

Quercetin, a Flavonoid with Great Pharmacological Capacity

 ,
,
Abstract
1. Introduction
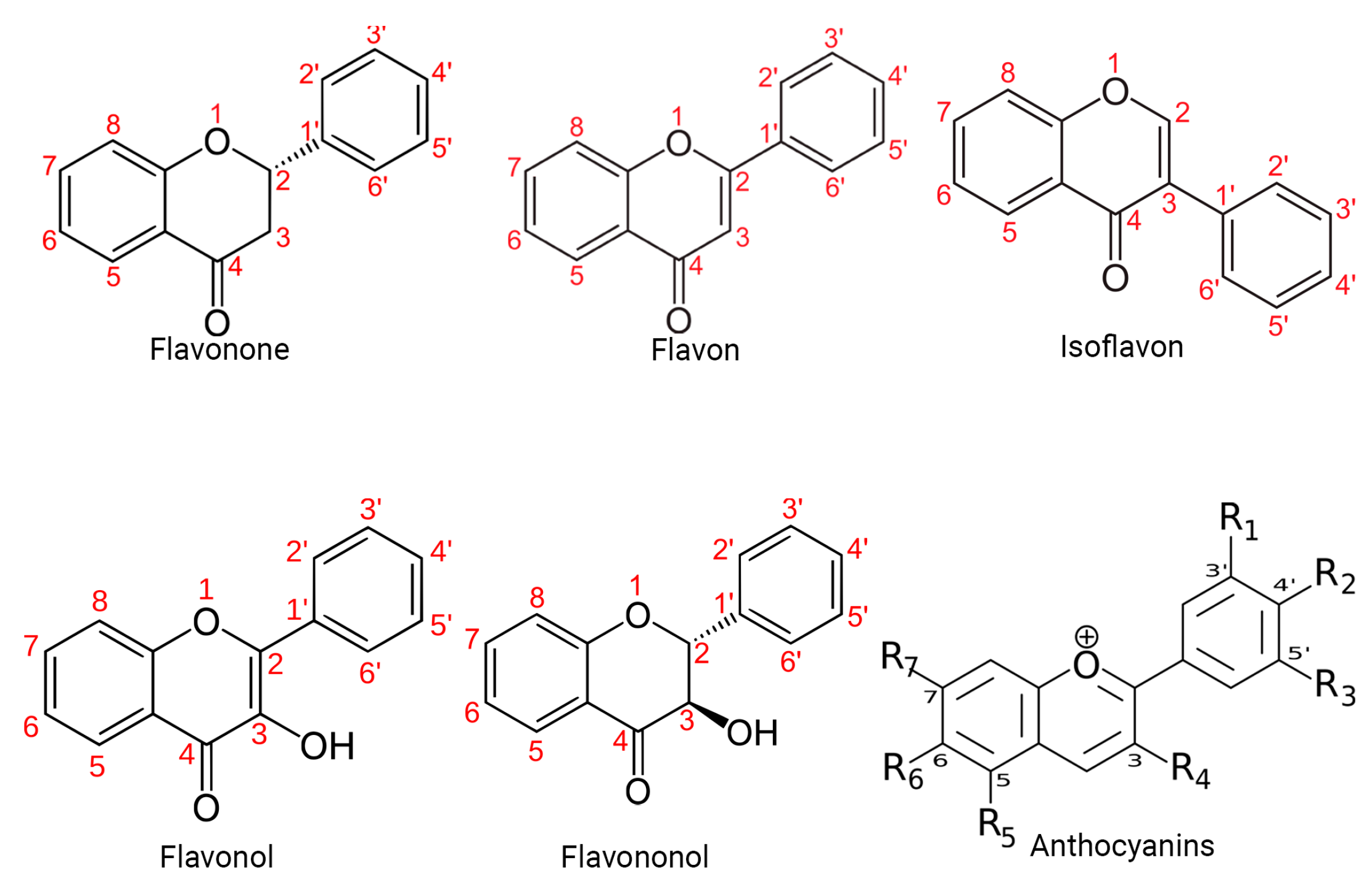
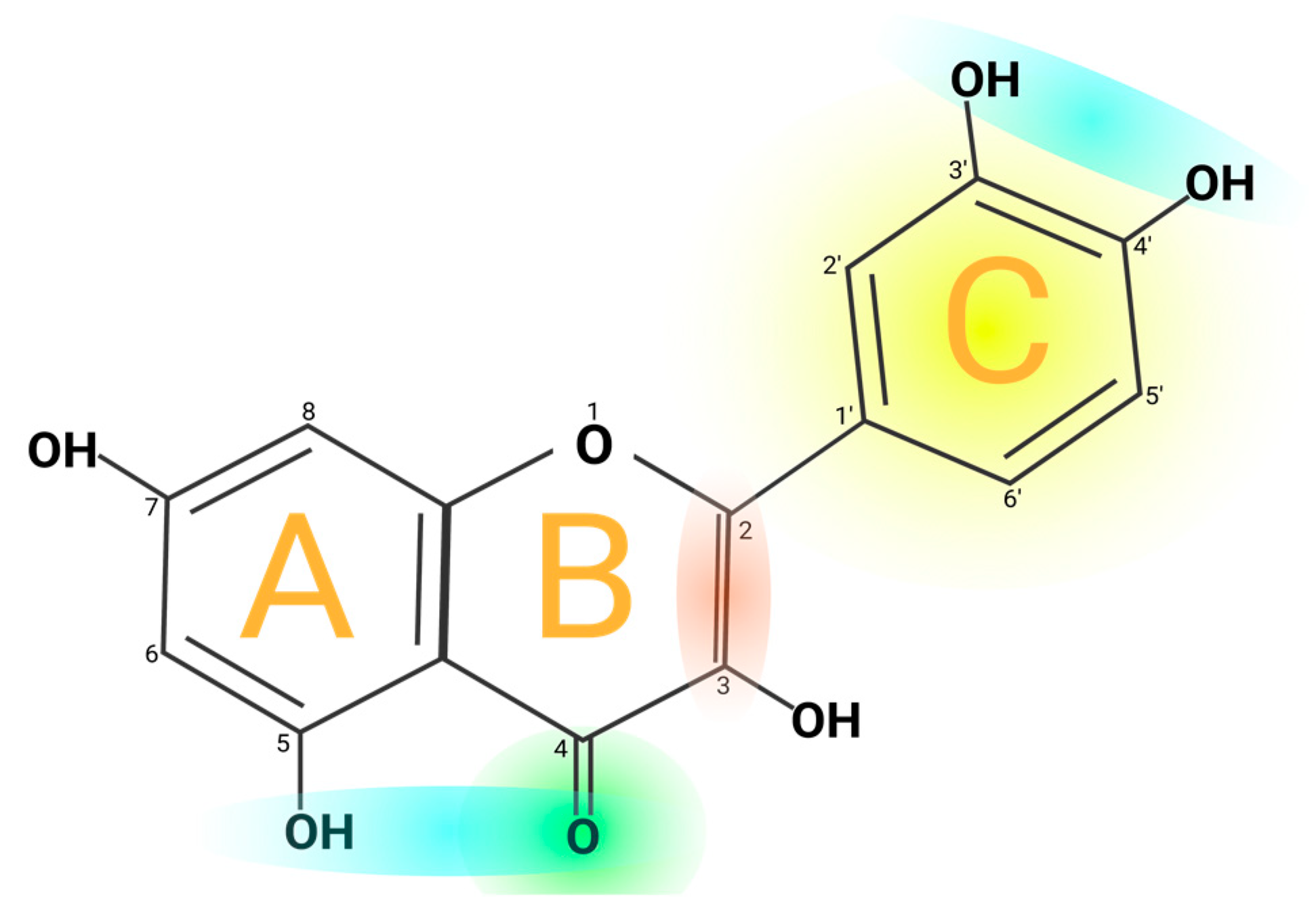
2. Quercetin Particularities
2.1. Sources
2.2. Physicochemical Properties
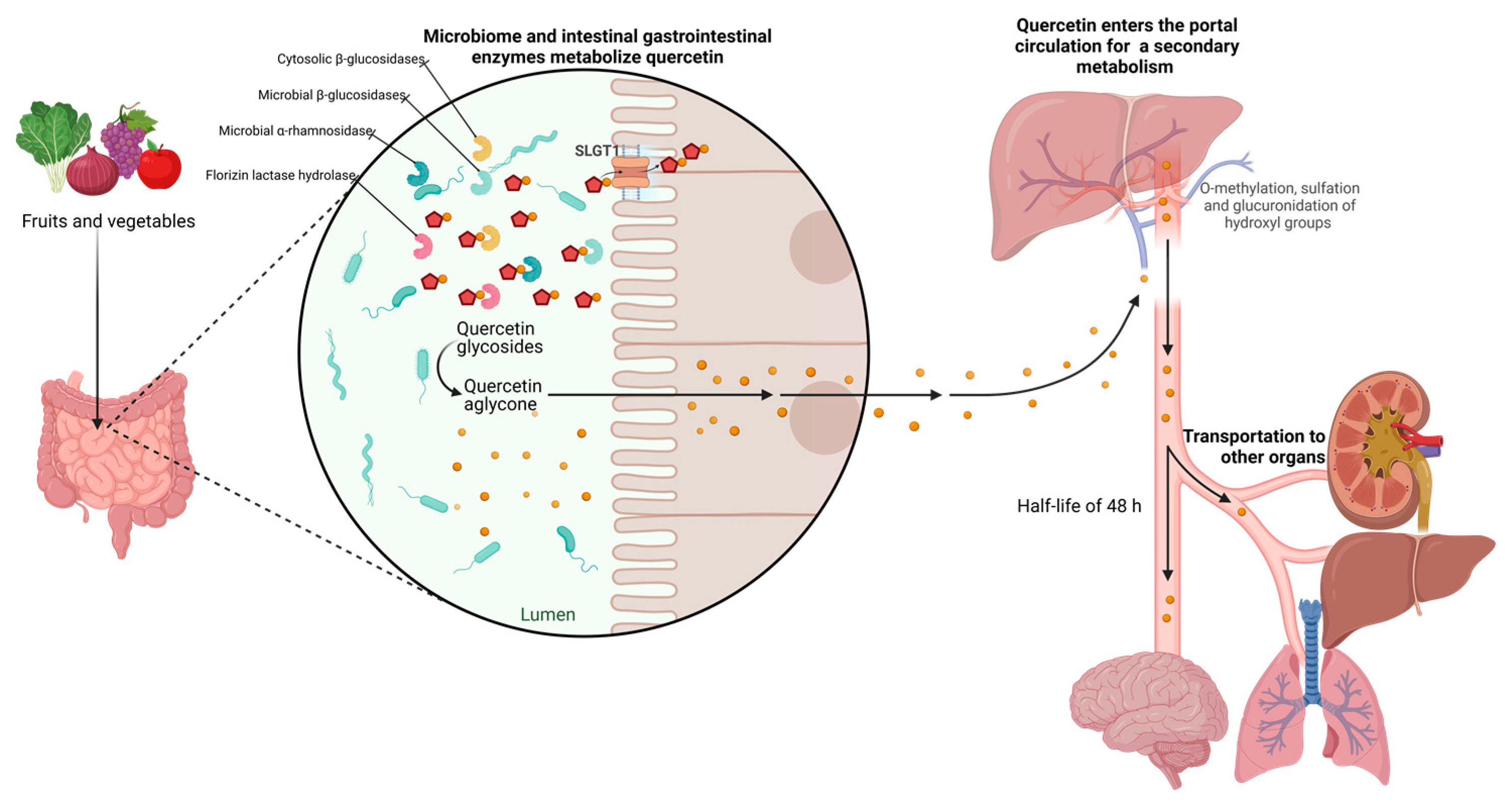
2.3. Bioavailability and Pharmacokinetics
2.4. Human Absorption and Metabolism of Quercetin
2.5. Quercetin Excretion
2.6. Pharmacological Properties
2.6.1. Antioxidant Properties
2.6.2. Cardiovascular Disease
Hypertensive Activity
Cardiovascular Protection
2.6.3. Alzheimer’s Disease
2.6.4. Antimicrobial Activity
2.6.5. Antiviral Activity
2.6.6. Hepatoprotective Activity
2.6.7. Oxidative Stress
2.6.8. Inflammation
2.6.9. Fibrosis
2.6.10. Cirrhosis
2.6.11. Diabetes
2.6.12. Arthritis
2.6.13. Cancer
Synergistic Effect
- Synergistic effect against breast cancer
- Synergistic effect against prostate cancer
- Synergistic effect against leukemia
- Synergistic effect in other types of cancer
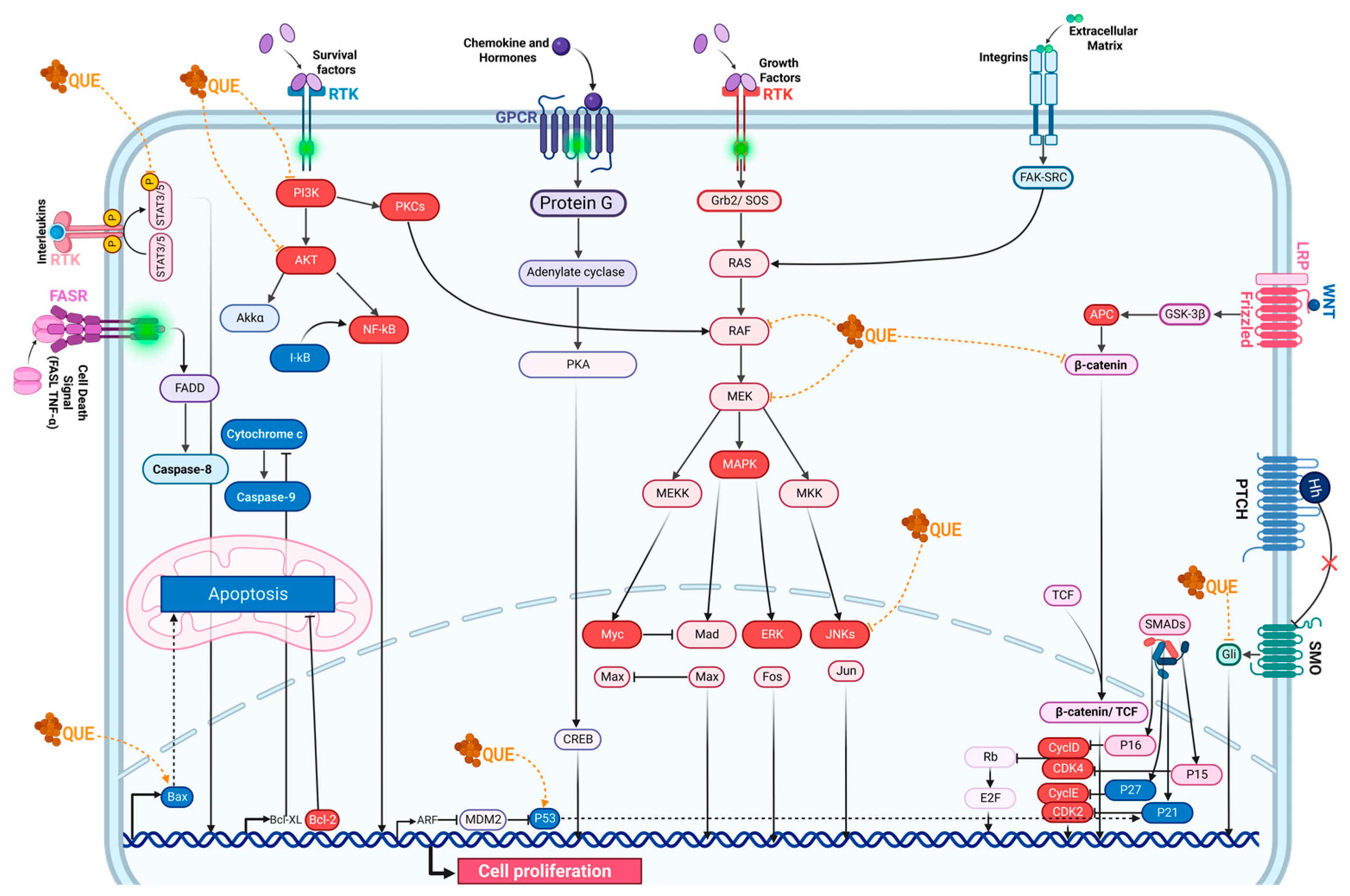
2.7. Molecular Pathways Targeted by Quercetin
2.7.1. Cell Cycle
2.7.2. Apoptosis
2.7.3. Wnt/β-Catenin Signaling
2.7.4. p53 Activity
2.7.5. Ras Expression
2.7.6. PI3K Signaling Pathway
2.7.7. NF-κB Signaling Pathway
2.7.8. Autophagy
2.8. Nanoparticles as Therapy
2.8.1. Liposomes
2.8.2. Lipid Nanoparticles
2.8.3. Polymeric Nanoparticles
PLGA Nanoparticles
2.8.4. Inorganic Nanoparticles
2.8.5. Silica Nanoparticles
2.8.6. Magnetic Nanoparticles
2.8.7. Extracellular Vesicles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Malignant Cell Line | Reference |
|---|---|
| HepG2 hepatocellular carcinoma cells | Parvez, Al-Dosari, Arbab, Al-Rehaily and Abdelwahid [54] |
| PC-3, LNCaP, and DU-145 human prostate cancer cells | Sharma, Raut, Baruah and Sharma [86] |
| MDA-MB-231 and AU565 human breast cancer cells | Molani Gol and Kheirouri [127] |
| Caco-2, DLD1, HT-29, SW620, HKE-3, HCT-116, FHC, DKO-4, and HKE-3 human colon cancer cells | Aziz, Lotfy, Said, El Ashry, El Tamany, Soliman, Abu-Serie, Teleb, Yousuf, Dömling, Domingo and Barakat [107] |
| HGC-27, NUGC-2, MKN-7, and MKN-28 human gastric cancer cells | SDF |
| 143B highly metastatic human osteosarcoma cell line | Doghish, Hegazy, Ismail, El-Mahdy, Elsakka, Elkhawaga, Elkady, Yehia, Abdelmaksoud and Mokhtar [94] |
| HPB-ALL chronic leukemia B-cells | Shi, Su, Cui, Yu, Du, and Han [87] |
| H460 lung cancer cell lines | Sul and Ra [112] |
| MNT1, M10, and M14 human melanoma cells | Córdoba-Moreno, Mendes, Markus and Fernandes [68] |
| A2780s and A2780cp ovarian cancer cell lines | Vafadar, Shabaninejad, Movahedpour, Fallahi, Taghavipour, Ghasemi, Akbari, Shafiee, Hajighadimi, Moradizarmehri, Razi, Savardashtaki and Mirzaei [124] |
| HeLa cervical cancer cell lines | Shorobi, Nisa, Saha, Chowdhury, Srisuphanunt, Hossain and Rahman [14] |
3. Conclusions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| RNS | Reactive nitrogen species |
| RCS | Reactive chlorine species |
| LDL | Low-density lipoprotein |
| FR | Free radical |
| COX | Cyclooxygenase |
| SGLT1 | Sodium glucose cotransporter 1 |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| LPH | Lactase-phlorizin hydrolase |
| Q4’G | Quercetin-4’-glucoside |
| MRP2 | Multidrug resistance-associated protein 2 |
| UDP | Uridine diphosphate |
| UGT-1A9 | UDP-glucuronosyltransferase 1A9 |
| HIV | Human immunodeficiency virus |
| HBV | Hepatitis B virus |
| HSV | Herpes simplex virus |
| GSH | Glutathione |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| ARE | Antioxidant response element |
| NQO1 | NAD (P)H dehydrogenase quinone 1 |
| GSTA | Glutathione S-transferase |
| HMOX1 | Heme oxygenase |
| FTL | Ferritin light chain |
| NOS | Nitric oxide synthase |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| ENOS | Endothelial nitric oxide synthase |
| NO | Nitric oxide |
| ACE | Angiotensin-converting enzyme |
| cGMP | Cyclic guanosine monophosphate |
| GTN | Glyceryl trinitrate |
| SNP | Sodium nitroprusside |
| CAD | Coronary artery disease |
| NF-Κb | Nuclear factor kappa B |
| TNF-α | Necrosis tumoral alfa |
| IL-10 | Interleukin 10 |
| IL-1β | Interleukin 1β |
| Ikβα | Βα kappa inhibitor |
| AD | Alzheimer’s disease |
| Aβ | Amyloid-beta |
| BACE1 | Beta-site amyloid precursor protein-cleaving enzyme 1 |
| NFTs | Neurofibrillary tangles |
| GSK | Glycogen synthase kinase |
| MRSA | Methicillin-resistant staphylococcus aureus |
| TEM | Transmission electron microscopy |
| HCV | Hepatitis C virus |
| IAV | Influenza A virus |
| CHIKV | Chikungunya virus |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| DNA | Deoxyribonucleic acid |
| RNA | Ribonucleic acid |
| NS3 | Nonstructural protein 3 |
| Akt | Serine/threonine protein kinase |
| SRC | Sarcoma |
| EGFR | Epidermal growth factor receptor |
| MMP | Matrix-metalloprotein |
| KDR | Kinase insert domain receptor |
| IGF1R | Insulin-like growth factor 1 receptor |
| PTK2 | Protein tyrosine kinase 2 |
| ABCG2 | ATP-binding cassette super-family G |
| BCRP | Breast cancer resistance protein |
| MET | Mesenchymal–epithelial transition |
| RMLV | Rauscher murine leukemia virus |
| HSCs | Hematopoietic stem cells |
| α-SMA | Alpha smooth muscle actin |
| TGF-Β1 | Transforming growth factor beta 1 |
| PDGF | Platelet-derived growth factor |
| CTGF | Connective tissue growth factor |
| FGF | Fibroblast growth factor |
| EGF | Epithelial growth factor |
| ILGF | Insulin-like growth factor |
| TGFβ3 | Transforming growth factor beta 3 |
| G1 | Growth 1 |
| P53 | Protein 53 |
| P21 | Protein 21 |
| P27 | Protein 27 |
| FAS | Apoptosis antigen 1 |
| STZ | Streptozotocin |
| GLUT4 | Glucose transporter type 4 |
| PI3K | Phosphoinositide 3-kinases |
| OA | Osteoarthritis |
| NSAIDs | Nonsteroidal anti-inflammatory drugs |
| COX-2 | Cyclooxygenase-2 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| HO-1 | Heme oxygenase-1 |
| MTOR | Mammalian target of rapamycin |
| ECM | Cartilage extracellular matrix |
| MCP-1 | Monocyte chemoattractant protein-1 |
| PKC | Protein kinase C |
| DAG | Diacylglycerol |
| HL-60 | Human leukemia 60 |
| TPKs | Tyrosine protein kinases |
| ERK1 | Extracellular signal-regulated kinase |
| BAX | Bcl-2-associated X-protein |
| C-MYC | Cellular myelocytomatosis |
| K-RAS | Kirsten rat sarcoma |
| MPT | Mitochondrial permeability transition |
| SMAC | Second mitochondria-derived activator of caspase |
| DIABLO | Direct inhibitor of apoptosis-binding protein with low pI |
| MCF7 | Michigan Cancer Foundation-7 |
| PI | Phosphatidylinositol |
| PIP | Phosphatidylinositol phosphate |
| AR | Androgen receptor |
| BCL-2 | B-cell lymphoma 2 |
| VEGF | Vascular endothelial growth factor |
| COMT | Catechol-O-methyltransferase |
| SOD | Superoxide dismutase |
| MDA | Malondialdehyde |
| ADR | Adriamycin |
| EGCG | Epigallocatechin gallate |
| JAK | Janus kinase |
| STAT | Signal transducer and activator of transcription |
| HSP | Heat shock protein |
| NQO1 | NAD (P)H:quinone oxidoreductase 1 |
| Dox | Doxorubicin |
| CDKs | Cyclin-dependent kinases |
| CDKIs | CDK inhibitors |
| PRB | Retinoblastoma protein |
| TOPOII | Type II topoisomerase |
| DSBs | DNA double-strand breaks |
| PARP | Poly ADP ribose polymerase |
| CYT C | Cytochrome complex |
| MMP | Mitochondrial membrane potential |
| COX | Cyclooxygenase |
| BCL-XL | B-cell lymphoma-extra-large |
| BCL-XS | B-cell lymphoma-extra-small |
| JNKs | Jun N-terminal kinases |
| WNT | Wingless and Int-1 |
| TCF | T cell factor |
| TNBC | Triple-negative breast cancer |
| EMT | Epithelial–mesenchymal transition |
| PUMA | P53-upregulated modulator of apoptosis |
| NOXA | Phorbol-12-myristate-13-acetate-induced protein 1 |
| MDM2 | Mouse double minute 2 |
| NAG-1 | Nonsteroidal anti-inflammatory drug-activated gene-1 |
| EGR-1 | Early growth response 1 |
| SP1 | Specificity protein 1 |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| H-RAS | Harvey rat sarcoma |
| N-RAS | Neuroblastoma rat sarcoma |
| SER | Serine |
| MTORC2 | Mammalian target of rapamycin complex 2 |
| HUVECs | Human umbilical vein endothelial cells |
| I-Κβ | Inhibitor of NF-Κβ |
| AP-1 | Activating protein-1 |
| MAPKs | Mitogen-activated protein kinases |
| CAMP | Cyclic adenosine monophosphate |
| CREB | Camp-response element-binding protein |
| ATF2 | Activating transcription factor 2 |
| PCD | Programmed cell death |
| 4E-BP1 | Eukaryotic translation initiation factor 4E-binding protein 1 |
| P70S6K | P70 ribosomal S6 kinase |
| HIF-1α | Hypoxia inducible factor 1 subunit alpha |
| TRAIL | Tumor necrosis factor-related apoptosis-inducing ligand |
| LC3B | Microtubule-associated protein 1 light chain 3B |
| PLGA | Poly-lactic-co-glycolic acid |
| MPA | Mycophenolic acid |
| LNPs | Lipid nanoparticles |
| P-Gp | P-glycoprotein |
| MLVs | Multilamellar vesicles |
| ATP | Adenosine triphosphate |
| PLA | Poly-lactic acid |
| NLCs | Nanostructured lipid carriers |
| SLNs | Solid lipid nanoparticles |
| QU | Quercetin |
| TQ | Thymoquinone |
| PEG | Polyethylene glycol |
| Mab | Monoclonal antibody |
| MDR | Multidrug resistance |
| ABC | ATP-binding cassette |
| FDA | Food And Drug Administration |
| NPs | Nanoparticles |
| MSNS | Mesoporous silica nanoparticles |
| FA | Folic acid |
| SBA-15 | Santa Barbara amorphous-15 |
| H2AX | H2A histone family member X |
| MRI | Magnetic resonance imaging |
| Sio2 | Silicon dioxide |
| BBB | Blood–brain barrier |
| BTN | Biotin |
| FITC | Fluorescein isothiocyanate |
| EVS | Extracellular vesicles |
| GAP | Growth-associated protein |
References
- Flores, G.P. Relaciones entre el Estrés Oxidativo y la Salud. Soc. Rural. Prod. Medio Ambiente 2019, 19, 34. [Google Scholar]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Futur. J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef]
- Terao, J. Potential Role of Quercetin Glycosides as Anti-Atherosclerotic Food-Derived Factors for Human Health. Antioxidants 2023, 12, 258. [Google Scholar] [CrossRef]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2020, 60, 3290–3303. [Google Scholar] [CrossRef] [PubMed]
- Heřmánková, E.; Zatloukalová, M.; Biler, M.; Sokolová, R.; Bancířová, M.; Tzakos, A.G.; Křen, V.; Kuzma, M.; Trouillas, P.; Vacek, J. Redox properties of individual quercetin moieties. Free Radic. Biol. Med. 2019, 143, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Pozo, C.; Cires, M.J.; Gotteland, M. Quercetin and epigallocatechin gallate in the prevention and treatment of obesity: From molecular to clinical studies. J. Med. Food 2019, 22, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Ye, H.; Kamaraj, R.; Zhang, T.; Zhang, J.; Pavek, P. A review on pharmacological activities and synergistic effect of quercetin with small molecule agents. Phytomedicine 2021, 92, 153736. [Google Scholar] [CrossRef] [PubMed]
- Cattivelli, A.; Conte, A.; Martini, S.; Tagliazucchi, D. Influence of Cooking Methods on Onion Phenolic Compounds Bioaccessibility. Foods 2021, 10, 1023. [Google Scholar] [CrossRef] [PubMed]
- Mourabit, Y.; El Hajjaji, S.; Taha, D.; Badaoui, B.; El Yadini, M.; Rusu, M.E.; Lee, L.-H.; Bouyahya, A.; Bourais, I. HPLC-DAD-ESI/MS phytochemical investigation, antioxidant, and antidiabetic activities of Moroccan Rosa canina L. extracts. Biocatal. Agric. Biotechnol. 2023, 52, 102817. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, M.R. Flavonoids a Bioactive Compound from Medicinal Plants and Its Therapeutic Applications. Biomed Res. Int. 2022, 2022, 5445291. [Google Scholar] [CrossRef]
- Tronina, T.; Łużny, M.; Dymarska, M.; Urbaniak, M.; Kozłowska, E.; Piegza, M.; Stępień, Ł.; Janeczko, T. Glycosylation of Quercetin by Selected Entomopathogenic Filamentous Fungi and Prediction of Its Products’ Bioactivity. Int. J. Mol. Sci. 2023, 24, 11857. [Google Scholar] [CrossRef] [PubMed]
- Shorobi, F.M.; Nisa, F.Y.; Saha, S.; Chowdhury, M.A.H.; Srisuphanunt, M.; Hossain, K.H.; Rahman, M.A. Quercetin: A Functional Food-Flavonoid Incredibly Attenuates Emerging and Re-Emerging Viral Infections through Immunomodulatory Actions. Molecules 2023, 28, 938. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Russo, C.; Statti, G.; Argentieri, M.P.; Meleleo, D.; Mallamaci, R.; Avato, P.; Conforti, F. Phytochemical and biological characterization of dry outer scales extract from Tropea red onion (Allium cepa L. var. Tropea)–A promising inhibitor of pancreatic lipase. Phytomed. Plus 2022, 2, 100235. [Google Scholar] [CrossRef]
- Roszkowski, S. Application of Polyphenols and Flavonoids in Oncological Therapy. Molecules 2023, 28, 4080. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, R.U.; Dhanaraj, S. A critical review on quercetin bioflavonoid and its derivatives: Scope, synthesis, and biological applications with future prospects. Arab. J. Chem. 2023, 16, 104881. [Google Scholar] [CrossRef]
- Kooshki, L.; Zarneshan, S.N.; Fakhri, S.; Moradi, S.Z.; Echeverria, J. The pivotal role of JAK/STAT and IRS/PI3K signaling pathways in neurodegenerative diseases: Mechanistic approaches to polyphenols and alkaloids. Phytomedicine 2023, 112, 154686. [Google Scholar] [CrossRef]
- George, J.; Edwards, D.; Pun, S.; Williams, D. Evaluation of Antioxidant Capacity (ABTS and CUPRAC) and Total Phenolic Content (Folin-Ciocalteu) Assays of Selected Fruit, Vegetables, and Spices. Int. J. Food Sci. 2022, 2022, 2581470. [Google Scholar] [CrossRef]
- Golmohammadi, M.; Elmaghraby, D.A.; Ramírez-Coronel, A.A.; Rakhimov, N.; Mohammed, S.S.; Romero-Parra, R.M.; Jawad, M.A.; Zamanian, M.Y.; Soltani, A.; Taheri, N.; et al. A comprehensive view on the quercetin impact on bladder cancer: Focusing on oxidative stress, cellular, and molecular mechanisms. Fundam. Clin. Pharmacol. 2023, 37, 900–909. [Google Scholar] [CrossRef]
- Xiong, H.H.; Lin, S.Y.; Chen, L.L.; Ouyang, K.H.; Wang, W.J. The Interaction between Flavonoids and Intestinal Microbes: A Review. Foods 2023, 12, 320. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Chen, H.Y.; Tang, Q.Q.; Li, Y.F.; Liu, X.S.; Lu, F.H.; Gu, Y.Y. Protective effect of quercetin on kidney diseases: From chemistry to herbal medicines. Front. Pharmacol. 2022, 13, 968226. [Google Scholar] [CrossRef]
- Kandemir, K.; Tomas, M.; McClements, D.J.; Capanoglu, E. Recent advances on the improvement of quercetin bioavailability. Trends Food Sci. Technol. 2022, 119, 192–200. [Google Scholar] [CrossRef]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef]
- Owczarek-Januszkiewicz, A.; Magiera, A.; Olszewska, M.A. Enzymatically Modified Isoquercitrin: Production, Metabolism, Bioavailability, Toxicity, Pharmacology, and Related Molecular Mechanisms. Int. J. Mol. Sci. 2022, 23, 14784. [Google Scholar] [CrossRef] [PubMed]
- Billowria, K.; Ali, R.; Rangra, N.K.; Kumar, R.; Chawla, P.A. Bioactive Flavonoids: A Comprehensive Review on Pharmacokinetics and Analytical Aspects. Crit. Rev. Anal. Chem. 2022, 1–5. [Google Scholar] [CrossRef]
- Ullah, H.; Minno, A.D.; Santarcangelo, C.; Tantipongpiradet, A.; Dacrema, M.; Matteo, R.D.; El-Seedi, H.R.; Khalifa, S.A.M.; Baldi, A.; Rossi, A.; et al. In Vitro Bioaccessibility and Anti-Inflammatory Activity of a Chemically Characterized Allium cepa L. Extract Rich in Quercetin Derivatives Optimized by the Design of Experiments. Molecules 2022, 27, 9065. [Google Scholar] [CrossRef]
- Rha, C.-S.; Choi, J.-M.; Jung, Y.S.; Kim, E.-R.; Ko, M.J.; Seo, D.-H.; Kim, D.-O.; Park, C.-S. High-efficiency enzymatic production of α-isoquercitrin glucosides by amylosucrase from Deinococcus geothermalis. Enzym. Microb. Technol. 2019, 120, 84–90. [Google Scholar] [CrossRef]
- Zhang, Y.; Mu, T.; Deng, X.; Guo, R.; Xia, B.; Jiang, L.; Wu, Z.; Liu, M. New Insights of Biological Functions of Natural Polyphenols in Inflammatory Intestinal Diseases. Int. J. Mol. Sci. 2023, 24, 9581. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.-H.; Lee, J.H.; Lee, Y.-J. Evaluation of the Mrp2-mediated flavonoid-drug interaction potential of quercetin in rats and in vitro models. Asian J. Pharm. Sci. 2019, 14, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.H.; Eslami, A.; Jelodar, S.k.; Ranjbar, M.; Hasantabar, V. Preventive effect of quercetin-Loaded nanophytosome against autistic-like damage in maternal separation model: The possible role of Caspase-3, Bax/Bcl-2 and Nrf2. Behav. Brain Res. 2023, 441, 114300. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, N.; Yaghoobi, A.; Salami, M.; Gholinezhad, Y.; Aghadavood, F.; Eshraghi, R.; Aarabi, M.H.; Homayoonfal, M.; Asemi, Z.; Mirzaei, H.; et al. Impact of polyphenols on heart failure and cardiac hypertrophy: Clinical effects and molecular mechanisms. Front. Cardiovasc. Med. 2023, 10, 1174816. [Google Scholar] [CrossRef] [PubMed]
- Bešlo, D.; Golubić, N.; Rastija, V.; Agić, D.; Karnaš, M.; Šubarić, D.; Lučić, B. Antioxidant Activity, Metabolism, and Bioavailability of Polyphenols in the Diet of Animals. Antioxidants 2023, 12, 1141. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.L.; Zhou, H.F.; Yang, J.; Wang, F.X.; Sun, F.; Li, J.Y. Biological Activities Underlying the Therapeutic Effect of Quercetin on Inflammatory Bowel Disease. Mediat. Inflamm. 2022, 2022, 5665778. [Google Scholar] [CrossRef]
- Cheng, S.C.; Huang, W.C.; JH, S.P.; Wu, Y.H.; Cheng, C.Y. Quercetin Inhibits the Production of IL-1β-Induced Inflammatory Cytokines and Chemokines in ARPE-19 Cells via the MAPK and NF-κB Signaling Pathways. Int. J. Mol. Sci. 2019, 20, 2957. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, J.; Tang, S.; Yan, J.; Chen, H.; Li, D.; Yan, X. Quercetin regulates inflammation, oxidative stress, apoptosis, and mitochondrial structure and function in H9C2 cells by promoting PVT1 expression. Acta Histochem. 2021, 123, 151819. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef]
- Pan, E.; Feng, H.; Yang, Z.; Xin, Y.; Ji, X.; Ping, K.; Sun, Y.; Dong, J. Quercetin dietary supplementation protects against difenoconazole-induced carp spleen inflammatory damage via regulating ROS/NF-κB/NLRP3 inflammasome axis. Aquaculture 2024, 579, 740162. [Google Scholar] [CrossRef]
- Wuputra, K.; Tsai, M.H.; Kato, K.; Ku, C.C.; Pan, J.B.; Yang, Y.H.; Saito, S.; Wu, C.C.; Lin, Y.C.; Cheng, K.H.; et al. Jdp2 is a spatiotemporal transcriptional activator of the AhR via the Nrf2 gene battery. Inflamm. Regen. 2023, 43, 42. [Google Scholar] [CrossRef]
- Da Rocha, E.V.; Falchetti, F.; Pernomian, L.; De Mello, M.M.B.; Parente, J.M.; Nogueira, R.C.; Gomes, B.Q.; Bertozi, G.; Sanches-Lopes, J.M.; Tanus-Santos, J.E. Quercetin decreases cardiac hypertrophic mediators and maladaptive coronary arterial remodeling in renovascular hypertensive rats without improving cardiac function. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023, 396, 939–949. [Google Scholar] [CrossRef]
- Abdelghffar, E.A.; Obaid, W.A.; Elgamal, A.M.; Daoud, R.; Sobeh, M.; El Raey, M.A. Pea (Pisum sativum) peel extract attenuates DOX-induced oxidative myocardial injury. Biomed. Pharmacother. 2021, 143, 112120. [Google Scholar] [CrossRef]
- Popiolek-Kalisz, J.; Fornal, E. The Effects of Quercetin Supplementation on Blood Pressure—Meta-Analysis. Curr. Probl. Cardiol. 2022, 47, 101350. [Google Scholar] [CrossRef]
- Das, M.; Devi, K.P.; Belwal, T.; Devkota, H.P.; Tewari, D.; Sahebnasagh, A.; Nabavi, S.F.; Khayat Kashani, H.R.; Rasekhian, M.; Xu, S.; et al. Harnessing polyphenol power by targeting eNOS for vascular diseases. Crit. Rev. Food Sci. Nutr. 2023, 63, 2093–2118. [Google Scholar] [CrossRef]
- Ra, J.-E.; Woo, S.-Y.; Jin, H.; Lee, M.J.; Kim, H.Y.; Ham, H.; Chung, I.-M.; Seo, W.D. Evaluation of antihypertensive polyphenols of barley (Hordeum vulgare L.) seedlings via their effects on angiotensin-converting enzyme (ACE) inhibition. Appl. Biol. Chem. 2020, 63, 1–9. [Google Scholar] [CrossRef]
- Fikriah, I.; Ismail, S.; Kosala, K. In vitro evaluation of the vasodilatory activity of ethanol extracts of Eleutherine bulbosa bulbs and leaves. J. Appl. Pharm. Sci. 2021, 11, 135–140. [Google Scholar]
- Solfaine, R.; Muniroh, L.; Irawan, A. Roles of averrhoa bilimbi extract in increasing serum nitric oxide concentration and vascular dilatation of ethanol-induced hypertensive rats. Prev. Nutr. Food Sci. 2021, 26, 186. [Google Scholar] [CrossRef]
- Chekalina, N.; Burmak, Y.; Petrov, Y.; Borisova, Z.; Manusha, Y.; Kazakov, Y.; Kaidashev, I. Quercetin reduces the transcriptional activity of NF-kB in stable coronary artery disease. Indian Heart J. 2018, 70, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, Y.; Zhang, J.; Zhang, X.; Yang, G. Molecular mechanism of autophagy: Its role in the therapy of Alzheimer’s disease. Curr. Neuropharmacol. 2020, 18, 720–739. [Google Scholar] [CrossRef]
- Guo, M.; Zhu, F.; Qiu, W.; Qiao, G.; Law, B.Y.-K.; Yu, L.; Wu, J.; Tang, Y.; Yu, C.; Qin, D.; et al. High-throughput screening for amyloid-β binding natural small-molecules based on the combinational use of biolayer interferometry and UHPLC−DAD-Q/TOF-MS/MS. Acta Pharm. Sin. B 2022, 12, 1723–1739. [Google Scholar] [CrossRef]
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer’s disease. Life Sci. 2019, 224, 109–119. [Google Scholar] [CrossRef]
- Wang, S.; Yao, J.; Zhou, B.; Yang, J.; Chaudry, M.T.; Wang, M.; Xiao, F.; Li, Y.; Yin, W. Bacteriostatic effect of quercetin as an antibiotic alternative in vivo and its antibacterial mechanism in vitro. J. Food Prot. 2018, 81, 68–78. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Bhattacharya, D. Antimicrobial Activity of Quercetin: An Approach to Its Mechanistic Principle. Molecules 2022, 27, 2494. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Shorobi, F.M.; Uddin, M.N.; Saha, S.; Hossain, M.A. Quercetin attenuates viral infections by interacting with target proteins and linked genes in chemicobiological models. Silico Pharmacol. 2022, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.K.; Al-Dosari, M.S.; Arbab, A.H.; Al-Rehaily, A.J.; Abdelwahid, M.A.S. Bioassay-guided isolation of anti-hepatitis B virus flavonoid myricetin-3-O-rhamnoside along with quercetin from Guiera senegalensis leaves. Saudi Pharm. J. 2020, 28, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.F.d.; Salatino, A.; Motta, L.B.d.; Negri, G.; Salatino, M.L.F. Chemical characterization, antioxidant and anti-HIV activities of a Brazilian propolis from Ceará state. Rev. Bras. Farmacogn. 2019, 29, 309–318. [Google Scholar] [CrossRef]
- Upadhyay, R.; Tiwari, K.N. The antiviral potential of Phyllanthus species: A systematic review. Arch. Virol. 2023, 168, 177. [Google Scholar] [CrossRef]
- Shao, C.; Xu, H.; Sun, X.; Huang, Y.; Guo, W.; He, Y.; Ye, L.; Wang, Z.; Huang, J.; Liang, X.; et al. New Perspectives on Chinese Medicine in Treating Hepatic Fibrosis: Lipid Droplets in Hepatic Stellate Cells. Am. J. Chin. Med. 2023, 51, 1413–1429. [Google Scholar] [CrossRef]
- Alshammari, G.M.; Al-Qahtani, W.H.; AlFaris, N.A.; Alzahrani, N.S.; Alkhateeb, M.A.; Yahya, M.A. Quercetin prevents cadmium chloride-induced hepatic steatosis and fibrosis by downregulating the transcription of miR-21. BioFactors 2021, 47, 489–505. [Google Scholar] [CrossRef]
- Shakerian, E.; Akbari, R.; Mohammadtaghvaei, N.; Mohammadi Gahrooie, M.; Afarin, R. Quercetin Reduces Hepatic Fibrogenesis by Inhibiting TGF-β/Smad3 Signaling Pathway in LX-2 Cell Line. Jundishapur. J. Nat. Pharm. Prod. 2022, 17, e113484. [Google Scholar] [CrossRef]
- Serna-Salas, S.A.; Arroyave-Ospina, J.C.; Zhang, M.; Damba, T.; Buist-Homan, M.; Muñoz-Ortega, M.H.; Ventura-Juárez, J.; Moshage, H. α-1 Adrenergic receptor antagonist doxazosin reverses hepatic stellate cells activation via induction of senescence. Mech. Ageing Dev. 2022, 201, 111617. [Google Scholar] [CrossRef]
- Owen, T.; Carpino, G.; Chen, L.; Kundu, D.; Wills, P.; Ekser, B.; Onori, P.; Gaudio, E.; Alpini, G.; Francis, H.; et al. Endothelin Receptor-A Inhibition Decreases Ductular Reaction, Liver Fibrosis, and Angiogenesis in a Model of Cholangitis. Cell. Mol. Gastroenterol. Hepatol. 2023, 16, 513–540. [Google Scholar] [CrossRef]
- Ansari, P.; Choudhury, S.T.; Seidel, V.; Rahman, A.B.; Aziz, M.A.; Richi, A.E.; Rahman, A.; Jafrin, U.H.; Hannan, J.M.A.; Abdel-Wahab, Y.H.A. Therapeutic Potential of Quercetin in the Management of Type-2 Diabetes Mellitus. Life 2022, 12, 1146. [Google Scholar] [CrossRef]
- Shi, G.-J.; Li, Y.; Cao, Q.-H.; Wu, H.-X.; Tang, X.-Y.; Gao, X.-H.; Yu, J.-Q.; Chen, Z.; Yang, Y. In vitro and in vivo evidence that quercetin protects against diabetes and its complications: A systematic review of the literature. Biomed. Pharmacother. 2019, 109, 1085–1099. [Google Scholar] [CrossRef]
- Hamilton, K.E.; Rekman, J.F.; Gunnink, L.K.; Busscher, B.M.; Scott, J.L.; Tidball, A.M.; Stehouwer, N.R.; Johnecheck, G.N.; Looyenga, B.D.; Louters, L.L. Quercetin inhibits glucose transport by binding to an exofacial site on GLUT1. Biochimie 2018, 151, 107–114. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; El Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic Potential of Quercetin: New Insights and Perspectives for Human Health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef] [PubMed]
- Borghi, S.M.; Mizokami, S.S.; Pinho-Ribeiro, F.A.; Fattori, V.; Crespigio, J.; Clemente-Napimoga, J.T.; Napimoga, M.H.; Pitol, D.L.; Issa, J.P.; Fukada, S.Y. The flavonoid quercetin inhibits titanium dioxide (TiO2)-induced chronic arthritis in mice. J. Nutr. Biochem. 2018, 53, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Yin, W.; Zou, Z.; Zhang, C.; Sun, M.; Min, L.; Yang, L.; Kong, L. Quercitrin alleviates cartilage extracellular matrix degradation and delays ACLT rat osteoarthritis development: An in vivo and in vitro study. J. Adv. Res. 2021, 28, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Córdoba-Moreno, M.O.; Mendes, M.T.; Markus, R.P.; Fernandes, P.A. Rat resistance to rheumatoid arthritis induction as a function of the early-phase adrenal–pineal crosstalk. J. Physiol. 2023, 601, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Das, S.; Patra, S.K.; Efferth, T.; Jena, M.; Bhutia, S.K. Dietary polyphenols in chemoprevention and synergistic effect in cancer: Clinical evidences and molecular mechanisms of action. Phytomedicine 2021, 90, 153554. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Z.; Gao, Y.; Liu, S.; Li, K.; Wang, S.; Gao, L.; Shi, M.; Liu, Z.; Han, Z.; et al. Effect of a Drug Delivery System Made of Quercetin Formulated into PEGylation Liposomes on Cervical Carcinoma In Vitro and In vivo. J. Nanomater. 2021, 2021, 9389934. [Google Scholar] [CrossRef]
- Tang, S.-M.; Deng, X.-T.; Zhou, J.; Li, Q.-P.; Ge, X.-X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed. Pharmacother. 2020, 121, 109604. [Google Scholar] [CrossRef]
- Joyner, P.M. Protein Adducts and Protein Oxidation as Molecular Mechanisms of Flavonoid Bioactivity. Molecules 2021, 26, 5102. [Google Scholar] [CrossRef]
- Shah, S.; Narang, R.; Singh, V.J.; Pilli, G.; Nayak, S.K. A Review on Anticancer Profile of Flavonoids: Sources, Chemistry, Mechanisms, Structure-activity Relationship and Anticancer Activity. Curr. Drug Res. Rev. Former. Curr. Drug Abus. Rev. 2023, 15, 122–148. [Google Scholar] [CrossRef]
- Zhu, S.; Yu, W.; Bi, L.; Qin, F.; Li, J.; Zeng, H.; Lu, L. Quercetin induces apoptosis of human breast cancer cells by activiting PTEN and inhibiting PI3K/AKT and JNK signaling pathways. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2022, 38, 714–720. [Google Scholar] [PubMed]
- Mohammadinejad, S.; Jafari-Gharabaghlou, D.; Zarghami, N. Development of PEGylated PLGA Nanoparticles Co-Loaded with Bioactive Compounds: Potential Anticancer Effect on Breast Cancer Cell Lines. Asian Pac. J. Cancer Prev. 2022, 23, 4063–4072. [Google Scholar] [CrossRef] [PubMed]
- Svitina, H.; Hamman, J.H.; Gouws, C. Molecular mechanisms and associated cell signalling pathways underlying the anticancer properties of phytochemical compounds from Aloe species (Review). Exp. Ther. Med. 2021, 22, 852. [Google Scholar] [CrossRef]
- Akhtar, M.F.; Saleem, A.; Rasul, A.; Faran Ashraf Baig, M.M.; Bin-Jumah, M.; Abdel Daim, M.M. Anticancer natural medicines: An overview of cell signaling and other targets of anticancer phytochemicals. Eur. J. Pharmacol. 2020, 888, 173488. [Google Scholar] [CrossRef]
- Mumtaz, P.T.; Bashir, S.M.; Rather, M.A.; Dar, K.B.; Taban, Q.; Sajood, S.; Ali, A.; Rather, Z.A.; Amin, I.; Dar, M.A. Antiproliferative and Apoptotic Activities of Natural Honey. In Therapeutic Applications of Honey and its Phytochemicals; Rehman, M.U., Majid, S., Eds.; Springer: Singapore, 2020; Volume 1, pp. 345–360. [Google Scholar]
- Boccellino, M.; Quagliuolo, L.; D’Angelo, S. Annurca Apple Biophenols’ Effects in Combination with Cisplatin on A549 Cells. Curr. Nutr. Food Sci. 2021, 17, 111–120. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Alsahli, M.A.; Almatroudi, A.; Verma, A.K.; Aloliqi, A.; Allemailem, K.S.; Khan, A.A.; Rahmani, A.H. Potential Therapeutic Targets of Quercetin, a Plant Flavonol, and Its Role in the Therapy of Various Types of Cancer through the Modulation of Various Cell Signaling Pathways. Molecules 2021, 26, 1315. [Google Scholar] [CrossRef]
- Kicinska, A.; Jarmuszkiewicz, W. Flavonoids and Mitochondria: Activation of Cytoprotective Pathways? Molecules 2020, 25, 3060. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, R.; Qian, J.; Sun, J.; Li, G.; Shen, J.; Xie, Y. Combination therapy of doxorubicin and quercetin on multidrug-resistant breast cancer and their sequential delivery by reduction-sensitive hyaluronic acid-based conjugate/d-α-tocopheryl poly (ethylene glycol) 1000 succinate mixed micelles. Mol. Pharm. 2020, 17, 1415–1427. [Google Scholar] [CrossRef] [PubMed]
- Kantapan, J.; Paksee, S.; Chawapun, P.; Sangthong, P.; Dechsupa, N. Pentagalloyl Glucose- and Ethyl Gallate-Rich Extract from Maprang Seeds Induce Apoptosis in MCF-7 Breast Cancer Cells through Mitochondria-Mediated Pathway. Evid. Based Complement. Altern. Med. 2020, 2020, 5686029. [Google Scholar] [CrossRef]
- Gupta, P.; Neupane, Y.R.; Aqil, M.; Kohli, K.; Sultana, Y. Lipid-based nanoparticle-mediated combination therapy for breast cancer management: A comprehensive review. Drug Deliv. Transl. Res. 2023, 13, 2739–2766. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Calderaro, A.; Patanè, G.T.; Navarra, M.; Barreca, D.; Cirmi, S.; Felice, M.R. Targets Involved in the Anti-Cancer Activity of Quercetin in Breast, Colorectal and Liver Neoplasms. Int. J. Mol. Sci. 2023, 24, 2952. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Raut, P.W.; Baruah, M.M.; Sharma, A. Combination of quercetin and 2-methoxyestradiol inhibits epithelial–mesenchymal transition in PC-3 cell line via Wnt signaling pathway. Future Sci. OA 2021, 7, FSO747. [Google Scholar] [CrossRef]
- Shi, Y.; Su, X.; Cui, H.; Yu, L.; Du, H.; Han, Y. Combination of quercetin and Adriamycin effectively suppresses the growth of refractory acute leukemia. Oncol. Lett. 2019, 18, 153–160. [Google Scholar] [CrossRef]
- Chen, K.T.J.; Anantha, M.; Leung, A.W.Y.; Kulkarni, J.A.; Militao, G.G.C.; Wehbe, M.; Sutherland, B.; Cullis, P.R.; Bally, M.B. Characterization of a liposomal copper(II)-quercetin formulation suitable for parenteral use. Drug Deliv. Transl. Res. 2020, 10, 202–215. [Google Scholar] [CrossRef]
- Jaisamut, P.; Wanna, S.; Limsuwan, S.; Chusri, S.; Wiwattanawongsa, K.; Wiwattanapatapee, R. Enhanced Oral Bioavailability and Improved Biological Activities of a Quercetin/Resveratrol Combination Using a Liquid Self-Microemulsifying Drug Delivery System. Planta Med. 2020, 87, 336–346. [Google Scholar] [CrossRef]
- Feriotto, G.; Tagliati, F.; Giriolo, R.; Casciano, F.; Tabolacci, C.; Beninati, S.; Khan, M.T.H.; Mischiati, C. Caffeic Acid Enhances the Anti-Leukemic Effect of Imatinib on Chronic Myeloid Leukemia Cells and Triggers Apoptosis in Cells Sensitive and Resistant to Imatinib. Int. J. Mol. Sci. 2021, 22, 1644. [Google Scholar] [CrossRef] [PubMed]
- Maleki Dana, P.; Sadoughi, F.; Asemi, Z.; Yousefi, B. The role of polyphenols in overcoming cancer drug resistance: A comprehensive review. Cell. Mol. Biol. Lett. 2022, 27, 1. [Google Scholar] [CrossRef] [PubMed]
- Mahbub, A.A.; Maitre, C.L.L.; Haywood-Small, S.; Cross, N.A.; Jordan-Mahy, N. Polyphenols enhance the activity of alkylating agents in leukaemia cell lines. Oncotarget 2019, 10, 4570–4586. [Google Scholar] [CrossRef][Green Version]
- Cione, E.; La Torre, C.; Cannataro, R.; Caroleo, M.C.; Plastina, P.; Gallelli, L. Quercetin, Epigallocatechin Gallate, Curcumin, and Resveratrol: From Dietary Sources to Human MicroRNA Modulation. Molecules 2020, 25, 63. [Google Scholar] [CrossRef] [PubMed]
- Doghish, A.S.; Hegazy, M.; Ismail, A.; El-Mahdy, H.A.; Elsakka, E.G.E.; Elkhawaga, S.Y.; Elkady, M.A.; Yehia, A.M.; Abdelmaksoud, N.M.; Mokhtar, M.M. A spotlight on the interplay of signaling pathways and the role of miRNAs in osteosarcoma pathogenesis and therapeutic resistance. Pathol. Res. Pract. 2023, 245, 154442. [Google Scholar] [CrossRef]
- Tavana, E.; Mollazadeh, H.; Mohtashami, E.; Modaresi, S.M.S.; Hosseini, A.; Sabri, H.; Soltani, A.; Javid, H.; Afshari, A.R.; Sahebkar, A. Quercetin: A promising phytochemical for the treatment of glioblastoma multiforme. BioFactors 2020, 46, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Rather, R.A.; Bhagat, M. Quercetin as an innovative therapeutic tool for cancer chemoprevention: Molecular mechanisms and implications in human health. Cancer Med. 2020, 9, 9181–9192. [Google Scholar] [CrossRef]
- Soofiyani, S.R.; Hosseini, K.; Forouhandeh, H.; Ghasemnejad, T.; Tarhriz, V.; Asgharian, P.; Reiner, Ž.; Sharifi-Rad, J.; Cho, W.C. Quercetin as a Novel Therapeutic Approach for Lymphoma. Oxidative Med. Cell. Longev. 2021, 2021, 3157867. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.; Dey, D.; Biswas, P.K.; Rahaman, T.I.; Saha, S.; Parvez, A.; Khan, D.A.; Lily, N.J.; Saha, K.; Sohel, M.; et al. A Comprehensive Analysis and Anti-Cancer Activities of Quercetin in ROS-Mediated Cancer and Cancer Stem Cells. Int. J. Mol. Sci. 2022, 23, 11746. [Google Scholar] [CrossRef]
- Li, W.; Cheng, M.; Zhang, W.; He, R.; Yang, H. New Insights into the Mechanisms of Polyphenol from Plum Fruit Inducing Apoptosis in Human Lung Cancer A549 Cells Via PI3K/AKT/FOXO1 Pathway. Plant Foods Hum. Nutr. 2021, 76, 125–132. [Google Scholar] [CrossRef]
- Vinayak, M.; Maurya, A.K. Quercetin Loaded Nanoparticles in Targeting Cancer: Recent Development. Anti-Cancer Agents Med. Chem. Anti-Cancer Agents 2019, 19, 1560–1576. [Google Scholar] [CrossRef]
- Zhang, C.; Hao, Y.; Sun, Y.; Liu, P. Quercetin suppresses the tumorigenesis of oral squamous cell carcinoma by regulating microRNA-22/WNT1/β-catenin axis. J. Pharmacol. Sci. 2019, 140, 128–136. [Google Scholar] [CrossRef]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A review on anti-cancer properties of Quercetin in breast cancer. Life Sci. 2020, 248, 117463. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, J.; Deng, Y.; Liao, L.; Zhou, M.; Peng, C.; Li, Y. Quercetin as a protective agent for liver diseases: A comprehensive descriptive review of the molecular mechanism. Phytother. Res. 2021, 35, 4727–4747. [Google Scholar] [CrossRef] [PubMed]
- Rizzotto, D.; Englmaier, L.; Villunger, A. At a Crossroads to Cancer: How p53-Induced Cell Fate Decisions Secure Genome Integrity. Int. J. Mol. Sci. 2021, 22, 10883. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, S.-D.; Jiao, J.; Wang, W.; Yu, L.; Zhao, X.-L.; Wang, L.-T.; Meng, D.; Fu, Y.-J. ROS -mediated p53 activation by juglone enhances apoptosis and autophagy in vivo and in vitro. Toxicol. Appl. Pharmacol. 2019, 379, 114647. [Google Scholar] [CrossRef] [PubMed]
- Aziz, Y.M.A.; Lotfy, G.; Said, M.M.; El Ashry, E.S.H.; El Tamany, E.S.H.; Soliman, S.M.; Abu-Serie, M.M.; Teleb, M.; Yousuf, S.; Dömling, A.; et al. Design, Synthesis, Chemical and Biochemical Insights Into Novel Hybrid Spirooxindole-Based p53-MDM2 Inhibitors with Potential Bcl2 Signaling Attenuation. Front. Chem. 2021, 9, 735236. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Lee, J.; Moon, H.; Ryu, C.H.; Seok, J.; Jung, Y.-S.; Ryu, J.; Baek, S.J. Quercetin Induces Anticancer Activity by Upregulating Pro-NAG-1/GDF15 in Differentiated Thyroid Cancer Cells. Cancers 2021, 13, 3022. [Google Scholar] [CrossRef]
- Tisi, R.; Gaponenko, V.; Vanoni, M.; Sacco, E. Natural Products Attenuating Biosynthesis, Processing, and Activity of Ras Oncoproteins: State of the Art and Future Perspectives. Biomolecules 2020, 10, 1535. [Google Scholar] [CrossRef]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, X.; Zhu, G.; Liu, H.; Chen, J.; Wang, Y.; He, X. Quercetin inhibits TNF-α induced HUVECs apoptosis and inflammation via downregulating NF-kB and AP-1 signaling pathway in vitro. Medicine 2020, 99, e22241. [Google Scholar] [CrossRef]
- Sul, O.J.; Ra, S.W. Quercetin Prevents LPS-Induced Oxidative Stress and Inflammation by Modulating NOX2/ROS/NF-kB in Lung Epithelial Cells. Molecules 2021, 26, 6949. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ding, H.; Tang, X.; Liang, M.; Li, S.; Zhang, J.; Cao, J. Quercetin induces pro-apoptotic autophagy via SIRT1/AMPK signaling pathway in human lung cancer cell lines A549 and H1299 in vitro. Thorac. Cancer 2021, 12, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Bayda, S.; Adeel, M.; Tuccinardi, T.; Cordani, M.; Rizzolio, F. The History of Nanoscience and Nanotechnology: From Chemical–Physical Applications to Nanomedicine. Molecules 2020, 25, 112. [Google Scholar] [CrossRef] [PubMed]
- Feynman, R. There’s Plenty of Room at the Bottom. In Feynman and Computation; CRC Press: Boca Raton, FL, USA, 2018; pp. 63–76. [Google Scholar]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [PubMed]
- McGuckin, M.B.; Wang, J.; Ghanma, R.; Qin, N.; Palma, S.D.; Donnelly, R.F.; Paredes, A.J. Nanocrystals as a master key to deliver hydrophobic drugs via multiple administration routes. J. Control. Release 2022, 345, 334–353. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Cheng, M.; Zhang, X.; Chen, X. Quercetin nanoformulations: A promising strategy for tumor therapy. Food Funct. 2021, 12, 6664–6681. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Li, M.; Dey, R.; Chen, Y. Nanomaterials for cancer therapy: Current progress and perspectives. J. Hematol. Oncol. 2021, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Patel, G.; Thakur, N.S.; Kushwah, V.; Patil, M.D.; Nile, S.H.; Jain, S.; Kai, G.; Banerjee, U.C. Mycophenolate co-administration with quercetin via lipid-polymer hybrid nanoparticles for enhanced breast cancer management. Nanomed. Nanotechnol. Biol. Med. 2020, 24, 102147. [Google Scholar] [CrossRef]
- Zhou, H.; Yuan, Y.; Wang, Z.; Ren, Z.; Hu, M.; Lu, J.; Gao, H.; Pan, C.; Zhao, W.; Zhu, B. Co-delivery of doxorubicin and quercetin by Janus hollow silica nanomotors for overcoming multidrug resistance in breast MCF-7/Adr cells. Colloids Surf. A Physicochem. Eng. Asp. 2023, 658, 130654. [Google Scholar] [CrossRef]
- Pinheiro, R.G.R.; Pinheiro, M.; Neves, A.R. Nanotechnology Innovations to Enhance the Therapeutic Efficacy of Quercetin. Nanomaterials 2021, 11, 2658. [Google Scholar] [CrossRef]
- Manzoor, M.F.; Hussain, A.; Sameen, A.; Sahar, A.; Khan, S.; Siddique, R.; Aadil, R.M.; Xu, B. Novel extraction, rapid assessment and bioavailability improvement of quercetin: A review. Ultrason. Sonochemistry 2021, 78, 105686. [Google Scholar] [CrossRef]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Fallahi, F.; Taghavipour, M.; Ghasemi, Y.; Akbari, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; et al. Quercetin and cancer: New insights into its therapeutic effects on ovarian cancer cells. Cell Biosci. 2020, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Nasirizadeh, S.; Malaekeh-Nikouei, B. Solid lipid nanoparticles and nanostructured lipid carriers in oral cancer drug delivery. J. Drug Deliv. Sci. Technol. 2020, 55, 101458. [Google Scholar] [CrossRef]
- Marques, M.P.; Varela, C.; Mendonça, L.; Cabral, C. Nanotechnology-Based Topical Delivery of Natural Products for the Management of Atopic Dermatitis. Pharmaceutics 2023, 15, 1724. [Google Scholar] [CrossRef] [PubMed]
- Molani Gol, R.; Kheirouri, S. The Effects of Quercetin on the Apoptosis of Human Breast Cancer Cell Lines MCF-7 and MDA-MB-231: A Systematic Review. Nutr. Cancer 2022, 74, 405–422. [Google Scholar] [CrossRef] [PubMed]
- Lohan, S.; Sharma, T.; Saini, S.; Singh, A.; Kumar, A.; Raza, K.; Kaur, J.; Singh, B. Galactosylated nanoconstructs of Berberine with enhanced Biopharmaceutical and cognitive potential: A preclinical evidence in Alzheimer ‘s disease. J. Drug Deliv. Sci. Technol. 2021, 66, 102695. [Google Scholar] [CrossRef]
- Guo, S.; Liang, Y.; Liu, L.; Yin, M.; Wang, A.; Sun, K.; Li, Y.; Shi, Y. Research on the fate of polymeric nanoparticles in the process of the intestinal absorption based on model nanoparticles with various characteristics: Size, surface charge and pro-hydrophobics. J. Nanobiotechnol. 2021, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Das, S.S.; Verma, P.R.P.; Kar, S.; Singh, S.K. Quercetin-Loaded Nanomedicine as Oncotherapy. In Nanomedicine for Bioactives: Healthcare Applications; Rahman, M., Beg, S., Kumar, V., Ahmad, F.J., Eds.; Springer: Singapore, 2020; pp. 155–183. [Google Scholar]
- Gu, L.-Q.; Cui, P.-F.; Xing, L.; He, Y.-J.; Chang, X.; Zhou, T.-J.; Liu, Y.; Li, L.; Jiang, H.-L. An energy-blocking nanoparticle decorated with anti-VEGF antibody to reverse chemotherapeutic drug resistance. RSC Adv. 2019, 9, 12110–12123. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Xu, X.; Liu, J.; Jia, Q.; Ke, C.; Zhang, H.; Xu, C.; Ou, E.; Tan, W.; Zhao, Y. Mitochondria-Targeted Triphenylphosphonium Conjugated C-3 Modified Betulin: Synthesis, Antitumor Properties and Mechanism of Action. ChemMedChem 2022, 17, e202100659. [Google Scholar] [CrossRef]
- Rezvantalab, S.; Drude, N.I.; Moraveji, M.K.; Güvener, N.; Koons, E.K.; Shi, Y.; Lammers, T.; Kiessling, F. PLGA-Based Nanoparticles in Cancer Treatment. Front. Pharmacol. 2018, 9, 1260. [Google Scholar] [CrossRef]
- Ersoz, M.; Erdemir, A.; Derman, S.; Arasoglu, T.; Mansuroglu, B. Quercetin-loaded nanoparticles enhance cytotoxicity and antioxidant activity on C6 glioma cells. Pharm. Dev. Technol. 2020, 25, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Jha, N.K.; Arfin, S.; Jha, S.K.; Kar, R.; Dey, A.; Gundamaraju, R.; Ashraf, G.M.; Gupta, P.K.; Dhanasekaran, S.; Abomughaid, M.M.; et al. Re-establishing the comprehension of phytomedicine and nanomedicine in inflammation-mediated cancer signaling. Semin. Cancer Biol. 2022, 86, 1086–1104. [Google Scholar] [CrossRef] [PubMed]
- Gulin-Sarfraz, T.; Kalantzopoulos, G.N.; Kvalvåg Pettersen, M.; Wold Åsli, A.; Tho, I.; Axelsson, L.; Sarfraz, J. Inorganic Nanocarriers for Encapsulation of Natural Antimicrobial Compounds for Potential Food Packaging Application: A Comparative Study. Nanomaterials 2021, 11, 379. [Google Scholar] [CrossRef]
- Jafari, S.; Derakhshankhah, H.; Alaei, L.; Fattahi, A.; Varnamkhasti, B.S.; Saboury, A.A. Mesoporous silica nanoparticles for therapeutic/diagnostic applications. Biomed. Pharmacother. 2019, 109, 1100–1111. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Manna, K.; Kayal, U.; Saha, M.; Chatterjee, S.; Chandra, D.; Hara, M.; Datta, S.; Bhaumik, A.; Das Saha, K. Folic acid-conjugated magnetic mesoporous silica nanoparticles loaded with quercetin: A theranostic approach for cancer management. RSC Adv. 2020, 10, 23148–23164. [Google Scholar] [CrossRef]
- Zaharudin, N.S.; Mohamed Isa, E.D.; Ahmad, H.; Abdul Rahman, M.B.; Jumbri, K. Functionalized mesoporous silica nanoparticles templated by pyridinium ionic liquid for hydrophilic and hydrophobic drug release application. J. Saudi Chem. Soc. 2020, 24, 289–302. [Google Scholar] [CrossRef]
- Otieno, E.; Huang, Y.; Li, N.; Li, T.; Wang, M.; Qiu, X.; Xiao, X. Utilization of superparamagnetic iron oxide nanoparticles (SPIONs) as a vector for drug delivery. Appl. Nanosci. 2023, 13, 6191–6216. [Google Scholar] [CrossRef]
- Zandieh, M.A.; Farahani, M.H.; Daryab, M.; Motahari, A.; Gholami, S.; Salmani, F.; Karimi, F.; Samaei, S.S.; Rezaee, A.; Rahmanian, P.; et al. Stimuli-responsive (nano)architectures for phytochemical delivery in cancer therapy. Biomed. Pharmacother. 2023, 166, 115283. [Google Scholar] [CrossRef]
- Ghosh, M.; Godderis, L.; Hoet, P. Epigenetic Mechanisms in Understanding Nanomaterial-Induced Toxicity. In Nanotoxicology in Safety Assessment of Nanomaterials; Louro, H., Silva, M.J., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 195–223. [Google Scholar]
- Kondath, S.; Rajaram, R.; Anantanarayanan, R. Curcumin reduced gold nanoparticles synergistically induces ROS mediated apoptosis in MCF-7 cancer cells. Inorg. Nano-Met. Chem. 2020, 51, 601–613. [Google Scholar] [CrossRef]
- Raghav, A.; Giri, R.; Agarwal, S.; Kala, S.; Jeong, G.-B. Protective role of engineered extracellular vesicles loaded quercetin nanoparticles as anti-viral therapy against SARS-CoV-2 infection: A prospective review. Front. Immunol. 2022, 13, 1040027. [Google Scholar] [CrossRef]
- Qi, Y.; Guo, L.; Jiang, Y.; Shi, Y.; Sui, H.; Zhao, L. Brain delivery of quercetin-loaded exosomes improved cognitive function in AD mice by inhibiting phosphorylated Tau-mediated neurofibrillary tangles. Drug Deliv. 2020, 27, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Huang, Z.; Huang, L.; Liang, J.; Wang, P.; Zhao, L.; Shi, Y. Surface-modified engineered exosomes attenuated cerebral ischemia/reperfusion injury by targeting the delivery of quercetin towards impaired neurons. J. Nanobiotechnol. 2021, 19, 141. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Quezada, J.; Guajardo-Flores, D.; González-Valdez, J. Enhanced exosome-mediated delivery of black bean phytochemicals (Phaseolus vulgaris L.) for cancer treatment applications. Biomed. Pharmacother. 2020, 131, 110771. [Google Scholar] [CrossRef] [PubMed]




| Food Source | Quercetin Content (mg/100 g) |
|---|---|
| FRUITS | |
| Apple with skin (Malus domestica) | 4.42 |
| Acerola (Malpighia emarginata) | 4.74 |
| Arctic bramble (Rubus arcticus) | 9.1 |
| Blueberry (Vaccinium caesariense) | 7.67 |
| Cranberry (Vaccinium macrocarpon) | 14.84 |
| Elderberry (Sambucus spp.) | 26.77 |
| Fig (Ficus carica) | 5.47 |
| Plum (Prunus domestica) | 12.45 |
| Sea buckthorn (Hippophae rhamnoides) | 7.4 |
| Wolfberry (Lycium barbarum) | 13.6 |
| Common juniper (Juniperus communis) | 46.61 |
| Prickly pear (Opuntia spp.) | 4.86 |
| VEGETABLE PRODUCTS | |
| Sowthistle (Sonchus oleraceus) | 16 |
| Arugula (Eruca sativa) | 7.92 |
| Sparrow grass (Asparagus officinalis) | 13.98 |
| Swiss chard (Beta vulgaris) | 7.5 |
| Green chicory (Cichorium intybus) | 6.49 |
| Coriander (Coriandrum sativum) | 52.9 |
| Golden poppy (Eschscholzia californica) | 26.3 |
| Drumstick tree (Moringa oleifera) | 16.65 |
| Fennel (Foeniculum vulgare) | 48.80 |
| Leaf cabbage (Brassica oleracea) | 7.71 |
| Red lettuce (Lactuca sativa) | 7.61 |
| Mustard greens (Brassica juncea) | 8.8 |
| Okra (Abelmoschus esculentus) | 20.97 |
| Onions (Allium cepa) | 20.3 |
| Perennial wall-rocket (Diplotaxis tenuifolia) | 66.19 |
| New Mexico chile (Capsicum annuum) | 15 |
| Sweet potato (Ipomoea batatas) | 16.94 |
| SPICES AND HERBS | |
| Caper bush (Capparis spinosa) | 180.77 |
| Dill (Anethum graveolens) | 55.15 |
| Oregano (Origanum vulgare) | 7.3 |
| Tarragon (Artemisia dracunculus) | 11 |
| Turmeric (Curcuma longa) | 4.92 |
| Buckwheat (Fagopyrum esculentum) | 15.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrillo-Martinez, E.J.; Flores-Hernández, F.Y.; Salazar-Montes, A.M.; Nario-Chaidez, H.F.; Hernández-Ortega, L.D. Quercetin, a Flavonoid with Great Pharmacological Capacity. Molecules 2024, 29, 1000. https://doi.org/10.3390/molecules29051000
Carrillo-Martinez EJ, Flores-Hernández FY, Salazar-Montes AM, Nario-Chaidez HF, Hernández-Ortega LD. Quercetin, a Flavonoid with Great Pharmacological Capacity. Molecules. 2024; 29(5):1000. https://doi.org/10.3390/molecules29051000
Chicago/Turabian StyleCarrillo-Martinez, Eber Josue, Flor Yohana Flores-Hernández, Adriana María Salazar-Montes, Hector Fabián Nario-Chaidez, and Luis Daniel Hernández-Ortega. 2024. "Quercetin, a Flavonoid with Great Pharmacological Capacity" Molecules 29, no. 5: 1000. https://doi.org/10.3390/molecules29051000
APA StyleCarrillo-Martinez, E. J., Flores-Hernández, F. Y., Salazar-Montes, A. M., Nario-Chaidez, H. F., & Hernández-Ortega, L. D. (2024). Quercetin, a Flavonoid with Great Pharmacological Capacity. Molecules, 29(5), 1000. https://doi.org/10.3390/molecules29051000