

Enhancement of Female Rat Fertility via Ethanolic Extract from Nigella sativa L. (Black Cumin) Seeds Assessed via HPLC-ESI-MS/MS and Molecular Docking
 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Qualitative Characterization of Phytoconstituents in N. sativa seed Ethanol Fractional Extract through HPLC/ESI-MS/MS Analysis
2.1.1. Alkaloids
2.1.2. Saponins
2.1.3. Monoterpenes
2.1.4. Phenolic Acids
2.1.5. Fatty Acids
2.2. Biological Results
2.2.1. Evaluation of FSH-like Activity
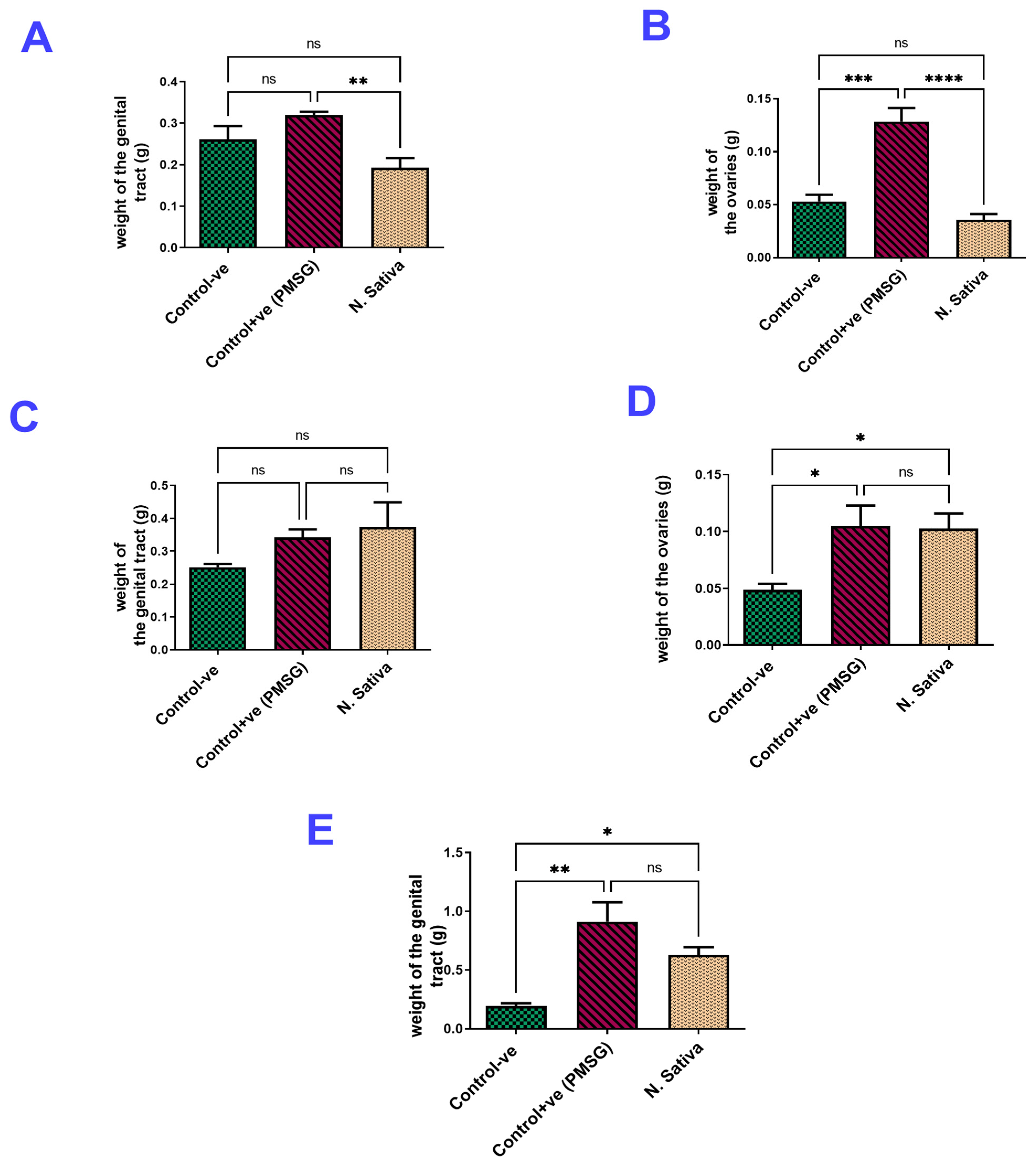
Ovarian and Uterine Weights
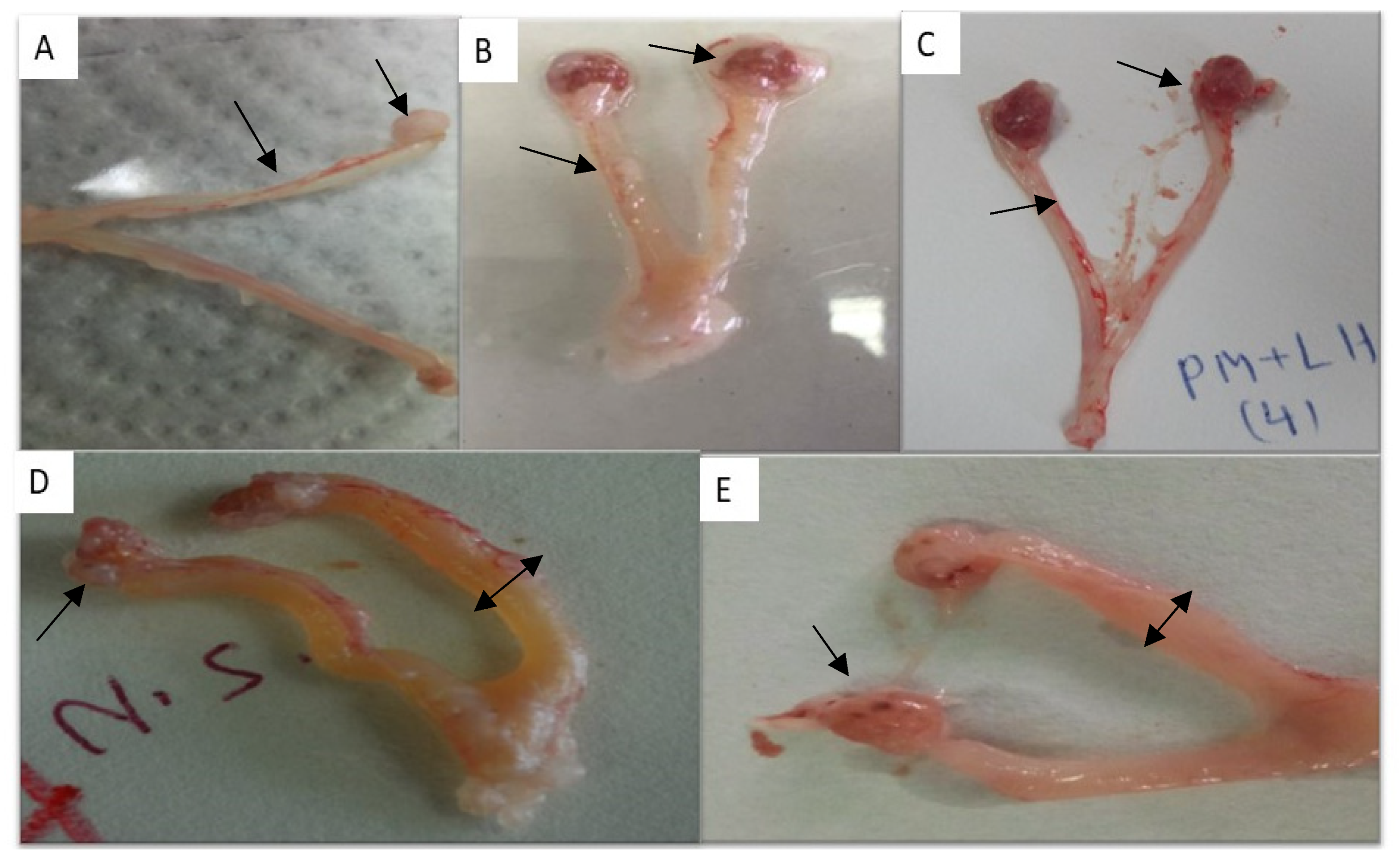
Morphological Examination of Genitalia
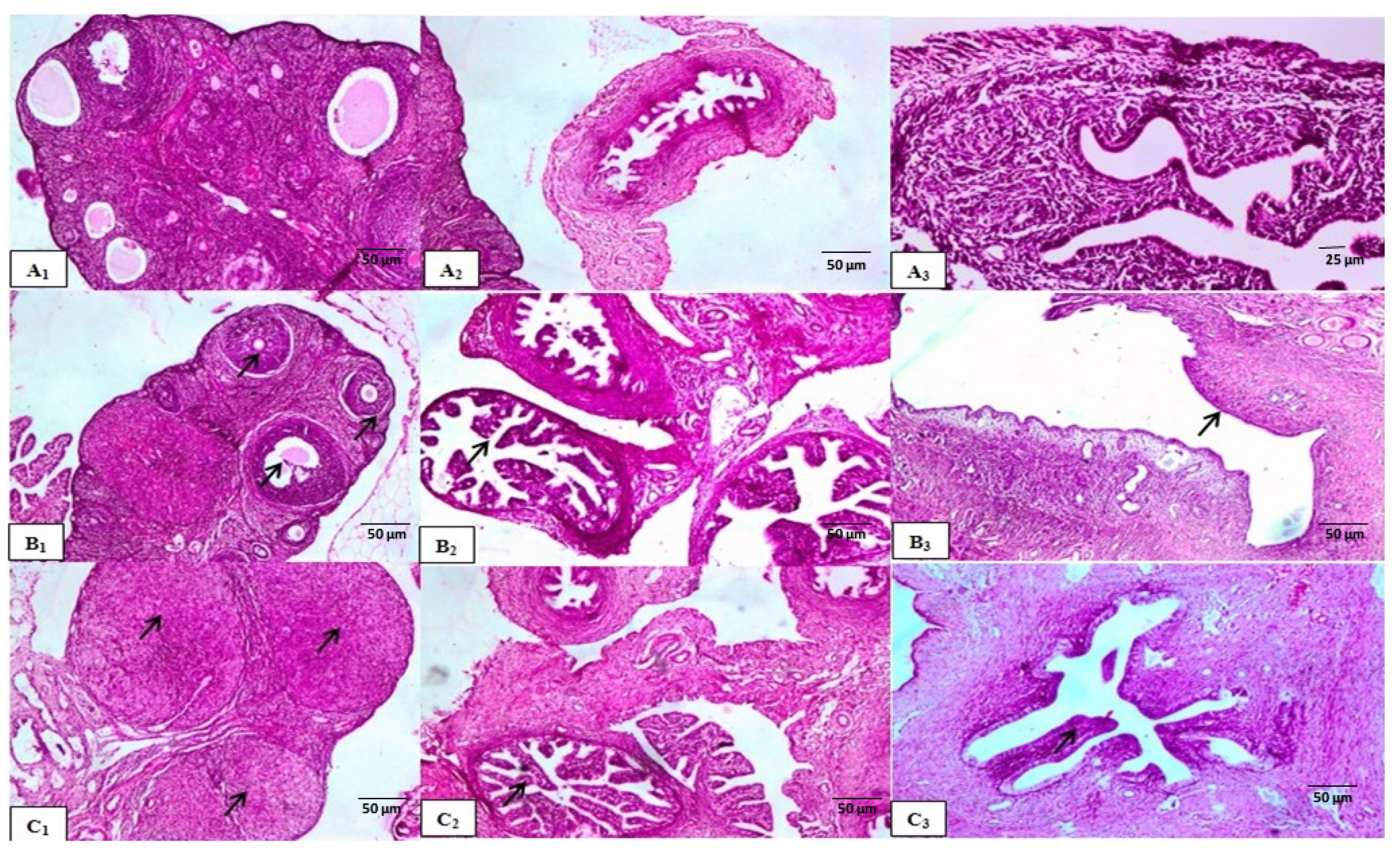
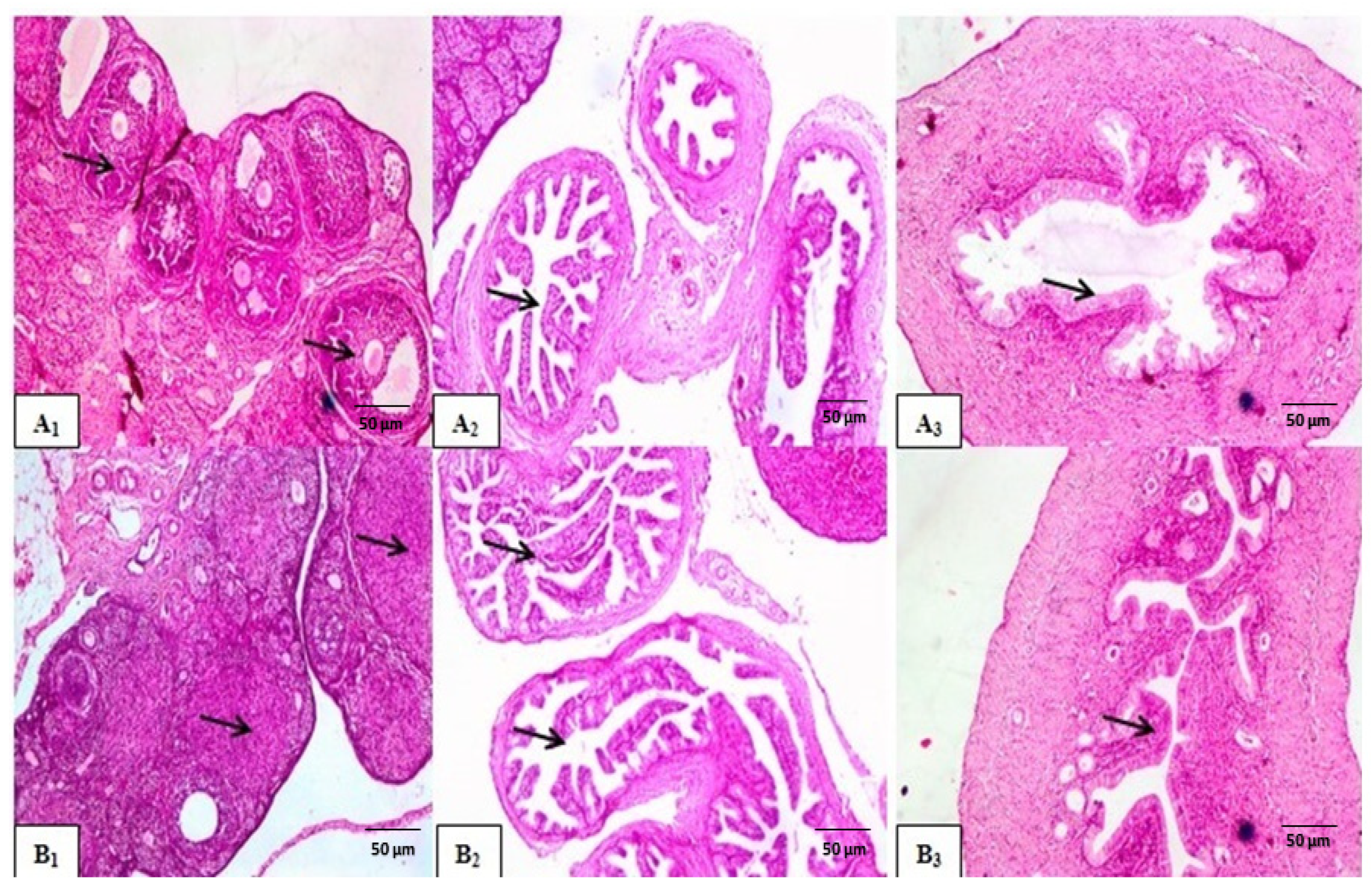
Histological Examination
2.2.2. Evaluation of LH-like Activity
Ovarian and Uterine Weights
Morphological Examination of Genitalia
Histological Examination
2.2.3. Evaluation of Estrogen-like Activity
Ovarian and Uterine Weights
Morphological Examination of Genitalia
Histological Examination
Determination of Serum Hormones
Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction Processes
4.3. HPLC-ESI-MS/MS Analysis
4.4. Biological Assays
4.4.1. Chemicals, Drugs, and Kits
4.4.2. Experimental Animals
4.4.3. Preliminary Study
4.4.4. FSH-like Activity
4.4.5. LH-like Activity
4.4.6. Estrogenic Activity (Ovariectomized Rats)
4.4.7. Blood Sample Collection and Morphological and Metric Assessment of the Reproductive Tract
- The Rat FSH ELISA Kit from Wuhan Fine Biotech in Wuhan, Hubei, China was used to measure FSH levels.
- LH levels were measured using an LH ELISA kit (Ref. no. E29-118, Lot. Number: 117041404) from IMMUNOSPEC in Seattle, Washington, DC, USA.
- An Estradiol (E2) ELISA kit (Catalog No. 10009, Lot. Number: 118031903) from BIOS, Microwell Diagnostic Systems Chemux Bioscience, Inc. in the Netherlands was used to measure estradiol levels.
- Progesterone levels were determined using an ELISA kit (Ref. no. 10005, Lot. Number: 118042304) from Chemux BioScience, INC in South San Francisco, CA, USA.
4.4.8. Histological Examination
4.5. Molecular Docking Studies
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petok, W.D. Infertility Counseling (or the Lack Thereof) of the Forgotten Male Partner. Fertil. Steril. 2015, 104, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Adriaens, I.; Cortvrindt, R.; Smitz, J. Differential FSH Exposure in Preantral Follicle Culture Has Marked Effects on Folliculogenesis and Oocyte Developmental Competence. Hum. Reprod. 2004, 19, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Hammam, A.M.; El-Gendy, A.M.; Nagya, A.M.; Elkhadrawy, H.H.; Zaki, M.S.; Ali, A.H. Extraction and Purification of Buffalo Pituitary FSH (BuFSH) with Emphasis on Its Biological Activity and Histological Changes in Ovaries of Mice Hammam. Life Sci. J. 2014, 11, 1258–1267. [Google Scholar]
- Hammam, A.; Mohamed, A.N.; Ahmed, M.M.; Kandeil, A.M.G. Preparation of Biologically Active Recombinant Buffalo Follicle Stimulating Hormone (RbuFSH) from Buffalo Pituitaries. Egypt. J. Vet. Sci. 2018, 49, 91–102. [Google Scholar] [CrossRef]
- Fernandez-Valdivia, R.; Jeong, J.; Mukherjee, A.; Soyal, S.M.; Li, J.; Ying, Y.; DeMayo, F.J.; Lydon, J.P. A Mouse Model to Dissect Progesterone Signaling in the Female Reproductive Tract and Mammary Gland. Genesis 2010, 48, 106–113. [Google Scholar] [CrossRef]
- Conneely, O.M. Progesterone Receptors and Ovulation. Handb. Exp. Pharmacol. 2010, 198, 37–44. [Google Scholar] [CrossRef]
- Nagy, A.M.; Sathe, S.R.; Atta, A.H.; Hammam, A.M.M.; Hsu, W.H. Characterization of Nuclear Progesterone Receptor Isoforms in the Term Equine Placenta. Front. Vet. Sci. 2021, 8, 660177. [Google Scholar] [CrossRef]
- Parhizkar, S.; Latiff, L.A.; Rahman, S.A.; Dollah, M.A.; Parichehr, H. Assessing Estrogenic Activity of Nigella sativa in Ovariectomized Rats Using Vaginal Cornification Assay. Afr. J. Pharm. Pharmacol. 2011, 5, 137–142. [Google Scholar] [CrossRef]
- Aziz, M.A.; Khan, A.H.; Ullah, H.; Adnan, M.; Hashem, A.; Abd Allah, E.F. Traditional Phytomedicines for Gynecological Problems Used by Tribal Communities of Mohmand Agency near the Pak-Afghan Border Area. Rev. Bras. Farmacogn. 2018, 28, 503–511. [Google Scholar] [CrossRef]
- Leisegang, K.; Almaghrawi, W.; Henkel, R. Biomedicine & Pharmacotherapy The Effect of Nigella sativa Oil and Metformin on Male Seminal Parameters and Testosterone in Wistar Rats Exposed to an Obesogenic Diet. Biomed. Pharmacother. 2021, 133, 111085. [Google Scholar] [CrossRef]
- Farzaneh, E.; Nia, F.R.; Mehrtash, M.; Mirmoeini, F.S.; Jalilvand, M. The Effects of 8-Week Nigella sativa Supplementation and Aerobic Training on Lipid Profile and VO2 Max in Sedentary Overweight Females. Int. J. Prev. Med. 2014, 5, 210–216. [Google Scholar] [PubMed]
- Kooti, W.; Hasanzadeh-Noohi, Z.; Sharafi-Ahvazi, N.; Asadi-Samani, M.; Ashtary-Larky, D. Phytochemistry, Pharmacology, and Therapeutic Uses of Black Seed (Nigella sativa). Chin. J. Nat. Med. 2016, 14, 732–745. [Google Scholar] [CrossRef]
- Gilani, A.-H.; Jabeen, Q.; Khan, M.A.U. A Review of Medicinal Uses and Pharmacological Activities of of Nigella sativa. Pak. J. Biol. Sci. 2004, 7, 441–451. [Google Scholar]
- Forouzanfar, F.; Bazzaz, B.S.F.; Hosseinzadeh, H. Black Cumin (Nigella sativa) and Its Constituent (Thymoquinone): A Review on Antimicrobial Effects. Iran. J. Basic Med. Sci. 2014, 17, 929–938. [Google Scholar]
- Ahmad, M.F.; Ahmad, F.A.; Ashraf, S.A.; Saad, H.H.; Wahab, S.; Khan, M.I.; Ali, M.; Mohan, S.; Hakeem, K.R.; Athar, M.T. An Updated Knowledge of Black Seed (Nigella sativa Linn.): Review of Phytochemical Constituents and Pharmacological Properties. J. Herb. Med. 2021, 25, 100404. [Google Scholar] [CrossRef]
- Anaeigoudari, A.; Safari, H.; Khazdair, M.R. Effects of Nigella sativa, Camellia Sinensis, and Allium Sativum as Food Additives on Metabolic Disorders, a Literature Review. Front. Pharmacol. 2021, 12, 762182. [Google Scholar] [CrossRef]
- Hannan, M.A.; Rahman, M.A.; Sohag, A.A.M.; Uddin, M.J.; Dash, R.; Sikder, M.H.; Rahman, M.S.; Timalsina, B.; Munni, Y.A.; Sarker, P.P.; et al. Black Cumin (Nigella sativa L.): A Comprehensive Review on Phytochemistry, Health Benefits, Molecular Pharmacology, and Safety. Nutrients 2021, 13, 1784. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Xu, Q.; Ma, J.; Li, X.; Yan, J.; Tian, Y.; Wen, Y.; Chen, T. Nigella sativa and Health Outcomes: An Overview of Systematic Reviews and Meta-Analyses. Front. Nutr. 2023, 10, 1107750. [Google Scholar] [CrossRef]
- Rashwan, H.K.; Mahgoub, S.; Abuelezz, N.Z.; Amin, H.K. Black Cumin Seed (Nigella sativa) in Inflammatory Disorders: Therapeutic Potential and Promising Molecular Mechanisms. Drugs Drug Candidates 2023, 2, 516–537. [Google Scholar] [CrossRef]
- Ciesielska-Figlon, K.; Wojciechowicz, K.; Wardowska, A.; Lisowska, K.A. The Immunomodulatory Effect of Nigella sativa. Antioxidants 2023, 12, 1340. [Google Scholar] [CrossRef]
- Gholamnezhad, Z.; Keyhanmanesh, R.; Boskabady, M.H. Anti-Inflammatory, Antioxidant, and Immunomodulatory Aspects of Nigella sativa for Its Preventive and Bronchodilatory Effects on Obstructive Respiratory Diseases: A Review of Basic and Clinical Evidence. J. Funct. Foods 2015, 17, 910–927. [Google Scholar] [CrossRef]
- Gholamnezhad, Z.; Havakhah, S.; Boskabady, M.H. Preclinical and Clinical Effects of Nigella sativa and Its Constituent, Thymoquinone: A Review. J. Ethnopharmacol. 2016, 190, 372–386. [Google Scholar] [CrossRef] [PubMed]
- Dajani, E.Z.; Shahwan, T.G.; Dajani, N.E. Overview of the Preclinical Pharmacological Properties of Nigella sativa (Black Seeds): A Complementary Drug with Historical and Clinical Significance. J. Physiol. Pharmacol. 2016, 67, 801–817. [Google Scholar] [PubMed]
- El-Far, A.H.; Korshom, M.A.; Mandour, A.A.; El-Bessoumy, A.A.; El-Sayed, Y.S. Hepatoprotective Efficacy of Nigella sativa Seeds Dietary Supplementation against Lead Acetate-Induced Oxidative Damage in Rabbit—Purification and Characterization of Glutathione Peroxidase. Biomed. Pharmacother. 2017, 89, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Gad, H.A.; Heiss, A.G.; Wessjohann, L.A. Metabolomics Driven Analysis of Six Nigella Species Seeds via UPLC-QTOF-MS and GC–MS Coupled to Chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; El Sayed, A.M.; El Banna, A.; Ruehmann, S. Metabolomics Reveals Distinct Methylation Reaction in MeJA Elicited Nigella sativa Callus via UPLC–MS and Chemometrics. Plant Cell Tissue Organ Cult. 2015, 122, 453–463. [Google Scholar] [CrossRef]
- Alta’ee, A.H.; Ewadh, M.J.; Zaidan, H.K. Hormonal Contents of Two Types of Black Seed (Nigella sativa) Oil: Comparative Study. Med. J. Babylon 2006, 3, 17–21. [Google Scholar]
- Ali, Z.; Ferreira, D.; Carvalho, P.; Avery, M.A.; Khan, I.A. Nigellidine-4-O-Sulfite, the First Sulfated Indazole-Type Alkaloid from the Seeds of Nigella sativa. J. Nat. Prod. 2008, 71, 1111–1112. [Google Scholar] [CrossRef]
- Pham, H.N.; Tran, C.A.; Trinh, T.D.; Nguyen Thi, N.L.; Tran Phan, H.N.; Le, V.N.; Le, N.H.; Phung, V.T. UHPLC-Q-TOF-MS/MS Dereplication to Identify Chemical Constituents of Hedera Helix Leaves in Vietnam. J. Anal. Methods Chem. 2022, 8, 1167265. [Google Scholar] [CrossRef]
- Kadam, D.; Lele, S.S. Extraction, Characterization and Bioactive Properties of Nigella sativa Seedcake. J. Food Sci. Technol. 2017, 54, 3936–3947. [Google Scholar] [CrossRef]
- Avula, B.; Wang, Y.; Ali, Z.; Smillie, T.; Khan, I. Quantitative Determination of Chemical Constituents from Seeds of Nigella sativa L. by Using HPLC-UV and Identification by LC-ESI-TOF. Planta Med. 2009, 93, 1778–1787. [Google Scholar] [CrossRef]
- Venkatachallam, S.K.T.; Pattekhan, H.; Divakar, S.; Kadimi, U.S. Chemical Composition of Nigella sativa L. Seed Extracts Obtained by Supercritical Carbon Dioxide. J. Food Sci. Technol. 2010, 47, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Heitman, L.H.; Oosterom, J.; Bonger, K.M.; Timmers, C.M.; Wiegerinck, P.H.G.; IJzerman, A.P. [3H]Org 43553, the First Low-Molecular-Weight Agonistic and Allosteric Radioligand for the Human Luteinizing Hormone Receptor. Mol. Pharmacol. 2008, 73, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Sahu, P. Plants Altering Hormonal Milieu: A Review. Asian Pac. J. Reprod. 2017, 6, 49–53. [Google Scholar] [CrossRef]
- Jasim, W.K.; Hassan, M.S.; Keam, G.G. Study the Effect of Nigella sativa on Thyroid Function and Reproductive Hormone of Female Rat. J. Contemp. Med. Sci. 2016, 2, 67–69. [Google Scholar]
- Juma, F.T.; Abdulrahman, H.M.A. The Effects of Nigella sativa Oil Administration on Some Physiological and Histological Values of Reproductive Aspects of Rats. Iraqi J. Vet. Med. 2011, 35, 52–60. [Google Scholar] [CrossRef]
- Boukhliq, R.; Martin, G.B.; White, C.L.; Blackberry, M.A.; Murray, P.J. Role of Glucose, Fatty Acids and Protein in Regulation of Testicular Growth and Secretion of Gonadotrophin, Prolactin, Somatotrophin and Insulin in the Mature Ram. Reprod. Fertil. Dev. 1997, 9, 515–524. [Google Scholar] [CrossRef]
- Garrel, G.; Simon, V.; Denoyelle, C.; Ishaq, M.; Rouch, C.; Dairou, J.; Magnan, C.; Migrenne, S.; Cruciani-guglielmacci, C.; Cohen-tannoudji, J. Unsaturated Fatty Acids Disrupt Smad Signaling in Gonadotrope Cells Leading to Inhibition of FSHβ Gene Expression. Endocrinology 2014, 155, 592–604. [Google Scholar] [CrossRef]
- Garrel, G.; Simon, V.; Denoyelle, C.; Counis, R.; Magnan, C. Unsaturated Fatty Acids Stimulate LH Secretion via Novel PKCε and −θ in Gonadotrope Cells and Inhibit GnRH-Induced LH Release. Endocrinology 2011, 152, 3905–3916. [Google Scholar] [CrossRef]
- Moriyama, R.; Yamazaki, T.; Kato, T.; Kato, Y. Long-Chain Unsaturated Fatty Acids Reduce the Transcriptional Activity of the Rat Follicle-Stimulating Hormone β-Subunit Gene. J. Reprod. Dev. 2016, 62, 195–199. [Google Scholar] [CrossRef]
- Holt, E.H.; Lupsa, B.; Lee, G.S.; Bassyouni, H.; Peery, H.E. Hormonal Control of Pregnancy and Lactation. In Goodman’s Basic Medical Endocrinology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 473–527. [Google Scholar]
- Parhizkar, S.; Latiff, L.A.; Parsa, A. Effect of Nigella sativa on Reproductive System in Experimental. Avicenna J. Phytomed. 2016, 6, 95–103. [Google Scholar]
- Al-Saffar, R.A.S.; Al-Wiswasy, M.K.M. A Morphological Study of the Pharmacological Effects of the Nigella sativa on the Reproductive System in Experimental Rats. Jordan J. Biol. Sci. 2019, 12, 147–153. [Google Scholar]
- Piras, A.; Rosa, A.; Marongiu, B.; Porcedda, S.; Falconieri, D.; Dessì, M.A.; Ozcelik, B.; Koca, U. Chemical Composition and in Vitro Bioactivity of the Volatile and Fixed Oils of Nigella sativa L. Extracted by Supercritical Carbon Dioxide. Ind. Crop. Prod. 2013, 46, 317–323. [Google Scholar] [CrossRef]
- Ginther, O.J.; Utt, M.D.; Beg, M.A.; Gastal, E.L.; Gastal, M.O.; Foundation, E.; Plains, C. Negative Effect of Estradiol on Luteinizing Hormone Throughout the Ovulatory Luteinizing Hormone Surge in Mares 1. Biol. Reprod. 2007, 77, 543–550. [Google Scholar] [CrossRef] [PubMed]
- El-Fadaly, A.A.; Younis, I.Y.; Abdelhameed, M.F.; Ahmed, Y.H.; Ragab, T.I.M.; El Gendy, A.E.-N.G.; Farag, M.A.; Elshamy, A.I.; Elgamal, A.M. Protective Action Mechanisms of Launaea mucronata Extract and Its Nano-Formulation against Nephrotoxicity in Rats as Revealed via Biochemical, Histopathological, and UPLC-QTOF–MS/MS Analyses. Metabolites 2023, 13, 786. [Google Scholar] [CrossRef] [PubMed]
- Steelman, S.L.; Pohley, F.M. Assay of the Follicle Stimulating Hormone Based on the Augmentation with Human Chorionic Gonadotropin. Endocrinology 1953, 53, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Yang, D.K.; Kim, G.B.; Lee, S.J.; Kim, S.J. Nigella sativa Seed Extract Attenuates the Fatigue Induced by Exhaustive Swimming in Rats. Biomed. Rep. 2017, 6, 468–474. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Soliman, F.A. Assaying Luteinizing Hormone. Nature 1960, 185, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Salama, A.; Abdelhameed, M.F.; Mostafa, S.; Nada, S.A.; Taha, H.S.; Amer, A.A. Influence of Extract Derived Cell Cultures of Broccoli against Osteoporosis in Ovariectomized Rats. Egypt. J. Chem. 2021, 64, 3521–3539. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008; ISBN 0443102791. [Google Scholar]
- Yan, W.; Cheng, L.; Wang, W.; Wu, C.; Yang, X.; Du, X.; Ma, L.; Qi, S.; Wei, Y.; Lu, Z.; et al. Structure of the Human Gonadotropin-Releasing Hormone Receptor GnRH1R Reveals an Unusual Ligand Binding Mode. Nat. Commun. 2020, 11, 5287. [Google Scholar] [CrossRef]
- Manas, E.S.; Unwalla, R.J.; Xu, Z.B.; Malamas, M.S.; Miller, C.P.; Harris, H.A.; Hsiao, C.; Akopian, T.; Hum, W.T.; Malakian, K.; et al. Structure-Based Design of Estrogen Receptor-β Selective Ligands. J. Am. Chem. Soc. 2004, 126, 15106–15119. [Google Scholar] [CrossRef] [PubMed]
- Matias, P.M.; Donner, P.; Coelho, R.; Thomaz, M.; Peixoto, C.; Macedo, S.; Otto, N.; Joschko, S.; Scholz, P.; Wegg, A.; et al. Structural Evidence for Ligand Specificity in the Binding Domain of the Human Androgen Receptor: Implications for Pathogenic Gene Mutations. J. Biol. Chem. 2000, 275, 26164–26171. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Rt (min) | Tentatively Identified Compounds | Classes | [M-H]− m/z | MS2 Major Production | Refs. |
|---|---|---|---|---|---|---|
| 1 | 3.615 | Kaempferol | Flavanol | 285.04 | [30] | |
| 2 | 8.040 | Vanillic acid | Phenolic acid | 167.02 | [31] | |
| 3 | 8.180 | Magnoflorine | Alkaloid | 340.08 | [28] | |
| 4 | 8.423 | Thymoquinol glucoside | Monoterpene glycoside | 325.05 | 310.07, 282.09, 163.00 | [32] |
| 5 | 10.230 | Nigellidine | Alkaloid | 295.17 | 277.28, 171.17 | [28] |
| 6 | 13.592 | Hederagenin aglycone | Saponin | 471.73 | 393.02 | [32] |
| 7 | 14.090 | Hederagenin pentoside | Saponin | 603.31 | [32] | |
| 8 | 16.437 | Tauroside E | Saponin | 749.20 | 603.29, 471.36 | [32] |
| 9 | 18.390 | Hederagenin methyl ester | Saponin | 485.21 | 467.56, 439.83 | [32] |
| 10 | 27.012 | Dihydronigellidine | Alkaloid | 293.07 | 265.05, 237.05, 222.09 | [28] |
| 11 | 30.300 | Methoxycinnamic acid | Phenolic acid derivative | 177.87 | 159.90 | [31] |
| 12 | 31.689 | Thymoquinone | Monoterpene | 163.98 | 117.92 | [31,32] |
| 13 | 32.932 | Caffeic acid 3-O-glucuronide | Phenolic acid derivative | 355.3 | 179.14, 163.15 | [31] |
| 14 | 33.473 | (E)-p-Coumaric acid | Phenolic acid | 163.07 | 119.00 | [31] |
| 15 | 33.574 | Malic acid | Fatty acid | 132.84 | 115.88 | [25] |
| 16 | 33.622 | Thymoquinol glucuronic acid | Monoterpene glycoside | 339.16 | 163.12 | [32] |
| 17 | 34.931 | Octadecadienoic acid | Fatty acid | 279.24 | [26] | |
| 18 | 34.289 | Nigelanoid | Alkaloid | 312.11 | 293.86, 275.86, 183.04, 119.08 | [31] |
| 19 | 37.430 | 4-Vinylphenol | Phenol | 118.90 | 101.89 | [30] |
| 20 | 38.654 | Octadecenoic acid | Fatty acid | 281.22 | [25] | |
| 21 | 38.489 | Palmitic acid | Fatty acid | 255.29 | [26] |
| Compound | ΔG a (kcal/mol) | ||||
|---|---|---|---|---|---|
| E2 | FSH | LH | GnRH | P4 | |
| 4-Vinyl phenol | −4.89 | −3.31 | −4.37 | −4.32 | −4.66 |
| (E)-p-coumaric acid | −5.60 | −3.35 | −4.92 | −4.62 | −4.84 |
| E -Caffeic acid 3-O-glucuronide | −7.75 | −4.85 | −6.45 | −7.20 | −7.82 |
| Hederagenin methyl ester | ---- | ---- | ---- | ---- | ---- |
| Hederagenin pentoside | ---- | ---- | ---- | −7.53 | ---- |
| Hederagenin aglycone | ---- | ---- | ---- | ---- | ---- |
| Kaempferol | −7.34 | −4.15 | −6.09 | −6.13 | −6.57 |
| Magnoflorine | −8.15 | −4.45 | −6.26 | −7.00 | −8.57 |
| Malic acid | −4.93 | −3.84 | −4.58 | −4.21 | −4.44 |
| Methoxycinnamic acid | −5.71 | −3.78 | −5.08 | −4.78 | −5.12 |
| Nigelanoid | −8.80 | −4.36 | −6.04 | −6.00 | −6.92 |
| Nigellidine | −8.16 | −4.34 | −6.02 | −6.31 | −6.74 |
| Octadecadienoic acid | −9.20 | −4.74 | −6.66 | −7.49 | −7.17 |
| Octadecanoic acid | −8.56 | −5.11 | −7.31 | −7.28 | −6.57 |
| Palmitic acid | −9.00 | −4.88 | −6.75 | −6.52 | −7.63 |
| Tauroside E | ---- | ---- | ---- | ---- | ---- |
| Thymoquinol glucoside | −8.10 | −4.27 | −7.51 | −7.16 | −7.54 |
| Thymoquinol glucuronic acid | −8.23 | −4.27 | −6.95 | −6.98 | −7.53 |
| Thymoquinone | −6.05 | −3.54 | −5.40 | −4.92 | −5.27 |
| Vanillic acid | −5.72 | −3.21 | −4.89 | −4.65 | −5.06 |
| E2 ligand | −7.89 | ---- | ---- | ---- | ---- |
| FSH ligand | ---- | −6.21 | ---- | ---- | ---- |
| Org 43553 | ---- | ---- | −8.59 | ---- | ---- |
| GnRH ligand | ---- | ---- | ---- | −9.92 | ---- |
| P4 ligand | ---- | ---- | ---- | ---- | −7.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagy, A.M.; Abdelhameed, M.F.; Elkarim, A.S.A.; Sarker, T.C.; Abd-ElGawad, A.M.; Elshamy, A.I.; Hammam, A.M. Enhancement of Female Rat Fertility via Ethanolic Extract from Nigella sativa L. (Black Cumin) Seeds Assessed via HPLC-ESI-MS/MS and Molecular Docking. Molecules 2024, 29, 735. https://doi.org/10.3390/molecules29030735
Nagy AM, Abdelhameed MF, Elkarim ASA, Sarker TC, Abd-ElGawad AM, Elshamy AI, Hammam AM. Enhancement of Female Rat Fertility via Ethanolic Extract from Nigella sativa L. (Black Cumin) Seeds Assessed via HPLC-ESI-MS/MS and Molecular Docking. Molecules. 2024; 29(3):735. https://doi.org/10.3390/molecules29030735
Chicago/Turabian StyleNagy, Ahmed M., Mohamed F. Abdelhameed, Asmaa S. Abd Elkarim, Tushar C. Sarker, Ahmed M. Abd-ElGawad, Abdelsamed I. Elshamy, and Abdelmohsen M. Hammam. 2024. "Enhancement of Female Rat Fertility via Ethanolic Extract from Nigella sativa L. (Black Cumin) Seeds Assessed via HPLC-ESI-MS/MS and Molecular Docking" Molecules 29, no. 3: 735. https://doi.org/10.3390/molecules29030735
APA StyleNagy, A. M., Abdelhameed, M. F., Elkarim, A. S. A., Sarker, T. C., Abd-ElGawad, A. M., Elshamy, A. I., & Hammam, A. M. (2024). Enhancement of Female Rat Fertility via Ethanolic Extract from Nigella sativa L. (Black Cumin) Seeds Assessed via HPLC-ESI-MS/MS and Molecular Docking. Molecules, 29(3), 735. https://doi.org/10.3390/molecules29030735