Characterization of Neutral Lipids of the Oleaginous Alga Micractinum inermum



Abstract
1. Introduction
2. Results
2.1. M. inermum and Biomass Collection
2.2. Moisture and Total Lipid Content
2.3. Separation of Lipid Classes and 1H NMR Analysis
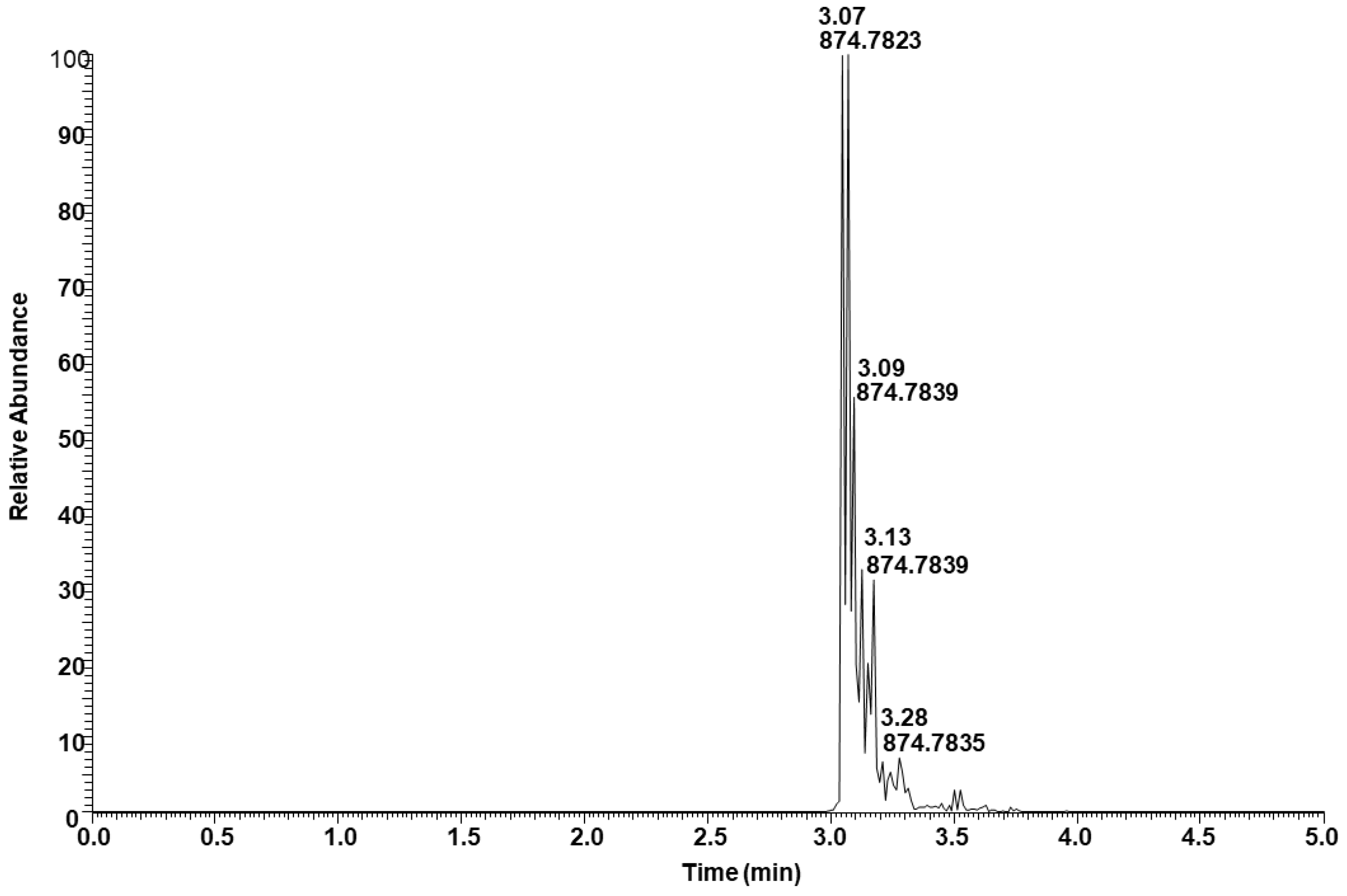
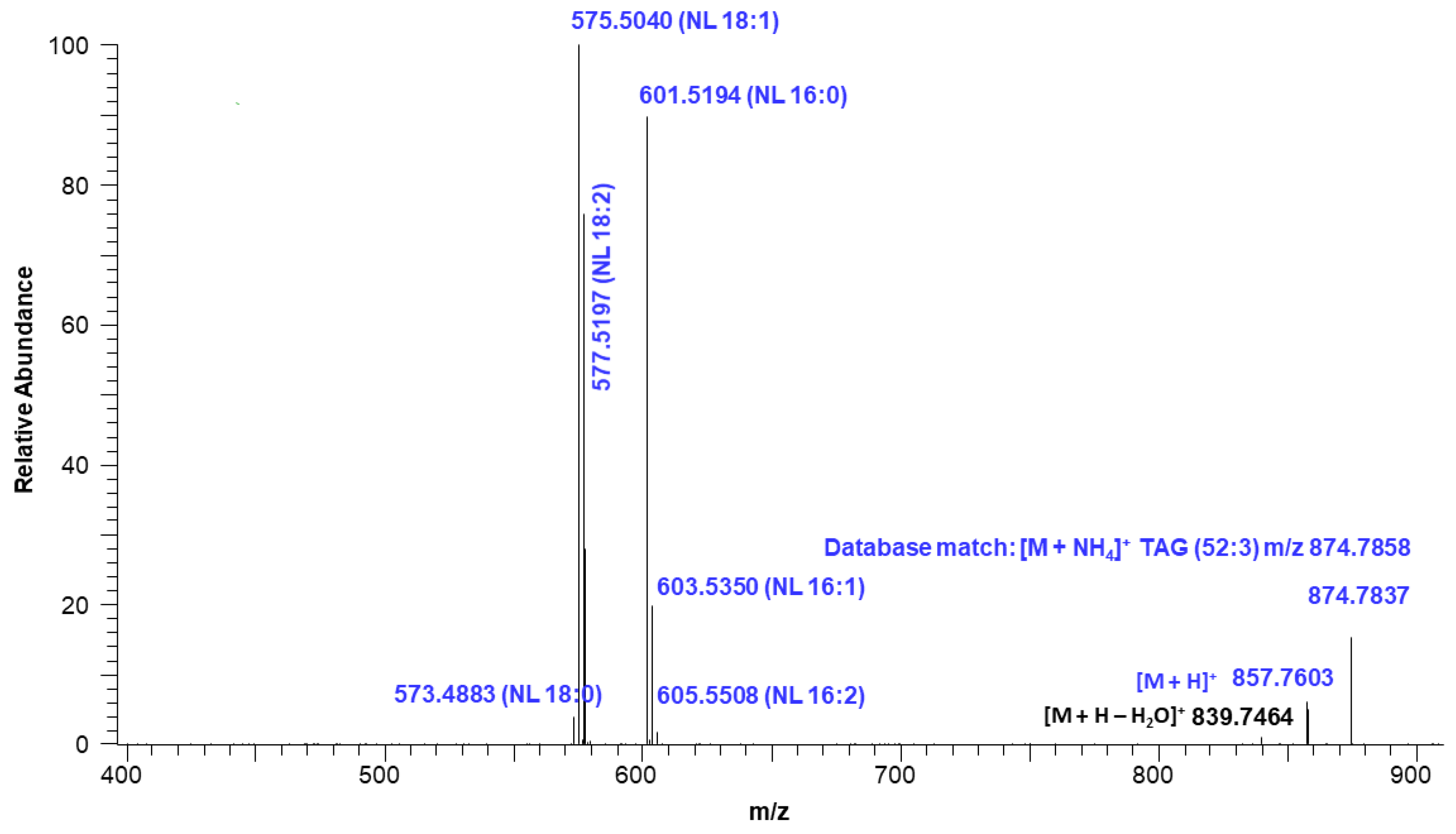
2.4. Triacylglycerols (TAGs) Analysis of Neutral Lipid Fraction
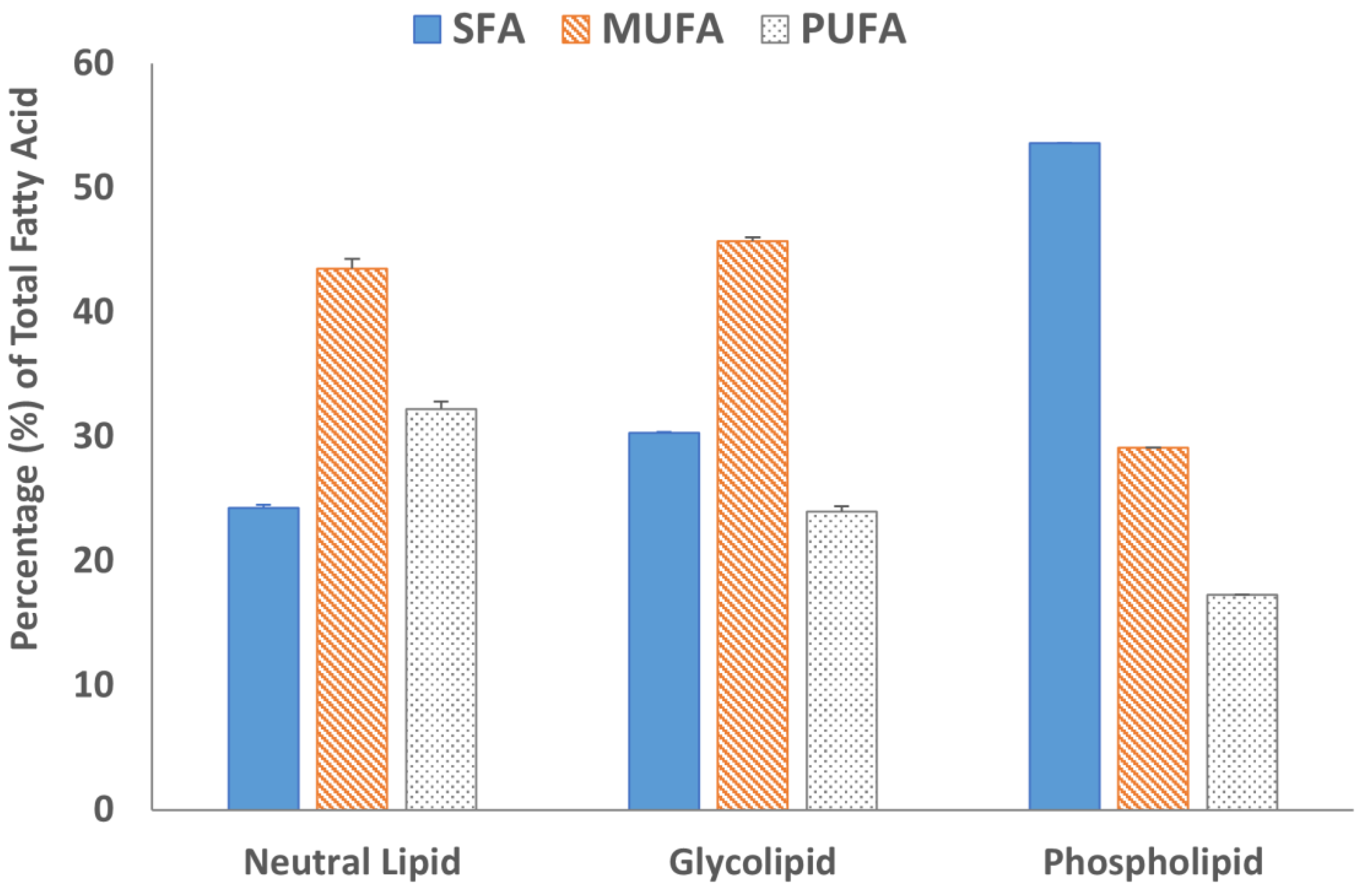
2.5. Fatty Acid Analysis of Lipid Fractions
3. Discussion
4. Materials and Methods
4.1. General
4.2. M. inermum Culture
4.3. Total Lipid Content
4.4. Lipid Class Separation by Solid Phase Extraction (SPE)
4.5. Triacylglycerols (TAGs) Analysis
4.6. Fatty Acid Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Skulberg, O.M. Microalgae as a source of bioactive molecules—Experience from cyanophyte research. J. Appl. Phycol. 2000, 12, 341–348. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococccus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef]
- Borowitzka, L.J.; Borowitzka, M.A. Commercial production of b-carotene by Dunaliella salina in open ponds. Bull. Mar. Sci. 1990, 47, 244–252. [Google Scholar]
- Eriksen, N.T. Research trends in the dominating microalgal pigments, b-carotene, astaxanthin, and phycocyanin used in feed, in foods, and in health applications. J. Nutr. Food Sci. 2016, 6, 507. [Google Scholar]
- Thanigaivel, S.; Priya, A.K.; Dutta, K.; Rajendran, S.; Vasseghian, Y. Engineering strategies and opportunities of next generation biofuel from microalgae: A perspective review on the potential bioenergy feedstock. Fuel 2022, 312, 122827. [Google Scholar] [CrossRef]
- Shi, M.; Wei, H.; Chen, Q.; Wang, X.; Zhou, W.; Liu, W. Exploring an isolate of the oleaginous alga Micractinium inermum for lipid production: Molecular characterization and physiochemical analysis under multiple growth conditions. J Appl. Phycol. 2019, 31, 1035–1046. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.; Park, Y.; Son, S.; Cho, S.; Kim, C.; Lee, T. Comparison of batch cultivation strategies for cost-effective biomass production of Micractinium inermum NLP-F014 using a blended wastewater medium. Bioresour. Technol. 2017, 234, 432–438. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.; Yoon, Y.; Park, Y.; Lee, T. Blending water- and nutrient-source wastewaters for cost-effective cultivation of high lipid content microalgal species Micractinium inermum NLP-F014. Bioresour. Technol. 2015, 198, 388–394. [Google Scholar] [CrossRef]
- Gibbons, G.F.; Islam, K.; Pease, R.J. Metabolism of triacylglycerol stores. Biochim. Biophys. Acta 2000, 1483, 37–57. [Google Scholar] [CrossRef]
- Prades, J.; Funari, S.S.; Escribá, P.V.; Barceló, F. Effects of unsaturated fatty acids and triacylglycerols on phosphatidylethanolamine membrane structure. J. Lipid Res. 2003, 44, 1720–1727. [Google Scholar] [CrossRef]
- Li, N.; Sancak, Y.; Frasor, J.; Atilla-Gokcumen, G.E. A protective role for triacylglycerols during apoptosis. Biochemistry 2018, 57, 72–80. [Google Scholar] [CrossRef]
- Stansell, G.R.; Gray, V.M.; Sym, S.D. Microalgal fatty acid composition: Implications for biodiesel quality. J. Appl. Phycol. 2012, 24, 791–801. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Kaewkannetra, P. Cultivation of microalga, Chlorella vulgaris under different auto-hetero-mixo trophic growths as a raw material during biodiesel production and cost evaluation. Energy 2014, 78, 4–8. [Google Scholar] [CrossRef]
- Lari, Z.; Moradi-kheibari, N.; Ahmadzadeh, H.; Abrishamchi, P.; Moheimani, N.R.; Murry, M.A. Bioprocess engineering of microalgae to optimize lipid production through nutrient management. J. Appl. Phycol. 2016, 28, 3235–3250. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Dujjanutat, P.; Muanruksa, P.; Kaewkannetra, P. Biocircular platform for third generation biodiesel production: Batch/fed batch mixotrophic cultivations of microalgae using glycerol waste as a carbon source. Biochem. Eng. J. 2021, 175, 108128. [Google Scholar] [CrossRef]
- Ryu, K.H.; Kim, B.; Lee, J.H. A model-based optimization of microalgal cultivation strategies for lipid production under photoautotrophic condition. Comput. Chem. Eng. 2019, 121, 57–66. [Google Scholar] [CrossRef]
- Dickinson, K.E.; Lalonde, C.G.; McGinn, P.J. Effects of spectral light quality and carbon dioxide on the physiology of Micractinium inermum: Growth, photosynthesis, and biochemical composition. J. Appl. Phycol. 2019, 31, 3385–3396. [Google Scholar] [CrossRef]
- Pruvost, J.; Vooren, G.V.; Gouic, B.L.; Couzinet-Mossion, A.; Legrand, J. Systematic investigation of biomass and lipid productivity by microalgae in photobioreactors for biodiesel application. Bioresour. Technol. 2011, 102, 150–158. [Google Scholar] [CrossRef]
- Quelhas, P.M.; Trovāo, M.; Silva, J.T.; Machado, A.; Santos, T.; Pereira, H.; Varela, J.; Simões, M.; Silva, J.L. Industrial production of Phaeodactylum tricornutum for CO2 mitigation: Biomass productivity and photosynthetic efficiency using photobioreactors of different volumes. J. Appl. Phycol. 2019, 31, 2187–2196. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Muylaert, K.; Foubert, I. Optimization of an analytical procedure for extraction of lipids from microalgae. J. Am. Oil Chem. Soc. 2012, 89, 189–198. [Google Scholar] [CrossRef]
- Nieva-Echevarría, B.; Goicoechea, E.; Manzanos, M.J.; Guillén, M.D. A method based on 1H NMR spectral data useful to evaluate the hydrolysis level in complex lipid mixtures. Food Res. Int. 2014, 66, 379–387. [Google Scholar] [CrossRef]
- MacDougall, K.M.; McNichol, J.; McGinn, P.J.; O’Leary, S.J.B.; Melanson, J.E. Triacylglycerol profiling of microalgae strains for biofuel feedstock by liquid chromatography-high-resolution mass spectrometry. Anal. Bioannal. Chem. 2011, 401, 2609–2616. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, Y.; Rasoul-Amini, S.; Naseri, A.T.; Montazeri-Najafabady, N.; Mobasher, M.A.; Dabbagh, F. Microalgae biofuel potentials (review). Appl. Biochem. Microbiol. 2012, 48, 126–144. [Google Scholar] [CrossRef]
- Sathya, A.B.; Thirunavukkarasu, A.; Nithya, R.; Nandan, A.; Sakthishobana, K.; Kola, A.K.; Sivashankar, R.; Tuan, H.A.; Deepanraj, B. Microalgal biofuel production: Potential challenges and prospective research. Fuel 2023, 332, 126199. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial application of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fei, X.; Deng, X. Novel molecular insights into nitrogen starvation-induced triacylglycerols accumulation revealed by differential gene expression analysis in green algae Micractinium pusillum. Biomass Bioenergy 2012, 42, 199–211. [Google Scholar] [CrossRef]
- Abu-Ghosh, S.; Ronen, B.; Feingold, D.; Banet, G.; Dubinsky, Z.; Iluz, D. Sethoxydim-resistant mutants of the thermotolerant microalga Micractinium sp. accumulate significant amounts of triacylglycerol in non-stressful conditions. J. Appl. Phycol. 2019, 31, 3433–3440. [Google Scholar] [CrossRef]
- Banskota, A.H.; Jones, A.; Hui, J.P.M.; Stefanova, R. Triacylglycerols and other lipids from hemp by-products. Molecules 2022, 27, 2339. [Google Scholar] [CrossRef]
- Marques, F.; Lopes, D.; Conde, T.; Melo, T.; Silva, J.; Abreu, M.H.; Domingues, P.; Domingues, M.R. Lipidomic characterization and antioxidant activity of macro- and microalgae blend. Life 2023, 13, 231. [Google Scholar] [CrossRef]
- Buré, C.; Solgadi, A.; Yen-Nicolaÿ, S.; Bardeau, T.; Libong, D.; Abreu, S.; Chaminade, P.; Subra-Paternault, P.; Cansell, M. Electrospray mass spectrometry as a tool to characterize phospholipid composition in plant cakes. Eur. J. Lipid. Technol. 2016, 118, 1282–1292. [Google Scholar] [CrossRef]
- Hsu, F.-F.; Turk, J. Electrospray ionization with low energy collosionally activated dissociation Tandem mass spectrometry of glycerophospholipids: Mechanisms of fragmentation and structure characterization. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2009, 877, 2673–2695. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS online tools for lipid research. Nucl. Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.L.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertases Animals; Smith, W.L., Chanely, M.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
- Park, K.C.; Whitney, C.; McNichol, J.C.; Dickinson, K.E.; MacQuarrie, S.; Skrupski, B.P.; Zou, J.; Wilson, K.E.; O’Leary, S.J.B.; McGinn, P.J. Mixotrophic and photoautotrophic cultivation of 14 microalgae isolates from Saskatchewan, Canada: Potential applications for wastewater remediation for biofuel production. J. Appl. Phycol. 2012, 24, 339–348. [Google Scholar] [CrossRef]
- Banskota, A.H.; Sperker, S.; Stefanova, R.; McGinn, P.J.; O’Leary, S.J.B. Antioxidant properties and lipid composition of selected microalgae. J. Appl. Phycol. 2019, 31, 309–318. [Google Scholar] [CrossRef]
- AOAC Official Method 991.39; Fatty Acids in Encapsulated Fish Oils and Fish Oil Methyl and Ethyl Esters, Chapter 41. AOAC International: Gaithersburg, MD, USA, 2000; pp. 26–28.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp 1 | Exp 2 | Exp 3 | Average ± SD | |
|---|---|---|---|---|
| Moisture Content | 5.6 | 6.0 | 5.9 | 5.8 ± 0.2 |
| Lipid Content | 44.0 | 44.5 | 44.0 | 44.2 ± 0.3 |
| Neutral Lipid Fraction | 79.0 | 74.4 | 78.6 | 77.3 ± 1.9 |
| Glycolipid Fraction | 14.3 | 14.2 | 11.0 | 13.2 ± 1.9 |
| Phospholipid Fraction | 4.0 | 6.2 | 6.6 | 5.6 ± 1.4 |
| TAG Relative Abundance | RT (min) | Measured (m/z) | Matched or Exact ms (m/z) | Error (ppm) * | Formula | TAG C:DB | TAG Identity |
|---|---|---|---|---|---|---|---|
| 0.41 | 1.45 | 838.6922 | 838.6919 | −0.30 | C53H88O6 | 50:7 | 18:3/16:3/16:1; 18:3/18:2/16:2; 18:2/16:3/16:2 |
| 0.60 | 1.45 | 864.7083 | 864.7076 | −0.80 | C55H90O6 | 52:8 | 18:3/18:1/16:4; 18:3/18:2/16:3; 18 |
| 0.77 | 1.67 | 930.7733 | 930.7756 | 2.48 | C57H100O8 | 54:5; O2 | NI |
| 2.21 | 1.67 | 840.7084 | 840.7076 | −0.95 | C53H90O6 | 50:6 | 18:3/16:3/16:0; 18:3/16:2/16:1; 18:2/16:3/16:1; 18:2/16:4/16:0 |
| 3.28 | 1.71 | 866.7239 | 866.7232 | −0.81 | C55H92O6 | 52:7 | 18:3/18:1/16:3; 18:2/18:1/16:4 |
| 0.69 | 1.94 | 894.7557 | 894.7545 | −1.34 | C57H96O6 | 54:7 | 18:3/18:2/18:3; 18:3/18:3/18:1 |
| 1.06 | 1.90 | 932.7922 | 932.7913 | −0.96 | C57H102O8 | 54:4; O2 | NI |
| 0.32 | 2.10 | 816.7093 | 816.7076 | −2.08 | C51H90O6 | 48:4 | 16:4/16:0/16:0; 16:3:16:1/16:0; 16:2/16:2/16:0 |
| 2.00 | 2.05 | 842.7249 | 842.7232 | −2.02 | C53H92O6 | 50:5 | 18:3/16:2/16:0; 18:3/16:1/16:1; 18:2/16:3/16:0; 18:2/16:2/16:1; 18:1/16:3/16:2; 18:1/16:4/16:0 |
| 3.09 | 2.06 | 868.7404 | 868.7389 | −1.73 | C55H94O6 | 52:6 | 18:3/18:2/16:1; 18:3/18:3/16:0; 18:1/18:0/16:5; 18:3/18:1/16:2; 18:1/18:1/16:4; 18:2/18:1/16:3 |
| 1.35 | 2.12 | 934.8069 | 934.8069 | 0.00 | C57H104O8 | 54:3;O2 | NI |
| 0.68 | 2.35 | 818.7235 | 818.7232 | −0.37 | C51H92O6 | 48:3 | 16:3/16:0/16:0; 16:2/16:1/16:0 |
| 3.26 | 2.35 | 844.7389 | 844.7389 | 0.00 | C53H94O6 | 50:4 | 18:1/16:3/16:0; 18:1/16:2/16:1; 18:2/16:2/16:0; 18:2/16:1/16:1; 18:3/16:1/16:0 |
| 5.08 | 2.35 | 870.7545 | 870.7545 | 0.00 | C55H96O6 | 52:5 | 18:3/18:2/16:0; 18:2/18:2/16:1; 18:3/18:1/16:1; 18:2/18:1/16:2; 18:1/18:1/16:3; 18:0/16:2/16:3 |
| 3.40 | 2.35 | 896.7697 | 896.7702 | 0.56 | C57H98O6 | 54:6 | 18:3/18:2/18:1 |
| 0.23 | 2.62 | 820.7403 | 820.7389 | −1.71 | C51H94O6 | 48:2 | 16:2/16:0/16:0 (major); 16:1/16:1/16:0 (minor); 18:2/16:0/14:0; 18:1/16:0/14:1; 18:1/16:1/14:0 |
| 4.70 | 2.75 | 898.7864 | 898.7858 | −0.67 | C57H100O6 | 54:5 | 18:3/18:1/18:1; 18:3/18:2/18:0; 18:2/18:2/18:1 |
| 1.75 | 2.78 | 846.7557 | 846.7545 | −1.42 | C53H96O6 | 50:3 | 18:3/16:0/16:0; 18:2/16:1/16:0; 18:1/16:1/16:1; 18:0/16:2/16:1 |
| 5.48 | 2.81 | 872.7709 | 872.7702 | −0.80 | C55H98O6 | 52:4 | 18:3/18:1/16:0; 18:2/18:2/16:0; 18:3/18:0/16:1; 18:3/18:0/16:1; 18:2/18:0/16:2; 18:1/18:0/16:3 |
| 11.46 | 3.07 | 900.8002 | 900.8015 | 1.44 | C57H102O6 | 54:4 | 18:2/18:2/18:0 (major); 18:3/18:1/18:0 (minor) |
| 13.01 | 3.07 | 874.7837 | 874.7858 | 2.40 | C55H100O6 | 52:3 | 18:2/18:1/16:0 (major); 18:2/18:0/16:1 (minor); 18:1/18:0/16:2 (minor) |
| 0.06 | 3.28 | 836.7695 | 836.7702 | 0.84 | C52H98O6 | 49:1 | 18:1/16:0/15:0 |
| 3.50 | 3.07 | 848.7690 | 848.7702 | 1.41 | C53H98O6 | 50:2 | 18:2/16:0/16:0; 18:1/16:1/16:0 |
| 0.43 | 3.28 | 862.7849 | 862.7858 | 1.04 | C54H100O6 | 51:2 | 18:1/17:1:16:0; 18:2/17:0:16:0; |
| 0.63 | 3.28 | 888.8000 | 888.8015 | 1.69 | C56H102O6 | 53:3 | 18:2:/18:1/17:0 |
| 3.22 | 3.50 | 850.7862 | 850.7858 | −0.47 | C53H100O6 | 50:1 | 18:1/16:0/16:0 |
| 0.25 | 3.66 | 864.8025 | 864.8015 | −1.16 | C54H102O6 | 51:1 | 18:1/17:0/16:0 |
| 12.77 | 3.50 | 876.8016 | 876.8015 | −0.11 | C55H102O6 | 52:2 | 18:1/18:1/16:0 |
| 0.59 | 3.73 | 890.8178 | 890.8171 | −0.79 | C56H104O6 | 53:2 | 18:1/18:1/17:0 |
| 11.03 | 3.50 | 902.8167 | 902.8171 | 0.44 | C57H104O6 | 54:3 | 18:1/18:1/18:1 (major); 18:2/18:1/18:0 (minor) |
| 0.97 | 3.90 | 878.8174 | 878.8171 | −0.34 | C55H104O6 | 52:1 | 18:1/18:0/16:0 |
| 0.16 | 3.96 | 930.8485 | 930.8484 | −0.11 | C59H108O6 | 56:3 | 20:1/18:1/18:1 ** |
| 1.47 | 3.96 | 904.8329 | 904.8328 | −0.11 | C57H106O6 | 54:2 | 18:1/18:1/18:0 (major); 18:2/18:0/18:0 (minor) |
 High abundance.
High abundance.Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banskota, A.H.; Hui, J.P.M.; Jones, A.; McGinn, P.J. Characterization of Neutral Lipids of the Oleaginous Alga Micractinum inermum. Molecules 2024, 29, 359. https://doi.org/10.3390/molecules29020359
Banskota AH, Hui JPM, Jones A, McGinn PJ. Characterization of Neutral Lipids of the Oleaginous Alga Micractinum inermum. Molecules. 2024; 29(2):359. https://doi.org/10.3390/molecules29020359
Chicago/Turabian StyleBanskota, Arjun H., Joseph P. M. Hui, Alysson Jones, and Patrick J. McGinn. 2024. "Characterization of Neutral Lipids of the Oleaginous Alga Micractinum inermum" Molecules 29, no. 2: 359. https://doi.org/10.3390/molecules29020359
APA StyleBanskota, A. H., Hui, J. P. M., Jones, A., & McGinn, P. J. (2024). Characterization of Neutral Lipids of the Oleaginous Alga Micractinum inermum. Molecules, 29(2), 359. https://doi.org/10.3390/molecules29020359