
Efficient Decolorization of the Poly-Azo Dye Sirius Grey by Coriolopsis gallica Laccase-Mediator System: Process Optimization and Toxicity Assessment


 and
and
Abstract
1. Introduction
2. Results
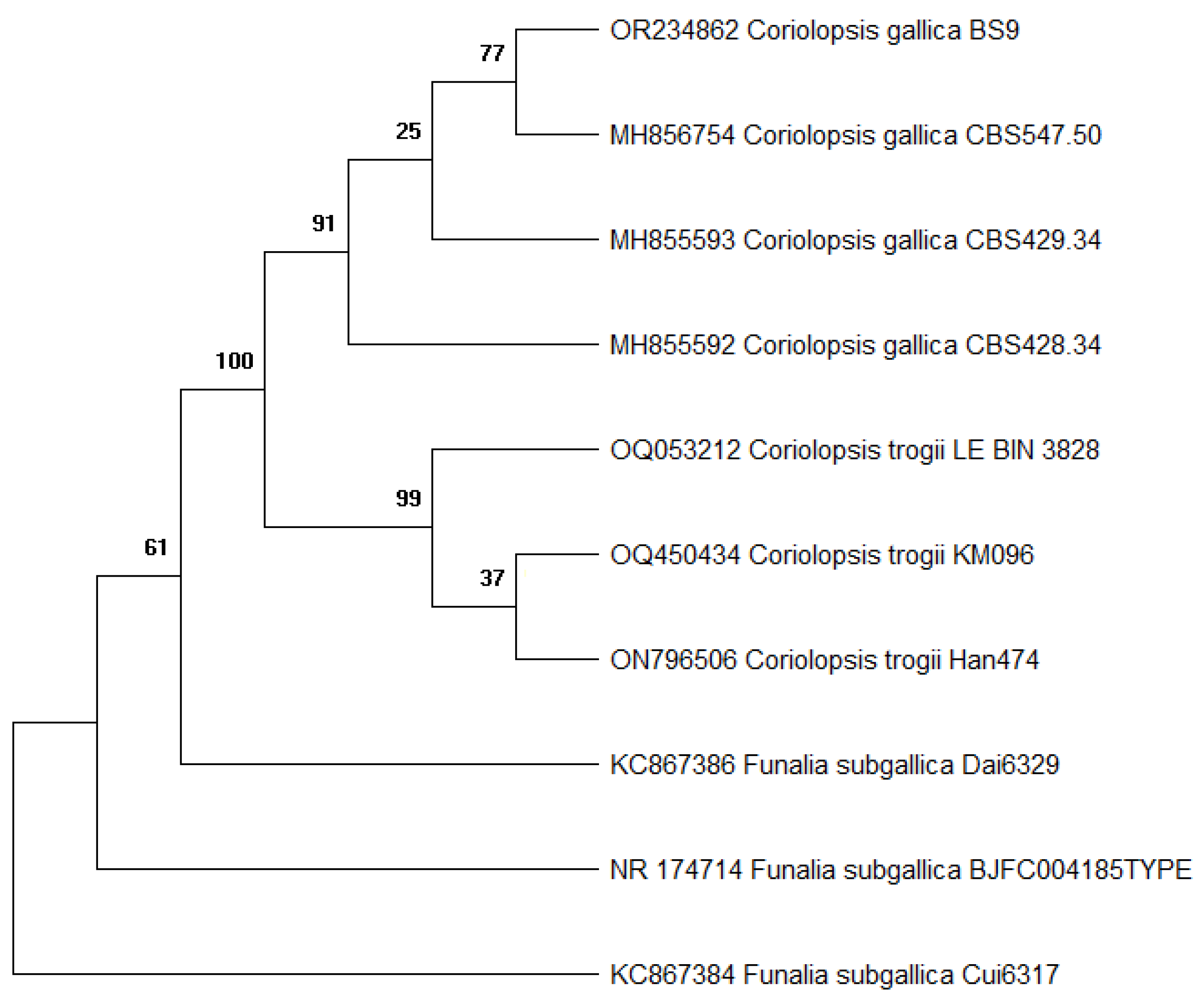
2.1. Fungi Isolation and Identification
2.2. Laccase-like Enzymes Detection and Production
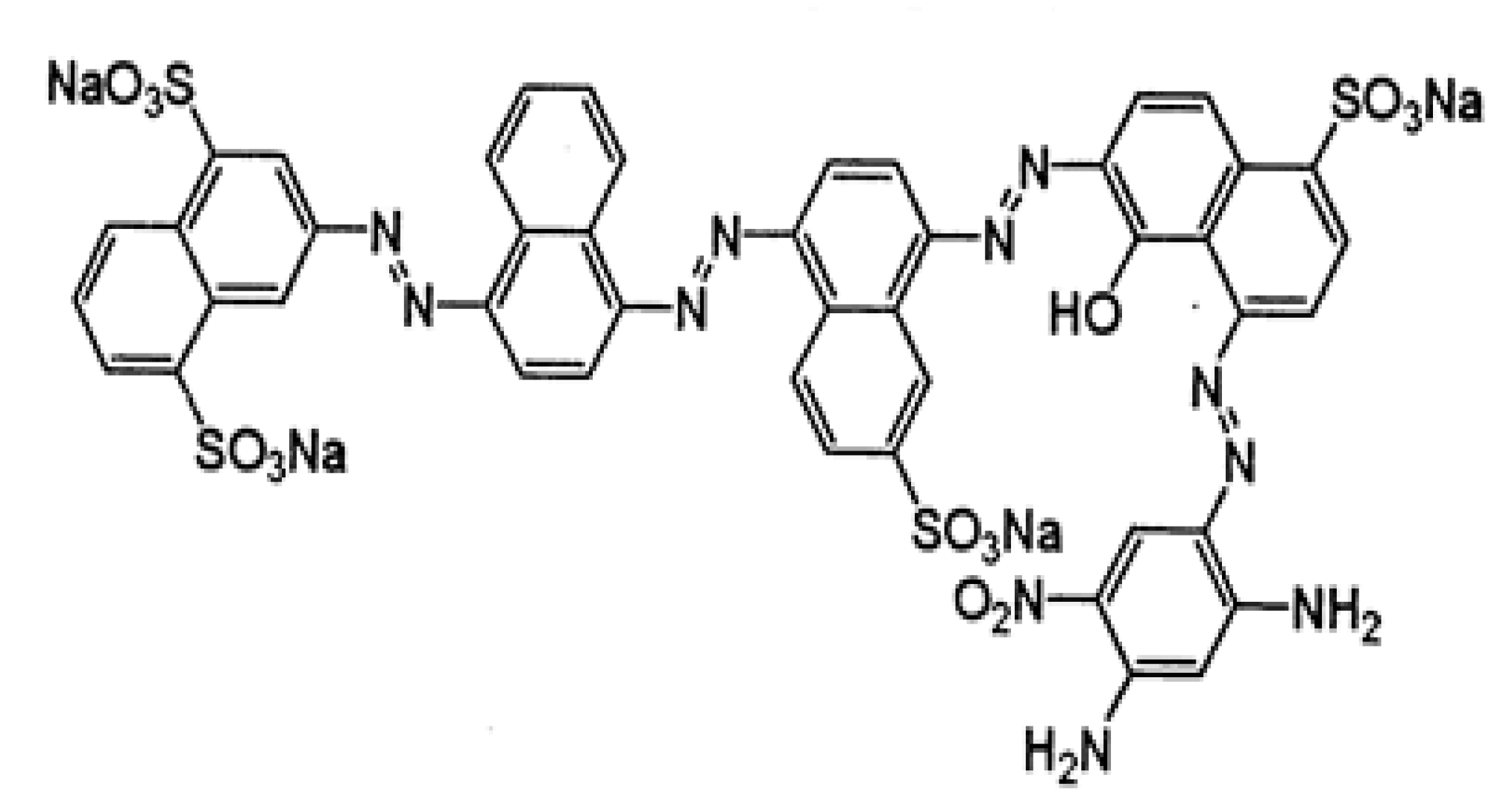
2.3. Dye Decolorization by Crude Laccase from C. gallica
2.4. Optimization of Dye Decolorization Conditions
2.5. Modeling Dye Decolorization Yield
2.6. Modeling Dye Decolorization Rate
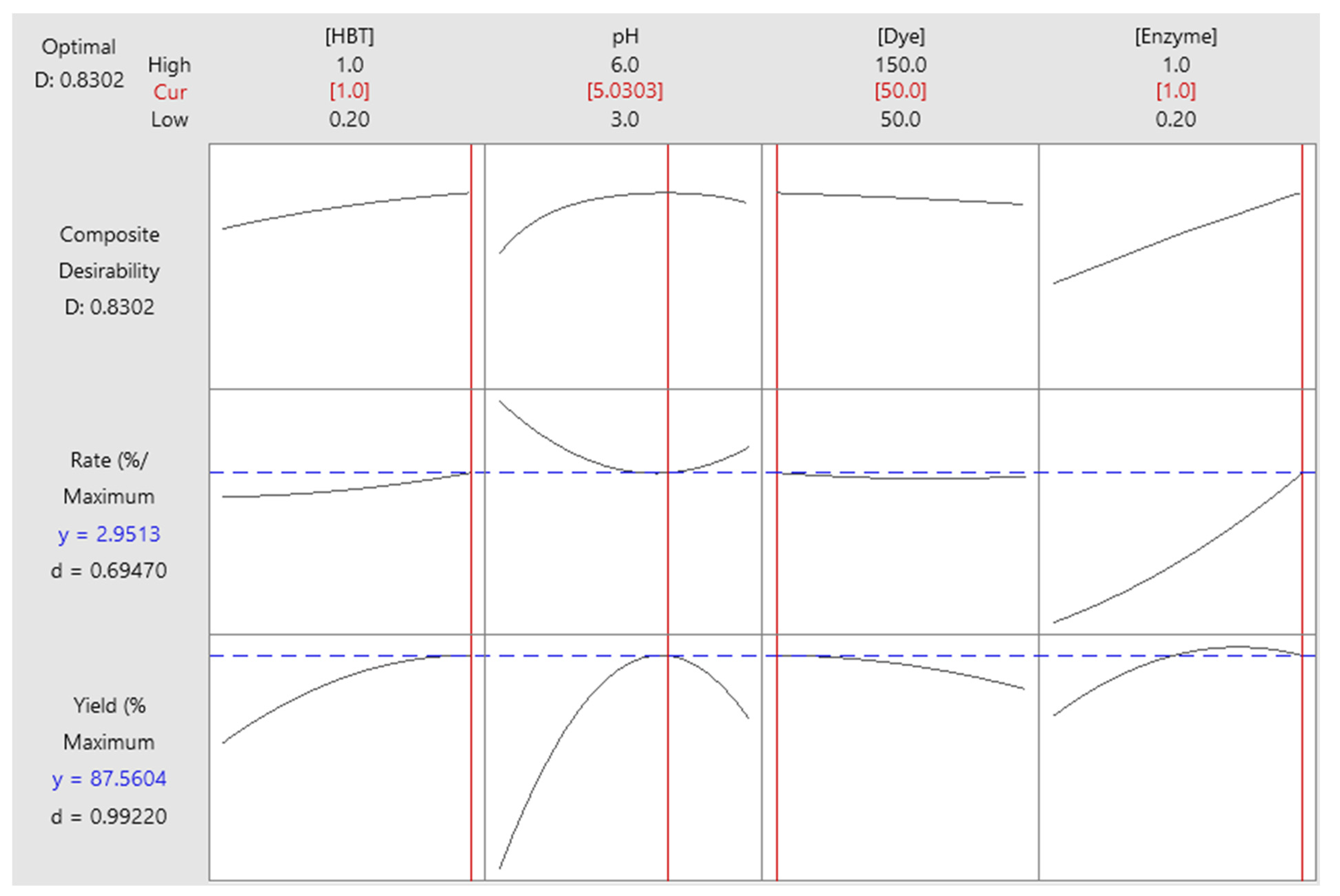
2.7. Optimization of Responses
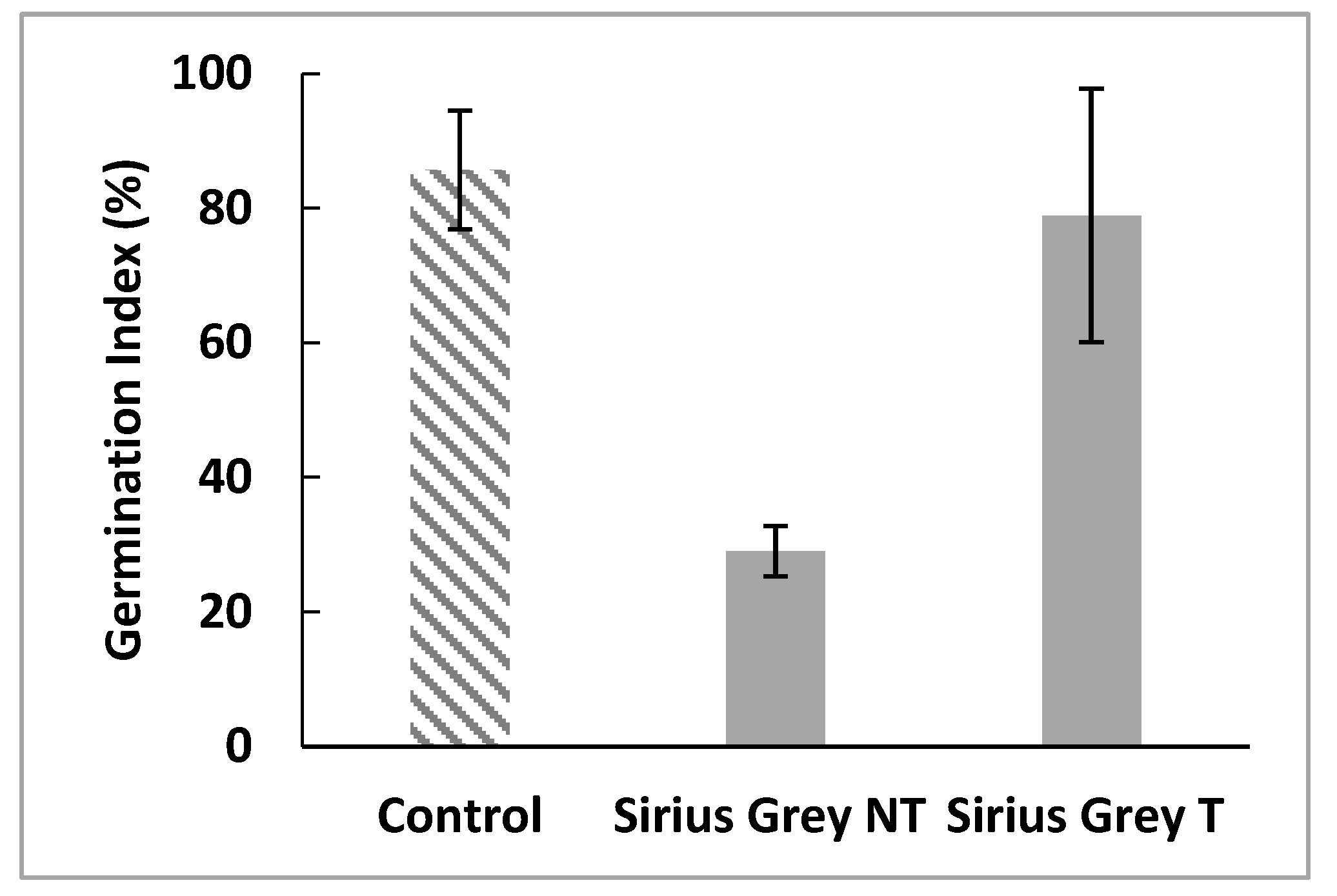
2.8. Evaluation of the Toxicity of Treated and Untreated Dye
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Media and Culture Conditions
4.3. Fungal Strain Isolation and Identification
4.3.1. Isolation
4.3.2. Fungal DNA Extraction, Amplification and Sequencing
4.3.3. Phylogenetic Analysis
4.4. Enzyme Assay
4.5. Dye Decolorization Experiments
4.6. Box-Behnken Design

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coded Levels | −1 | 0 | +1 | |
|---|---|---|---|---|
| Factor | ||||
| A: Initial HBT concentration (mM) | 0.2 | 0.6 | 1 | |
| B: pH | 3 | 4.5 | 6 | |
| C: Initial dye concentration (mg/L) | 50 | 100 | 150 | |
| D: Initial enzyme concentration (U/mL) | 0.2 | 0.6 | 1 | |
| Run | A:[HBT] * | B:pH * | C:[Dye] * | D:[Enzyme] * | y1: Yield (%) | y2: Rate (%/min) |
|---|---|---|---|---|---|---|
| 1 | −1 | −1 | 0 | 0 | 49.91 ± 2.73 | 2.93 ± 0.15 |
| 2 | 1 | −1 | 0 | 0 | 53.23 ± 1.41 | 2.87 ± 0.16 |
| 3 | −1 | 1 | 0 | 0 | 56.07 ± 0.53 | 1.81 ± 0.02 |
| 4 | 1 | 1 | 0 | 0 | 72.14 ± 0.09 | 2.09 ± 0.01 |
| 5 | 0 | 0 | −1 | −1 | 77.63 ± 0.90 | 1.81 ± 0.08 |
| 6 | 0 | 0 | 1 | −1 | 60.25 ± 0.88 | 1.29 ± 0.06 |
| 7 | 0 | 0 | −1 | 1 | 85.04 ± 1.28 | 2.90 ± 0.24 |
| 8 | 0 | 0 | 1 | 1 | 76.74 ± 1.39 | 2.67 ± 0.02 |
| 9 | −1 | 0 | 0 | −1 | 49.71 ± 1.63 | 1.43 ± 0.20 |
| 10 | 1 | 0 | 0 | −1 | 77.67 ± 0.04 | 1.68 ± 0.06 |
| 11 | −1 | 0 | 0 | 1 | 62.78 ± 0.77 | 2.63 ± 0.06 |
| 12 | 1 | 0 | 0 | 1 | 86.15 ± 0.14 | 2.96 ± 0.03 |
| 13 | 0 | −1 | −1 | 0 | 55.32 ± 0.93 | 2.68 ± 0.02 |
| 14 | 0 | +1 | −1 | 0 | 68.12 ± 2.81 | 2.13 ± 0.07 |
| 15 | 0 | −1 | +1 | 0 | 51.41 ± 1.03 | 2.82 ± 0.09 |
| 16 | 0 | +1 | +1 | 0 | 50.97 ± 0.52 | 2.03 ± 0.03 |
| 17 | −1 | 0 | −1 | 0 | 73.49 ± 1.05 | 2.38 ± 0.13 |
| 18 | 1 | 0 | −1 | 0 | 87.02 ± 1.33 | 2.51 ± 0.16 |
| 19 | −1 | 0 | +1 | 0 | 58.07 ± 0.80 | 1.83 ± 0.12 |
| 20 | 1 | 0 | +1 | 0 | 79.86 ± 1.53 | 2.21 ± 0.12 |
| 21 | 0 | −1 | 0 | −1 | 49.51 ± 1.01 | 2.82 ± 0.07 |
| 22 | 0 | +1 | 0 | −1 | 40.14 ± 2.03 | 1.37 ± 0.22 |
| 23 | 0 | −1 | 0 | +1 | 64.32 ± 1.21 | 3.63 ± 0.12 |
| 24 | 0 | +1 | 0 | +1 | 65.57 ± 1.33 | 2.74 ± 0.02 |
| 25 | 0 | 0 | 0 | 0 | 79.69 ± 1.01 | 2.17 ± 0.22 |
| 26 | 0 | 0 | 0 | 0 | 79.01 ± 0.51 | 1.98 ± 0.08 |
| 27 | 0 | 0 | 0 | 0 | 79.19 ± 0.83 | 2.04 ± 0.11 |
4.7. Design of Experiments and Statistical Analysis
4.8. Phytotoxicity Assay
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, M. Handbook of Textile and Industrial Dyeing: Principles, Processes and Types of Dyes; Elsevier: Amsterdam, The Netherlands, 2011; ISBN 0-85709-397-5. [Google Scholar]
- Abel, A. The History of Dyes and Pigments: From Natural Dyes to High Performance Pigments. In Colour Design; Elsevier: Amsterdam, The Netherlands, 2012; pp. 433–470. ISBN 978-1-84569-972-7. [Google Scholar]
- Benkhaya, S.; M’rabet, S.; Lgaz, H.; El Bachiri, A.; El Harfi, A. Dyes: Classification, Pollution, and Environmental Effects. In Dye Biodegradation, Mechanisms and Techniques; Muthu, S.S., Khadir, A., Eds.; Sustainable Textiles: Production, Processing, Manufacturing & Chemistry; Springer: Singapore, 2022; pp. 1–50. ISBN 9789811659317. [Google Scholar]
- Benkhaya, B.; El Harfi, S.; El Harfi, A. Classifications, Properties and Applications of Textile Dyes: A Review. Appl. J. Environ. Eng. Sci. 2017, 3, 311–320. [Google Scholar] [CrossRef]
- Benkhaya, S.; M’Rabet, S.; El Harfi, A. A Review on Classifications, Recent Synthesis and Applications of Textile Dyes. Inorg. Chem. Commun. 2020, 115, 107891. [Google Scholar] [CrossRef]
- Mokhtar, A.; Abdelkrim, S.; Boukoussa, B.; Hachemaoui, M.; Djelad, A.; Sassi, M.; Abboud, M. Elimination of Toxic Azo Dye Using a Calcium Alginate Beads Impregnated with NiO/Activated Carbon: Preparation, Characterization and RSM Optimization. Int. J. Biol. Macromol. 2023, 233, 123582. [Google Scholar] [CrossRef]
- Ben Ayed, A.; Hadrich, B.; Sciara, G.; Lomascolo, A.; Bertrand, E.; Faulds, C.B.; Zouari-Mechichi, H.; Record, E.; Mechichi, T. Optimization of the Decolorization of the Reactive Black 5 by a Laccase-like Active Cell-Free Supernatant from Coriolopsis gallica. Microorganisms 2022, 10, 1137. [Google Scholar] [CrossRef] [PubMed]
- Brüschweiler, B.J.; Merlot, C. Azo Dyes in Clothing Textiles Can Be Cleaved into a Series of Mutagenic Aromatic Amines Which Are Not Regulated Yet. Regul. Toxicol. Pharmacol. 2017, 88, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, U.; Shakoori, F.R.; Zahid, M.T.; Majeed, W.; Zahra, I.; Abbas, S.Z.; Hedfi, A.; Hassan, S.; Shakoori, A.R.; Mutery, A.A. Biodegradation and Decolorization of Textile Azo Dyes by Paramecium caudatum Isolated from Industrial Wastewater. Water 2022, 14, 3553. [Google Scholar] [CrossRef]
- Chung, K.-T. Azo Dyes and Human Health: A Review. J. Environ. Sci. Health Part C 2016, 34, 233–261. [Google Scholar] [CrossRef]
- Sarkar, S.; Banerjee, A.; Halder, U.; Biswas, R.; Bandopadhyay, R. Degradation of Synthetic Azo Dyes of Textile Industry: A Sustainable Approach Using Microbial Enzymes. Water Conserv Sci Eng 2017, 2, 121–131. [Google Scholar] [CrossRef]
- Souza, M.C.O.; González, N.; Herrero, M.; Marquès, M.; Rovira, J.; Nadal, M.; Barbosa, F.; Domingo, J.L. Screening of Regulated Aromatic Amines in Clothing Marketed in Brazil and Spain: Assessment of Human Health Risks. Environ. Res. 2023, 221, 115264. [Google Scholar] [CrossRef] [PubMed]
- Lellis, B.; Fávaro-Polonio, C.Z.; Pamphile, J.A.; Polonio, J.C. Effects of Textile Dyes on Health and the Environment and Bioremediation Potential of Living Organisms. Biotechnol. Res. Innov. 2019, 3, 275–290. [Google Scholar] [CrossRef]
- Ngo, A.C.R.; Devanadera, M.K.P.; Dedeles, G.R. Decolorization of Selected Synthetic Textile Dyes by Yeasts from Leaves and Fruit Peels. J. Health Pollut. 2016, 6, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Moyo, S.; Makhanya, B.P.; Zwane, P.E. Use of Bacterial Isolates in the Treatment of Textile Dye Wastewater: A Review. Heliyon 2022, 8, e09632. [Google Scholar] [CrossRef]
- Ngo, A.C.R.; Tischler, D. Microbial Degradation of Azo Dyes: Approaches and Prospects for a Hazard-Free Conversion by Microorganisms. Int. J. Environ. Res. Public Health 2022, 19, 4740. [Google Scholar] [CrossRef] [PubMed]
- Ghazal, F.; Mahdy, E.-S.; Abdelfattah, M.; EL-Sadany, A.; Doha, N. The Use of Microalgae in Bioremediation of the Textile Wastewater Effluent. Nat. Sci. 2018, 16, 98–104. [Google Scholar]
- Rajhans, G.; Barik, A.; Sen, S.; Raut, S. Degradation of Dyes by Fungi: An Insight into Mycoremediation. BioTechnologia 2021, 102, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Sun, K.; Wang, F.; Zhao, L.; Hu, J.; Ma, H.; Ding, Z. Laccase Production by Trametes versicolor in Solid-State Fermentation Using Tea Residues as Substrate and Its Application in Dye Decolorization. J. Environ. Manag. 2020, 270, 110904. [Google Scholar] [CrossRef]
- Levin, L.; Grassi, E.; Carballo, R. Efficient Azoic Dye Degradation by Trametes trogii and a Novel Strategy to Evaluate Products Released. Int. Biodeterior. Biodegrad. 2012, 75, 214–222. [Google Scholar] [CrossRef]
- Daâssi, D.; Rodríguez-Couto, S.; Nasri, M.; Mechichi, T. Biodegradation of Textile Dyes by Immobilized Laccase from Coriolopsis gallica into Ca-Alginate Beads. Int. Biodeterior. Biodegrad. 2014, 90, 71–78. [Google Scholar] [CrossRef]
- Alihosseini, F.; Azimi, S.H.; Mostajeran, M.; Feiz, M. Removal of Different Vat Dyes by Aspergillus niger: A Comparative Study Focusing on the Molecular Structure. J. Microbiol. Methods 2023, 208, 106720. [Google Scholar] [CrossRef]
- Grace Barrios-Gutiérrez, S.; Inés Vélez-Mercado, M.; Rodrigues Ortega, J.; Da Silva Lima, A.; Luiza Da Rocha Fortes Saraiva, A.; Leila Berto, G.; Segato, F. Oxidative Machinery of Basidiomycetes as Potential Enhancers in Lignocellulosic Biorefineries: A Lytic Polysaccharide Monooxygenases Approach. Bioresour. Technol. 2023, 386, 129481. [Google Scholar] [CrossRef]
- Singh, A.K.; Iqbal, H.M.N.; Cardullo, N.; Muccilli, V.; Fernández-Lucas, J.; Schmidt, J.E.; Jesionowski, T.; Bilal, M. Structural Insights, Biocatalytic Characteristics, and Application Prospects of Lignin-Modifying Enzymes for Sustainable Biotechnology. Int. J. Biol. Macromol. 2023, 242, 124968. [Google Scholar] [CrossRef] [PubMed]
- Mate, D.M.; Alcalde, M. Laccase Engineering: From Rational Design to Directed Evolution. Biotechnol. Adv. 2015, 33, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kaur, K.; Puri, S.; Sharma, P. Critical Factors Affecting Laccase-Mediated Biobleaching of Pulp in Paper Industry. Appl. Microbiol. Biotechnol. 2015, 99, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Yuan, L.; Jia, L.; Xue, P.; Yao, H. Recent Developments of a Co-Immobilized Laccase–Mediator System: A Review. RSC Adv. 2021, 11, 29498–29506. [Google Scholar] [CrossRef]
- Daâssi, D.; Zouari-Mechichi, H.; Belbahri, L.; Barriuso, J.; Martínez, M.J.; Nasri, M.; Mechichi, T. Phylogenetic and Metabolic Diversity of Tunisian Forest Wood-Degrading Fungi: A Wealth of Novelties and Opportunities for Biotechnology. 3 Biotech 2016, 6, 46. [Google Scholar] [CrossRef]
- Agrawal, N.; Shahi, S.K. Degradation of Polycyclic Aromatic Hydrocarbon (Pyrene) Using Novel Fungal Strain Coriolopsis byrsina Strain APC5. Int. Biodeterior. Biodegrad. 2017, 122, 69–81. [Google Scholar] [CrossRef]
- Cen, Q.; Wu, X.; Cao, L.; Lu, Y.; Lu, X.; Chen, J.; Fu, G.; Liu, Y.; Ruan, R. Green Production of a Yellow Laccase by Coriolopsis gallica for Phenolic Pollutants Removal. AMB Express 2022, 12, 96. [Google Scholar] [CrossRef]
- Ben Ayed, A.; Akrout, I.; Albert, Q.; Greff, S.; Simmler, C.; Armengaud, J.; Kielbasa, M.; Turbé-Doan, A.; Chaduli, D.; Navarro, D.; et al. Biotransformation of the Fluoroquinolone, Levofloxacin, by the White-Rot Fungus Coriolopsis Gall. J. Fungi 2022, 8, 965. [Google Scholar] [CrossRef]
- Misal, S.A.; Gawai, K.R. Azoreductase: A Key Player of Xenobiotic Metabolism. Bioresour. Bioprocess. 2018, 5, 17. [Google Scholar] [CrossRef]
- Forootanfar, H.; Rezaei, S.; Zeinvand-Lorestani, H.; Tahmasbi, H.; Mogharabi, M.; Ameri, A.; Faramarzi, M.A. Studies on the Laccase-Mediated Decolorization, Kinetic, and Microtoxicity of Some Synthetic Azo Dyes. J. Environ. Health Sci. Eng. 2016, 14, 7. [Google Scholar] [CrossRef]
- Aksu, Z.; Tezer, S. Biosorption of Reactive Dyes on the Green Alga Chlorella vulgaris. Process Biochem. 2005, 40, 1347–1361. [Google Scholar] [CrossRef]
- Kumar, R.; Kaur, J.; Jain, S.; Kumar, A. Optimization of Laccase Production from Aspergillus flavus by Design of Experiment Technique: Partial Purification and Characterization. J. Genet. Eng. Biotechnol. 2016, 14, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Zhou, G.; Peng, C.; Zhang, Y.; Kües, U.; Liu, J.; Xiao, Y.; Fang, Z. The First Fungal Laccase with an Alkaline pH Optimum Obtained by Directed Evolution and Its Application in Indigo Dye Decolorization. AMB Express 2019, 9, 151. [Google Scholar] [CrossRef]
- Çifçi, D.İ.; Atav, R.; Güneş, Y.; Güneş, E. Determination of the Color Removal Efficiency of Laccase Enzyme Depending on Dye Class and Chromophore. Water Sci. Technol. 2019, 80, 134–143. [Google Scholar] [CrossRef]
- Munk, L.; Andersen, M.L.; Meyer, A.S. Influence of Mediators on Laccase Catalyzed Radical Formation in Lignin. Enzym. Microb. Technol. 2018, 116, 48–56. [Google Scholar] [CrossRef]
- Robinson, P.K. Enzymes: Principles and Biotechnological Applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef]
- Hassan, J.; Rajib, M.M.R.; Sarker, U.; Akter, M.; Khan, M.N.-E.-A.; Khandaker, S.; Khalid, F.; Rahman, G.K.M.M.; Ercisli, S.; Muresan, C.C.; et al. Optimizing Textile Dyeing Wastewater for Tomato Irrigation through Physiochemical, Plant Nutrient Uses and Pollution Load Index of Irrigated Soil. Sci. Rep. 2022, 12, 10088. [Google Scholar] [CrossRef]
- Singh, R.; Rathore, D. Effects of Fertilization with Textile Effluent on Germination, Growth and Metabolites of Chilli (Capsicum annum L) Cultivars. Environ. Process. 2021, 8, 1249–1266. [Google Scholar] [CrossRef]
- Ostrowski, G.; Jaworska, D.; Płecha, M.; Przybylski, W.; Sałek, P.; Sawicki, K.; Pawłowska, J. Cold Adapted and Closely Related Mucoraceae Species Colonise Dry-Aged Beef (DAB). Fungal Biol. 2023, 127, 1397–1404. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; The UGENE team Unipro. UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.; Guillén, F.; Martínez, A.T.; Martínez, M.J. Induction and Characterization of Laccase in the Ligninolytic Fungus Pleurotus eryngii. Curr. Microbiol. 1997, 34, 1–5. [Google Scholar] [CrossRef]
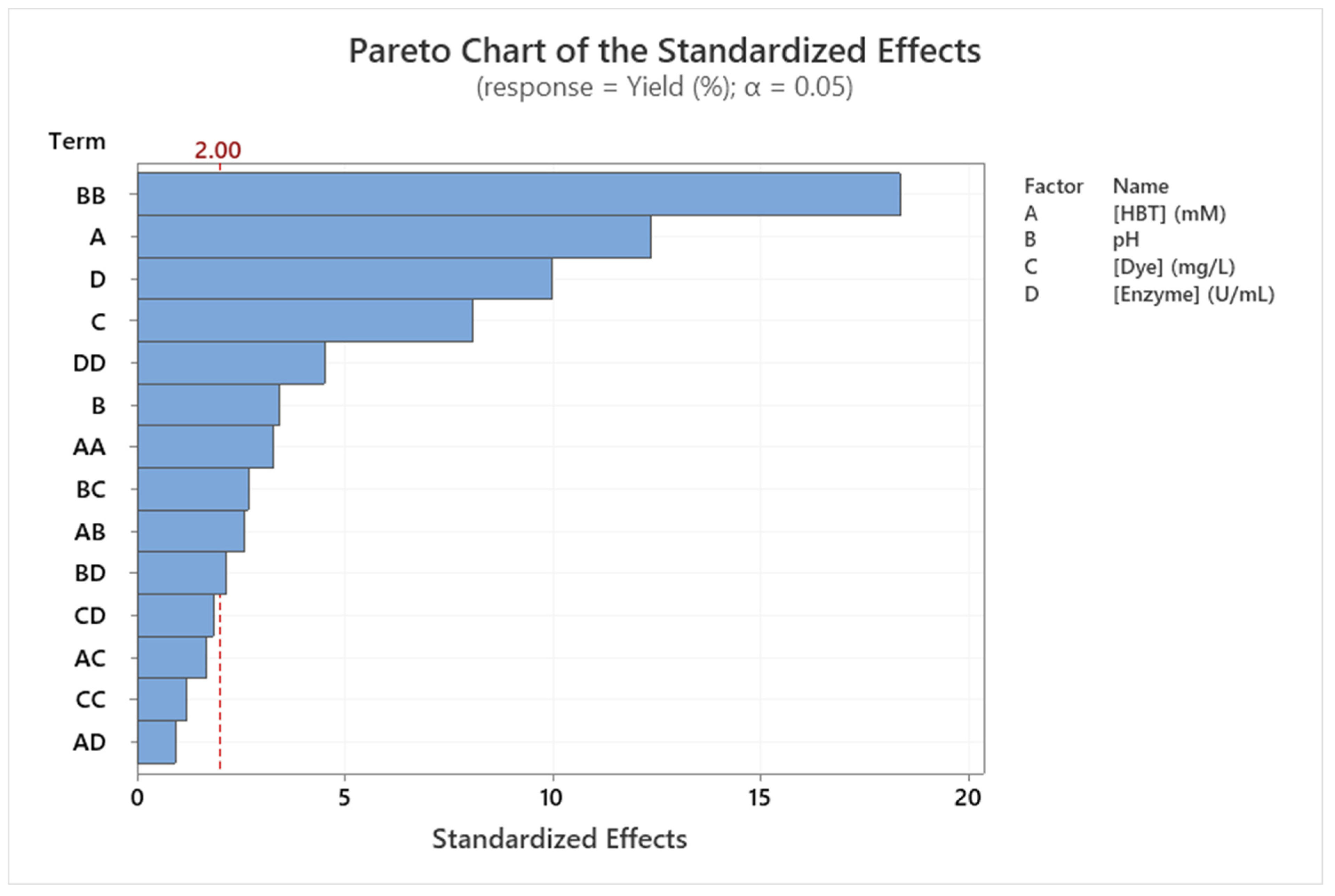
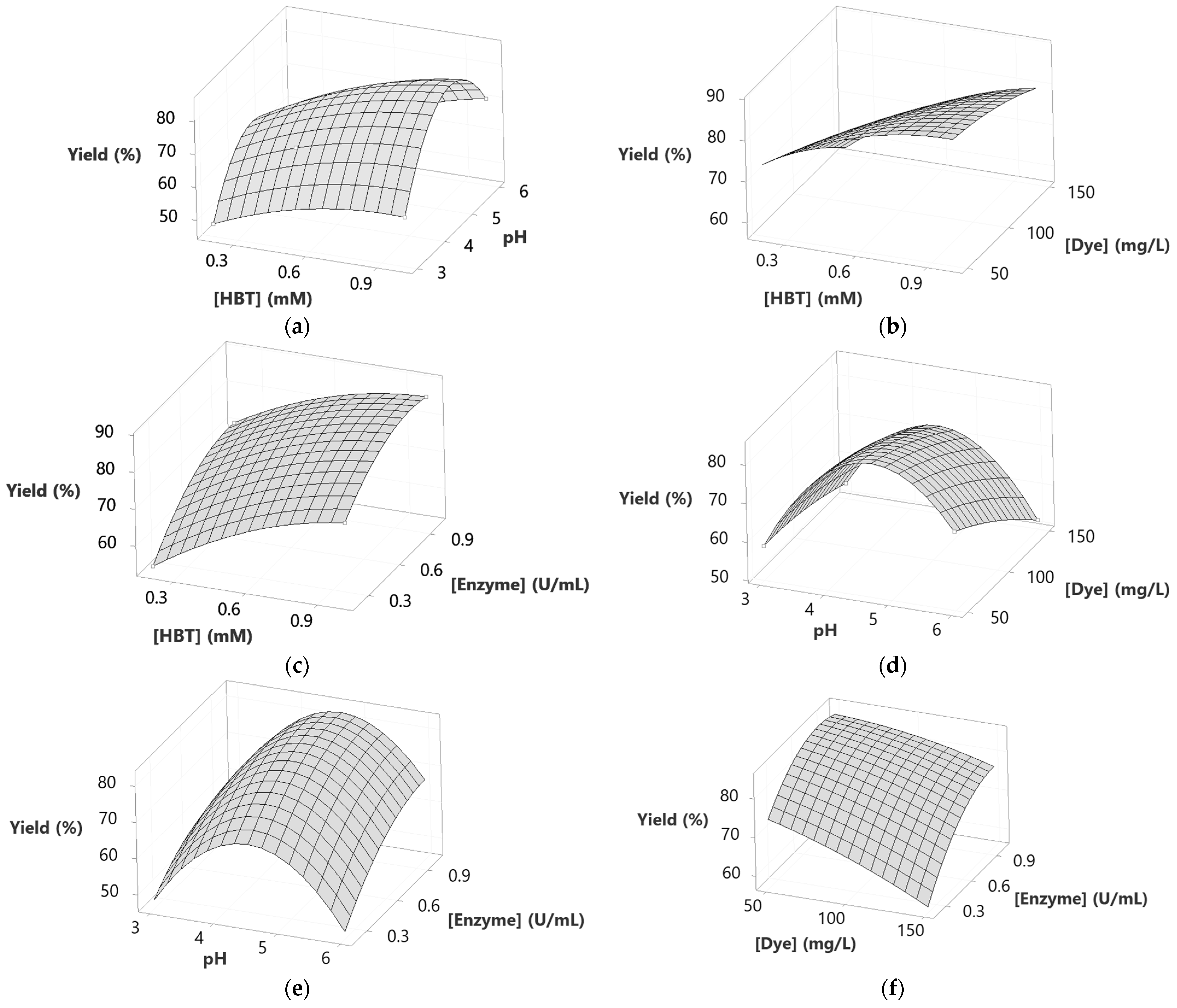

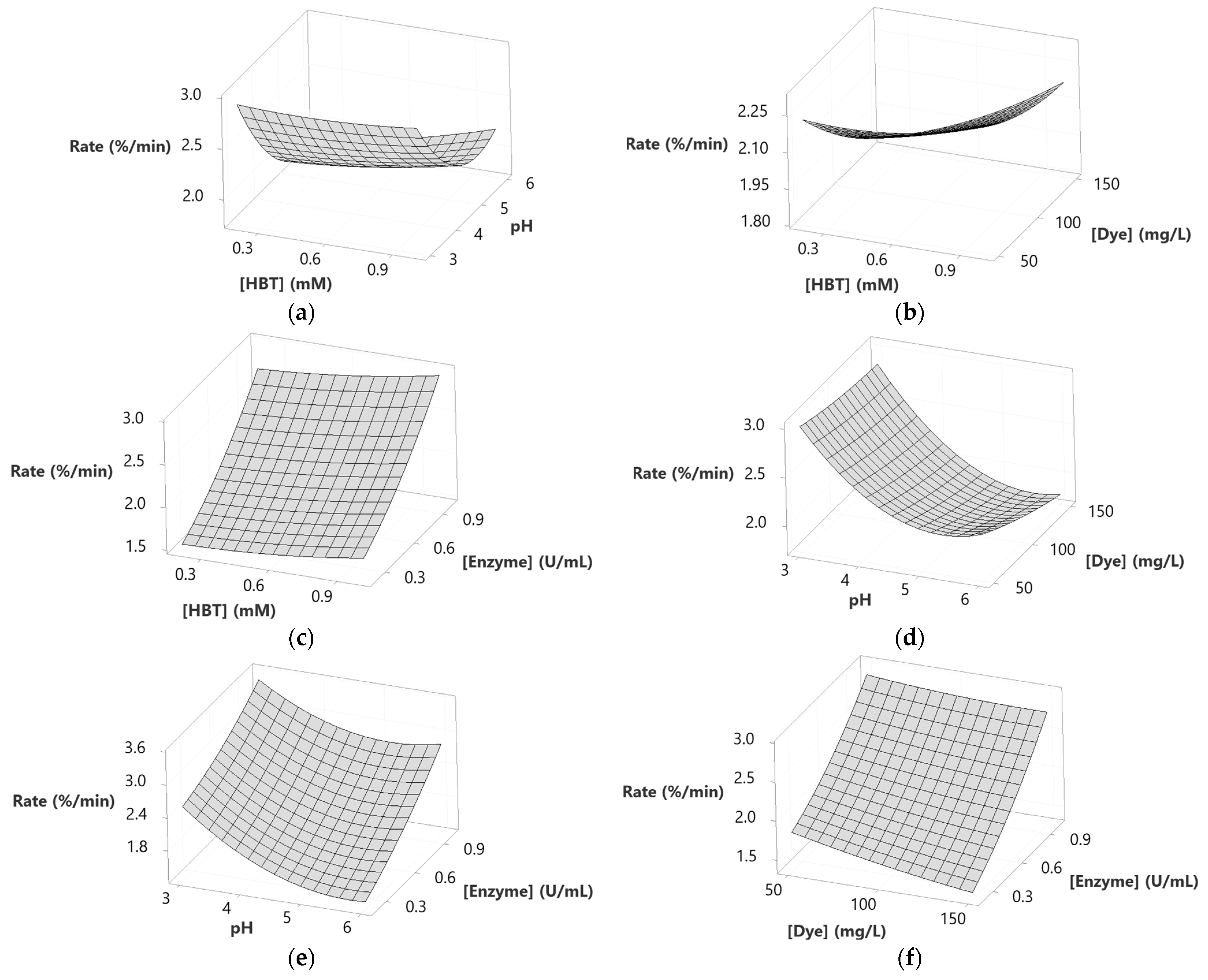
| Source | DF | SS | MS | F | p |
|---|---|---|---|---|---|
| Regression | 14 | 13290.4 | 949.3 | 51.63 | <0.001 *** |
| Linear | 4 | 6062.2 | 1515.5 | 82.43 | <0.001 *** |
| A: Initial [HBT] | 1 | 2810.8 | 2810.8 | 152.87 | <0.001 *** |
| B: pH | 1 | 214.7 | 214.7 | 11.68 | 0.001 *** |
| C: Initial [Dye] | 1 | 1200.9 | 1200.9 | 65.31 | <0.001 *** |
| D: Initial [Enzyme] | 1 | 1835.8 | 1835.8 | 99.85 | <0.001 *** |
| Square | 4 | 6761.1 | 1690.3 | 91.93 | <0.001 *** |
| A × A | 1 | 197.8 | 197.8 | 10.76 | 0.002 ** |
| B × B | 1 | 6202.5 | 6202.5 | 337.34 | <0.001 *** |
| C × C | 1 | 26.5 | 26.5 | 1.44 | 0.234 |
| D × D | 1 | 374.7 | 374.66 | 20.38 | <0.001 *** |
| Interaction | 6 | 467.1 | 77.86 | 4.23 | 0.001 *** |
| A × B | 1 | 121.9 | 121.92 | 6.63 | 0.012 * |
| A × C | 1 | 51.3 | 51.25 | 2.79 | 0.100 |
| A × D | 1 | 15.8 | 15.80 | 0.86 | 0.357 |
| B × C | 1 | 131.6 | 131.61 | 7.16 | 0.009 ** |
| B × D | 1 | 84.6 | 84.64 | 4.60 | 0.036 * |
| C × D | 1 | 61.9 | 61.93 | 3.37 | 0.071 |
| Error | 66 | 1213.5 | 18.39 | ||
| Total | 80 | 14503.9 |
| Source | DF | SS | MS | F | p |
|---|---|---|---|---|---|
| Regression | 14 | 24.348 | 1.739 | 57.96 | <0.001 *** |
| Linear | 4 | 21.498 | 5.375 | 179.11 | <0.001 *** |
| A: Initial [HBT] | 1 | 0.420 | 0.420 | 13.99 | <0.001 *** |
| B: pH | 1 | 7.767 | 7.767 | 258.85 | <0.001 *** |
| C: Initial [Dye] | 1 | 0.612 | 0.612 | 20.41 | <0.001 *** |
| D: Initial [Enzyme] | 1 | 12.699 | 12.699 | 423.19 | <0.001 *** |
| Square | 4 | 2.369 | 0.592 | 19.73 | <0.001 *** |
| A × A | 1 | 0.026 | 0.026 | 0.86 | 0.357 |
| B × B | 1 | 2.137 | 2.137 | 71.22 | <0.001 *** |
| C × C | 1 | 0.015 | 0.015 | 0.51 | 0.479 |
| D × D | 1 | 0.214 | 0.214 | 7.14 | 0.009 ** |
| Interaction | 6 | 0.481 | 0.080 | 2.67 | 0.022 * |
| A × B | 1 | 0.086 | 0.086 | 2.85 | 0.096 |
| A × C | 1 | 0.048 | 0.048 | 1.61 | 0.209 |
| A × D | 1 | 0.005 | 0.005 | 0.18 | 0.672 |
| B × C | 1 | 0.041 | 0.041 | 1.36 | 0.248 |
| B × D | 1 | 0.236 | 0.236 | 7.88 | 0.007 ** |
| C × D | 1 | 0.065 | 0.065 | 2.15 | 0.147 |
| Error | 66 | 1.981 | 0.030 | ||
| Total | 80 | 26.329 |
| Properties | Sirius Grey GB | DMP | HBT |
|---|---|---|---|
| CAS number | 6409-87-6 | 91-10-1 | 12333-53-9 |
| Molecular weight (g mol−1) | 1193.99 | 154.16 | 135.12 |
| EC Number | 241-164-5 | 202-041-1 | 219-989-7 |
| CI | 35,865 | - | - |
| Empirical formula | C46H27N11O15S4.4Na | (CH3O)2C6H3OH | C6H5N3O · xH2O |
| Other names | Direct black 76 | Pyrogallol 1,3-dimethyl ether | HOBt Hydrate |
| λmax (nm) | 610 | - | - |
| Number azo bonds | 4 | - | - |
| Purity (%) | 99 | 97 |
| Species | Culture Collection Designation | ITS Accession Number |
|---|---|---|
| C. gallica | BS9 | OR234862 |
| C. gallica | CBS547.50 | MH856754 |
| C. gallica | CBS429.34 | MH855593 |
| C. gallica | CBS428.34 | MH855592 |
| C. trogii | LE-BIN_3828 | OQ053212 |
| C. trogii | KM096 | OQ450434 |
| C. trogii | Han474 | ON796506 |
| Funalia subgallica | Dai6329 | KC867386 |
| F. subgallica | Cui6317 | KC867384 |
| F. subgallica | BJFC004185 | NR_174714 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zouari-Mechichi, H.; Benali, J.; Alessa, A.H.; Hadrich, B.; Mechichi, T. Efficient Decolorization of the Poly-Azo Dye Sirius Grey by Coriolopsis gallica Laccase-Mediator System: Process Optimization and Toxicity Assessment. Molecules 2024, 29, 477. https://doi.org/10.3390/molecules29020477
Zouari-Mechichi H, Benali J, Alessa AH, Hadrich B, Mechichi T. Efficient Decolorization of the Poly-Azo Dye Sirius Grey by Coriolopsis gallica Laccase-Mediator System: Process Optimization and Toxicity Assessment. Molecules. 2024; 29(2):477. https://doi.org/10.3390/molecules29020477
Chicago/Turabian StyleZouari-Mechichi, Héla, Jihen Benali, Abdulrahman H. Alessa, Bilel Hadrich, and Tahar Mechichi. 2024. "Efficient Decolorization of the Poly-Azo Dye Sirius Grey by Coriolopsis gallica Laccase-Mediator System: Process Optimization and Toxicity Assessment" Molecules 29, no. 2: 477. https://doi.org/10.3390/molecules29020477
APA StyleZouari-Mechichi, H., Benali, J., Alessa, A. H., Hadrich, B., & Mechichi, T. (2024). Efficient Decolorization of the Poly-Azo Dye Sirius Grey by Coriolopsis gallica Laccase-Mediator System: Process Optimization and Toxicity Assessment. Molecules, 29(2), 477. https://doi.org/10.3390/molecules29020477