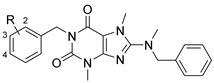
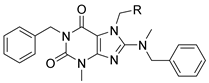
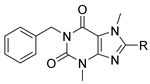
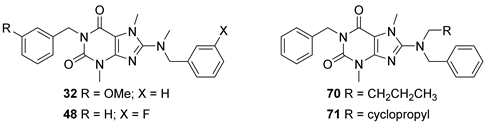
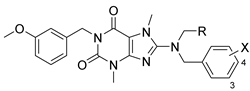
3.2. Analog Synthesis
Synthesis of analogs
16–
18,
21,
8, and
67 are described here as representative of the routes used to access the most active SLACK inhibitor compounds. Synthetic methods and characterization of all samples submitted for biological testing are available in the
Supplementary Material.
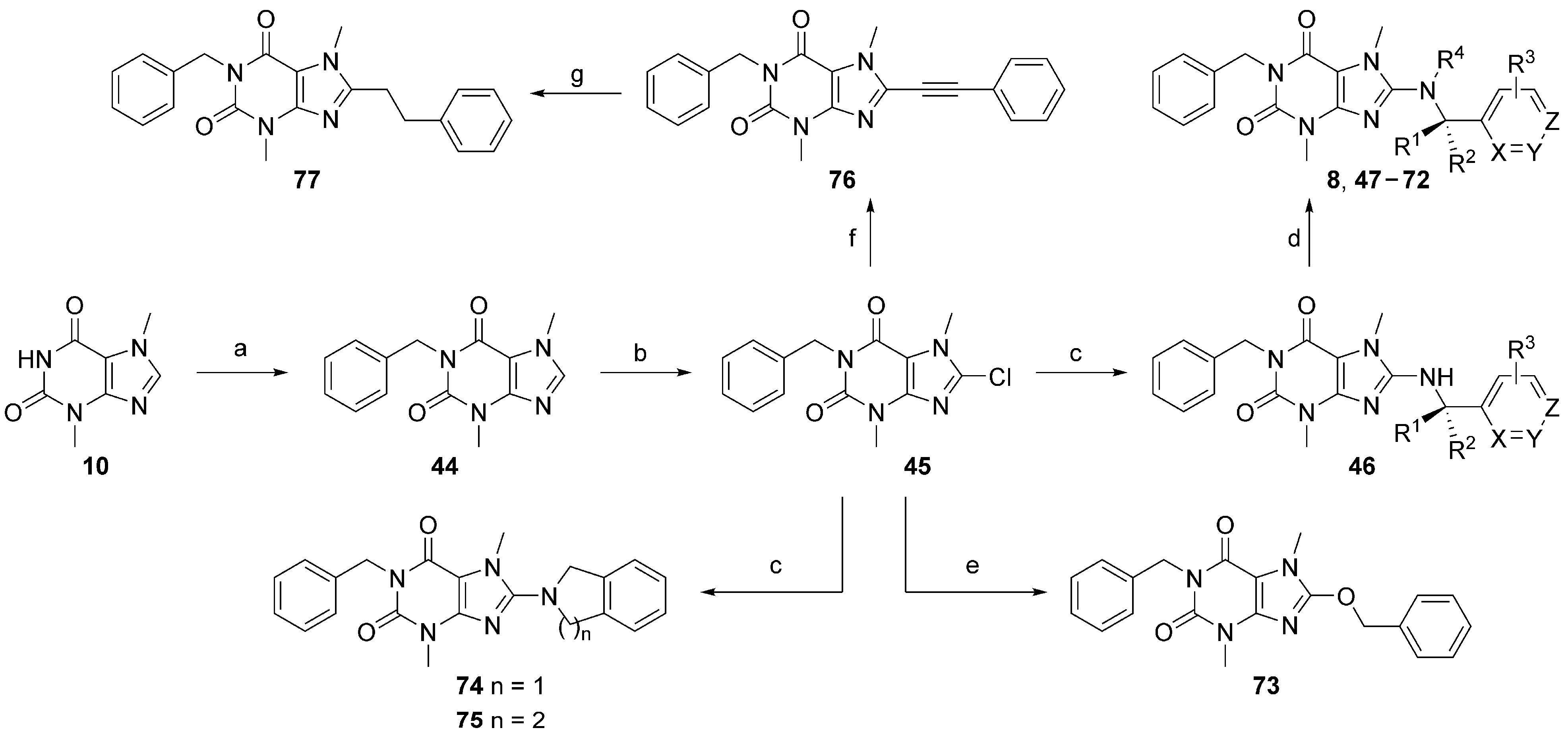
8-Chloro-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (14). Theobromine (10) (5.00 g, 27.8 mmol, 1.0 eq) and NCS (7.41 g, 55.5 mmol, 2.0 eq) were suspended in THF (25 mL), and the reaction was heated at 60 °C overnight. THF was removed in vacuo, and the residue was filtered and washed with water. The product was dried at 40 °C to provide 4.4 g (74%) of the title compound as a beige solid that was used further without purification. LCMS RT = 2.62 min; HRMS, calc’d for C7H8ClN4O2+ [M + H], 215.0330; found 215.0330.
8-(Benzyl(methyl)amino)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (15). Intermediate 14 (1.00 g, 4.66 mmol, 1.0 eq), N-methylbenzyl amine (1.13 g, 9.32 mmol, 2.0 eq), Cs2CO3 (3.03 g, 9.32 mmol, 2.0 eq), and DMF (5.0 mL) were added to a microwave vial and heated in a microwave reactor at 130 °C for 30 min. The reaction was cooled to room temperature, water was added, and the mixture was extracted with ethyl acetate (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 600 mg (43%) of the title compound. LCMS RT = 4.0 min; HRMS, calc’d for C15H18N5O2+ [M + H], 300.1455; found 300.1460.
8-(Benzyl(methyl)amino)-1-(2-fluorobenzyl)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (16). Sodium hydride (8.0 mg, 0.34 mmol, 2.0 eq) was added to an ice-cold solution of 15 (50 mg, 0.17 mmol, 1.0 eq) and was stirred for 15 min. Afterwards, 2-fluorobenzyl methanesulfonate (69 mg, 0.34 mmol, 2.0 eq) was added, and the reaction was heated at 130 °C for one hour. The reaction was cooled to room temperature, water was added, and the mixture was extracted with DCM (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 25 mg (36%) of the title compound. 1H NMR (300 MHz, CDCl3) δ 7.43–7.14 (m, 7H), 7.09–6.97 (m, 2H), 5.28 (s, 2H), 4.47 (s, 2H), 3.80 (s, 3H), 3.54 (s, 3H), 2.91 (s, 3H); 13C NMR (75 MHz, CDCl3) δ 162.33, 159.05, 157.58, 154.43, 151.61, 148.00, 136.77, 128.86 (d, J(C,F) = 4.07 Hz), 128.64 (d, J(C,F) = 8.29 Hz), 128.31 (d, J(C,F) = 69.62 Hz), 127.78, 124.68 (d, J(C,F) = 14.40 Hz), 124.00 (d, J(C,F) = 3.65 Hz), 115.35 (d, J(C,F) = 21.73 Hz), 105.03, 57.45, 38.91, 38.00 (d, J(C,F) = 4.97 Hz), 32.92, 29.76 ppm. LCMS RT = 5.25 min; HRMS, calc’d for C22H23FN5O2+ [M + H], 408.1830; found 408.1836.
8-(Benzyl(methyl)amino)-1-(3-fluorobenzyl)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (17). 3-Fluorobenzyl 4-methylbenzenesulfonate (95.3 mg, 0.34 mmol, 2.0 eq) was added to a solution of 15 (50 mg, 0.17 mmol, 1.0 eq) and Cs2CO3 (110 mg, 0.34 mmol, 2.0 eq) in THF (3 mL) and stirred at 60 °C overnight. The reaction was cooled to room temperature, water was added, and the mixture was extracted with DCM (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 15 mg (22%) of the title compound. 1H NMR (300 MHz, CDCl3) δ 7.41–7.21 (m, 7H), 7.17 (m, 1H), 6.93 (m, 1H), 5.17 (s, 2H), 4.47 (s, 2H), 3.81 (s, 3H), 3.53 (s, 3H), 2.91 (s, 3H); 13C NMR (75 MHz, CDCl3) δ 162.77 (d, J(C,F) = 245.42 Hz), 157.60, 154.42, 151.63, 147.97, 140.25 (d, J(C,F) = 7.40 Hz), 136.75, 129.79 (d, J(C,F) = 8.25 Hz), 128.77, 127.84, 127.78, 124.21 (d, J(C,F) = 2.80 Hz), 115.41 (d, J(C,F) = 21.82 Hz), 114.23 (d, J(C,F) = 21.04 Hz), 105.05, 57.44, 42.74 (d, J(C,F) = 1.67 Hz), 38.91, 32.91, 29.74 ppm. LCMS RT = 5.33 min; HRMS, calc’d for C22H23FN5O2+ [M + H], 408.1830; found 408.1835.
8-(Benzyl(methyl)amino)-1-(4-fluorobenzyl)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (18). 4-Fluorobenzylchloride (49 mg, 0.34 mmol, 2.0 eq) was added to a solution of 15 (50 mg, 0.17 mmol, 1.0 eq) and Cs2CO3 (110 mg, 0.34 mmol, 2.0 eq) in DMF (0.5 mL) and stirred at 100 °C overnight. The reaction was cooled to room temperature, water was added, and the mixture was extracted with DCM (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 28 mg (40%) of the title compound. 1H NMR (300 MHz, CDCl3) δ 7.49 (m, 2H), 7.41–7.27 (m, 5H), 6.97 (m, 2H), 5.14 (s, 2H), 4.46 (s, 2H), 3.80 (s, 3H), 3.52 (s, 3H), 2.90 (s, 3H); 13C NMR (75 MHz, CDCl3) δ 162.18 (d, J(C,F) = 245.07 Hz), 157.55, 154.50, 151.65, 147.88, 136.75, 133.65 (d, J(C,F) = 3.20 Hz), 130.74 (d, J(C,F) = 8.16 Hz), 128.76, 127.84, 127.77, 115.11 (d, J(C,F) = 21.19 Hz), 105.10, 57.45, 43.49, 38.92, 32.88, 29.71 ppm. LCMS RT = 5.38 min; HRMS, calc’d for C22H23FN5O2+ [M + H], 408.1830; found 408.1830.
8-(Benzyl(methyl)amino)-1-(4-chlorobenzyl)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (21). 4-Chlorobenzyl bromide (33 mg, 0.16 mmol, 2.0 eq) was added to a solution of 15 (25 mg, 0.08 mmol, 1.0 eq) and K2CO3 (22 mg, 0.16 mmol, 2.0 eq) in DMF (0.5 mL) and stirred at 100 °C overnight. The reaction was cooled to room temperature, water was added, and the mixture was extracted with DCM (2×). The combined organics were washed with brine, dried over sodium sulfate (Na2SO4), filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 24 mg (71%) of the title compound. 1H NMR (300 MHz, CDCl3) δ 7.43 (m, 2H), 7.40–7.21 (m, 7H), 5.13 (s, 2H), 4.46 (s, 2H), 3.80 (s, 3H), 3.52 (s, 3H), 2.91 (s, 3H); 13C NMR (75 MHz, CDCl3) δ 157.58, 154.44, 151.63, 147.93, 139.74, 136.34, 133.15, 130.31, 128.77, 128.47, 127.84, 127.78, 105.07, 57.44, 43.57, 38.91, 32.90, 29.72 ppm. LCMS RT = 5.59 min; HRMS, calc’d for C22H23ClN5O2+ [M + H], 424.1535; found 424.1540.
1-Benzyl-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (44). Theobromine (10) (1.58 g, 8.78 mmol, 1.5 eq) and K2CO3 (1.62 g, 11.7 mmol, 2.0 eq) were suspended in DMF (8.0 mL), followed by the addition of benzyl bromide (1.00 g, 5.85 mmol, 1.0 eq). The reaction was heated at 90 °C overnight. The reaction was cooled to room temperature, water was added, and the residue was filtered and rinsed with water. The product was dried at 40 °C to provide 1.50 g (95%) of the title compound as a white powder that was used further without purification. LCMS RT = 3.98 min; HRMS, calc’d for C14H15N4O2+ [M + H], 271.1190; found 271.1192.
1-Benzyl-8-chloro-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (45). Intermediate 44 (1.50 g, 5.55, 1.0 eq) and NCS (1.48 g, 11.1 mmol, 2.0 eq) were suspended in THF (15 mL), and the reaction was heated at 60 °C overnight. THF was removed in vacuo, and the residue was filtered and washed with water. The product was dried at 40 °C to provide 1.31 g (78%) of the title compound as a beige solid that was used further without purification. 1H NMR (300 MHz, CDCl3) δ 7.47 (m, 2H), 7.40–7.18 (m, 3H), 5.18 (s, 2H), 3.95 (s, 3H), 3.53 (s, 3H). LCMS RT = 4.64 min; HRMS, calc’d for C14H14ClN4O2+ [M + H], 305.0800; found 305.0799.
1-Benzyl-8-(benzyl(methyl)amino)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione [VU0607689] (8). Intermediate 45 (50 mg, 0.16 mmol, 1.0 eq), benzyl amine (34 mg, 0.32 mmol, 2.0 eq), Cs2CO3 (104 mg, 0.32 mmol, 2.0 eq), and DMF (0.5 mL) were added to a microwave vial and heated in a microwave reactor at 130 °C for 30 min. The reaction was cooled to room temperature, water was added, and the mixture was extracted with ethyl acetate (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded impure product that was dissolved in THF (2.0 mL) and cooled to 0 °C, followed by the addition of sodium hydride (15 mg, 0.64 mmol, 4.0 eq). The solution was stirred for 30 min, followed by the addition of methyl iodide. The reaction temperature was brought to 50 °C and stirred at that temperature for 1 h. The reaction was cooled to room temperature, water was added, and the mixture was extracted with DCM (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 20 mg (32%) of the title compound. 1H NMR (300 MHz, CDCl3) δ 7.49 (m, 2H), 7.41–7.18 (m, 8H), 5.19 (s, 2H), 4.45 (s, 2H), 3.80 (s, 3H), 3.52 (s, 3H), 2.90 (s, 3H); 13C NMR (75 MHz, CDCl3) δ 157.49, 154.61, 151.70, 147.83, 137.83, 136.79, 128.76, 128.74, 128.35, 127.86, 127.76, 127.32, 105.14, 57.48, 44.22, 38.93, 32.86, 29.70 ppm. LCMS RT = 5.26 min; HRMS, calc’d for C22H24N5O2+ [M + H], 390.1925; found 390.1932.
1-Benzyl-8-(benzylamino)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (46a). Intermediate 45 (700 mg, 2.30 mmol, 1.0 eq), benzyl amine (493 mg, 4.60 mmol, 2.0 eq), Cs2CO3 (1.50 g, 4.60 mmol, 2.0 eq), and DMF (5.0 mL) were added to a microwave vial and heated in a microwave reactor at 130 °C for 30 min. The reaction was cooled to room temperature, water was added, and the mixture was extracted with ethyl acetate (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 650 mg (38%) of the title compound. 1H NMR (300 MHz, CDCl3) δ 7.47 (m, 2H), 7.41–7.16 (m, 8H), 5.18 (s, 2H), 4.66 (d, J = Hz, 2H), 4.40 (m, 1H), 3.66 (s, 3H), 3.52 (s, 3H); 13C NMR (75 MHz, CDCl3) δ 154.12, 153.21, 151.69, 148.73, 138.13, 137.92, 128.87, 128.61, 128.34, 127.98, 127.26, 103.37, 47.43, 44.11, 29.72, 29.63 ppm. LCMS RT = 4.87 min; HRMS, calc’d for C21H22N5O2+ [M + H], 376.1768; found 376.1774.
1-Benzyl-8-(benzyl(ethyl)amino)-3,7-dimethyl-3,7-dihydro-1H-purine-2,6-dione (67). Compound 46a (50 mg, 0.13 mmol, 1.0 eq) was dissolved in THF (2.0 mL) and cooled to 0 °C, followed by the addition of NaH (12.5 mg, 0.52 mmol, 4.0 eq). The solution was stirred for 30 min, followed by the addition of iodoethane. The reaction temperature was brought to 50 °C and stirred for 1 h. The reaction was cooled to room temperature, water was added, and the mixture was extracted with DCM (2×). The combined organics were washed with brine, dried over Na2SO4, filtered, and concentrated in vacuo. Purification of the residue by flash chromatography on silica gel yielded 21 mg (40%) of the title compound. 1H NMR (300 MHz, CDCl3) δ 7.49 (m, 2H), 7.39–7.18 (m, 8H), 5.18 (s, 2H), 4.54 (s, 2H), 3.76 (s, 3H), 3.51 (s, 3H), 3.27 (q, J = 7.11 Hz, 2H), 1.15 (t, J = 7.11 Hz, 3H); 13C NMR (75 MHz, CDCl3) δ 156.56, 154.62, 151.69, 147.78, 137.83, 137.44, 128.77, 128.62, 128.34, 127.95, 127.59, 127.32, 105.10, 54.71, 46.00, 44.21, 32.58, 29.71, 12.68 ppm. LCMS RT = 5.51 min; HRMS, calc’d for C23H26N5O2+ [M + H], 404.2081; found 404.2088.



 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}