

State of the Art in the Development of Human Serum Carnosinase Inhibitors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
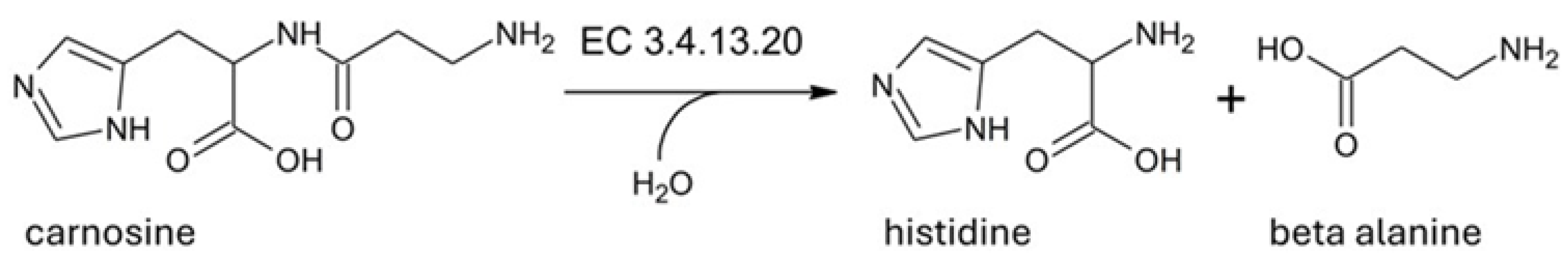
1. Enzymatic Activity of Human Serum Carnosinase
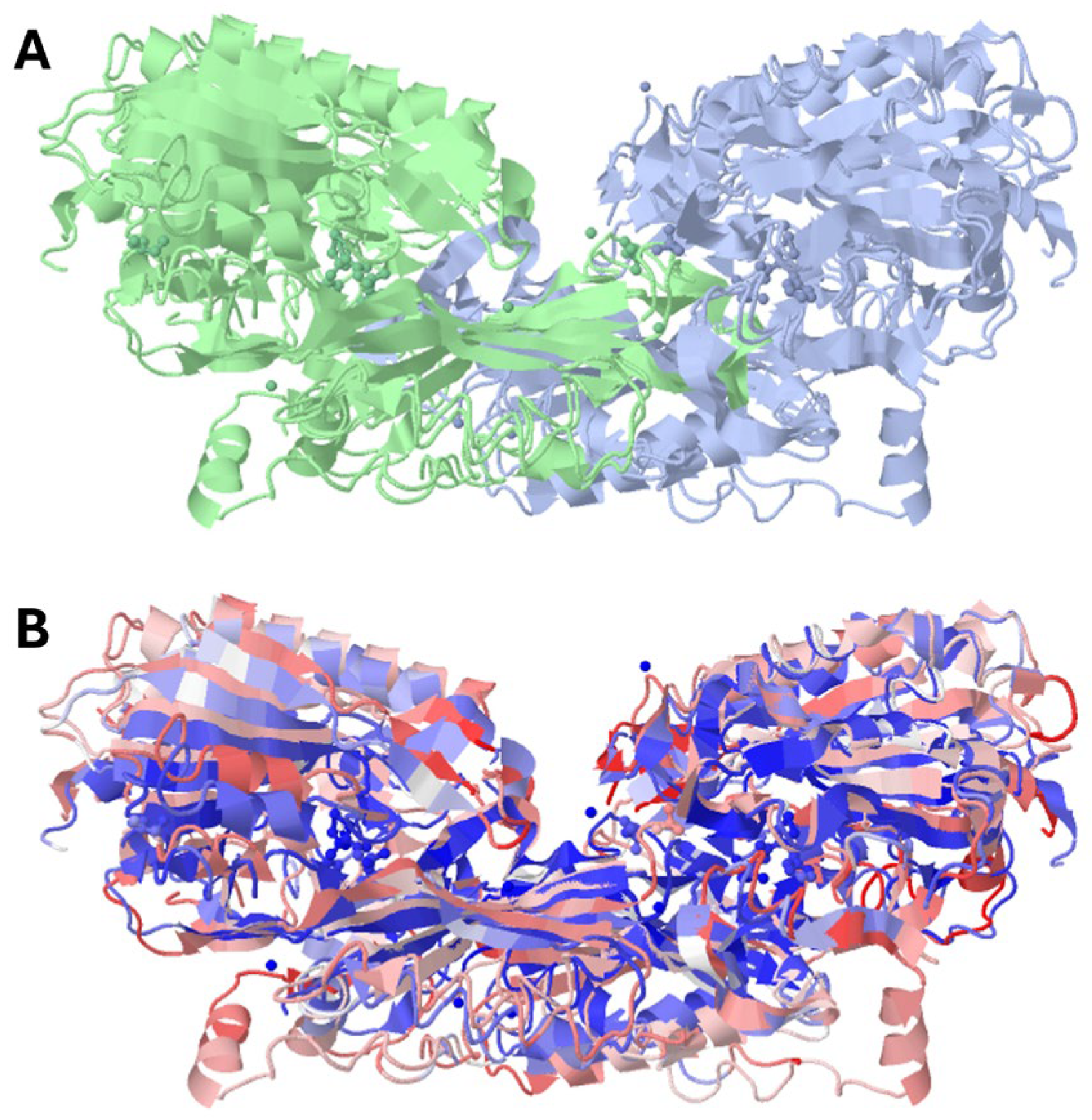
1.1. Enzyme Classification
1.2. Closely Related Enzymes
1.3. Expression and Localization
1.4. Substrate Selectivity
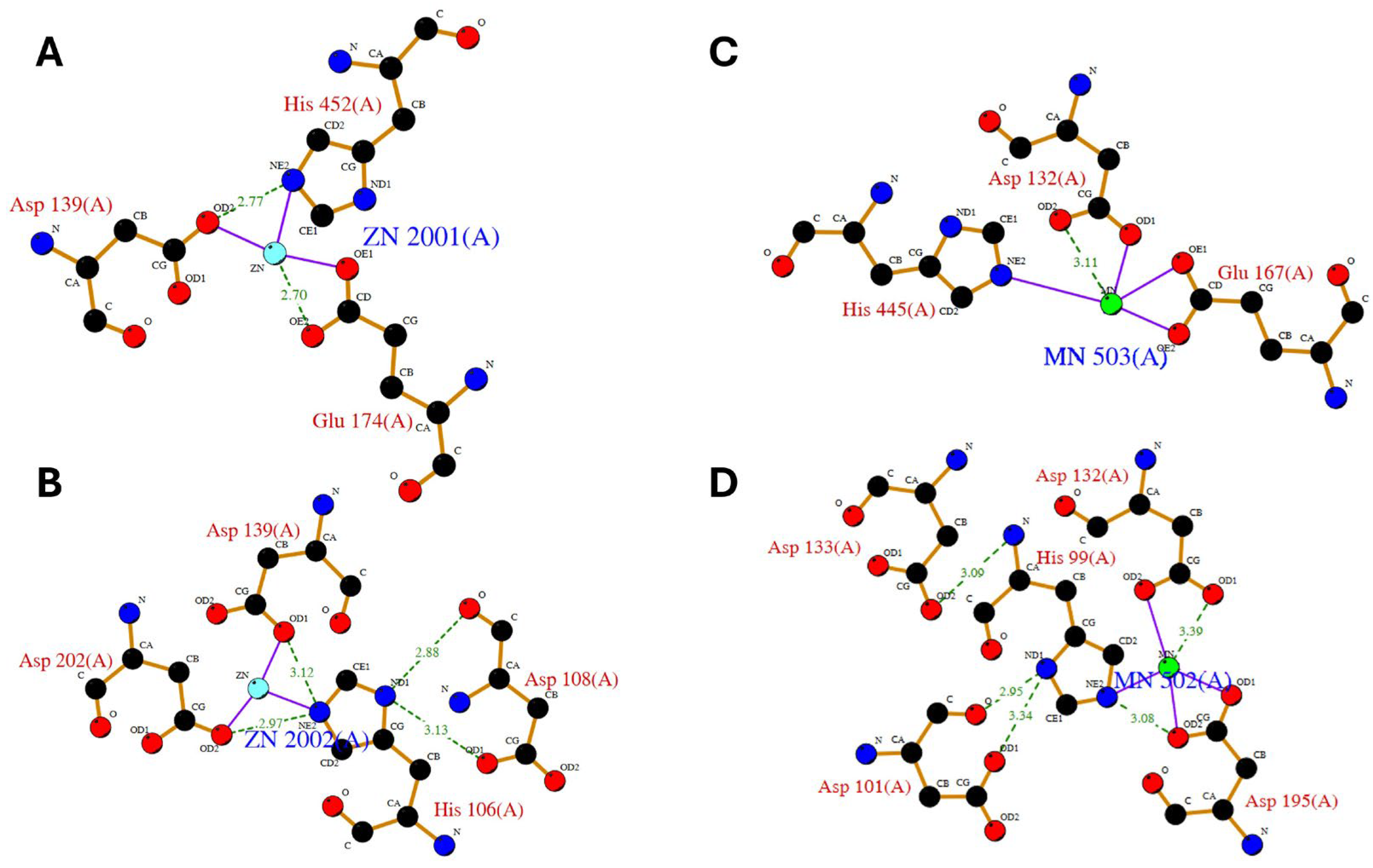
1.5. Substrate Recognition and Structure Activity Relationships
2. Compounds Able to Alter the Activity of Human Serum Carnosinase
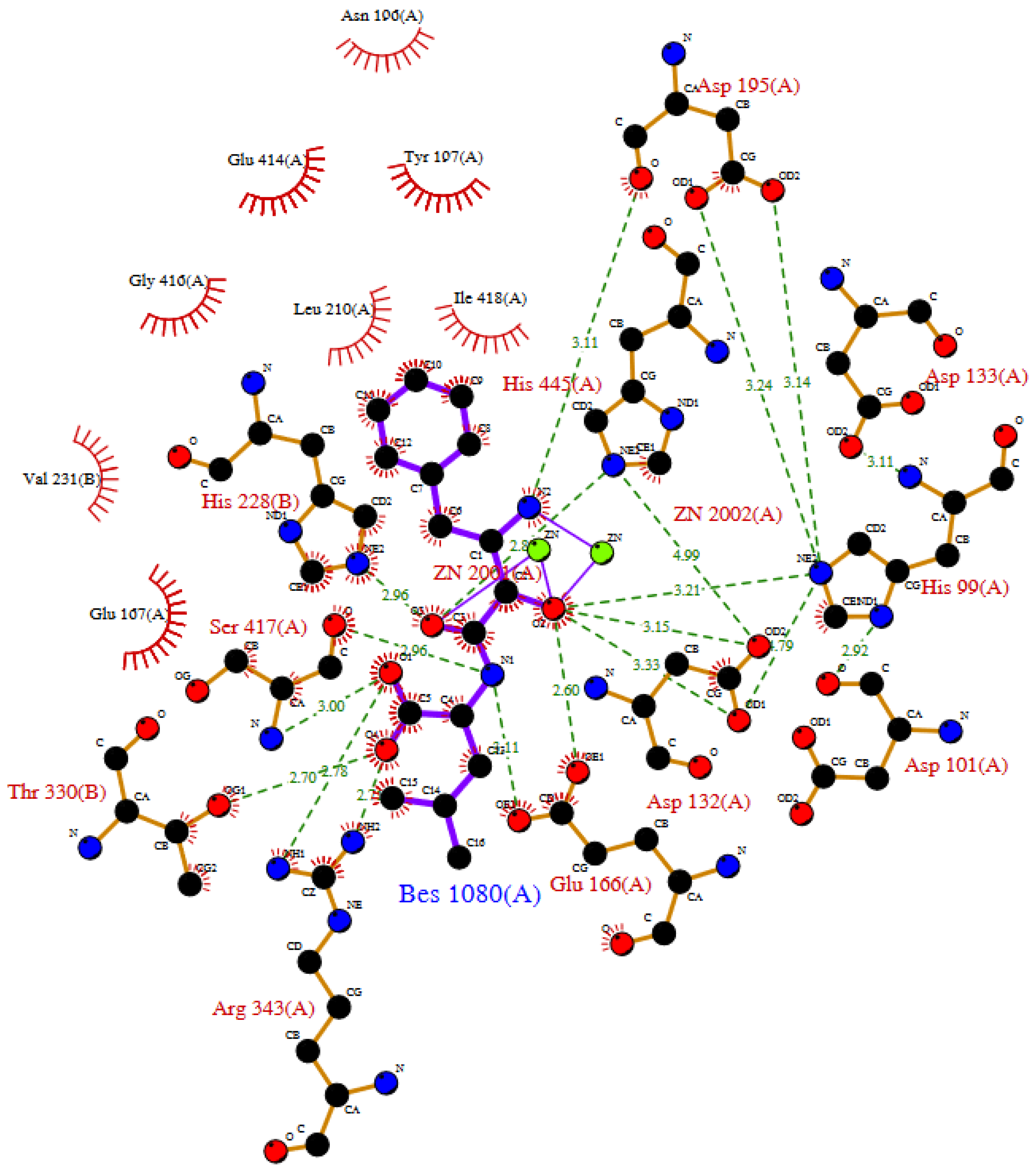
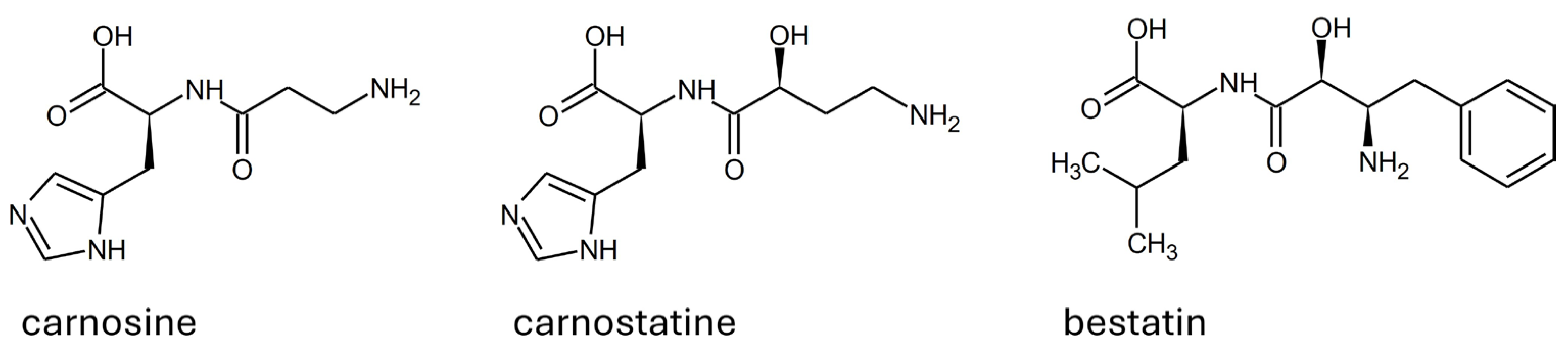
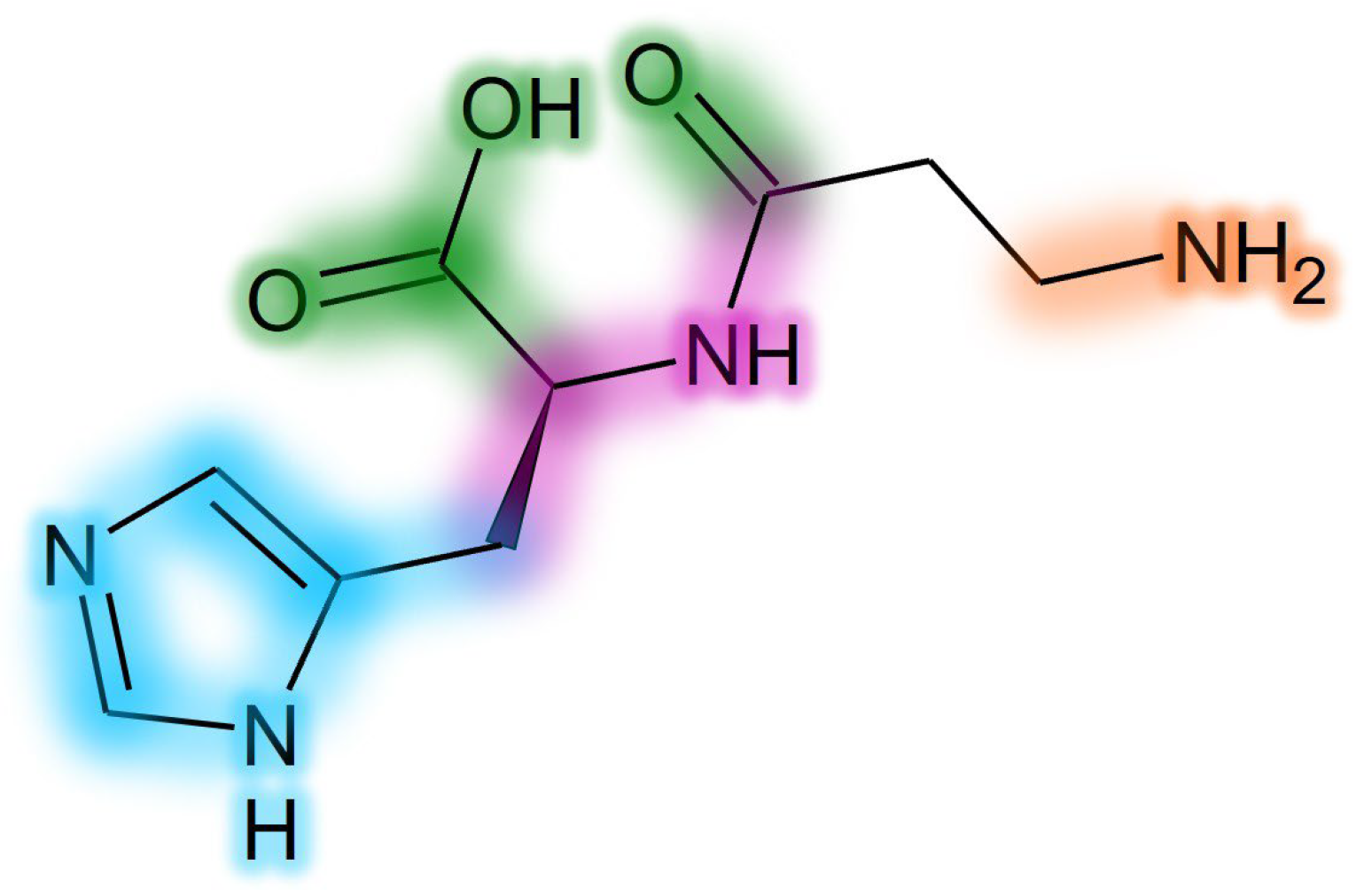
2.1. Competitive Inhibitors
2.2. Allosteric Modulators
2.3. Compounds Interfering with Zinc
3. Human Serum Carnosinase in Human Disease
3.1. Human Serum Carnosinase and Central Nervous System Diseases
3.2. Human Serum Carnosinase in Other Diseases
3.3. The Role of Carnosine and Homocarnosine in Diseases with Altered Carnosinase Activity
4. Human Serum Carnosinase as a Druggable Target
Funding
Conflicts of Interest
References
- McDonald, A.G.; Tipton, K.F. Enzyme nomenclature and classification: The state of the art. FEBS J. 2023, 290, 2214–2231. [Google Scholar] [CrossRef]
- Murphey, W.H.; Patchen, L.; Lindmark, D.G. Carnosinase: A fluorometric assay and demonstration of two electrophoretic forms in human tissue extracts. Clin. Chim. Acta 1972, 42, 309–314. [Google Scholar] [CrossRef]
- Teufel, M.; Saudek, V.; Ledig, J.P.; Bernhardt, A.; Boularand, S.; Carreau, A.; Cairns, N.J.; Carter, C.; Cowley, D.J.; Duverger, D.; et al. Sequence identification and characterization of human carnosinase and a closely related non-specific dipeptidase. J. Biol. Chem. 2003, 278, 6521–6531. [Google Scholar] [CrossRef]
- Lenney, J.F.; George, R.P.; Weiss, A.M.; Kucera, C.M.; Chan, P.W.; Rinzler, G.S. Human serum carnosinase: Characterization, distinction from cellular carnosinase, and activation by cadmium. Clin. Chim. Acta 1982, 123, 221–231. [Google Scholar] [CrossRef]
- Rawlings, N.D. Twenty-five years of nomenclature and Classif. of proteolytic enzymes. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2020, 1868, 140345. [Google Scholar] [CrossRef]
- Pavlin, M.; Rossetti, G.; De Vivo, M.; Carloni, P. Carnosine and Homocarnosine Degradation Mechanisms by the Human Carnosinase Enzyme CN1: Insights from Multiscale Simulations. Biochemistry 2016, 55, 2772–2784. [Google Scholar] [CrossRef]
- Toviwek, B.; Koonawootrittriron, S.; Suwanasopee, T.; Pongprayoon, P. Molecular insights into the binding of carnosine and anserine to human serum carnosinase 1 (CN1). PeerJ Phys. Chem. 2022, 4, e25. [Google Scholar] [CrossRef]
- Pandya, V.K.; Shankar, S.S.; Sonwane, B.P.; Rajesh, S.; Rathore, R.; Kumaran, S.; Kulkarni, M.J. Mechanistic insights on anserine hydrolyzing activities of human carnosinases. Biochim. Biophys. Acta Gen. Subj. 2023, 1867, 130290. [Google Scholar] [CrossRef]
- Toviwek, B.; Suwanasopee, T.; Koonawootrittriron, S.; Jattawa, D.; Pongprayoon, P. Binding Modes of Carnostatine, Homocarnosine, and Ophidine to Human Carnosinase 1. ACS Omega 2023, 8, 42966–42975. [Google Scholar] [CrossRef]
- Tancharoen, C.; Tovivek, B.; Niramitranon, J.; Kityakarn, S.; Luksirikul, P.; Gorinstein, S.; Pongprayoon, P. Exploring the structural and dynamic differences between human carnosinase I (CN1) and II (CN2). Proteins 2023, 91, 822–830. [Google Scholar] [CrossRef]
- Vistoli, G.; Pedretti, A.; Cattaneo, M.; Aldini, G.; Testa, B. Homology modeling of human serum carnosinase, a potential medicinal target, and MD simulations of its allosteric activation by citrate. J. Med. Chem. 2006, 49, 3269–3277. [Google Scholar] [CrossRef]
- Chmielewska, K.; Vittorio, S.; Gervasoni, S.; Dzierzbicka, K.; Inkielewicz-Stepniak, I.; Vistoli, G. Human carnosinases: A brief history, medicinal relevance, and in silico analyses. Drug Discov. Today 2024, 29, 103860. [Google Scholar] [CrossRef]
- Peters, V.; Kebbewar, M.; Jansen, E.W.; Jakobs, C.; Riedl, E.; Koeppel, H.; Frey, D.; Adelmann, K.; Klingbeil, K.; Mack, M.; et al. Relevance of allosteric conformations and homocarnosine concentration on carnosinase activity. Amino Acids 2010, 38, 1607–1615. [Google Scholar] [CrossRef]
- Jackson, M.C.; Kucera, C.M.; Lenney, J.F. Purification and properties of human serum carnosinase. Clin. Chim. Acta 1991, 196, 193–205. [Google Scholar] [CrossRef]
- Priestman, D.A.; Butterworth, J. Prolinase and non-specific dipeptidase of human kidney. Biochem. J. 1985, 231, 689–694. [Google Scholar] [CrossRef]
- Lenney, J.F. Human cytosolic carnosinase: Evidence of identity with prolinase, a non-specific dipeptidase. Biol. Chem. Hoppe-Seyler 1990, 371, 167–171. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Lenney, J.F.; Peppers, S.C.; Kucera-Orallo, C.M.; George, R.P. Characterization of human tissue carnosinase. Biochem. J. 1985, 228, 653–660. [Google Scholar] [CrossRef]
- Pandya, V.K.; Sonwane, B.; Rathore, R.; Unnikrishnan, A.G.; Kumaran, S.; Kulkarni, M.J. Development of multiple reaction monitoring assay for quantification of carnosine in human plasma. RSC Adv. 2020, 10, 763–769. [Google Scholar] [CrossRef]
- Peters, V.; Klessens, C.Q.; Baelde, H.J.; Singler, B.; Veraar, K.A.; Zutinic, A.; Drozak, J.; Zschocke, J.; Schmitt, C.P.; de Heer, E. Intrinsic carnosine metabolism in the human kidney. Amino Acids 2015, 47, 2541–2550. [Google Scholar] [CrossRef]
- Lenney, J.F. Separation and characterization of two carnosine-splitting cytosolic dipeptidases from hog kidney (carnosinase and non-specific dipeptidase). Biol. Chem. Hoppe-Seyler 1990, 371, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Margolis, F.L.; Grillo, M.; Brown, C.E.; Williams, T.H.; Pitcher, R.G.; Elgar, G.J. Enzymatic and immunological evidence for two forms of carnosinase in the mouse. Biochim. Biophys. Acta 1979, 570, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Margolis, F.L.; Grillo, M.; Grannot-Reisfeld, N.; Farbman, A.I. Purification, characterization and immunocytochemical localization of mouse kidney carnosinase. Biochim. Biophys. Acta 1983, 744, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Janssen, B.; Hohenadel, D.; Brinkkoetter, P.; Peters, V.; Rind, N.; Fischer, C.; Rychlik, I.; Cerna, M.; Romzova, M.; de Heer, E.; et al. Carnosine as a protective factor in diabetic nephropathy: Association with a leucine repeat of the carnosinase gene CNDP1. Diabetes 2005, 54, 2320–2327. [Google Scholar] [CrossRef] [PubMed]
- Riedl, E.; Koeppel, H.; Brinkkoetter, P.; Sternik, P.; Steinbeisser, H.; Sauerhoefer, S.; Janssen, B.; van der Woude, F.J.; Yard, B.A. A CTG polymorphism in the CNDP1 gene determines the secretion of serum carnosinase in Cos-7 transfected cells. Diabetes 2007, 56, 2410–2413. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Kebbewar, M.; Janssen, B.; Hoffmann, G.F.; Moller, K.; Wygoda, S.; Charbit, M.; Fernandes-Teixeira, A.; Jeck, N.; Zschocke, J.; et al. CNDP1 genotype and renal survival in pediatric nephropathies. J. Pediatr. Endocrinol. Metab. 2016, 29, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Albrecht, T.; Rodriguez-Nino, A.; Qiu, J.; Schnuelle, P.; Peters, V.; Schmitt, C.P.; van den Born, J.; Bakker, S.J.L.; Lammert, A.; et al. Carnosinase concentration, activity, and CNDP1 genotype in patients with type 2 diabetes with and without nephropathy. Amino Acids 2019, 51, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Freedman, B.I.; Hicks, P.J.; Sale, M.M.; Pierson, E.D.; Langefeld, C.D.; Rich, S.S.; Xu, J.; McDonough, C.; Janssen, B.; Yard, B.A.; et al. A leucine repeat in the carnosinase gene CNDP1 is associated with diabetic end-stage renal disease in European Americans. Nephrol. Dial. Transpl. 2007, 22, 1131–1135. [Google Scholar] [CrossRef] [PubMed]
- Mooyaart, A.L.; van Valkengoed, I.G.; Shaw, P.K.; Peters, V.; Baelde, H.J.; Rabelink, T.J.; Bruijn, J.A.; Stronks, K.; de Heer, E. Lower frequency of the 5/5 homozygous CNDP1 genotype in South Asian Surinamese. Diabetes Res. Clin. Pract. 2009, 85, 272–278. [Google Scholar] [CrossRef]
- Albrecht, T.; Zhang, S.; Braun, J.D.; Xia, L.; Rodriquez, A.; Qiu, J.; Peters, V.; Schmitt, C.P.; van den Born, J.; Bakker, S.J.L.; et al. The CNDP1 (CTG)(5) Polymorphism Is Associated with Biopsy-Proven Diabetic Nephropathy, Time on Hemodialysis, and Diabetes Duration. J. Diabetes Res. 2017, 2017, 9506730. [Google Scholar] [CrossRef]
- Riedl, E.; Koeppel, H.; Pfister, F.; Peters, V.; Sauerhoefer, S.; Sternik, P.; Brinkkoetter, P.; Zentgraf, H.; Navis, G.; Henning, R.H.; et al. N-glycosylation of carnosinase influences protein secretion and enzyme activity: Implications for hyperglycemia. Diabetes 2010, 59, 1984–1990. [Google Scholar] [CrossRef]
- Yadav, A.K.; Sinha, N.; Kumar, V.; Bhansali, A.; Dutta, P.; Jha, V. Association of CTG repeat polymorphism in carnosine dipeptidase 1 (CNDP1) gene with diabetic nephropathy in north Indians. Indian. J. Med. Res. 2016, 144, 32–37. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, D.; Tang, M.; Yang, G.; Yard, B.; Hu, H.; Wu, Y.; Zhang, Q. Serum and urinary carnosinase-1 correlate with kidney function and inflammation. Amino Acids 2023, 55, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Mooyaart, A.L.; Zutinic, A.; Bakker, S.J.; Grootendorst, D.C.; Kleefstra, N.; van Valkengoed, I.G.; Bohringer, S.; Bilo, H.J.; Dekker, F.W.; Bruijn, J.A.; et al. Association between CNDP1 genotype and diabetic nephropathy is sex specific. Diabetes 2010, 59, 1555–1559. [Google Scholar] [CrossRef] [PubMed]
- Alkhalaf, A.; Landman, G.W.; van Hateren, K.J.; Groenier, K.H.; Mooyaart, A.L.; De Heer, E.; Gans, R.O.; Navis, G.J.; Bakker, S.J.; Kleefstra, N.; et al. Sex specific association between carnosinase gene CNDP1 and cardiovascular mortality in patients with type 2 diabetes (ZODIAC-22). J. Nephrol. 2015, 28, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Artioli, G.G.; Sale, C.; Jones, R.L. Carnosine in health and disease. Eur. J. Sport. Sci. 2019, 19, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, J.; Lancha, A.H., Jr. Single Nucleotide Polymorphisms in Carnosinase Genes (CNDP1 and CNDP2) are Associated with Power Athletic Status. Int. J. Sport. Nutr. Exerc. Metab. 2017, 27, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Baguet, A.; Everaert, I.; Yard, B.; Peters, V.; Zschocke, J.; Zutinic, A.; De Heer, E.; Podgorski, T.; Domaszewska, K.; Derave, W. Does low serum carnosinase activity favor high-intensity exercise capacity? J. Appl. Physiol. 2014, 116, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Gilardoni, E.; Gervasoni, S.; Maspero, M.; Dallanoce, C.; Vistoli, G.; Carini, M.; Aldini, G.; Regazzoni, L. Development of a direct LC-ESI-MS method for the measurement of human serum carnosinase activity. J. Pharm. Biomed. Anal. 2020, 189, 113440. [Google Scholar] [CrossRef]
- Pegova, A.; Abe, H.; Boldyrev, A. Hydrolysis of carnosine and related compounds by mammalian carnosinases. Comp. Biochem. Physiol. Part. B Biochem. Mol. Biol. 2000, 127, 443–446. [Google Scholar] [CrossRef]
- Everaert, I.; Baron, G.; Barbaresi, S.; Gilardoni, E.; Coppa, C.; Carini, M.; Vistoli, G.; Bex, T.; Stautemas, J.; Blancquaert, L.; et al. Development and validation of a sensitive LC-MS/MS assay for the quantification of anserine in human plasma and urine and its application to pharmacokinetic study. Amino Acids 2019, 51, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Cacciatore, I.; Cocco, A.; Costa, M.; Fontana, M.; Lucente, G.; Pecci, L.; Pinnen, F. Biochemical properties of new synthetic carnosine analogues containing the residue of 2,3-diaminopropionic acid: The effect of N-acetylation. Amino Acids 2005, 28, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Orioli, M.; Vistoli, G.; Regazzoni, L.; Pedretti, A.; Lapolla, A.; Rossoni, G.; Canevotti, R.; Gamberoni, L.; Previtali, M.; Carini, M.; et al. Design, synthesis, ADME properties, and pharmacological activities of beta-alanyl-D-histidine (D-carnosine) prodrugs with improved bioavailability. ChemMedChem 2011, 6, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Iacobini, C.; Menini, S.; Blasetti Fantauzzi, C.; Pesce, C.M.; Giaccari, A.; Salomone, E.; Lapolla, A.; Orioli, M.; Aldini, G.; Pugliese, G. FL-926-16, a novel bioavailable carnosinase-resistant carnosine derivative, prevents onset and stops progression of diabetic nephropathy in db/db mice. Br. J. Pharmacol. 2018, 175, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Vistoli, G.; Katunga, L.A.; Funai, K.; Regazzoni, L.; Monroe, T.B.; Gilardoni, E.; Cannizzaro, L.; Colzani, M.; De Maddis, D.; et al. A carnosine analog mitigates metabolic disorders of obesity by reducing carbonyl stress. J. Clin. Investig. 2018, 128, 5280–5293. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Jansen, E.E.; Jakobs, C.; Riedl, E.; Janssen, B.; Yard, B.A.; Wedel, J.; Hoffmann, G.F.; Zschocke, J.; Gotthardt, D.; et al. Anserine inhibits carnosine degradation but in human serum carnosinase (CN1) is not correlated with histidine dipeptide concentration. Clin. Chim. Acta 2011, 412, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; Orioli, M.; Rossoni, G.; Savi, F.; Braidotti, P.; Vistoli, G.; Yeum, K.J.; Negrisoli, G.; Carini, M. The carbonyl scavenger carnosine ameliorates dyslipidaemia and renal function in Zucker obese rats. J. Cell. Mol. Med. 2011, 15, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Jozic, D.; Bourenkow, G.; Bartunik, H.; Scholze, H.; Dive, V.; Henrich, B.; Huber, R.; Bode, W.; Maskos, K. Crystal structure of the dinuclear zinc aminopeptidase PepV from Lactobacillus delbrueckii unravels its preference for dipeptides. Structure 2002, 10, 1097–1106. [Google Scholar] [CrossRef]
- Bando, K.; Shimotsuji, T.; Toyoshima, H.; Hayashi, C.; Miyai, K. Fluorometric assay of human serum carnosinase activity in normal children, adults and patients with myopathy. Ann. Clin. Biochem. 1984, 21 Pt 6, 510–514. [Google Scholar] [CrossRef]
- Bellia, F.; Calabrese, V.; Guarino, F.; Cavallaro, M.; Cornelius, C.; De Pinto, V.; Rizzarelli, E. Carnosinase levels in aging brain: Redox state induction and cellular stress response. Antioxid. Redox Signal. 2009, 11, 2759–2775. [Google Scholar] [CrossRef]
- Lindner, H.A.; Lunin, V.V.; Alary, A.; Hecker, R.; Cygler, M.; Menard, R. Essential roles of zinc ligation and enzyme dimerization for catalysis in the aminoacylase-1/M20 family. J. Biol. Chem. 2003, 278, 44496–44504. [Google Scholar] [CrossRef] [PubMed]
- de Giacomi, C.; Regazzoni, L. A Green and Cost-Effective Chromatographic Method for the Measurement of the Activity of Human Serum Carnosinase. Separations 2023, 10, 460. [Google Scholar] [CrossRef]
- Peters, V.; Schmitt, C.P.; Weigand, T.; Klingbeil, K.; Thiel, C.; van den Berg, A.; Calabrese, V.; Nawroth, P.; Fleming, T.; Forsberg, E.; et al. Allosteric inhibition of carnosinase (CN1) by inducing a conformational shift. J. Enzym. Inhib. Med. Chem. 2017, 32, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Hauske, S.J.; Zhang, S.; Rodriguez-Nino, A.; Albrecht, T.; Pastene, D.O.; van den Born, J.; van Goor, H.; Ruf, S.; Kohlmann, M.; et al. Identification and characterisation of carnostatine (SAN9812), a potent and selective carnosinase (CN1) inhibitor with in vivo activity. Amino Acids 2019, 51, 7–16. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, X.Q.; Zhang, S.Q.; Qi, X.M.; Zhang, Q.; Yard, B.; Wu, Y.G. Correlation between serum carnosinase concentration and renal damage in diabetic nephropathy patients. Amino Acids 2021, 53, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Schoen, P.; Everts, H.; de Boer, T.; van Oeveren, W. Serum carnosinase activity in plasma and serum: Validation of a method and values in cardiopulmonary bypass surgery. Clin. Chem. 2003, 49, 1930–1932. [Google Scholar] [CrossRef] [PubMed]
- Everaert, I.; Mooyaart, A.; Baguet, A.; Zutinic, A.; Baelde, H.; Achten, E.; Taes, Y.; De Heer, E.; Derave, W. Vegetarianism, female gender and increasing age, but not CNDP1 genotype, are associated with reduced muscle carnosine levels in humans. Amino Acids 2011, 40, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- de Jager, S.; Blancquaert, L.; Van der Stede, T.; Lievens, E.; De Baere, S.; Croubels, S.; Gilardoni, E.; Regazzoni, L.G.; Aldini, G.; Bourgois, J.G.; et al. The ergogenic effect of acute carnosine and anserine supplementation: Dosing, timing, and underlying mechanism. J. Int. Soc. Sports Nutr. 2022, 19, 70–91. [Google Scholar] [CrossRef] [PubMed]
- Blancquaert, L.; Everaert, I.; Baguet, A.; Bex, T.; Barbaresi, S.; de Jager, S.; Lievens, E.; Stautemas, J.; De Smet, S.; Baron, G.; et al. Acute preexercise supplementation of combined carnosine and anserine enhances initial maximal power of Wingate tests in humans. J. Appl. Physiol. 2021, 130, 1868–1878. [Google Scholar] [CrossRef]
- Albrecht, T.; Schilperoort, M.; Zhang, S.; Braun, J.D.; Qiu, J.; Rodriguez, A.; Pastene, D.O.; Kramer, B.K.; Koppel, H.; Baelde, H.; et al. Carnosine Attenuates the Development of both Type 2 Diabetes and Diabetic Nephropathy in BTBR ob/ob Mice. Sci. Rep. 2017, 7, 44492. [Google Scholar] [CrossRef]
- Brown, B.E.; Kim, C.H.; Torpy, F.R.; Bursill, C.A.; McRobb, L.S.; Heather, A.K.; Davies, M.J.; van Reyk, D.M. Supplementation with carnosine decreases plasma triglycerides and modulates atherosclerotic plaque composition in diabetic apo E(-/-) mice. Atherosclerosis 2014, 232, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Grasso, G.I.; Arena, G.; Bellia, F.; Rizzarelli, E.; Vecchio, G. Copper(II)-chelating homocarnosine glycoconjugate as a new multifunctional compound. J. Inorg. Biochem. 2014, 131, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Scornik, O.A.; Botbol, V. Bestatin as an experimental tool in mammals. Curr. Drug Metab. 2001, 2, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Peppers, S.C.; Lenney, J.F. Bestatin inhibition of human tissue carnosinase, a non-specific cytosolic dipeptidase. Biol. Chem. Hoppe-Seyler 1988, 369, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Lanthaler, B.; Amberger, A.; Fleming, T.; Forsberg, E.; Hecker, M.; Wagner, A.H.; Yue, W.W.; Hoffmann, G.F.; Nawroth, P.; et al. Carnosine metabolism in diabetes is altered by reactive metabolites. Amino Acids 2015, 47, 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.; Meyerstein, D.; Czapski, G. The Fenton reagents. Free Radic. Biol. Med. 1993, 15, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Abordo, E.A.; Minhas, H.S.; Thornalley, P.J. Accumulation of alpha-oxoaldehydes during oxidative stress: A role in cytotoxicity. Biochem. Pharmacol. 1999, 58, 641–648. [Google Scholar] [CrossRef]
- Lange, J.N.; Wood, K.D.; Knight, J.; Assimos, D.G.; Holmes, R.P. Glyoxal formation and its role in endogenous oxalate synthesis. Adv. Urol. 2012, 2012, 819202. [Google Scholar] [CrossRef]
- Thornalley, P.J.; Langborg, A.; Minhas, H.S. Formation of glyoxal, methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose. Biochem. J. 1999, 344 Pt 1, 109–116. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Giustarini, D.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation in human diseases. Trends Mol. Med. 2003, 9, 169–176. [Google Scholar] [CrossRef]
- Foster, M.W.; Hess, D.T.; Stamler, J.S. Protein S-nitrosylation in health and disease: A current perspective. Trends Mol. Med. 2009, 15, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Bellia, F.; Vecchio, G.; Rizzarelli, E. Carnosinases, their substrates and diseases. Molecules 2014, 19, 2299–2329. [Google Scholar] [CrossRef] [PubMed]
- Perry, T.L.; Hansen, S.; Tischler, B.; Bunting, R.; Berry, K. Carnosinemia. A new metabolic disorder associated with neurologic disease and mental defect. N. Engl. J. Med. 1967, 277, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Perry, T.L.; Hansen, S.; Love, D.L. Serum-carnosinase deficiency in carnosinaemia. Lancet 1968, 1, 1229–1230. [Google Scholar] [CrossRef] [PubMed]
- Fleisher, L.D.; Rassin, D.K.; Wisniewski, K.; Salwen, H.R. Carnosinase deficiency: A new variant with high residual activity. Pediatr. Res. 1980, 14, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Hartlage, P.L.; Krawiecki, N.; Roesel, R.A.; Carter, A.L.; Hommes, F.A. Serum carnosinase deficiency: A non-disabling phenotype? J. Ment. Defic. Res. 1985, 29 Pt 4, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Willi, S.M.; Zhang, Y.; Hill, J.B.; Phelan, M.C.; Michaelis, R.C.; Holden, K.R. A deletion in the long arm of chromosome 18 in a child with serum carnosinase deficiency. Pediatr. Res. 1997, 41, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Lunde, H.A.; Gjessing, L.R.; Sjaastad, O. Homocarnosinosis: Influence of dietary restriction of histidine. Neurochem. Res. 1986, 11, 825–838. [Google Scholar] [CrossRef]
- Balion, C.M.; Benson, C.; Raina, P.S.; Papaioannou, A.; Patterson, C.; Ismaila, A.S. Brain type carnosinase in dementia: A pilot study. BMC Neurol. 2007, 7, 38. [Google Scholar] [CrossRef]
- Wassif, W.S.; Sherwood, R.A.; Amir, A.; Idowu, B.; Summers, B.; Leigh, N.; Peters, T.J. Serum carnosinase activities in central nervous system disorders. Clin. Chim. Acta 1994, 225, 57–64. [Google Scholar] [CrossRef]
- Wisniewski, K.; Fleisher, L.; Rassin, D.; Lassmann, H. Neurological disease in a child with carnosinase deficiency. Neuropediatrics 1981, 12, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Godos, J.; Castellano, S.; Micek, A.; Murabito, P.; Galvano, F.; Ferri, R.; Grosso, G.; Caraci, F. The Therapeutic Potential of Carnosine/Anserine Supplementation against Cognitive Decline: A Systematic Review with Meta-Analysis. Biomedicines 2021, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, K.; Dzierzbicka, K.; Inkielewicz-Stepniak, I.; Przybylowska, M. Therapeutic Potential of Carnosine and Its Derivatives in the Treatment of Human Diseases. Chem. Res. Toxicol. 2020, 33, 1561–1578. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Albrecht, T.; Zhang, S.; Hauske, S.J.; Rodriguez-Nino, A.; Zhang, X.; Nosan, D.; Pastene, D.O.; Sticht, C.; Delatorre, C.; et al. Human carnosinase 1 overexpression aggravates diabetes and renal impairment in BTBR(Ob/Ob) mice. J. Mol. Med. 2020, 98, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Limonta, E.; Invernizzi, P.L.; Rampichini, S.; Bisconti, A.V.; Ce, E.; Longo, S.; Coratella, G.; Iaia, F.M.; Esposito, F. Acute carnosine and beta-alanine supplementation increase the compensated part of the ventilation versus work rate relationship during a ramp incremental cycle test in physically active men. J. Sports Med. Phys. Fit. 2021, 61, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Dolan, E.; Saunders, B.; Harris, R.C.; Bicudo, J.; Bishop, D.J.; Sale, C.; Gualano, B. Comparative physiology investigations support a role for histidine-containing dipeptides in intracellular acid-base regulation of skeletal muscle. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 234, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Stegen, S.; Sigal, R.J.; Kenny, G.P.; Khandwala, F.; Yard, B.; De Heer, E.; Baelde, H.; Peersman, W.; Derave, W. Aerobic and resistance training do not influence plasma carnosinase content or activity in type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E663–E669. [Google Scholar] [CrossRef] [PubMed]
- Schon, M.; Just, I.; Krumpolec, P.; Blazicek, P.; Valkovic, L.; Aldini, G.; Tsai, C.L.; De Courten, B.; Krssak, M.; Ukropcova, B.; et al. Supplementation-induced change in muscle carnosine is paralleled by changes in muscle metabolism, protein glycation and reactive carbonyl species sequestering. Physiol. Res. 2023, 72, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Freitas, P.A.C.; Ehlert, L.R.; Camargo, J.L. Glycated albumin: A potential biomarker in diabetes. Arch. Endocrinol. Metab. 2017, 61, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Baye, E.; Ukropcova, B.; Ukropec, J.; Hipkiss, A.; Aldini, G.; de Courten, B. Physiological and therapeutic effects of carnosine on cardiometabolic risk and disease. Amino Acids 2016, 48, 1131–1149. [Google Scholar] [CrossRef]
- Gualano, B.; Everaert, I.; Stegen, S.; Artioli, G.G.; Taes, Y.; Roschel, H.; Achten, E.; Otaduy, M.C.; Junior, A.H.; Harris, R.; et al. Reduced muscle carnosine content in type 2, but not in type 1 diabetic patients. Amino Acids 2012, 43, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Sauerhofer, S.; Yuan, G.; Braun, G.S.; Deinzer, M.; Neumaier, M.; Gretz, N.; Floege, J.; Kriz, W.; van der Woude, F.; Moeller, M.J. L-carnosine, a substrate of carnosinase-1, influences glucose metabolism. Diabetes 2007, 56, 2425–2432. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Zschocke, J.; Schmitt, C.P. Carnosinase, diabetes mellitus and the potential relevance of carnosinase deficiency. J. Inherit. Metab. Dis. 2018, 41, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, S.; Sheetal, M.M.; Gnanaprakash, V.; Chidambaram, M.; Liju, S.; Kanthi Mathi, S.; Shanthirani, C.S.; Mohan, V.; Radha, V.; Bodhini, D. IDF21-0406 Genetic variants in the Carnosine-Carnosinase system and their impact on diabetic nephropathy. Diabetes Res. Clin. Pract. 2022, 186, 109650. [Google Scholar] [CrossRef]
- Qiu, J.; Yard, B.A.; Kramer, B.K.; van Goor, H.; van Dijk, P.; Kannt, A. Association Between Serum Carnosinase Concentration and Activity and Renal Function Impairment in a Type-2 Diabetes Cohort. Front. Pharmacol. 2022, 13, 899057. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Nino, A.; Pastene, D.O.; Post, A.; Said, M.Y.; Gomes-Neto, A.W.; Kieneker, L.M.; Heiner-Fokkema, M.R.; Esatbeyoglu, T.; Rimbach, G.; Schnuelle, P.; et al. Urinary Carnosinase-1 Excretion is Associated with Urinary Carnosine Depletion and Risk of Graft Failure in Kidney Transplant Recipients: Results of the Transplant Lines Cohort Study. Antioxid 2021, 10, 1102. [Google Scholar] [CrossRef] [PubMed]
- Peters, V.; Yard, B.; Schmitt, C.P. Carnosine and Diabetic Nephropathy. Curr. Med. Chem. 2020, 27, 1801–1812. [Google Scholar] [CrossRef]
- Rodriguez-Nino, A.; Pastene, D.O.; Hettler, S.A.; Qiu, J.; Albrecht, T.; Vajpayee, S.; Perciaccante, R.; Gretz, N.; Bakker, S.J.L.; Kramer, B.K.; et al. Influence of carnosine and carnosinase-1 on diabetes-induced afferent arteriole vasodilation: Implications for glomerular hemodynamics. Am. J. Physiol. Ren. Physiol. 2022, 323, F69–F80. [Google Scholar] [CrossRef]
- Everaert, I.; He, J.; Hanssens, M.; Stautemas, J.; Bakker, K.; Albrecht, T.; Zhang, S.; Van der Stede, T.; Vanhove, K.; Hoetker, D.; et al. Carnosinase-1 overexpression, but not aerobic exercise training, affects the development of diabetic nephropathy in BTBR ob/ob mice. Am. J. Physiol. Ren. Physiol. 2020, 318, F1030–F1040. [Google Scholar] [CrossRef]
- Weigand, T.; Colbatzky, F.; Pfeffer, T.; Garbade, S.F.; Klingbeil, K.; Colbatzky, F.; Becker, M.; Zemva, J.; Bulkescher, R.; Schurfeld, R.; et al. A Global Cndp1-Knock-Out Selectively Increases Renal Carnosine and Anserine Concentrations in an Age- and Gender-Specific Manner in Mice. Int. J. Mol. Sci. 2020, 21, 4887. [Google Scholar] [CrossRef]
- Pfeffer, T.; Wetzel, C.; Kirschner, P.; Bartosova, M.; Poth, T.; Schwab, C.; Poschet, G.; Zemva, J.; Bulkescher, R.; Damgov, I.; et al. Carnosinase-1 Knock-Out Reduces Kidney Fibrosis in Type-1 Diabetic Mice on High Fat Diet. Antioxid 2023, 12, 1270. [Google Scholar] [CrossRef]
- Everaert, I.; Taes, Y.; De Heer, E.; Baelde, H.; Zutinic, A.; Yard, B.; Sauerhofer, S.; Vanhee, L.; Delanghe, J.; Aldini, G.; et al. Low plasma carnosinase activity promotes carnosinemia after carnosine ingestion in humans. Am. J. Physiol. Ren. Physiol. 2012, 302, F1537–F1544. [Google Scholar] [CrossRef]
- Cesak, O.; Vostalova, J.; Vidlar, A.; Bastlova, P.; Student, V., Jr. Carnosine and Beta-Alanine Supplementation in Human Medicine: Narrative Review and Critical Assessment. Nutrients 2023, 15, 1770. [Google Scholar] [CrossRef]
- Caruso, G. Unveiling the Hidden Therapeutic Potential of Carnosine, a Molecule with a Multimodal Mechanism of Action: A Position Paper. Molecules 2022, 27, 3303. [Google Scholar] [CrossRef]
- Caruso, G.; Di Pietro, L.; Cardaci, V.; Maugeri, S.; Caraci, F. The therapeutic potential of carnosine: Focus on cellular and molecular mechanisms. Curr. Res. Pharmacol. Drug Discov. 2023, 4, 100153. [Google Scholar] [CrossRef]
- Caruso, G.; Caraci, F.; Jolivet, R.B. Pivotal role of carnosine in the modulation of brain cells activity: Multimodal mechanism of action and therapeutic potential in neurodegenerative disorders. Prog. Neurobiol. 2019, 175, 35–53. [Google Scholar] [CrossRef]
- Chez, M.G.; Buchanan, C.P.; Aimonovitch, M.C.; Becker, M.; Schaefer, K.; Black, C.; Komen, J. Double-blind, placebo-controlled study of L-carnosine supplementation in children with autistic spectrum disorders. J. Child. Neurol. 2002, 17, 833–837. [Google Scholar] [CrossRef]
- Menini, S.; Iacobini, C.; Ricci, C.; Scipioni, A.; Blasetti Fantauzzi, C.; Giaccari, A.; Salomone, E.; Canevotti, R.; Lapolla, A.; Orioli, M.; et al. D-Carnosine octylester attenuates atherosclerosis and renal disease in ApoE null mice fed a Western diet through reduction of carbonyl stress and inflammation. Br. J. Pharmacol. 2012, 166, 1344–1356. [Google Scholar] [CrossRef]
- Al-Sawalha, N.A.; Alshogran, O.Y.; Awawdeh, M.S.; Almomani, B.A. The effects of l-Carnosine on development of metabolic syndrome in rats. Life Sci. 2019, 237, 116905. [Google Scholar] [CrossRef]
- Schon, M.; Mousa, A.; Berk, M.; Chia, W.L.; Ukropec, J.; Majid, A.; Ukropcova, B.; de Courten, B. The Potential of Carnosine in Brain-Related Disorders: A Comprehensive Review of Current Evidence. Nutrients 2019, 11, 1196. [Google Scholar] [CrossRef]
- Tardy, M.; Rolland, B.; Bardakdjian, J.; Gonnard, P. Action of homocarnosine, carnosine and anserine on uptake and metabolism of GABA in different subcellular fractions of rat brain. Experientia 1978, 34, 823–824. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H. Studies on homocarnosine in cerebrospinal fluid in infancy and childhood. Part II. Homocarnosine levels in cerebrospinal fluid from children with epilepsy, febrile convulsion or meningitis. Brain Dev. 1981, 3, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Grove, J.; Schechter, P.J.; Tell, G.; Rumbach, L.; Marescaux, C.; Warter, J.M.; Koch-Weser, J. Artifactual increases in the concentration of free GABA in samples of human cerebrospinal fluid are due to degradation of homocarnosine. J. Neurochem. 1982, 39, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, C.; Sato, Y.; Takahashi, H. Homocarnosine levels in cerebrospinal fluid of patients with infantile spasms under ACTH therapy. Brain Dev. 1983, 5, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, A.M.; Tell, G.; Schechter, P.J.; Grove, J.; Saint-Hilaire, M.H.; De Smet, Y.; Agid, Y. Cerebrospinal fluid GABA and homocarnosine concentrations in patients with Friedreich’s ataxia, Parkinson’s disease, and Huntington’s chorea. Mov. Disord. 1987, 2, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Perry, T.L.; Yong, V.W.; Bergeron, C.; Hansen, S.; Jones, K. Amino acids, glutathione, and glutathione transferase activity in the brains of patients with Alzheimer’s disease. Ann. Neurol. 1987, 21, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Jakobs, C.; Jaeken, J.; Gibson, K.M. Inherited disorders of GABA metabolism. J. Inherit. Metab. Dis. 1993, 16, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Lenney, J.F.; Peppers, S.C.; Kucera, C.M.; Sjaastad, O. Homocarnosinosis: Lack of serum carnosinase is the defect probably responsible for elevated brain and CSF homocarnosine. Clin. Chim. Acta 1983, 132, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Freund, M.A.; Chen, B.; Decker, E.A. The Inhibition of Advanced Glycation End Products by Carnosine and Other Natural Dipeptides to Reduce Diabetic and Age-Related Complications. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1367–1378. [Google Scholar] [CrossRef]
- Qi, Z.; Yu, X.; Xu, P.; Hao, Y.; Pan, X.; Zhang, C. l-Homocarnosine, l-carnosine, and anserine attenuate brain oxidative damage in a pentylenetetrazole-induced epilepsy model of ovariectomized rats. 3 Biotech 2018, 8, 363. [Google Scholar] [CrossRef]
- Huang, J.; Wang, T.; Yu, D.; Fang, X.; Fan, H.; Liu, Q.; Yi, G.; Yi, X.; Liu, Q. l-Homocarnosine attenuates inflammation in cerebral ischemia-reperfusion injury through inhibition of nod-like receptor protein 3 inflammasome. Int. J. Biol. Macromol. 2018, 118, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H. Protective effects of carnosine and homocarnosine on ferritin and hydrogen peroxide-mediated DNA damage. BMB Rep. 2010, 43, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kang, J.H. Protective effects of histidine dipeptides on the modification of neurofilament-L by the cytochrome c/hydrogen peroxide system. J. Biochem. Mol. Biol. 2007, 40, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Petroff, O.A.; Hyder, F.; Rothman, D.L.; Mattson, R.H. Brain homocarnosine and seizure control of patients taking gabapentin or topiramate. Epilepsia 2006, 47, 495–498. [Google Scholar] [CrossRef] [PubMed]
- de Jager, S.; Vermeulen, A.; De Baere, S.; Van der Stede, T.; Lievens, E.; Croubels, S.; Jager, R.; Purpura, M.; Bourgois, J.G.; Derave, W. Acute balenine supplementation in humans as a natural carnosinase-resistant alternative to carnosine. Sci. Rep. 2023, 13, 6484. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Senut, M.C.; Rajanikant, K.; Greenberg, E.; Bandagi, R.; Zemke, D.; Mousa, A.; Kassab, M.; Farooq, M.U.; Gupta, R.; et al. Differential neuroprotective effects of carnosine, anserine, and N-acetyl carnosine against permanent focal ischemia. J. Neurosci. Res. 2008, 86, 2984–2991. [Google Scholar] [CrossRef] [PubMed]
- de Jager, S.; Van Damme, S.; De Baere, S.; Croubels, S.; Jager, R.; Purpura, M.; Lievens, E.; Bourgois, J.G.; Derave, W. No Effect of Acute Balenine Supplementation on Maximal and Submaximal Exercise Performance in Recreational Cyclists. Int. J. Sport. Nutr. Exerc. Metab. 2023, 33, 84–92. [Google Scholar] [CrossRef]
- Everaert, I.; Van der Stede, T.; Stautemas, J.; Hanssens, M.; van Aanhold, C.; Baelde, H.; Vanhaecke, L.; Derave, W. Oral anserine supplementation does not attenuate type-2 diabetes or diabetic nephropathy in BTBR ob/ob mice. Amino Acids 2021, 53, 1269–1277. [Google Scholar] [CrossRef]
- Bonaccorso, A.; Privitera, A.; Grasso, M.; Salamone, S.; Carbone, C.; Pignatello, R.; Musumeci, T.; Caraci, F.; Caruso, G. The Therapeutic Potential of Novel Carnosine Formulations: Perspectives for Drug Development. Pharmaceuticals 2023, 16, 778. [Google Scholar] [CrossRef]
- Habra, K.; Morris, R.H.; McArdle, S.E.B.; Cave, G.W.V. Controlled release of carnosine from poly(lactic-co-glycolic acid) beads using nanomechanical magnetic trigger towards the treatment of glioblastoma. Nanoscale Adv. 2022, 4, 2242–2249. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, Y.; Wang, L.; Li, G.; Gao, J.; Wang, Y. Development of L-carnosine functionalized iron oxide nanoparticles loaded with dexamethasone for simultaneous therapeutic potential of blood brain barrier crossing and ischemic stroke treatment. Drug Deliv. 2021, 28, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Farid, R.M.; Gaafar, P.M.E.; Hazzah, H.A.; Helmy, M.W.; Abdallah, O.Y. Chemotherapeutic potential of L-carnosine from stimuli-responsive magnetic nanoparticles against breast cancer model. Nanomedicine 2020, 15, 891–911. [Google Scholar] [CrossRef] [PubMed]
- Moulahoum, H.; Sanli, S.; Timur, S.; Zihnioglu, F. Potential effect of carnosine encapsulated niosomes in bovine serum albumin modifications. Int. J. Biol. Macromol. 2019, 137, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Slovak, L.; Ponist, S.; Fedorova, T.; Logvinenko, A.; Levacheva, I.; Samsonova, O.; Bakowsky, U.; Paskova, L.; Cavojsky, T.; Tsiklauri, L.; et al. Evaluation of liposomal carnosine in adjuvant arthritis. Gen. Physiol. Biophys. 2017, 36, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Kim, D.; Nyberg, S.; Poma, A.; Cecchin, D.; Jain, S.A.; Kim, K.A.; Shin, Y.J.; Kim, E.H.; Kim, M.; et al. LRP-1 functionalized polymersomes enhance the efficacy of carnosine in experimental stroke. Sci. Rep. 2020, 10, 699. [Google Scholar] [CrossRef] [PubMed]
- Darvishi, B.; Dinarvand, R.; Mohammadpour, H.; Kamarul, T.; Sharifi, A.M. Dual l-Carnosine/Aloe vera Nanophytosomes with Synergistically Enhanced Protective Effects against Methylglyoxal-Induced Angiogenesis Impairment. Mol. Pharm. 2021, 18, 3302–3325. [Google Scholar] [CrossRef] [PubMed]
- Vistoli, G.; Carini, M.; Aldini, G. Transforming dietary peptides in promising lead compounds: The case of bioavailable carnosine analogs. Amino Acids 2012, 43, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Bellia, F.; Amorini, A.M.; La Mendola, D.; Vecchio, G.; Tavazzi, B.; Giardina, B.; Di Pietro, V.; Lazzarino, G.; Rizzarelli, E. New glycosidic derivatives of histidine-containing dipeptides with antioxidant properties and resistant to carnosinase activity. Eur. J. Med. Chem. 2008, 43, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Lanza, V.; Bellia, F.; D’Agata, R.; Grasso, G.; Rizzarelli, E.; Vecchio, G. New glycoside derivatives of carnosine and analogs resistant to carnosinase hydrolysis: Synthesis and characterization of their copper(II) complexes. J. Inorg. Biochem. 2011, 105, 181–188. [Google Scholar] [CrossRef]
- Kulikova, O.I.; Stvolinsky, S.L.; Migulin, V.A.; Andreeva, L.A.; Nagaev, I.Y.; Lopacheva, O.M.; Kulichenkova, K.N.; Lopachev, A.V.; Trubitsina, I.E.; Fedorova, T.N. A new derivative of acetylsalicylic acid and carnosine: Synthesis, physical and chemical properties, biological activity. Daru 2020, 28, 119–130. [Google Scholar] [CrossRef]
- Kobayashi, S.; Homma, T.; Okumura, N.; Han, J.; Nagaoka, K.; Sato, H.; Konno, H.; Yamada, S.; Takao, T.; Fujii, J. Carnosine dipeptidase II (CNDP2) protects cells under cysteine insufficiency by hydrolyzing glutathione-related peptides. Free Radic. Biol. Med. 2021, 174, 12–27. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regazzoni, L. State of the Art in the Development of Human Serum Carnosinase Inhibitors. Molecules 2024, 29, 2488. https://doi.org/10.3390/molecules29112488
Regazzoni L. State of the Art in the Development of Human Serum Carnosinase Inhibitors. Molecules. 2024; 29(11):2488. https://doi.org/10.3390/molecules29112488
Chicago/Turabian StyleRegazzoni, Luca. 2024. "State of the Art in the Development of Human Serum Carnosinase Inhibitors" Molecules 29, no. 11: 2488. https://doi.org/10.3390/molecules29112488
APA StyleRegazzoni, L. (2024). State of the Art in the Development of Human Serum Carnosinase Inhibitors. Molecules, 29(11), 2488. https://doi.org/10.3390/molecules29112488