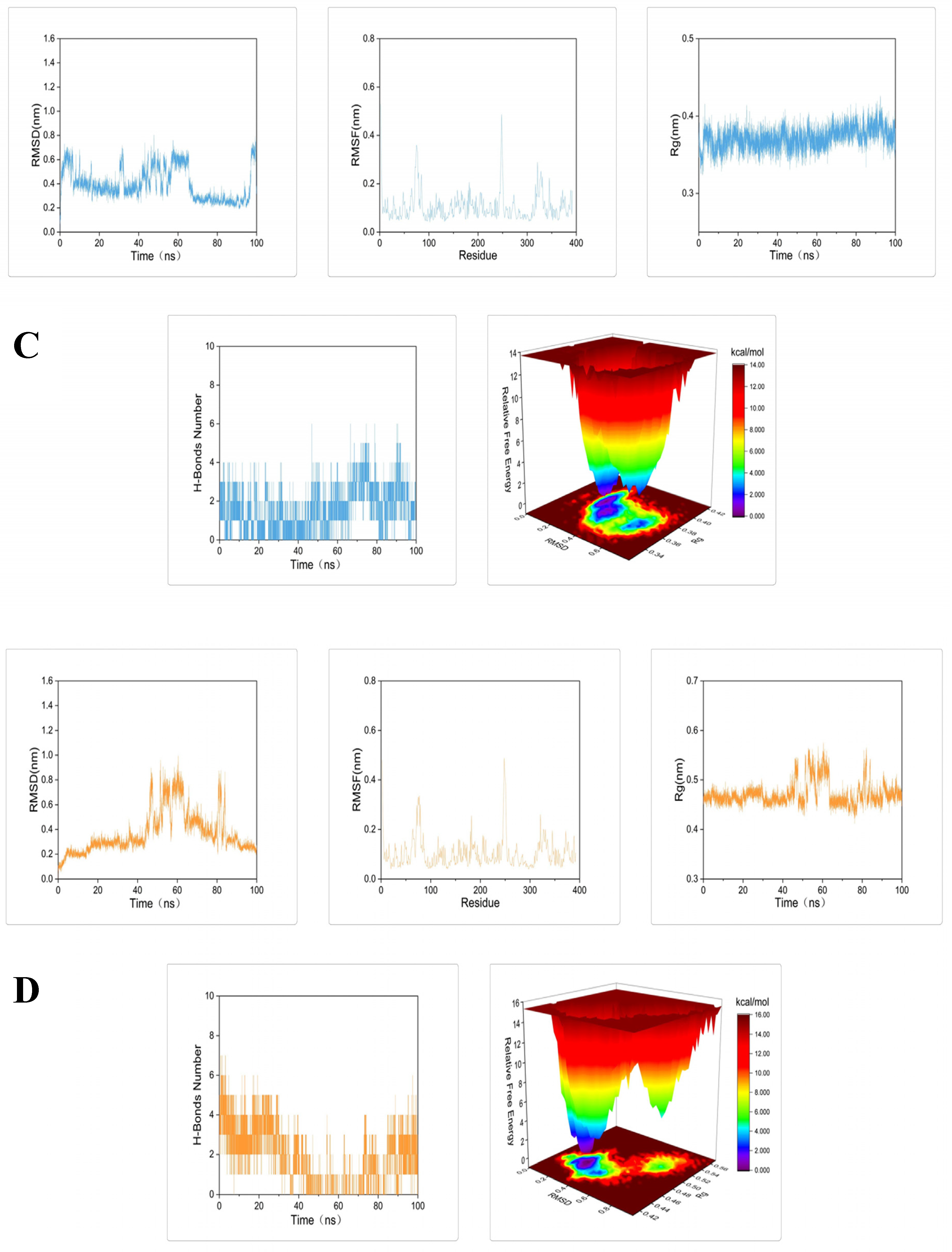
2.3. Molecular Docking and Dynamic Simulation of Tyrosinase Inhibitors
3. Materials and Methods
3.1. Plant Materials
3.2. General Experimental Procedures
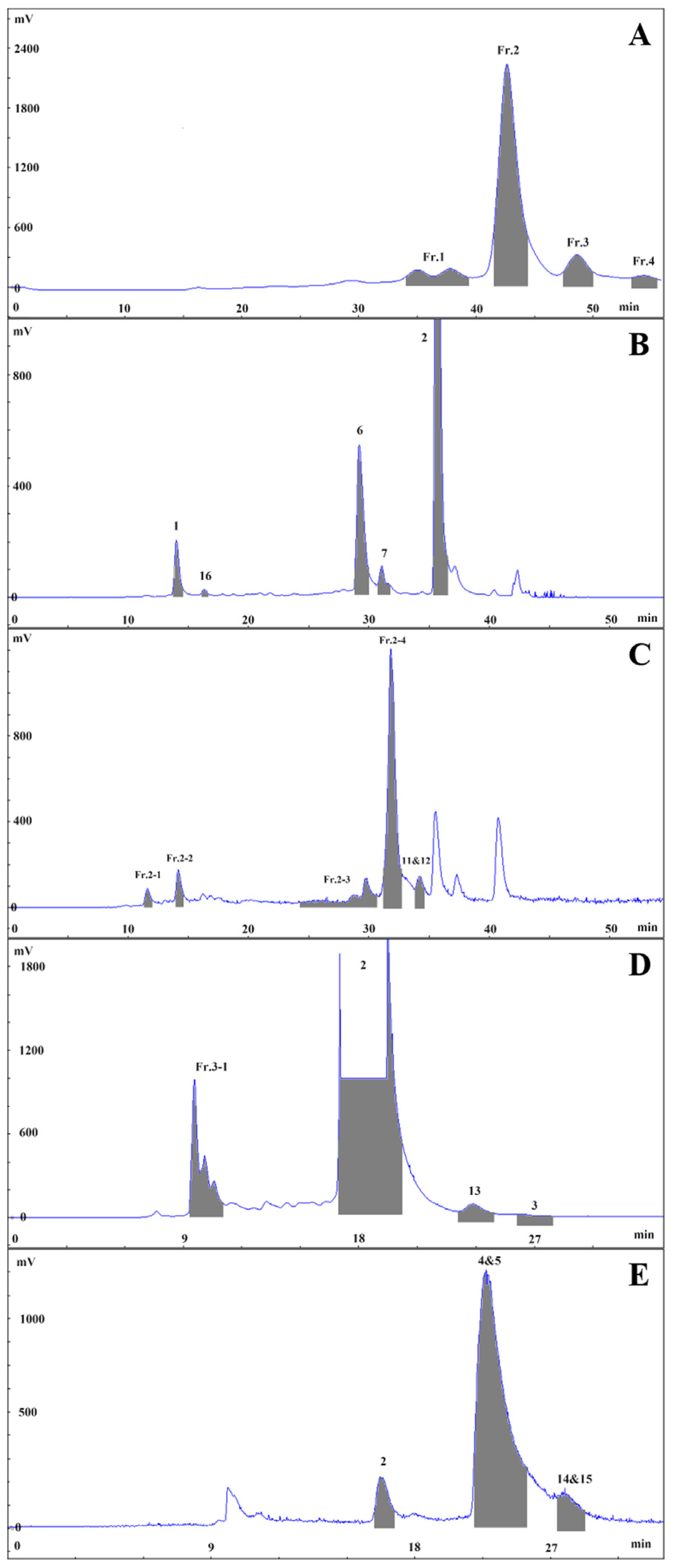
3.3. Extraction and Isolation
3.3.1. Isolation and Purification of Fr.3
3.3.2. Isolation and Purification of Fr.4
3.3.3. Isolation and Purification of Fr.1
3.3.4. Isolation and Purification of Fr.2
3.4. Screening and Validation of Inhibitory Tyrosinase Activity
3.4.1. Screening for Tyrosinase Inhibitory Activity
3.4.2. Safety Evaluation and Verification of Anti-Melanogenic Effect
3.5. Molecular Docking and Dynamics Simulation
3.5.1. Molecular Docking
3.5.2. Dynamics Simulation
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutaker, R.M.; Weiß, C.L.; Ellis, D.; Anglin, N.L.; Knapp, S.; Luis Fernández-Alonso, J.; Prat, S.; Burbano, H.A. The origins and adaptation of European potatoes reconstructed from historical genomes. Nat. Ecol. Evol. 2019, 3, 1093–1101. [Google Scholar] [CrossRef]
- Liu, B.; Gu, W.; Yang, Y.; Lu, B.; Wang, F.; Zhang, B.; Bi, J. Promoting potato as staple food can reduce the carbon–land–water impacts of crops in China. Nat. Food 2021, 2, 570–577. [Google Scholar] [CrossRef]
- Stewart, L.; Indukuri, V.V.; Charepalli, V.; Chrisfield, B.J.; Anantheswaran, R.C.; Lambert, J.D.; Vanamala, J.K. Comparative effects of vacuum or conventional frying on the polyphenol chemistry and in vitro colon cancer stem cell inhibitory activity of purple-flesh potatoes. J. Food Sci. 2022, 87, 3260–3267. [Google Scholar] [CrossRef]
- Mori, M.; Hayashi, K.; Ohara-Takada, A.; Watanuki, H.; Katahira, R.; Ono, H.; Terahara, N. Anthocyanins from Skins and Fleshes of Potato Varieties. Food Sci. Technol. Res. 2010, 16, 115–122. [Google Scholar] [CrossRef]
- Oertel, A.; Matros, A.; Hartmann, A.; Arapitsas, P.; Dehmer, K.J.; Martens, S.; Mock, H.-P. Metabolite profiling of red and blue potatoes revealed cultivar and tissue specific patterns for anthocyanins and other polyphenols. Planta 2017, 246, 281–297. [Google Scholar] [CrossRef]
- Yin, L.; Chen, T.; Li, Y.; Fu, S.; Li, L.; Xu, M.; Niu, Y. A Comparative Study on Total Anthocyanin Content, Composition of Anthocyanidin, Total Phenolic Content and Antioxidant Activity of Pigmented Potato Peel and Flesh. Food Sci. Technol. Res. 2016, 22, 219–226. [Google Scholar] [CrossRef]
- Eichhorn, S.; Winterhalter, P. Anthocyanins from pigmented potato (Solanum tuberosum L.) varieties. Food Res. Int. 2005, 38, 943–948. [Google Scholar] [CrossRef]
- Lewis, C.E.; Walker, J.R.; Lancaster, J.E.; Sutton, K.H. Determination of anthocyanins, flavonoids and phenolic acids in potatoes. I: Coloured cultivars of Solanum tuberosum L. J. Sci. Food Agric. 1998, 77, 45–57. [Google Scholar] [CrossRef]
- Fossen, T.; Øvstedal, D.O.; Slimestad, R.; Andersen, Ø.M. Anthocyanins from a Norwegian potato cultivar. Food Chem. 2003, 81, 433–437. [Google Scholar] [CrossRef]
- Sampaio, S.L.; Petropoulos, S.A.; Dias, M.I.; Pereira, C.; Calhelha, R.C.; Fernandes, Â.; Leme, C.M.; Alexopoulos, A.; Santos-Buelga, C.; Ferreira, I.C. Phenolic composition and cell-based biological activities of ten coloured potato peels (Solanum tuberosum L.). Food Chem. 2021, 363, 130360. [Google Scholar] [CrossRef]
- Sampaio, S.L.; Lonchamp, J.; Dias, M.I.; Liddle, C.; Petropoulos, S.A.; Glamočlija, J.; Alexopoulos, A.; Santos-Buelga, C.; Ferreira, I.C.; Barros, L. Anthocyanin-rich extracts from purple and red potatoes as natural colourants: Bioactive properties, application in a soft drink formulation and sensory analysis. Food Chem. 2021, 342, 128526. [Google Scholar] [CrossRef]
- Qi, Z.; Wang, W.; Liu, Z.; Niu, N.; Li, Z.; Chen, L.; Zhu, J.; Li, D.; Liu, Y. Anthocyanin Profiles in Colored Potato Tubers at Different Altitudes by HPLC–MS Analysis with Optimized Ultrasound-Assisted Extraction. Foods 2023, 12, 4175. [Google Scholar] [CrossRef]
- Maeda-Yamamoto, M.; Honmou, O.; Sasaki, M.; Haseda, A.; Kagami-Katsuyama, H.; Shoji, T.; Namioka, A.; Namioka, T.; Magota, H.; Oka, S. The impact of purple-flesh Potato (Solanum tuberosum L.) cv.“Shadow Queen” on minor health complaints in healthy adults: A randomized, double-blind, placebo-controlled study. Nutrients 2022, 14, 2446. [Google Scholar] [CrossRef] [PubMed]
- Strugała, P.; Urbaniak, A.; Kuryś, P.; Włoch, A.; Kral, T.; Ugorski, M.; Hof, M.; Gabrielska, J. Antitumor and antioxidant activities of purple potato ethanolic extract and its interaction with liposomes, albumin and plasmid DNA. Food Funct. 2021, 12, 1271–1290. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.; Blesso, C.N. Antioxidant properties of anthocyanins and their mechanism of action in atherosclerosis. Free Radic. Biol. Med. 2021, 172, 152–166. [Google Scholar] [CrossRef]
- Li, S.; Wang, T.; Wu, B.; Fu, W.; Xu, B.; Pamuru, R.R.; Kennett, M.; Vanamala, J.K.; Reddivari, L. Anthocyanin-containing purple potatoes ameliorate DSS-induced colitis in mice. J. Nutr. Biochem. 2021, 93, 108616. [Google Scholar] [CrossRef]
- Jokioja, J.; Yang, B.; Linderborg, K.M. Acylated anthocyanins: A review on their bioavailability and effects on postprandial carbohydrate metabolism and inflammation. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5570–5615. [Google Scholar] [CrossRef]
- Lee, E.H.; Hong, S.-C.; Kim, S.M.; Pan, C.-H. Inhibitory effect of anthocyanin-enriched fractions from colored potatoes against lipase and α-glucosidase. LWT 2023, 184, 114952. [Google Scholar] [CrossRef]
- Chen, K.; Kortesniemi, M.K.; Linderborg, K.M.; Yang, B. Anthocyanins as promising molecules affecting energy homeostasis, inflammation, and gut microbiota in type 2 diabetes with special reference to impact of acylation. J. Agric. Food Chem. 2022, 71, 1002–1017. [Google Scholar] [CrossRef]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Zhang, N.; Jing, P. Anthocyanins in Brassicaceae: Composition, stability, bioavailability, and potential health benefits. Crit. Rev. Food Sci. Nutr. 2022, 62, 2205–2220. [Google Scholar] [CrossRef]
- Qi, Q.; Chu, M.; Yu, X.; Xie, Y.; Li, Y.; Du, Y.; Liu, X.; Zhang, Z.; Shi, J.; Yan, N. Anthocyanins and proanthocyanidins: Chemical structures, food sources, bioactivities, and product development. Food Rev. Int. 2023, 39, 4581–4609. [Google Scholar] [CrossRef]
- Şöhretoğlu, D.; Sari, S.; Barut, B.; Özel, A. Tyrosinase inhibition by some flavonoids: Inhibitory activity, mechanism by in vitro and in silico studies. Bioorganic Chem. 2018, 81, 168–174. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Jhan, J.K.; Chung, Y.C.; Chen, G.H.; Chang, C.H.; Lu, Y.C.; Hsu, C.K. Anthocyanin contents in the seed coat of black soya bean and their anti-human tyrosinase activity and antioxidative activity. Int. J. Cosmet. Sci. 2016, 38, 319–324. [Google Scholar] [CrossRef]
- Jiang, Y.; Ding, Y.; Wang, D.; Deng, Y.; Zhao, Y. Radio frequency-assisted enzymatic extraction of anthocyanins from Akebia trifoliata (Thunb.) Koidz. flowers: Process optimization, structure, and bioactivity determination. Ind. Crops Prod. 2020, 149, 112327. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, L.; Zhang, R.; Yang, X.; Sun, Y.; Shi, L.; Xue, P. Optimization of ultrasound-assisted extraction by response surface methodology, antioxidant capacity, and tyrosinase inhibitory activity of anthocyanins from red rice bran. Food Sci. Nutr. 2020, 8, 921–932. [Google Scholar] [CrossRef]
- Shen, M.; Liu, K.; Liang, Y.; Liu, G.; Sang, J.; Li, C. Extraction optimization and purification of anthocyanins from Lycium ruthenicum Murr. and evaluation of tyrosinase inhibitory activity of the anthocyanins. J. Food Sci. 2020, 85, 696–706. [Google Scholar] [CrossRef]
- Yang, S.Y.; Kim, J.H.; Su, X.; Kim, J.A. The Luteolinidin and Petunidin 3-O-Glucoside: A Competitive Inhibitor of Tyrosinase. Molecules 2022, 27, 5703. [Google Scholar] [CrossRef]
- Karunarathne, W.A.H.M.; Molagoda, I.M.N.; Park, S.R.; Kim, J.W.; Lee, O.-K.; Kwon, H.Y.; Oren, M.; Choi, Y.H.; Ryu, H.W.; Oh, S.-R. Anthocyanins from Hibiscus syriacus L. inhibit melanogenesis by activating the ERK signaling pathway. Biomolecules 2019, 9, 645. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Ø.M.; Opheim, S.; Aksnes, D.W.; Frøystein, N.Å. Structure of petanin, an acylated anthocyanin isolated from Solanum tuberosum, using homo-and hetero-nuclear two-dimensional nuclear magnetic resonance techniques. Phytochem. Anal. 1991, 2, 230–236. [Google Scholar] [CrossRef]
- Bontempo, P.; Carafa, V.; Grassi, R.; Basile, A.; Tenore, G.C.; Formisano, C.; Rigano, D.; Altucci, L. Antioxidant, antimicrobial and anti-proliferative activities of Solanum tuberosum L. var. Vitelotte. Food Chem. Toxicol. 2013, 55, 304–312. [Google Scholar] [CrossRef]
- Ando, T.; Tatsuzawa, F.; Toya, Y.; Watanabe, H.; Hirayama, Y.; Shinoda, K.; Hara, R.; Seki, H. Acylated peonidin 3-rutinoside-5-glucosides from commercial petunia cultivars with pink flowers. Heterocycles 2004, 63, 509. [Google Scholar] [CrossRef]
- Fossen, T.; Andersen, Ø.M. Anthocyanins from tubers and shoots of the purple potato, Solanum tuberosum. J. Hortic. Sci. Biotechnol. 2000, 75, 360–363. [Google Scholar] [CrossRef]
- Ando, T.; Saito, N.; Tatsuzawa, F.; Kakefuda, T.; Yamakage, K.; Ohtani, E.; Koshi-ishi, M.; Matsusake, Y.; Kokubun, H.; Watanabe, H. Floral anthocyanins in wild taxa of Petunia (Solanaceae). Biochem. Syst. Ecol. 1999, 27, 623–650. [Google Scholar] [CrossRef]
- Ichiyanagi, T.; Kashiwada, Y.; Shida, Y.; Ikeshiro, Y.; Kaneyuki, T.; Konishi, T. Nasunin from Eggplant Consists of Cis–Trans Isomers of Delphinidin 3-[4-(p-Coumaroyl)-l-rhamnosyl (1→6) glucopyranoside]-5-glucopyranoside. J. Agric. Food Chem. 2005, 53, 9472–9477. [Google Scholar] [CrossRef]
- Slimestad, R.; Aaberg, A.; Andersen, O.M. Acylated anthocyanins from petunia flowers. Phytochemistry 1999, 50, 1081–1086. [Google Scholar] [CrossRef]
- Yoshida, K.; Sato, Y.; Okuno, R.; Kameda, K.; Isobe, M.; Kondo, T. Structural analysis and measurement of anthocyanins from colored seed coats of Vigna, Phaseolus, and Glycine legumes. Biosci. Biotechnol. Biochem. 1996, 60, 589–593. [Google Scholar] [CrossRef]
- Zhao, S.-S.; Li, S.; Luo, Z.-H.; Zhou, Z.-Q.; Li, N.; Wang, Y.; Yao, X.-S.; Gao, H. Bioactive phenylpropanoid derivatives from the fruits of Lycium ruthenicum Murr. Bioorganic Chem. 2021, 116, 105307. [Google Scholar] [CrossRef]
- Yang, Z.; Li, X.; Ma, J.; Han, H.; Liu, S.; Gao, Y. Anthocyanin Composition and Content in “Heijingang” Purple Potato. Food Sci. 2013, 34, 271–275. [Google Scholar]
- Patil, R.; Das, S.; Stanley, A.; Yadav, L.; Sudhakar, A.; Varma, A.K. Optimized hydrophobic interactions and hydrogen bonding at the target-ligand interface leads the pathways of drug-designing. PLoS ONE 2010, 5, e12029. [Google Scholar] [CrossRef] [PubMed]
- Brüschweiler, R. Efficient RMSD measures for the comparison of two molecular ensembles. Proteins Struct. Funct. Bioinform. 2003, 50, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Baskin, L.S. Electric conductance and pH measurements of isoionic salt-free bovine mercaptalbumin solutions. An evaluation of root-mean-square proton fluctuations. J. Phys. Chem. 1968, 72, 2958–2962. [Google Scholar] [CrossRef] [PubMed]
- MIu, L.; Bogatyreva, N.; Galzitskaia, O. Radius of gyration is indicator of compactness of protein structure. Mol. Biol. 2008, 42, 701–706. [Google Scholar]
- Bharatiy, S.K.; Hazra, M.; Paul, M.; Mohapatra, S.; Samantaray, D.; Dubey, R.C.; Sanyal, S.; Datta, S.; Hazra, S. In silico designing of an industrially sustainable carbonic anhydrase using molecular dynamics simulation. ACS Omega 2016, 1, 1081–1103. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | Content Range (mg g−1) |
|---|---|---|
| 1 | petunidin 3-O-p-coumaroylrutinoside-5-O-glucoside | 0.282~0.843 |
| 2 | petunidin 3-O-feruloylrutinoside-5-O-glucoside | 0.0963~0.269 |
| 3 | pelargonidin 3-O-p-coumaroylrutinoside-5-O-glucoside | 0~0.0387 |
| 4 | peonidin 3-O-p-coumaroylrutinoside-5-O-glucoside | 0~0.241 |
| 5 | malvidin 3-O-p-coumaroylrutinoside-5-O-glucoside | 0~0.656 |
| 6 | peonidin 3-O-feruloylrutinoside-5-O-glucoside | 0~0.218 |
| 7 | malvidin 3-O-feruloylrutinoside-5-O-glucoside | 0~0.212 |
| 8 | petunidin 3-O-rutinoside-5-O-glucoside | 0.510~1.34 |
| 9 | petunidin 3-O-rutinoside-5-O-rhamnoside | 0~1.13 |
| 10 | petunidin 3-O-caffeoylrutinoside-5-O-glucoside | 0~1.07 |
| 11 | petunidin 3-O-p-coumaroylrutinoside-5-O-glucoside | 1.12~2.08 |
| 12 | petunidin 3-O-feruloylrutinoside-5-O-glucoside | 0.520~1.13 |
| 14 | petunidin 3-O-p-coumaroylrutinoside | 0~0.400 |
| No. | 1H NMR | 13C NMR | No. | 1H NMR | 13C NMR |
|---|---|---|---|---|---|
| 2 | ― | 160.4 (s) | 3′′ | 3.90 (1H, overlapped) | 70.3 (d) |
| 3 | ― | 139.0 (s) | 4′′ | 4.94 (1H, t-like, 9.5) | 75.3 (d) |
| 4 | 7.69 (1H, s) | 117.2 (d) | 5′′ | 3.82 (1H, dq, 9.5, 6.2) | 67.6 (d) |
| 5 | ― | 155.4 (s) | 6′′ | 1.00 (3H, d, 6.2) | 17.8 (q) |
| 6 | 6.60 (1H, d, 1.4) | 101.2 (d) | Glc-1′′′ | 4.95 (1H, d, 7.8) | 102.7 (d) |
| 7 | ― | 161.4 (s) | 2′′′ | 3.57 (1H, dd, 8.5, 7.8) | 74.9 (d) |
| 8 | 6.37 (1H, d, 1.4) | 97.6 (d) | 3′′′ | 3.48 (1H, overlapped) | 78.3 (d) |
| 9 | ― | 153.3 (s) | 4′′′ | 3.49 (1H, overlapped) | 71.0 (d) |
| 10 | ― | 104.4 (s) | 5′′′ | 3.51 (1H, m) | 77.5 (d) |
| Glc-1′ | 5.07 (1H, d, 7.0) | 101.8 (d) | 6′′′ | 3.74 (1H, br d, 12.1) 3.88 (1H, overlapped) | 62.2 (t) |
| 2′ | 3.51 (1H, overlapped) | 74.4 (d) | 1′′′′ | ― | 127.2 (s) |
| 3′ | 3.49 (1H, overlapped) | 78.1 (d) | 2′′′′, 6′′′′ | 7.48 (2H, d, 8.4) | 131.3 (d) |
| 4′ | 3.41 (1H, t-like, 8.6) | 71.3 (d) | 3′′′′, 5′′′′ | 6.80 (2H, d, 8.4) | 116.8 (d) |
| 5′ | 3.69 (1H, m) | 76.9 (d) | 4′′′′ | ― | 161.3 (s) |
| 6′ | 3.69 (1H, overlapped) 4.01 (1H, br d, 9.6) | 67.0 (t) | 7′′′′ | 7.60 (1H, d, 15.9) | 147.0 (d) |
| Rha-1′′ | 4.76 (1H, br s) | 101.7 (d) | 8′′′′ | 6.34 (1H, d, 15.9) | 115.1 (d) |
| 2′′ | 3.88 (1H, br s) | 72.1 (d) | 9′′′′ | ― | 169.1 (s) |
| No. | 1H NMR | 13C NMR |
|---|---|---|
| Rha-1 | 5.04 (1H, br s) | 95.8 (d) |
| 2 | 3.83 (1H, br s) | 73.1 (d) |
| 3 | 3.93 (1H, dd, 9.7, 2.9) | 70.3 (d) |
| 4 | 5.02 (1H, t-like, 9.7) | 75.7 (d) |
| 5 | 4.01 (1H, dq, 9.7, 6.3) | 67.3 (d) |
| 6 | 1.14 (3H, d, 6.3) | 18.0 (q) |
| 1′ | ― | 127.2 (s) |
| 2′, 6′ | 7.47 (2H, d, 8.5) | 131.2 (d) |
| 3′, 5′ | 6.80 (2H, d, 8.5) | 116.8 (d) |
| 4′ | ― | 161.3 (s) |
| 7′ | 7.64 (1H, d, 15.9) | 146.8 (d) |
| 8′ | 6.36 (1H, d, 15.9) | 115.2 (d) |
| 9′ | ― | 169.0 (s) |
| NO. | Name | IC50 (μM) | Affinity (kcal/mol) | Amino Acids |
|---|---|---|---|---|
| 2 | Petanin | 122.37 ± 8.03 | −8.5 | GLN-307/THR-308/ASP-312/ASP-357/GLU-359/LYS-369/LYS-376/LYS-379 |
| 9 | Norpetanin | 115.53 ± 7.51 | −8.5 | TYR-65/ASN-81/HIS-85/HIS-244/ASN-260/MET-280 |
| 10 | 4-O-(p-coumaryl) rhamnose | 335.03 ± 12.99 | −7.6 | GLN-307 |
| 18 and 19 | Lyciruthephenylpropanoid D/E | 156.27 ± 11.22 | −8.4/−8.5 | ASN-81/HIS-85/ASN-260 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, J.; Hu, N.; Wang, H. Isolation, Purification and Tyrosinase Inhibitory Activity of Anthocyanins and Their Novel Degradation Compounds from Solanum tuberosum L. Molecules 2024, 29, 1492. https://doi.org/10.3390/molecules29071492
Ouyang J, Hu N, Wang H. Isolation, Purification and Tyrosinase Inhibitory Activity of Anthocyanins and Their Novel Degradation Compounds from Solanum tuberosum L. Molecules. 2024; 29(7):1492. https://doi.org/10.3390/molecules29071492
Chicago/Turabian StyleOuyang, Jian, Na Hu, and Honglun Wang. 2024. "Isolation, Purification and Tyrosinase Inhibitory Activity of Anthocyanins and Their Novel Degradation Compounds from Solanum tuberosum L." Molecules 29, no. 7: 1492. https://doi.org/10.3390/molecules29071492
APA StyleOuyang, J., Hu, N., & Wang, H. (2024). Isolation, Purification and Tyrosinase Inhibitory Activity of Anthocyanins and Their Novel Degradation Compounds from Solanum tuberosum L. Molecules, 29(7), 1492. https://doi.org/10.3390/molecules29071492