
A Comparison of Phenolic, Flavonoid, and Amino Acid Compositions and In Vitro Antioxidant and Neuroprotective Activities in Thai Plant Protein Extracts



Abstract
1. Introduction
2. Results
2.1. Amino Acid Profiles
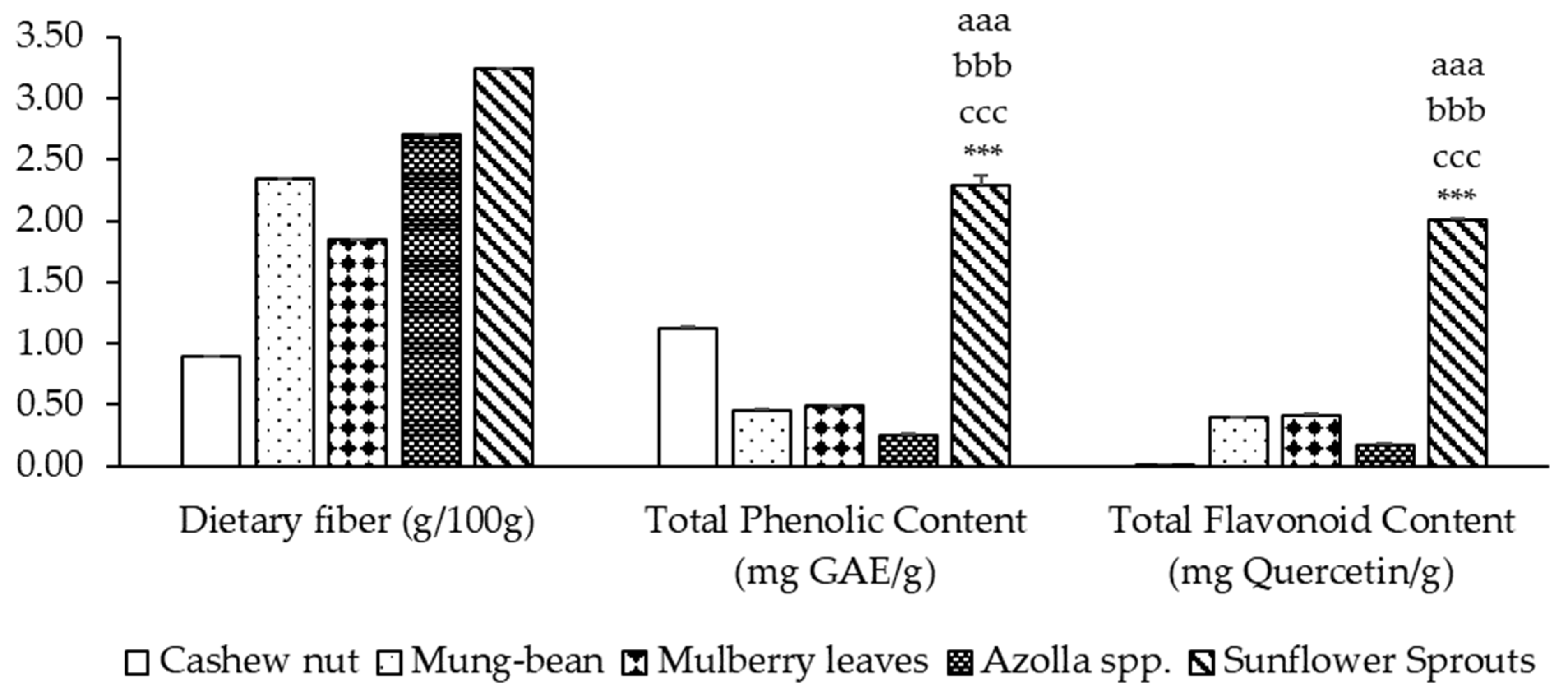
2.2. Dietary Fiber, Total Phenolic Content (TPC), and Total Flavonoid Content (TFC)
2.3. Antioxidant Activity
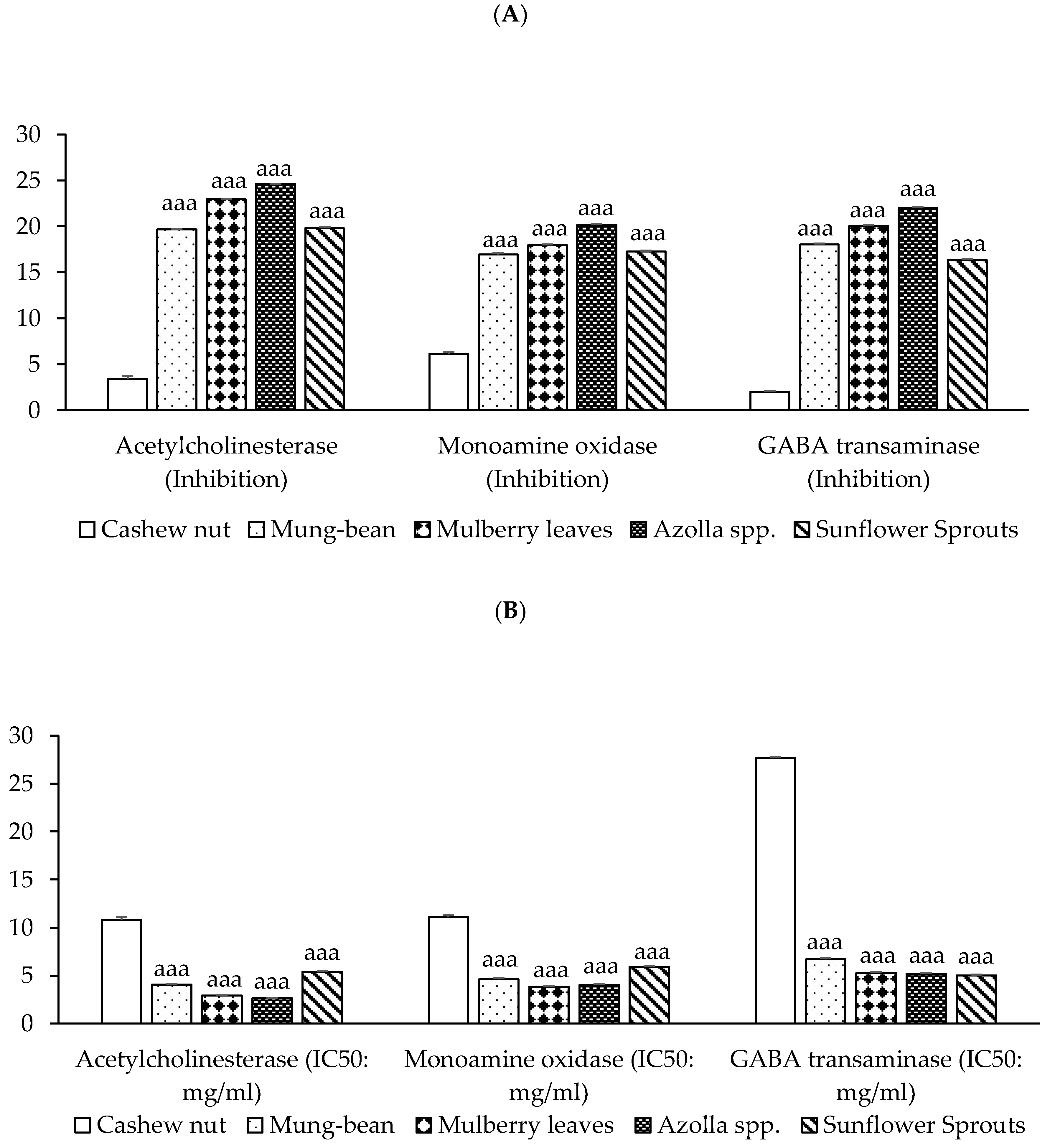
2.4. Neuroprotective Activity
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extractions
4.2. Identification of the Amino Acid Profile
4.3. Determination of Dietary Fiber
4.4. Determination of Total Phenolic Contents
4.5. Determination of Total Flavonoid Content
4.6. Determination of Antioxidant Activity
4.6.1. The 1,1-Diphenyl-2-picryl hydrazyl Radical (DPPH) Assay
4.6.2. The 2,2′-azinobis-3-ethylbenzothiazoline-6-sulfonic Acid (ABTS) Assay
4.7. Determination of Neuroprotective Activity
4.7.1. Acetylcholinesterase (AChE) Inhibition Assay
4.7.2. Monoamine Oxidase (MAO) Inhibition Assay
4.7.3. γ-Aminobutyric Acid Transaminase (GABA-T) Inhibition
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alcorta, A.; Porta, A.; Tárrega, A.; Alvarez, M.D.; Pilar Vaquero, M. Foods for Plant-Based Diets: Challenges and Innovations. Foods 2021, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Wootton-Beard, P.C.; Ryan, L. Improving Public Health?: The Role of Antioxidant-Rich Fruit and Vegetable Beverages. Food Res. Int. 2011, 44, 3135–3148. [Google Scholar] [CrossRef]
- Dreiseitel, A.; Korte, G.; Schreier, P.; Oehme, A.; Locher, S.; Domani, M.; Hajak, G.; Sand, P.G. Berry Anthocyanins and Their Aglycons Inhibit Monoamine Oxidases A and B. Pharmacol. Res. 2009, 59, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Han, X.H.; Hong, S.S.; Hwang, J.S.; Lee, M.K.; Hwang, B.Y.; Ro, J.S. Monoamine Oxidase Inhibitory Components from Cayratia Japonica. Arch. Pharm. Res. 2007, 30, 13–17. [Google Scholar] [CrossRef]
- Papandreou, M.A.; Dimakopoulou, A.; Linardaki, Z.I.; Cordopatis, P.; Klimis-Zacas, D.; Margarity, M.; Lamari, F.N. Effect of a Polyphenol-Rich Wild Blueberry Extract on Cognitive Performance of Mice, Brain Antioxidant Markers and Acetylcholinesterase Activity. Behav. Brain Res. 2009, 198, 352–358. [Google Scholar] [CrossRef]
- Qoms, M.S.; Arulrajah, B.; Shamsudin, R.; Ibadullah, W.Z.W.; Saari, N. Valorization of Green Biomass Azolla Pinnata Fern: Multi-Parameter Evaluation of Processing Conditions on Protein Extractability and Their Influence on the Physicochemical, Structural, Techno-Functional Properties and Protein Quality. J. Sci. Food Agric. 2022, 102, 6974–6983. [Google Scholar] [CrossRef]
- Mosha, S.S. A Review on Significance of Azolla Meal as a Protein Plant Source in Finfish Culture. J. Aquac. Res. Dev. 2018, 9, 7. [Google Scholar] [CrossRef]
- Qian, W.; Wu, W.; Kang, Y.; Wang, Y.; Yang, P.; Deng, Y.; Ni, C.; Huang, J. Comprehensive Identification of Minor Components and Bioassay-Guided Isolation of an Unusual Antioxidant from Azolla Imbricata Using Ultra-High Performance Liquid Chromatography—Quadrupole Time-of-Flight Mass Spectrometry Combined with Multicomponent Knoc. J. Chromatogr. A 2020, 1609, 460435. [Google Scholar] [CrossRef]
- Kunjiappan, S.; Bhattacharjee, C.; Chowdhury, R. In Vitro Antioxidant and Hepatoprotective Potential of Azolla Microphylla Phytochemically Synthesized Gold Nanoparticles on Acetaminophen—Induced Hepatocyte Damage in Cyprinus carpio L. Vitr. Cell. Dev. Biol.-Anim. 2015, 51, 630–643. [Google Scholar] [CrossRef]
- Bhaskaran, S.K.; Kannappan, P.; Muneeswari, P.; Madathil, R. Toxicological Evaluation of the Repeated Dose Administration of the Ethanolic Extract of Azolla Microphylla in Wistar Albino Rats. Toxicol. Int. 2021, 28, 39–48. [Google Scholar] [CrossRef]
- Iqbal, S.; Younas, U.; Sirajuddin; Chan, K.W.; Sarfraz, R.A.; Uddin, M.K. Proximate Composition and Antioxidant Potential of Leaves from Three Varieties of Mulberry (Morus Sp.): A Comparative Study. Int. J. Mol. Sci. 2012, 13, 6651–6664. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, L.; Dai, Y.; Li, J.; Li, F.; Li, Q.; Yu, Z.; Chai, K.; Zhu, Y. Differential Metabolomics and Network Pharmacology Analysis of Silkworm Biotransformation between Mulberry Leaves and Silkworm Droppings. Evid.-Based Complement. Altern. Med. 2021, 2021, 8819538. [Google Scholar] [CrossRef] [PubMed]
- Wattanathorn, J.; Somboonporn, W.; Thukham-Mee, W.; Sungkamnee, S. Memory-Enhancing Effect of 8-Week Consumption of the Quercetin-Enriched Culinary Herbs-Derived Functional Ingredients: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Foods 2022, 11, 2678. [Google Scholar] [CrossRef]
- Hou, D.; Yousaf, L.; Xue, Y.; Hu, J.; Wu, J.; Hu, X.; Feng, N.; Shen, Q. Mung Bean (Vigna radiata L.): Bioactive Polyphenols, Polysaccharides, Peptides, and Health Benefits. Nutrients 2019, 11, 1238. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Yao, Y.; Zhu, Y.; Ren, G. Nutritional Composition and Antioxidant Activity of Twenty Mung Bean Cultivars in China. Crop J. 2016, 4, 398–406. [Google Scholar] [CrossRef]
- Li, G.H.; Le, G.W.; Liu, H.; Shi, Y.H. Mung-Bean Protein Hydrolysates Obtained with Alcalase Exhibit Angiotensin I-Converting Enzyme Inhibitory Activity. Food Sci. Technol. Int. 2005, 11, 281–287. [Google Scholar] [CrossRef]
- Wu, M.; Muchimapura, S.; Wattanathorn, J.; Thukhummee, W.; On-Ong-Arj, P.; Wannanon, P. Mung Bean-Derived Protein Protects against Neurodegeneration and Memory Impairment in Animal Model of Menopause with Obesity. J. Med. Assoc. Thail. 2019, 102, 43. [Google Scholar]
- Wattanathorn, J.; Thukham-Mee, W.; Muchimapura, S.; Wannanon, P.; Tong-Un, T.; Tiamkao, S. Preventive Effect of Cashew-Derived Protein Hydrolysate with High Fiber on Cerebral Ischemia. Biomed Res. Int. 2017, 2017, 6135023. [Google Scholar] [CrossRef] [PubMed]
- Ebert, A.W. Sprouts and Microgreens—Novel Food Sources for Healthy Diets. Plants 2022, 11, 571. [Google Scholar] [CrossRef]
- Pająk, P.; Socha, R.; Gałkowska, D.; Rożnowski, J.; Fortuna, T. Phenolic Profile and Antioxidant Activity in Selected Seeds and Sprouts. Food Chem. 2014, 143, 300–306. [Google Scholar] [CrossRef]
- Guo, S.; Ge, Y.; Na Jom, K. A Review of Phytochemistry, Metabolite Changes, and Medicinal Uses of the Common Sunflower Seed and Sprouts (Helianthus annuus L.). Chem. Cent. J. 2017, 11, 95. [Google Scholar] [CrossRef]
- Koodkaew, I.; Tungkasem, B. Effects of the Application Methods and Concentrations of Sucrose Solution on Growth and Antioxidant Activity of Sunflower Sprouts. Burapha Sci. J. 2021, 26, 561–571. [Google Scholar]
- Sun, A.Y.; Chen, Y.M. Oxidative Stress and Neurodegenerative Disorders. J. Biomed. Sci. 1998, 5, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Koleva, I.I.; Van Beek, T.A.; Linssen, J.P.H.; De Groot, A.; Evstatieva, L.N. Screening of Plant Extracts for Antioxidant Activity: A Comparative Study on Three Testing Methods. Phytochem. Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef]
- Ingkaninan, K.; Temkitthawon, P.; Chuenchom, K.; Yuyaem, T.; Thongnoi, W. Screening for Acetylcholinesterase Inhibitory Activity in Plants Used in Thai Traditional Rejuvenating and Neurotonic Remedies. J. Ethnopharmacol. 2003, 89, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zou, D.; Li, Y.; Gu, S.; Dong, J.; Ma, X.; Xu, S.; Wang, F.; Huang, J.H. Monoamine Neurotransmitters Control Basic Emotions and Affect Major Depressive Disorders. Pharmaceuticals 2022, 15, 1203. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.D.; Cheng, C.H.K.; Tan, R.X. Inhibition of MAO a and B by Some Plant-Derived Alkaloids, Phenols and Anthraquinones. J. Ethnopharmacol. 2004, 91, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Tarahi, M.; Abdolalizadeh, L.; Hedayati, S. Mung Bean Protein Isolate: Extraction, Structure, Physicochemical Properties, Modifications, and Food Applications. Food Chem. 2024, 444, 138626. [Google Scholar] [CrossRef] [PubMed]
- Joint WHO/FAO/UNU Expert Consultation. Protein and Amino Acid Requirements in Human Nutrition. World Health Organ Tech Rep Ser. 2007, 1–265. [Google Scholar]
- Xu, N.; Chen, G.; Liu, H. Antioxidative Categorization of Twenty Amino Acids Based on Experimental Evaluation. Molecules 2017, 22, 2066. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, Y.; Shi, Z.; Zhou, N.; Ren, G.; Hao, X.; Zou, L.; Yao, Y. Mung bean protein suppresses undernu-trition-induced growth deficits and cognitive dysfunction in rats via gut microbiota-TLR4/NF-kB pathway. J. Agric. Food Chem. 2021, 69, 12566–12577. [Google Scholar] [CrossRef] [PubMed]
- Pal, M.M. Glutamate: The Master Neurotransmitter and Its Implications in Chronic Stress and Mood Disorders. Front. Hum. Neurosci. 2021, 15, 722323. [Google Scholar] [CrossRef] [PubMed]
- Tarditi, A.M.; Klipfel, M.W.; Rodriguez, A.M.; Suvire, F.D.; Chasse, G.A.; Farkas, O.; Perczel, A.; Enriz, R.D. An ab initio exploratory study of side chain conformations for selected backbone conformations of N-acetyl-l-glutamine-N-methylamide. J. Mol. Struct. Theochem. 2001, 545, 29–47. [Google Scholar] [CrossRef]
- Xu, H.; Zhou, Q.; Liu, B.; Cheng, K.W.; Chen, F.; Wang, M. Neuroprotective Potential of Mung Bean (Vigna radiata L.) Polyphenols in Alzheimer’s Disease: A Review. J. Agric. Food Chem. 2021, 69, 11554–11571. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Mantas, A.; Deretey, E.; Ferretti, F.H.; Estrada, M.; Csizmadia, I.G. Ab initio conformational analysis of flavone and related compounds. J. Mol. Struct. Theochem. 2000, 504, 77–103. [Google Scholar] [CrossRef]
- Ichikawa, K.; Okabayashi, T.; Shima, Y.; Iiyama, T.; Takezaki, Y.; Munekage, M.; Namikawa, T.; Sugimoto, T.; Kobayashi, M.; Mimura, T.; et al. Branched-Chain Amino Acid-Enriched Nutrients Stimulate Antioxidant DNA Repair in a Rat Model of Liver Injury Induced by Carbon Tetrachloride. Mol. Biol. Rep. 2012, 39, 10803–10810. [Google Scholar] [CrossRef]
- Ullah, R.; Jo, M.H.; Riaz, M.; Alam, S.I.; Saeed, K.; Ali, W.; Rehman, I.U.; Ikram, M.; Kim, M.O. Glycine, the Smallest Amino Acid, Confers Neuroprotection against d-Galactose-Induced Neurodegeneration and Memory Impairment by Regulating c-Jun N-Terminal Kinase in the Mouse Brain. J. Neuroinflam. 2020, 17, 303. [Google Scholar] [CrossRef]
- Berding, K.; Carbia, C.; Cryan, J.F. Going with the grain: Fiber, cognition, and the microbiota-gut-brain-axis. Exp. Biol. Med. 2021, 246, 796–811. [Google Scholar] [CrossRef]
- Besten, G.D.; Eunen, K.V.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- McRae, M.P. Therapeutic Benefits of l-Arginine: An Umbrella Review of Meta-analyses. J. Chiropr. Med. 2016, 15, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Akomolafe, S.F.; Asowata-Ayodele, A.M. Roasted cashew (Anacardium occidentale L.) nut-enhanced diet forestalls cisplatin-initiated brain harm in rats. Heliyon 2022, 8, e11066. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.D.C.Q.; Madruga, M.S.; Almeida, G.H.O.; de Melo, M.F.F.T.; Viera, V.B.; Bertozzo, C.C.D.M.S.; Dutra, L.M.G.; Alves, A.P.V.; Dantas, F.A.; Bezerra, J.K.G.; et al. Consumption of cashew nut induced anxiolytic-like behavior in dyslipidemic rats consuming a high fat diet. Behav. Brain Res. 2023, 453, 114634. [Google Scholar] [CrossRef] [PubMed]
- Tam, D.N.H.; Nam, N.H.; Elhady, M.T.; Tran, L.; Hassan, O.G.; Sadik, M.; Tien, P.T.M.; Elshafei, G.A.; Huy, N.T. Effects of Mulberry on The Central Nervous System: A Literature Review. Curr. Neuropharmacol. 2021, 19, 193–219. [Google Scholar] [CrossRef] [PubMed]
- Ogunwolu, S.O.; Henshaw, F.O.; Mock, H.P.; Santros, A.; Awonorin, S.O. Functional Properties of Protein Concentrates and Isolates Produced from Cashew (Anacardium Occidentale L.) Nut. Food Chem. 2009, 115, 852–858. [Google Scholar] [CrossRef]
- Quettier-Deleu, C.; Gressier, B.; Vasseur, J.; Dine, T.; Brunet, C.; Luyckx, M.; Cazin, M.; Cazin, J.C.; Bailleul, F.; Trotin, F. Phenolic Compounds and Antioxidant Activities of Buckwheat (Fagopyrum Esculentum Moench) Hulls and Flour. J. Ethnopharmacol. 2000, 72, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Ebrahimzadeh, M.A.; Nabavi, S.M.; Nabavi, S.F.; Bahramian, F.; Bekhradnia, A.R. Antioxidant and Free Radical Scavenging Activity of H. Officinalis L. Var. Angustifolius, V. Odorata, B. Hyrcana and C. Speciosum. Pak. J. Pharm. Sci. 2010, 23, 29–34. [Google Scholar]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A Novel Method for Measuring Antioxidant Capacity and Its Application to Monitoring the Antioxidant Status in Premature Neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Holt, A.; Sharman, D.F.; Baker, G.B.; Palcic, M.M. A Continuous Spectrophotometric Assay for Monoamine Oxidase and Related Enzymes in Tissue Homogenates. Anal. Biochem. 1997, 244, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.J.; Lippert, B.; Metcalf, B.W.; Schechter, P.J.; Böhlen, P.; Sjoerdsma, A. The effect of 4-amino hex-5-ynoic acid (γ-acetylenic gaba, γ-ethynyl gaba) a catalytic inhibitor of gaba transaminase, on brain gaba metabolism in vivo. J. Neurochem. 1977, 28, 717–723. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Amino Acid Profile | Protein Concentrate (mg/100 g Protein) | |||||
|---|---|---|---|---|---|---|
| Soy Protein | Cashew Nut | Mung Bean | Mulberry Leaves | Azolla spp. | Sunflower Sprouts | |
| Essential amino acid | ||||||
| Threonine (Thr) | 2474 | 1302 | 2733 | 715 | 4487 | 5026 |
| Methionine (Met) | 814 | 651 | 1068 | 221 | 2243 | 1400 |
| Phenylalanine (Phe) | 3278 | 2605 | 6110 | 800 | 4829 | 5358 |
| Histidine (His) | 1578 | 3257 | 7362 | 1345 | 1414 | 2464 |
| Lysine (Lys) | 3929 | 2931 | 6736 | 1030 | 5853 | 5334 |
| Valine (Val) | 3064 | 2280 | 4134 | 810 | 5121 | 6996 |
| Isoleucine (Ile) | 2942 | 1302 | 3170 | 565 | 3512 | 4793 |
| Leucine (Leu) | 4917 | 4560 | 8023 | 1425 | 8926 | 8640 |
| Tryptophan (Trp) | 835 | ND | 664 | 605 | 682 | 1540 |
| Arginine (Arg) | 4642 | 7817 | 12,155 | 1415 | 5024 | 5831 |
| Total EAAs | 28,473 | 26,710 | 52,161 | 8931 | 42,097 | 47,386 |
| Non-essential amino acid | ||||||
| Serine (Ser) | 3369 | 2931 | 5434 | 675 | 5024 | 4625 |
| Glycine (Gly) | 2688 | 2280 | 1739 | 408 | 4829 | 5530 |
| Glutamic acid (Glu) | 12,013 | 12,377 | 15,269 | 1440 | 10,000 | 13,254 |
| Proline (Pro) | 3298 | 2931 | 4817 | 925 | 5658 | 4528 |
| Cysteine (Cys) | 886 | 1628 | 473 | ND | 1609 | ND |
| Alanine (Ala) | 2677 | 1628 | 3970 | 945 | 6390 | 6247 |
| Tyrosine (Tyr) | 2301 | 4234 | 2416 | 482 | 2829 | 3381 |
| Aspartic acid | 7249 | 1302 | 12,245 | 1735 | 8975 | 10,504 |
| Total NEAAs | 34,481 | 29,315 | 46,366 | 6611 | 45,316 | 48,073 |
| HAA | 24,513 | 18,241 | 33,700 | 6704 | 42,195 | 45,036 |
| AAA | 5579 | 6840 | 8526 | 1282 | 7658 | 10,281 |
| TAA | 62,954 | 56,026 | 98,527 | 15,542 | 87,414 | 95,460 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polyiam, P.; Thukhammee, W. A Comparison of Phenolic, Flavonoid, and Amino Acid Compositions and In Vitro Antioxidant and Neuroprotective Activities in Thai Plant Protein Extracts. Molecules 2024, 29, 2990. https://doi.org/10.3390/molecules29132990
Polyiam P, Thukhammee W. A Comparison of Phenolic, Flavonoid, and Amino Acid Compositions and In Vitro Antioxidant and Neuroprotective Activities in Thai Plant Protein Extracts. Molecules. 2024; 29(13):2990. https://doi.org/10.3390/molecules29132990
Chicago/Turabian StylePolyiam, Pontapan, and Wipawee Thukhammee. 2024. "A Comparison of Phenolic, Flavonoid, and Amino Acid Compositions and In Vitro Antioxidant and Neuroprotective Activities in Thai Plant Protein Extracts" Molecules 29, no. 13: 2990. https://doi.org/10.3390/molecules29132990
APA StylePolyiam, P., & Thukhammee, W. (2024). A Comparison of Phenolic, Flavonoid, and Amino Acid Compositions and In Vitro Antioxidant and Neuroprotective Activities in Thai Plant Protein Extracts. Molecules, 29(13), 2990. https://doi.org/10.3390/molecules29132990