
Unraveling the Binding Mode of Cyclic Adenosine–Inosine Monophosphate (cAIMP) to STING through Molecular Dynamics Simulations

 , , , , , ,
, , , , , ,
Abstract

1. Introduction
2. Results and Discussion
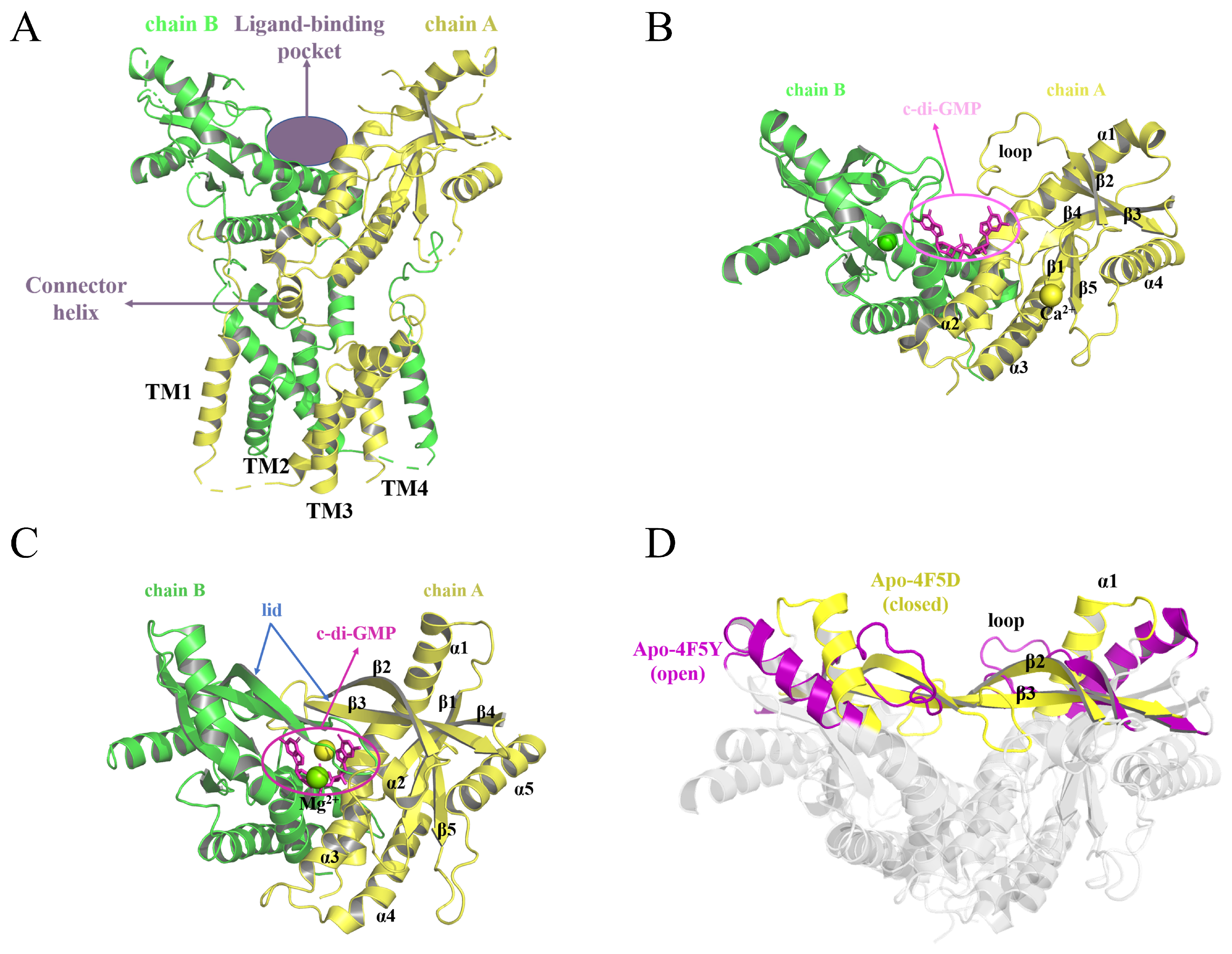
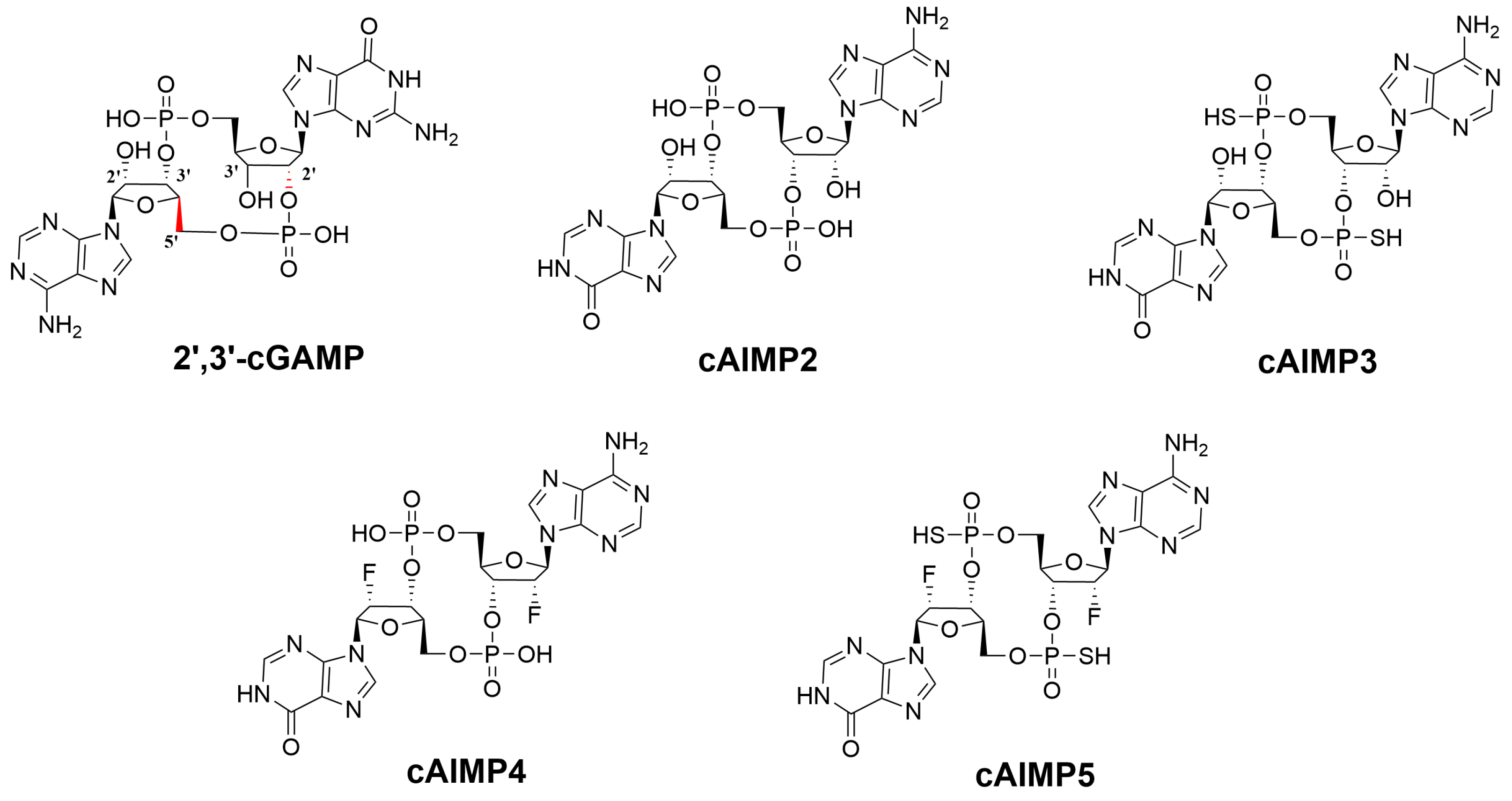
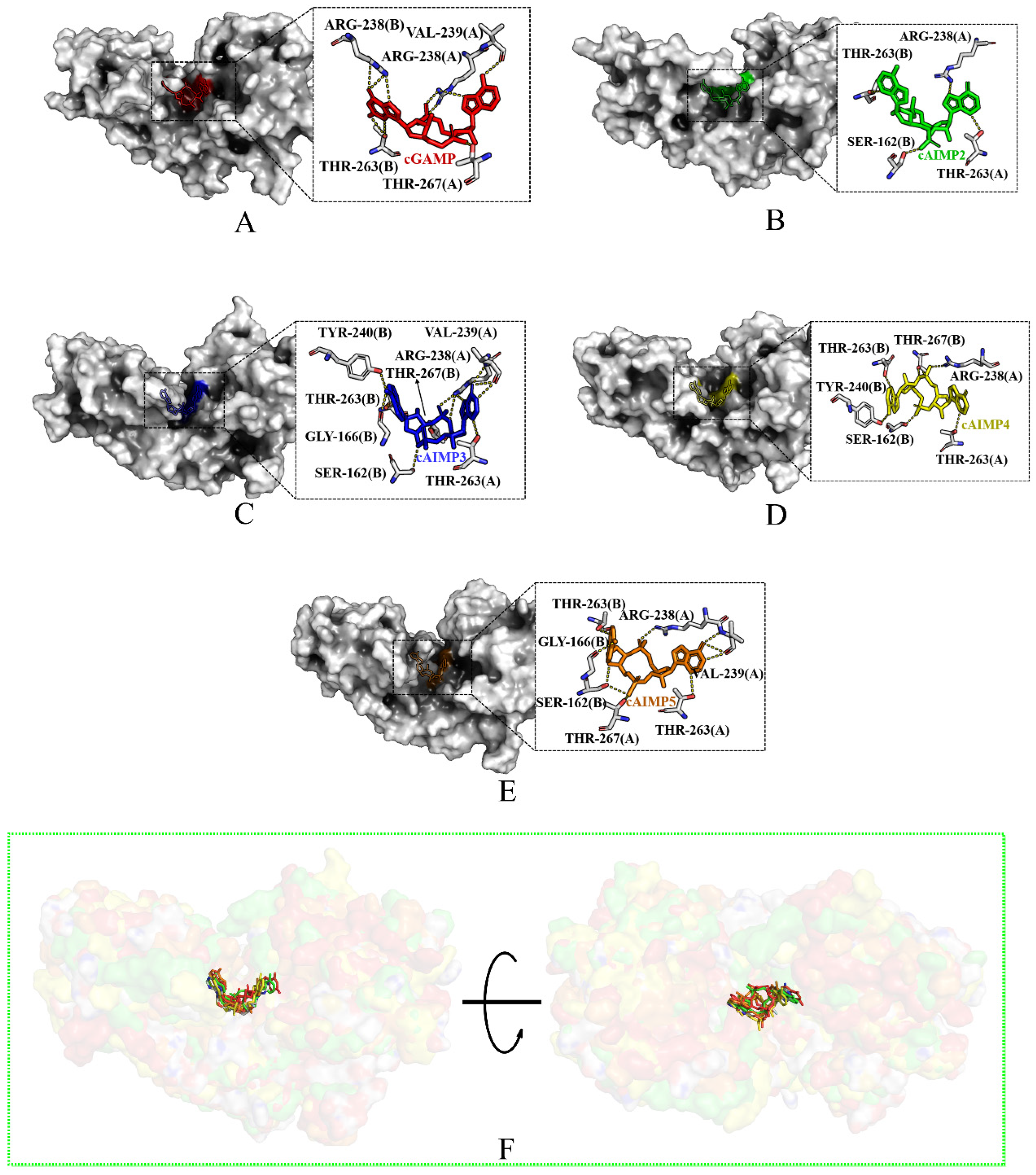
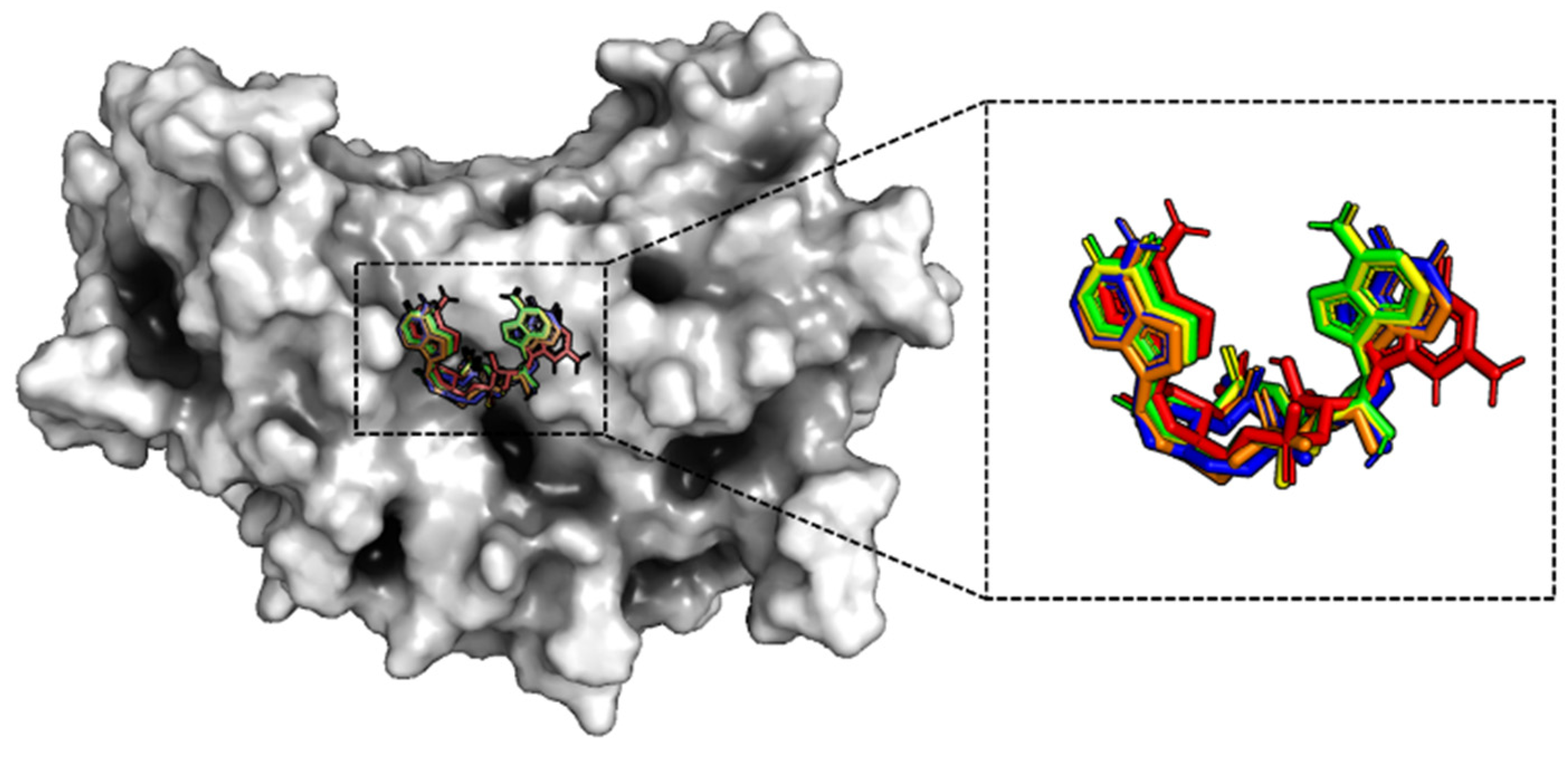
2.1. Binding Mode
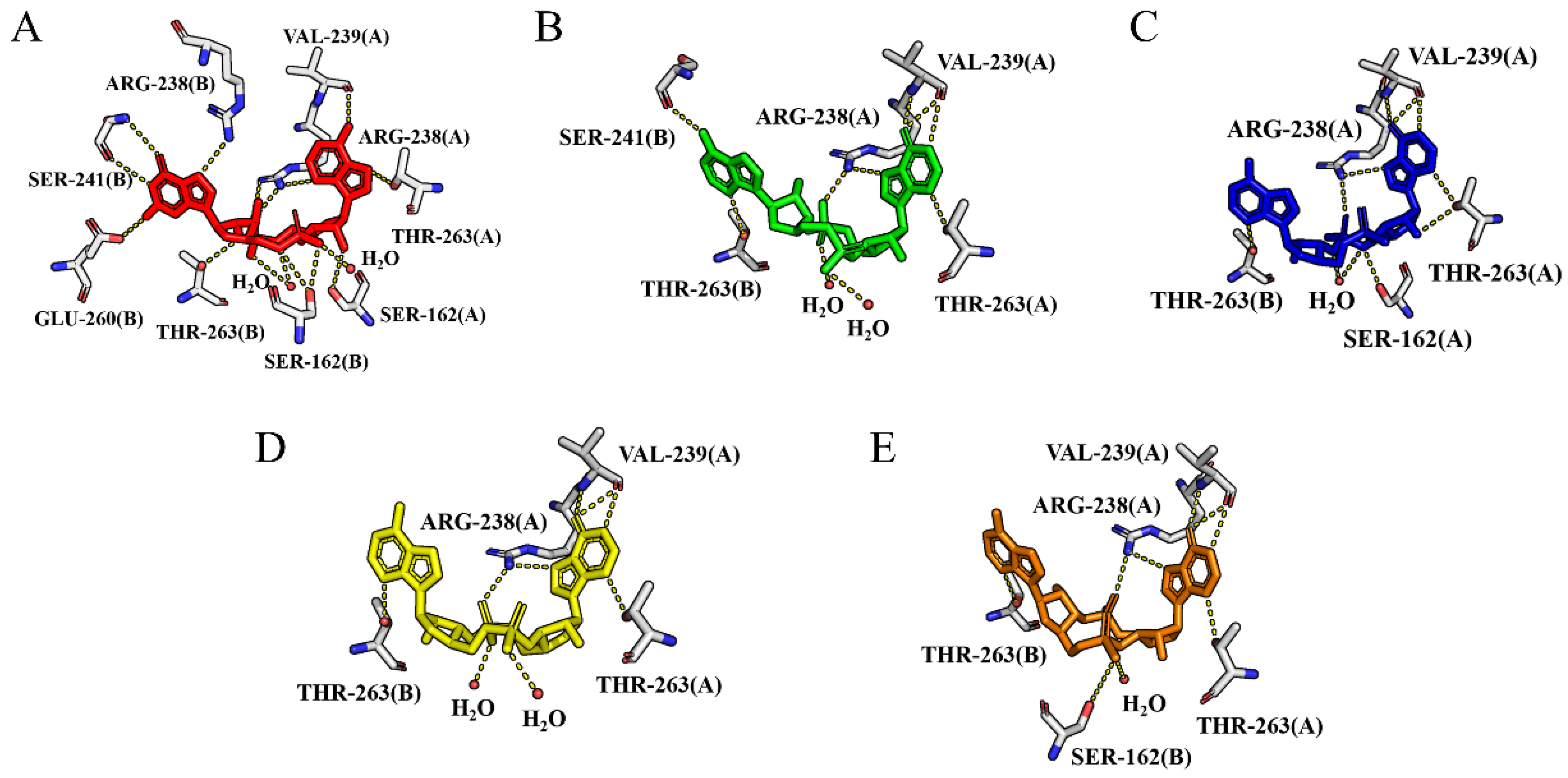
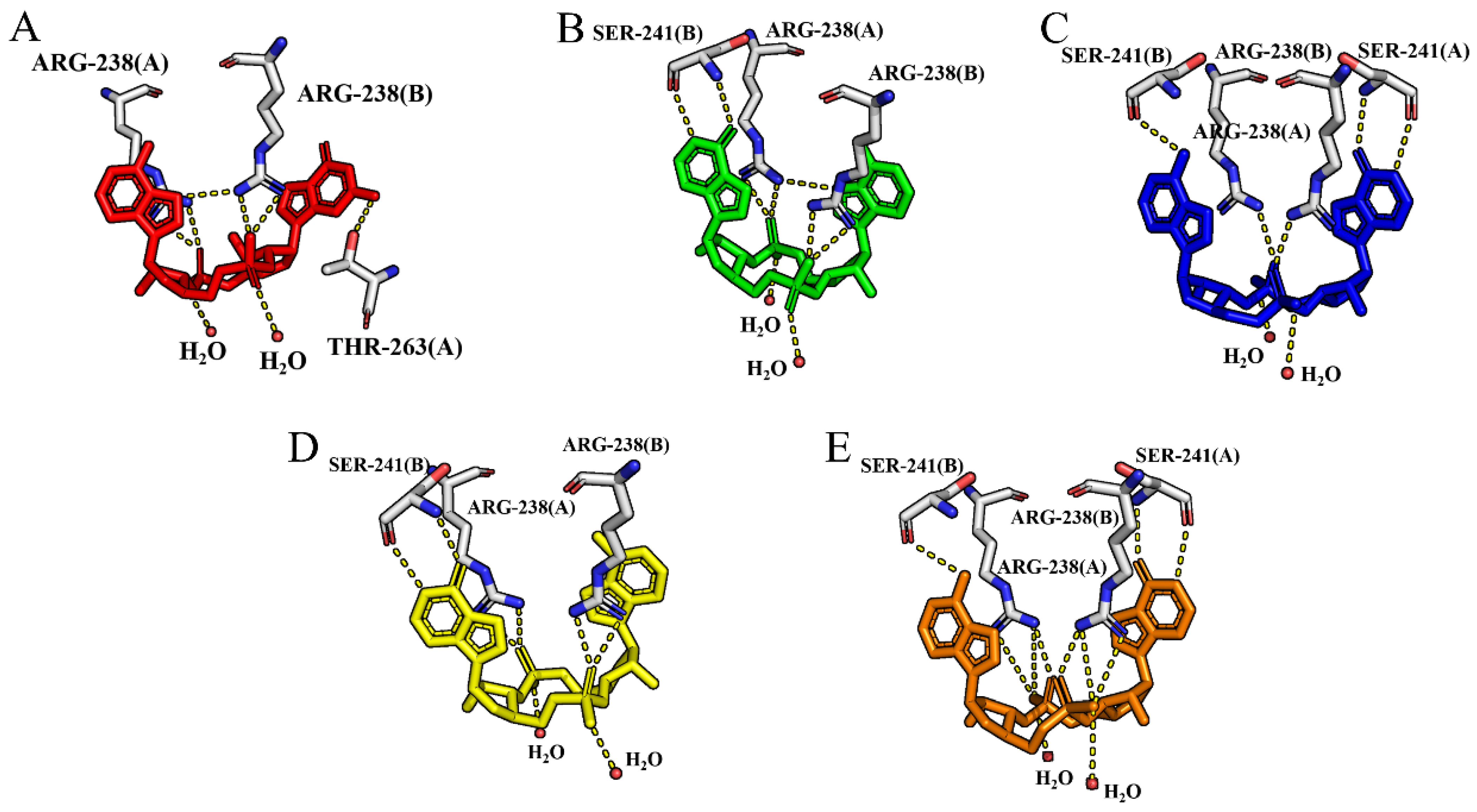
2.1.1. Binding to the Open State
2.1.2. Binding to the Closed State
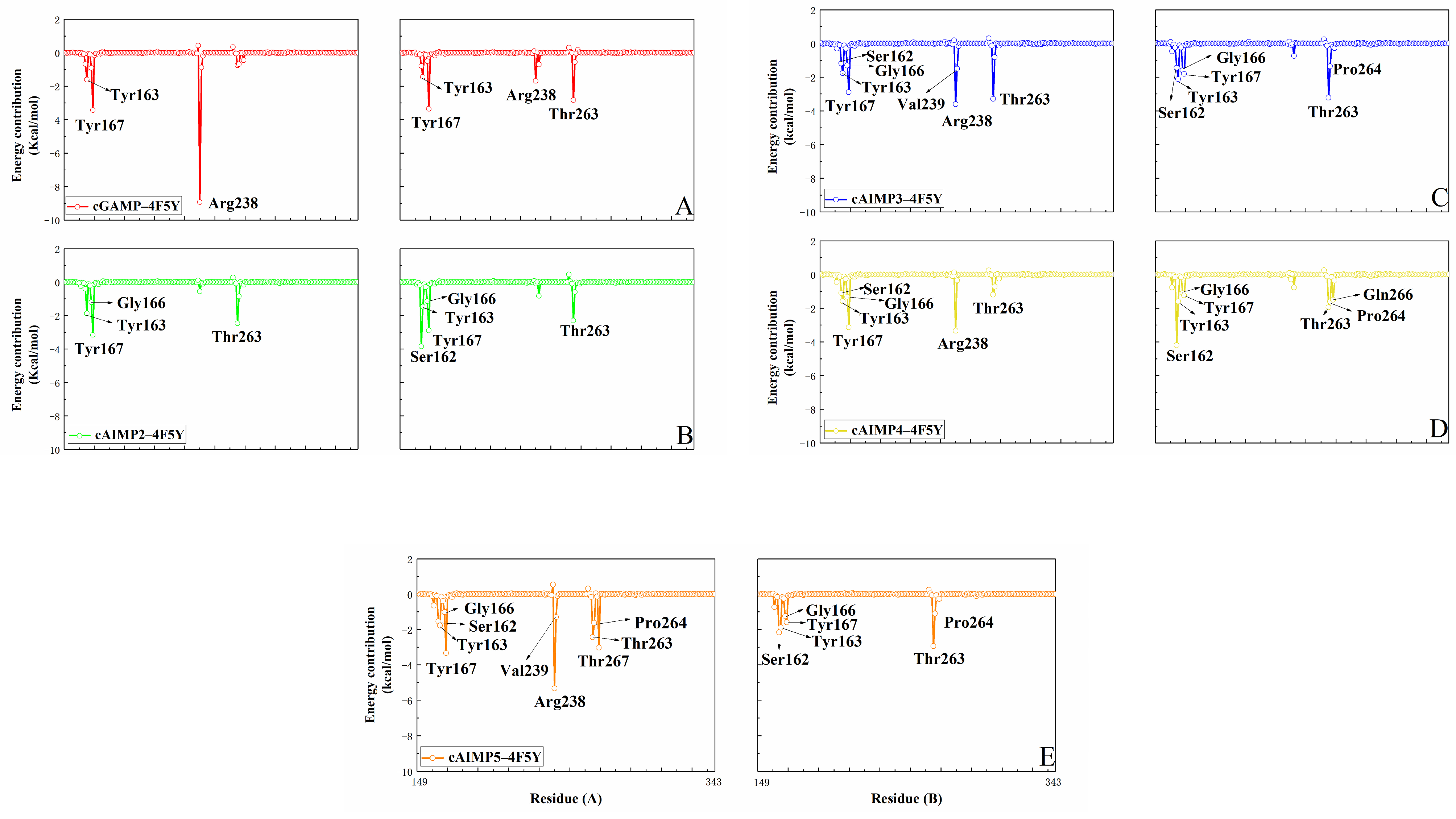
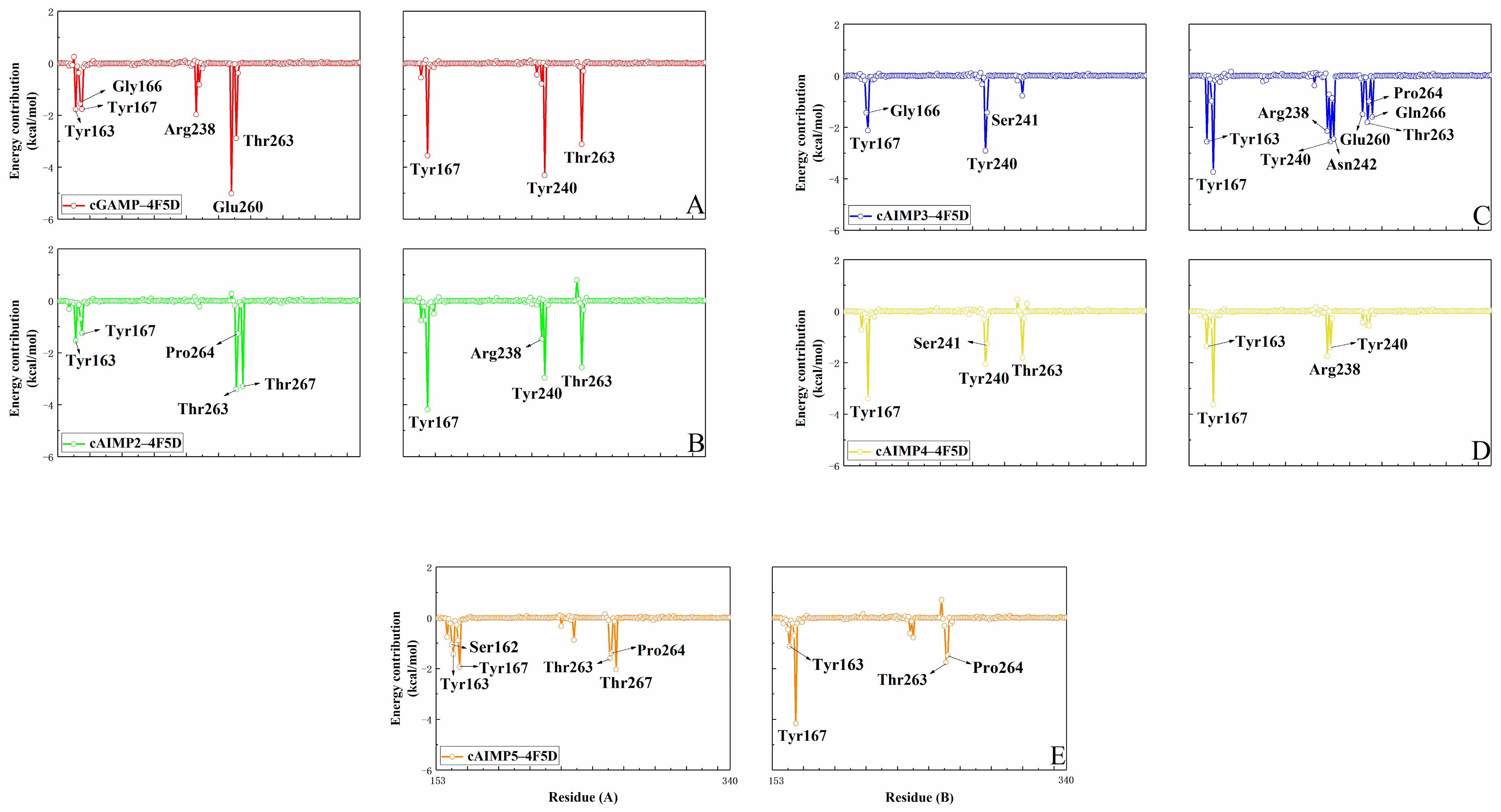
2.2. Binding Free Energy
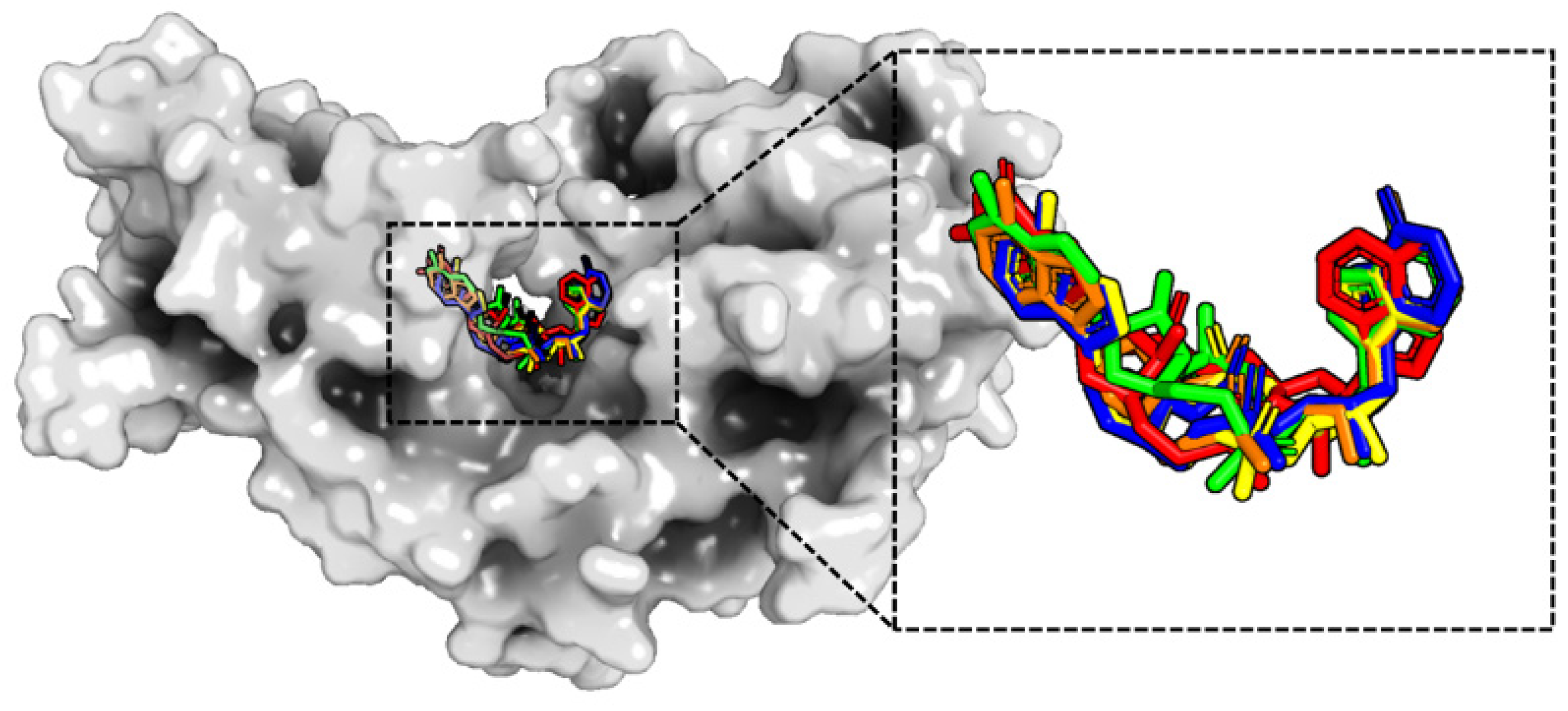
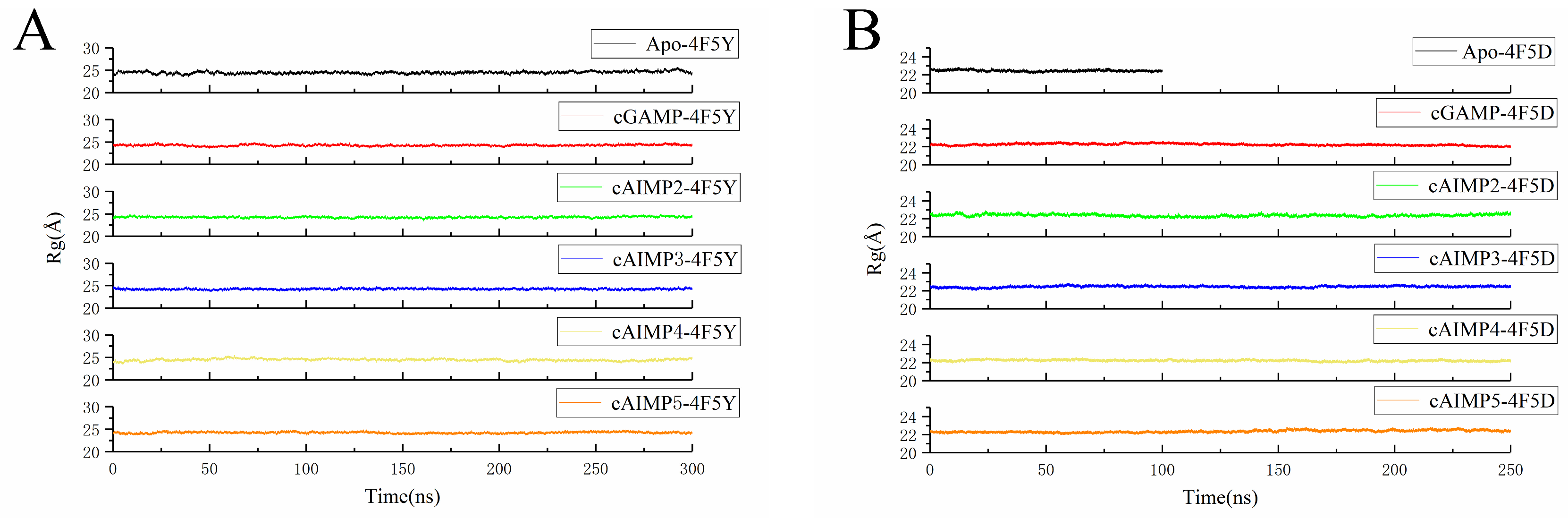
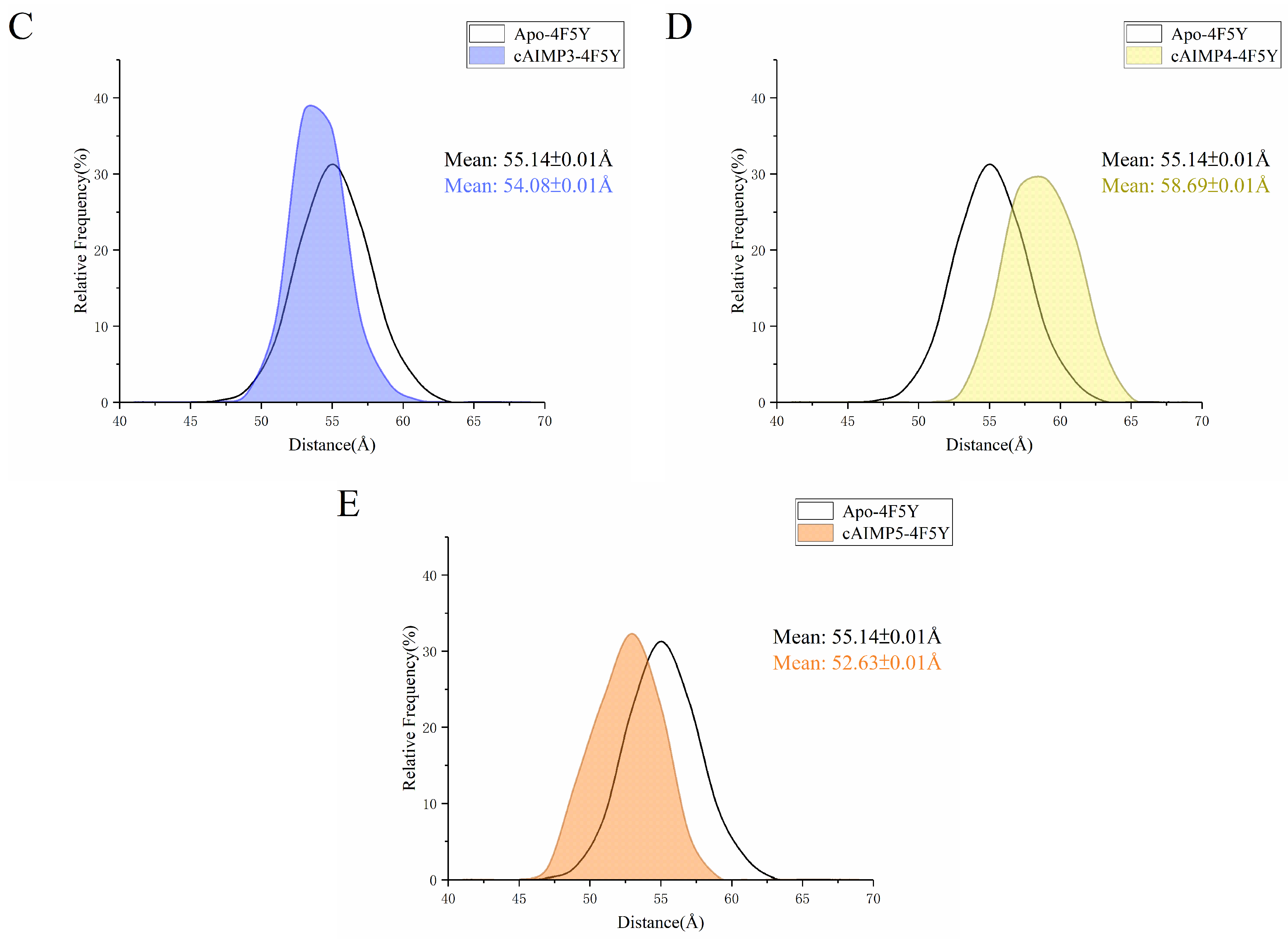
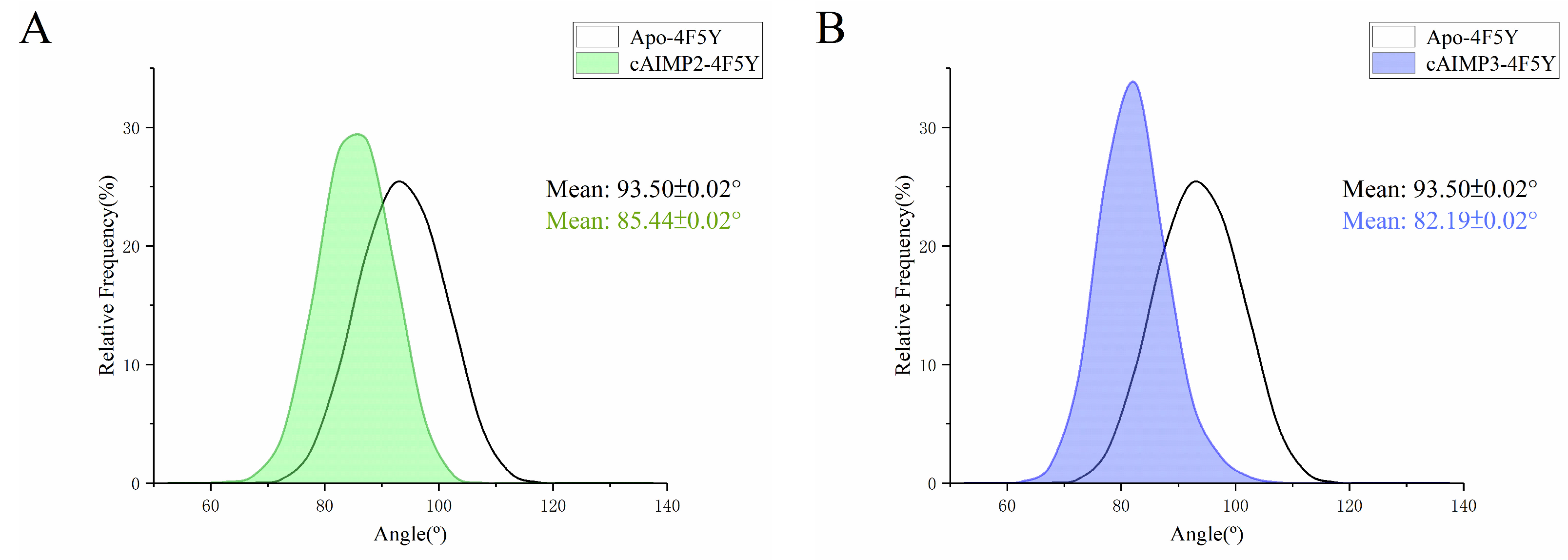
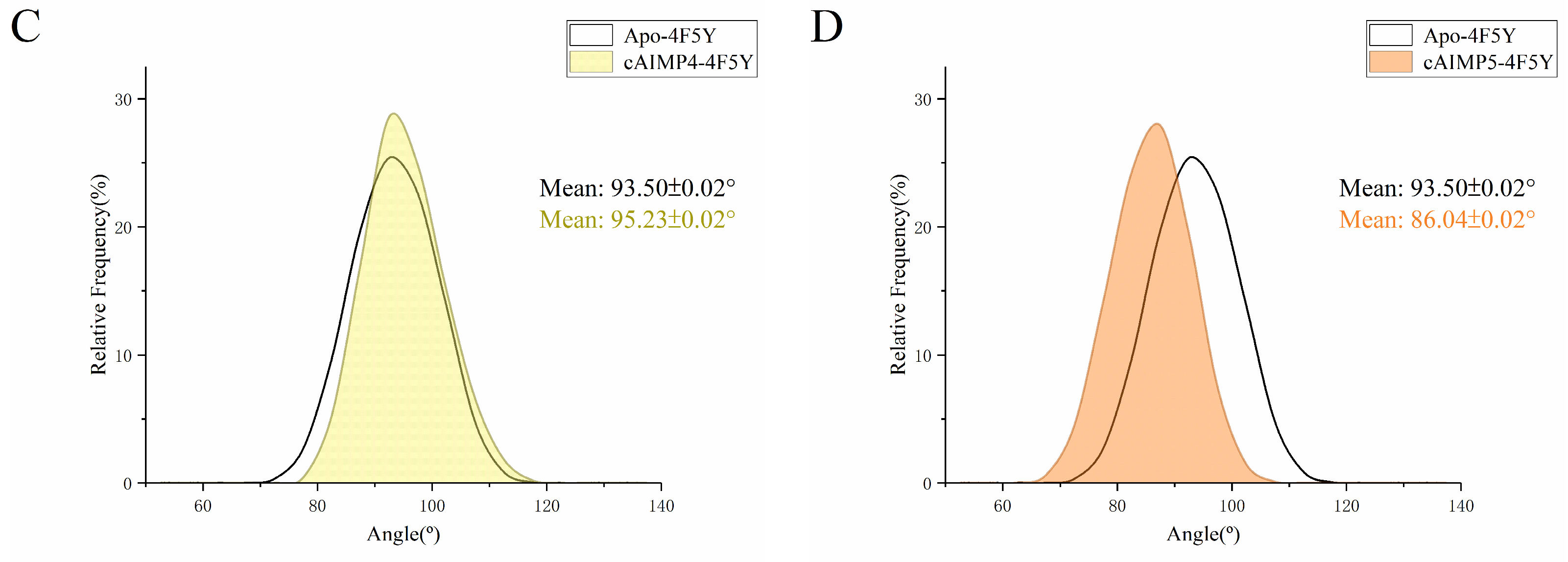
2.3. Flexibility and Conformational Change
3. Theory and Methods
3.1. System Preparation and Simulation Details
3.1.1. Molecular Docking
3.1.2. Molecular Dynamics Simulation
3.2. Methods
3.2.1. Trajectory Analysis
3.2.2. Binding Free Energy Calculation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ishikawa, H.; Barber, G.N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Cai, X.; Wu, J.; Cong, Q.; Chen, X.; Li, T.; Du, F.; Ren, J.; Wu, Y.-T.; Grishin, N.V.; et al. Phosphorylation of innate immune adaptor proteins MAVS, STING, and TRIF induces IRF3 activation. Science 2015, 347, aaa2630. [Google Scholar] [PubMed]
- Amouzegar, A.; Chelvanambi, M.; Filderman, J.N.; Storkus, W.J.; Luke, J.J. STING agonists as cancer therapeutics. Cancers 2021, 13, 2695. [Google Scholar] [CrossRef] [PubMed]
- Shang, G.; Zhang, C.; Chen, Z.J.; Bai, X.; Zhang, X. Cryo-EM structures of STING reveal its mechanism of activation by cyclic GMP–AMP. Nature 2019, 567, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.; Yi, G.; Watts, T.; Kao, C.C.; Li, P. Structure of STING bound to cyclic di-GMP reveals the mechanism of cyclic dinucleotide recognition by the immune system. Nat. Struct. Mol. Biol. 2012, 19, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, X.; Du, X.; Jiang, Z.; Su, X. The structural basis for the sensing and binding of cyclic di-GMP by STING. Nat. Struct. Mol. Biol. 2012, 19, 728–730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bai, X.; Chen, Z.J. Structures and mechanisms in the cGAS-STING innate immunity pathway. Immunity 2020, 53, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shang, G.; Gui, X.; Zhang, X.; Bai, X.-C.; Chen, Z.J. Structural basis of STING binding with and phosphorylation by TBK1. Nature 2019, 567, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Ergun, S.L.; Fernandez, D.; Weiss, T.M.; Li, L. STING polymer structure reveals mechanisms for activation, hyperactivation, and inhibition. Cell 2019, 178, 290–301.e10. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, S.; Song, X.; Wang, Y.; Ru, H.; Shaw, N.; Jiang, Y.; Niu, F.; Zhu, Y.; Qiu, W.; Parvatiyar, K.; et al. Structural analysis of the STING adaptor protein reveals a hydrophobic dimer interface and mode of cyclic di-GMP binding. Immunity 2012, 36, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, H.; Wu, J.; Zhang, X.; Sun, L.; Chen, C.; Chen, Z.J. Cyclic GMP-AMP containing mixed phosphodiester linkages is an endogenous high-affinity ligand for STING. Mol. Cell 2013, 51, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Aval, L.M.; Pease, J.E.; Sharma, R.; Pinato, D.J. Challenges and opportunities in the clinical development of STING agonists for cancer immunotherapy. J. Clin. Med. 2020, 9, 3323. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.K.; Kulkarni, M.M.; Chamberlain, B.; Dwight, T.; Koh, C.; Samant, R.; Jernigan, F.; Rice, J.; Tan, D.; Li, S.; et al. Design of a systemic small molecule clinical STING agonist using physics-based simulations and artificial intelligence. Sci. Rep. 2022, 12, 12179. [Google Scholar]
- Kim, D.S.; Endo, A.; Fang, F.G.; Huang, K.C.; Bao, X.; Choi, H.W.; Majumder, U.; Shen, Y.Y.; Mathieu, S.; Zhu, X.; et al. E7766, a macrocycle-bridged stimulator of interferon genes (STING) agonist with potent pan-genotypic activity. ChemMedChem 2021, 16, 1741–1744. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yang, Z.; Cheng, K.; Bi, H.; Chen, J. Small molecule-based immunomodulators for cancer therapy. Acta Pharm. Sin. B 2022, 12, 4287–4308. [Google Scholar] [CrossRef]
- Gao, P.; Ascano, M.; Zillinger, T.; Wang, W.; Dai, P.; Serganov, A.A.; Gaffney, B.L.; Shuman, S.; Jones, R.A.; Deng, L.; et al. Structure-function analysis of STING activation by c [G (2′, 5′) pA (3′, 5′) p] and targeting by antiviral DMXAA. Cell 2013, 154, 748–762. [Google Scholar] [CrossRef] [PubMed]
- Vavřina, Z.; Gutten, O.; Smola, M.; Zavřel, M.; Aliakbar Tehrani, Z.; Charvat, V.; Kožíšek, M.; Boura, E.; Birkuš, G.; Rulíšek, L. Protein–Ligand Interactions in the STING Binding Site Probed by Rationally Designed Single-Point Mutations: Experiment and Theory. Biochemistry 2021, 60, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; You, Q.; Xu, X. Targeting stimulator of interferon genes (STING): A medicinal chemistry perspective. J. Med. Chem. 2019, 63, 3785–3816. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhao, L.; Hu, H.; Li, W.; Li, Y. Agonists and inhibitors of the STING pathway: Potential agents for immunotherapy. Med. Res. Rev. 2020, 40, 1117–1141. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Zhang, H.; You, Q.; Xu, X. Agonist of stimulator of interferon genes as antitumor agents: A patent review (2008–2020). Expert Opin. Ther. Pat. 2021, 31, 563–584. [Google Scholar] [CrossRef] [PubMed]
- Pimková Polidarová, M.; Brehova, P.; Kaiser, M.M.; Smola, M.; Dracinsky, M.; Smith, J.; Marek, A.; Dejmek, M.; Šála, M.; Gutten, O.; et al. Synthesis and biological evaluation of phosphoester and phosphorothioate prodrugs of STING agonist 3′, 3′-c-Di (2′ F, 2′ dAMP). J. Med. Chem. 2021, 64, 7596–7616. [Google Scholar] [CrossRef] [PubMed]
- Meric-Bernstam, F.; Sweis, R.F.; Kasper, S.; Hamid, O.; Bhatia, S.; Dummer, R.; Stradella, A.; Long, G.V.; Spreafico, A.; Shimizu, T.; et al. Combination of the STING agonist MIW815 (ADU-S100) and PD-1 inhibitor spartalizumab in advanced/metastatic solid tumors or lymphomas: An open-label, multicenter, phase Ib study. Clin. Cancer Res. 2023, 29, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Ager, C.R.; Zhang, H.; Wei, Z.; Jones, P.; Curran, M.A.; Di Francesco, M.E. Discovery of IACS-8803 and IACS-8779, potent agonists of stimulator of interferon genes (STING) with robust systemic antitumor efficacy. Bioorg. Med. Chem. Lett. 2019, 29, 126640. [Google Scholar] [CrossRef] [PubMed]
- Maddess, M.; McIntosh, J.; Chang, W. Discovery and chemical development of uvelostinag (MK-1454): A therapeutic cyclic dinucleotide agonist of the stimulator of interferon gene. In Complete Accounts of Integrated Drug Discovery and Development: Recent Examples from the Pharmaceutical Industry; ACS Publications: Washington, DC, USA, 2022; Volume 4, pp. 1–94. [Google Scholar]
- Chang, W.; Altman, M.D.; Lesburg, C.A.; Perera, S.A.; Piesvaux, J.A.; Schroeder, G.K.; Wyss, D.F.; Cemerski, S.; Chen, Y.; DiNunzio, E.; et al. Discovery of MK-1454: A potent cyclic dinucleotide stimulator of interferon genes agonist for the treatment of cancer. J. Med. Chem. 2022, 65, 5675–5689. [Google Scholar] [CrossRef] [PubMed]
- Lioux, T.; Mauny, M.A.; Lamoureux, A.; Bascoul, N.; Hays, M.; Vernejoul, F.; Baudru, A.-S.; Boularan, C.; Lopes-Vicente, J.; Qushair, G.; et al. Design, synthesis, and biological evaluation of novel cyclic adenosine–inosine monophosphate (cAIMP) analogs that activate stimulator of interferon genes (STING). J. Med. Chem. 2016, 59, 10253–10267. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.H.F.; Olesen, R.; Jønsson, K.L.; Højen, J.F.; Krapp, C.; Mack, K.; Thomsen, M.K.; Østergaard, L.; Tolstrup, M.; Dagnæs-Hansen, F.; et al. cAIMP administration in humanized mice induces a chimerization-level-dependent STING response. Immunology 2019, 157, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.K.; Skouboe, M.K.; Boularan, C.; Vernejoul, F.; Lioux, T.; Leknes, S.L.; Berthelsen, M.F.; Riedel, M.; Cai, H.; Joseph, J.V.; et al. The cGAS-STING pathway is a therapeutic target in a preclinical model of hepatocellular carcinoma. Oncogene 2020, 39, 1652–1664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhai, Q.; Feng, X.; Chen, D.; Lu, Y.; Hu, J.; Xie, H.; Zhou, L.; Wu, J.; Zheng, S. Cancer cell-intrinsic STING is associated with CD8+ T-cell infiltration and might serve as a potential immunotherapeutic target in hepatocellular carcinoma. Clin. Transl. Oncol. 2021, 23, 1314–1324. [Google Scholar] [CrossRef] [PubMed]
- Skouboe, M.K.; Knudsen, A.; Reinert, L.S.; Boularan, C.; Lioux, T.; Perouzel, E.; Thomsen, M.K.; Paludan, S.R. STING agonists enable antiviral cross-talk between human cells and confer protection against genital herpes in mice. PLoS Pathog. 2018, 14, e1006976. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhao, S.; Zhu, Y.; Liu, Y.; Li, H.; Zhao, Q. Molecular dynamics simulations reveal the modulated mechanism of STING conformation. Interdiscip. Sci. Comput. Life Sci. 2021, 13, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Ramanjulu, J.M.; Pesiridis, G.S.; Yang, J.; Concha, N.; Singhaus, R.; Zhang, S.-Y.; Tran, J.-L.; Moore, P.; Lehmann, S.; Eberl, H.C.; et al. Design of amidobenzimidazole STING receptor agonists with systemic activity. Nature 2018, 564, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.T.; Crivelli, S.; Watanabe, M. All-Atom simulations uncover structural and dynamical properties of STING proteins in the membrane system. J. Chem. Inf. Model. 2022, 62, 4486–4499. [Google Scholar] [CrossRef] [PubMed]
- Smola, M.; Gutten, O.; Dejmek, M.; Kozisek, M.; Evangelidis, T.; Tehrani, Z.A.; Novotna, B.; Nencka, R.; Birkus, G.; Rulisek, L.; et al. Ligand strain and its conformational complexity is a major factor in the binding of cyclic dinucleotides to STING protein. Angew. Chem. Int. Ed. 2021, 60, 10172–10178. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, C.; Wang, C.; Zhang, H.; Ma, D.; Zhang, Q.; Wen, X.; Li, L.; Xi, Z. Synthesis and biological evaluation of all possible inosine-mixed cyclic dinucleotides that activate different hSTING variants. Biorg. Med. Chem. 2021, 29, 115899. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; He, Z.; Liu, H.; Chen, Y.; Gao, Y.; Zhang, S.; Wang, M.; Lu, X.; Wang, C.; Zhao, Z.; et al. 3D-QSAR, molecular docking, and molecular dynamics simulation study of thieno [3, 2-b] pyrrole-5-carboxamide derivatives as LSD1 inhibitors. RSC Adv. 2020, 10, 6927–6943. [Google Scholar] [CrossRef] [PubMed]
- Yanase, Y.; Tsuji, G.; Nakamura, M.; Shibata, N.; Demizu, Y. Control of STING agonistic/antagonistic activity using amine-skeleton-based c-di-GMP analogues. Int. J. Mol. Sci. 2022, 23, 6847. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yue, C.; Xie, S.; Shi, S.; Ye, S. Computational insights into the conformational transition of STING: Mechanistic, energetic considerations, and the influence of crucial mutations. J. Mol. Graphics Modell. 2024, 129, 108764. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Chen, L.; He, X.; Cao, D.; Zhang, Z.; Jiang, H.; Chen, K.; Cheng, X. Loops mediate agonist-induced activation of the stimulator of interferon genes protein. J. Chem. Inf. Model. 2023, 63, 7373–7381. [Google Scholar] [CrossRef] [PubMed]
- Tehrani, Z.A.; Rulíšek, L.; Černý, J. Molecular dynamics simulations provide structural insight into binding of cyclic dinucleotides to human STING protein. J. Biomol. Struct. Dyn. 2022, 40, 10250–10264. [Google Scholar] [CrossRef] [PubMed]
- Shang, G.; Zhu, D.; Li, N.; Zhang, J.; Zhu, C.; Lu, D.; Liu, C.; Yu, Q.; Zhao, Y.; Xu, S.; et al. Crystal structures of STING protein reveal basis for recognition of cyclic di-GMP. Nat. Struct. Mol. Biol. 2012, 19, 725–727. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Gao, Y.; Yang, M.; Wang, M.; Lu, J.; Wu, Z.; Zhao, J.; Yu, Y.; Wang, C.; Zhao, Z.; et al. Design and identification of two novel resveratrol derivatives as potential LSD1 inhibitors. Future Med. Chem. 2021, 13, 1415–1433. [Google Scholar] [CrossRef]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Giambasu, G.; et al. AMBER2020, University of California, San Fransisco. J. Amer. Chem. Soc 2020, 142, 3823–3835. [Google Scholar]
- Case, D.A.; Cheatham, T.E., III; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [PubMed]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Hub, J.S.; De Groot, B.L.; Grubmu, H.; Groenhof, G. Quantifying artifacts in Ewald simulations of inhomogeneous systems with a net charge. J. Chem. Theory Comput. 2014, 10, 381–390. [Google Scholar] [CrossRef] [PubMed]
- van Gunsteren, W.F.; Berendsen, H.J. Algorithms for macromolecular dynamics and constraint dynamics. Mol. Phys. 1977, 34, 1311–1327. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅ log (N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Sun, H.; Wang, J.; Wang, Z.; Liu, H.; Zhang, J.Z.H.; Hou, T. End-point binding free energy calculation with MM/PBSA and MM/GBSA: Strategies and applications in drug design. Chem. Rev. 2019, 119, 9478–9508. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fan, B.; Gao, Y.; Chen, Y.; Han, D.; Lu, J.; Liu, T.; Gao, Q.; Zhang, J.Z.; Wang, M. Design two novel tetrahydroquinoline derivatives against anticancer target LSD1 with 3D-QSAR model and molecular simulation. Molecules 2022, 27, 8358. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.R.; McGee, T.D.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Complex RMSD (Å) | Ligand RMSD (Å) | ||
|---|---|---|---|---|
| Mean | Max | Mean | Max | |
| Apo-4F5Y | 3.60 ± 0.00 | 5.05 | - | - |
| cGAMP-4F5Y | 3.25 ± 0.00 | 3.89 | 1.70 ± 0.00 | 2.03 |
| cAIMP2-4F5Y | 3.02 ± 0.00 | 3.67 | 0.51 ± 0.00 | 1.22 |
| cAIMP3-4F5Y | 3.06 ± 0.00 | 3.63 | 0.86 ± 0.00 | 1.64 |
| cAIMP4-4F5Y | 3.66 ± 0.00 | 4.53 | 1.46 ± 0.00 | 2.27 |
| cAIMP5-4F5Y | 3.27 ± 0.00 | 3.61 | 1.73 ± 0.00 | 2.39 |
| System | H-Bond | Docking | MD | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Acceptor | Donor | Length (Å) | Angle (°) | Energy (kcal/mol) | Length (Å) | Angle (°) | Energy (kcal/mol) | Hydrogen Bond Occupancy | |
| cGAMP-4F5Y | Lig-O11 | Arg238A-NH2 | 2.1 | 136.4 | −6.7 | 1.9 | 166.5 | −8.5 | 90% |
| Lig-O9 | Arg238A-NH1 | - | - | - | 1.9 | 165.6 | −8.2 | 83% | |
| cAIMP2-4F5Y | Lig-O4 | Ser162B-OG | - | - | - | 1.7 | 161.2 | −2.9 | 96% |
| Lig-N3 | Thr263B-OG1 | 2.6 | 159.9 | −1.7 | 2.1 | 147.4 | −1.7 | 29% | |
| cAIMP3-4F5Y | Lig-O11 | Thr263B-OG1 | - | - | - | 1.8 | 172.6 | −1.6 | 51% |
| Lig-N8 | Thr263A-OG1 | 2.2 | 160.1 | −2.0 | 2.1 | 165.4 | −2.3 | 47% | |
| Lig-O9 | Val239A-N | 2.4 | 144.4 | −1.3 | 2.0 | 155.5 | −3.8 | 34% | |
| Lig-N3 | Thr263B-OG1 | 2.3 | 141.4 | −1.2 | 2.5 | 149.5 | −2.4 | 32% | |
| cAIMP4-4F5Y | Lig-O4 | Ser162B-OG | - | - | - | 1.6 | 159.0 | −3.1 | 98% |
| Lig-N3 | Thr263B-OG1 | 2.3 | 144.8 | −1.4 | 1.8 | 162.7 | −1.8 | 36% | |
| cAIMP5-4F5Y | Lig-N3 | Thr263B-OG1 | 2.5 | 150.7 | −1.4 | 2.0 | 169.8 | −1.8 | 80% |
| Lig-S1 | Ser162B-OG | 3.7 | 127.6 | −0.6 | 2.2 | 157.5 | −3.7 | 41% | |
| Lig-N8 | Thr263A-OG1 | 2.2 | 159.9 | −2.1 | 2.3 | 148.0 | −1.3 | 34% | |
| Lig-O9 | Val239A-N | 2.2 | 150.2 | −2.6 | 1.9 | 161.0 | −2.6 | 32% | |
| Lig-S1 | Thr267A-OG1 | - | - | - | 2.0 | 168.4 | −3.7 | 30% | |
| Lig-O5 | Arg238A-NH1 | - | - | - | 1.9 | 138.4 | −6.4 | 25% | |
| System | H-Bond | Docking | MD | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Acceptor | Donor | Length (Å) | Angle (°) | Energy (kcal/mol) | Length (Å) | Angle (°) | Energy (kcal/mol) | Hydrogen Bond Occupancy | |
| cGAMP-4F5D | Lig-O9 | Tyr240B-OH | - | - | - | 1.7 | 161.5 | −4.4 | 97% |
| Lig-O8 | Thr263B-OG1 | - | - | - | 1.8 | 156.7 | −1.2 | 38% | |
| Lig-N6 | Thr263B-OG1 | - | - | - | 1.8 | 170.0 | −1.9 | 27% | |
| cAIMP2-4F5D | Lig-O7 | Thr267A-OG1 | - | - | - | 1.8 | 162.9 | −3.2 | 92% |
| Lig-N3 | Thr263A-OG1 | - | - | - | 1.9 | 156.7 | −1.1 | 82% | |
| Lig-N4 | Tyr240B-OH | - | - | - | 1.8 | 160.5 | −2.5 | 78% | |
| cAIMP3-4F5D | Lig-N3 | Tyr163B-OH | - | - | - | 2.4 | 160.5 | −3.2 | 37% |
| Lig-O9 | Ser241A-N | 2.1 | 148.1 | −2.2 | 1.9 | 164.1 | −5.9 | 35% | |
| cAIMP4-4F5D | Lig-O2 | Thr263A-OG1 | - | - | - | 2.0 | 175.7 | −1.1 | 44% |
| cAIMP5-4F5D | Lig-S1 | Thr267A-OG1 | - | - | - | 2.1 | 148.1 | −3.4 | 45% |
| Compound | THP1-Dual | Blood | Docking Score (kcal/mol) | Binding Free Energy (kcal/mol) | |||
|---|---|---|---|---|---|---|---|
| IRF EC50 (μM) | NF-κB EC50 (μM) | Type I IFN EC50 (μM) | Open | Closed | Open | Closed | |
| cGAMP | 7.2 ± 3.2 | 39.1 ± 22.6 | 19.6 ± 6.7 | −8.85 | −17.7 | −33.7 ± 0.2 | −37.6 ± 1.9 |
| cAIMP2 | 5.1 ± 1.4 | 15.9 ± 4.1 | 6.4 ± 2.3 | −9.18 | −19.6 | −34.5 ± 1.5 | −49.6 ± 2.0 |
| cAIMP3 | 1.6 ± 1.3 | 7.8 ± 1.3 | 10.6 ± 3.9 | −9.69 | −21.0 | −48.1 ± 1.8 | −34.0 ± 0.2 |
| cAIMP4 | 1.1 ± 0.4 | 15.4 ± 3.2 | 0.7 ± 0.1 | −9.92 | −19.6 | −33.9 ± 0.2 | −40.2 ± 2.4 |
| cAIMP5 | 0.3 ± 0.2 | 1.6 ± 0.3 | 0.4 ± 0.1 | −9.72 | −21.3 | −45.1 ± 0.8 | −35.9 ± 2.2 |
| System | Rg (Å) | System | Rg (Å) | ||
|---|---|---|---|---|---|
| Mean | Max | Mean | Max | ||
| Apo-4F5Y | 24.81 ± 0.00 | 25.66 | Apo-4F5D | 22.42 ± 0.00 | 22.64 |
| cGAMP-4F5Y | 24.41 ± 0.00 | 24.92 | cGAMP-4F5D | 22.06 ± 0.00 | 22.27 |
| cAIMP2-4F5Y | 24.40 ± 0.00 | 24.97 | cAIMP2-4F5D | 22.50 ± 0.00 | 22.86 |
| cAIMP3-4F5Y | 24.29 ± 0.00 | 24.82 | cAIMP3-4F5D | 22.47 ± 0.00 | 22.70 |
| cAIMP4-4F5Y | 24.54 ± 0.00 | 25.10 | cAIMP4-4F5D | 22.16 ± 0.00 | 22.42 |
| cAIMP5-4F5Y | 24.24 ± 0.00 | 24.71 | cAIMP5-4F5D | 22.42 ± 0.00 | 22.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Fan, B.; Lu, W.; Ryde, U.; Chang, Y.; Han, D.; Lu, J.; Liu, T.; Gao, Q.; Chen, C.; et al. Unraveling the Binding Mode of Cyclic Adenosine–Inosine Monophosphate (cAIMP) to STING through Molecular Dynamics Simulations. Molecules 2024, 29, 2650. https://doi.org/10.3390/molecules29112650
Wang M, Fan B, Lu W, Ryde U, Chang Y, Han D, Lu J, Liu T, Gao Q, Chen C, et al. Unraveling the Binding Mode of Cyclic Adenosine–Inosine Monophosphate (cAIMP) to STING through Molecular Dynamics Simulations. Molecules. 2024; 29(11):2650. https://doi.org/10.3390/molecules29112650
Chicago/Turabian StyleWang, Meiting, Baoyi Fan, Wenfeng Lu, Ulf Ryde, Yuxiao Chang, Di Han, Jiarui Lu, Taigang Liu, Qinghe Gao, Changpo Chen, and et al. 2024. "Unraveling the Binding Mode of Cyclic Adenosine–Inosine Monophosphate (cAIMP) to STING through Molecular Dynamics Simulations" Molecules 29, no. 11: 2650. https://doi.org/10.3390/molecules29112650
APA StyleWang, M., Fan, B., Lu, W., Ryde, U., Chang, Y., Han, D., Lu, J., Liu, T., Gao, Q., Chen, C., & Xu, Y. (2024). Unraveling the Binding Mode of Cyclic Adenosine–Inosine Monophosphate (cAIMP) to STING through Molecular Dynamics Simulations. Molecules, 29(11), 2650. https://doi.org/10.3390/molecules29112650