Abstract
Strigolactones (SLs) have potential to be used in sustainable agriculture to mitigate various stresses that plants have to deal with. The natural SLs, as well as the synthetic analogs, are difficult to obtain in sufficient amounts for practical applications. At the same time, fluorescent SLs would be useful for the mechanistic understanding of their effects based on bio-imaging or spectroscopic techniques. In this study, new fluorescent SL mimics containing a substituted 1,8-naphthalimide ring system connected through an ether link to a bioactive furan-2-one moiety were prepared. The structural, spectroscopic, and biological activity of the new SL mimics on phytopathogens were investigated and compared with previously synthetized fluorescent SL mimics. The chemical group at the C-6 position of the naphthalimide ring influences the fluorescence parameters. All SL mimics showed effects similar to GR24 on phytopathogens, indicating their suitability for practical applications. The pattern of the biological activity depended on the fungal species, SL mimic and concentration, and hyphal order. This dependence is probably related to the specificity of each fungal receptor–SL mimic interaction, which will have to be analyzed in-depth. Based on the biological properties and spectroscopic particularities, one SL mimic could be a good candidate for microscopic and spectroscopic investigations.
1. Introduction
Strigolactones (SLs) are carotenoid derivatives with multiple functions in plants [1,2]. Their initial function in the terrestrial plants seems to be related to rhizosphere communication [3]. In Physcomitrella patens, moss SL acts a neighbor signal, similar to bacterial quorum sensing signals [4]. P. patens SL defective mutants fail in the detection of neighboring colonies [4]. Similarly, pea (Pisum sativum) mutant, defective in SL biosynthesis, fails to detect neighboring plants [5]. Strigolactones are involved in the recruitment of the arbuscular mycorrhizal (AM) fungi [6,7,8] and symbiotic nitrogen-fixing rhizobia [9,10,11]. The difference in SL exudation between wild plants and SL-defective plant mutants shape differently the rhizosphere microbiome in Arabidopsis thaliana [12], soybean (Glycine max) [13], and rice (Oryza sativa) [14]. The rhizosphere SLs are also used as a cue for plant root presence by the parasitic plant seeds [15,16,17,18] and plant pathogenic fungi [19].
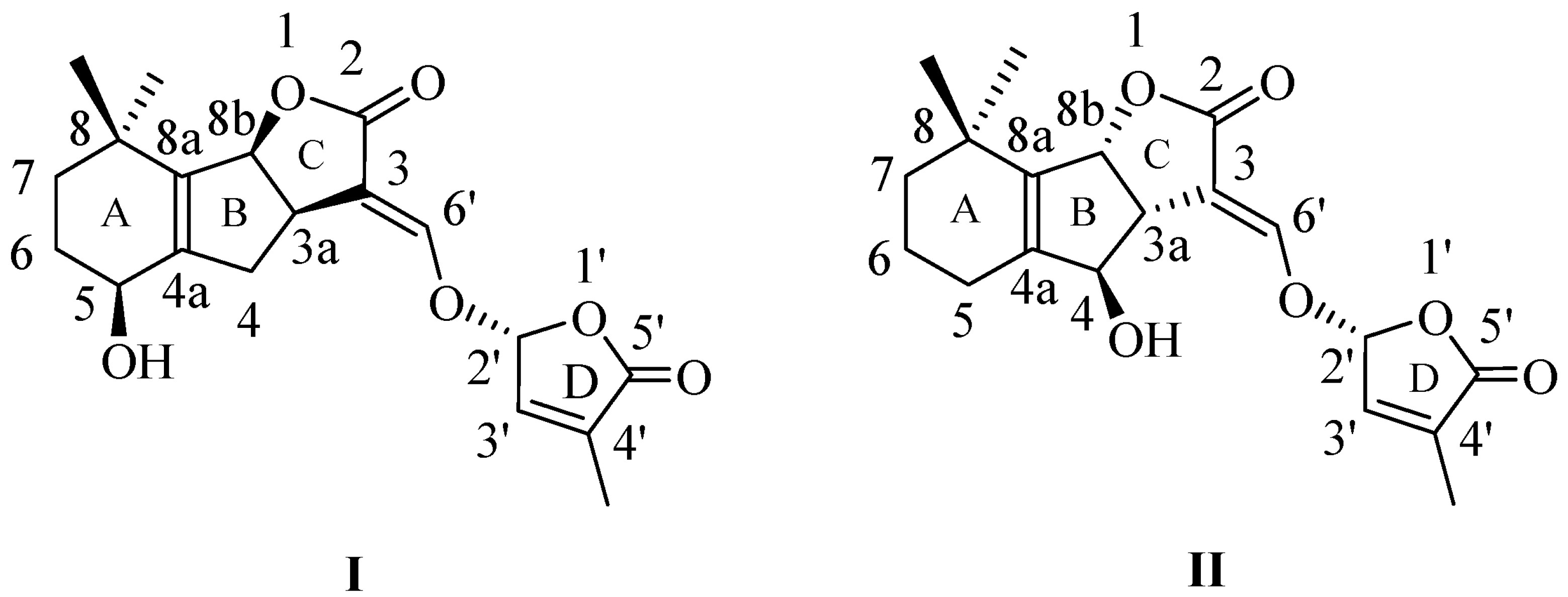
SLs have been recognized as a class of phytohormones involved in plant development [20,21,22,23,24,25,26,27,28,29,30] and in the orchestration of the plant responses to abiotic stresses [31,32,33]. The initial described natural SLs have complex structures containing a tricyclic lactone system (A, B, and C rings) connected through an enol-ether bond to a furan-2-one ring (D)—canonical SLs [34] (Figure 1).
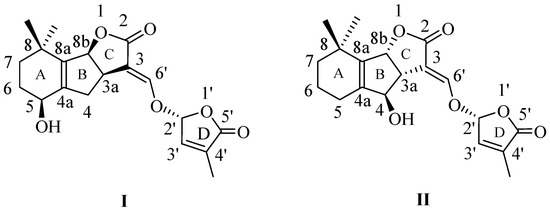
Figure 1.
Naturally occurring canonical strigolactone structures, from the families (+)-strigol (I) and (−)-orobanchol (II).
In the last decade, natural SLs that do not have the specific tricyclic lactone system (A, B, and C rings) were discovered—the non-canonical SLs, with the bioactiphore and the butenolide D-ring but without the A-, B-, or a closed C-ring [35]. The synthetic SLs further expand the structural varieties, SL analogs, and SL mimics demonstrating specific SL activities.
The SL multifaceted roles, as endo- and exo-signals, are related to the presence of C (open or closed) and D rings, connected via an enol ether bridge. This part of the SLs (canonical or non-canonical) bind to similar receptors from the (super)family of α/β hydrolase fold proteins and trigger the cascade reactions that determine the specific response to SLs in different organisms [36].
Naturally occurring SLs are released from plants in tiny amounts. Complex structures and laborious synthetic procedures for the multi-gram amounts of natural SLs have raised interest in synthesizing simplified molecules that display various SL bioactivities. Therefore, synthetic SL analogs that maintain the bioactiphore CD part of natural SLs [37,38,39,40,41,42,43,44,45] were synthetized. These SLs analogs are structurally more related to canonical SLs. Simpler SL mimics, more structurally related to non-canonical SLs, were proposed. In these SL mimics where the methyl-substituted D-ring is directly linked to an arylthiol, aryloxy or aroyloxy moieties have been synthesized [43,46,47,48].
The list of synthetic SL analogs includes GR24, (3E,3aR,8bS)-rel-3-[[[(2R)-2,5-dihydro-4-methyl-5-oxo-2-furanyl]oxy]methylene]-3,3a,4,8b-tetrahydro-2H-indeno[1,2-b]furan-2-one, named after its inventor Gerald Rosebery, which was first synthesized in 1981 [38] and largely used as a strigolactone (strigol type) synthetic replacer in laboratory studies; GR7, with the A-ring removed from GR24 [38]; Nijmegen-1, tested in field conditions as a suicidal germination stimulant for Orobanche ramosa [37]; CISA-1, a fluorescent alternative to GR24 [49]; and PLN65, PLO65, and PLS6—conjugated fluorescent strigolactone analogs [50]. Recently published were analogs of non-canonical SLs, cannalactone analogs named (±)-SdL19 and (±)-SdL118, that stimulate the germination of Phelipanche ramose and exhibit other SL activities, e.g., the inhibition of plant shoot branching and stimulation of hyphal branching in AM fungus Rhizophagus irregularis [51]. Strigolactone mimics include 4BD, 4-bromodebranone [52]; PL02, N-(4-methyl-5-oxo-2,5-dihydrofuran-2-yl)-3-nitrophthalimide, and other similar phthalimide-derived compounds [53]; SL-6, 2-(4-methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-benzo[de]isoquinoline-1,3-dione [54]; contalactone, i.e., contaminant lactone of GR24 [55]; and carlactonic acid, rac-/(R)-/(S)-(E,2E)-2-[(4-Methyl-5-oxo-2H-furan-2-yl)oxymethylene]-4-(2,6,6-trimethylcyclohexen-1-yl)but-3-enoic acid, and its methyl derivative [56]. A detailed list of SL analogs and mimics is presented in several reviews [28,57,58].
Structure–activity relationship data on SLs suggest the existence of different perception systems for their various biological activities. In plants, DWARF14 (D14), an α/β hydrolase, is the strigolactone main receptor [59], which evolved by duplication and differentiation from the karrikin (KAR) receptor KAI2 [3]. In parasitic plants, the SL perception as a cue is performed by a paralog of KAI2, α/β hydrolase ShHTLs/ShKAI2s [60,61]. In fungal plant pathogen Cryphonectria parasitica, CpD14, a structural homolog of plant receptor D14, is involved in the SL response. Many fluorescent SL analogs or mimics, most of which bear known fluorophore molecules, have already been prepared to reveal all protein receptors and the distribution of SLs in plants, and to elucidate their mode of action [49,50,62,63,64,65,66].
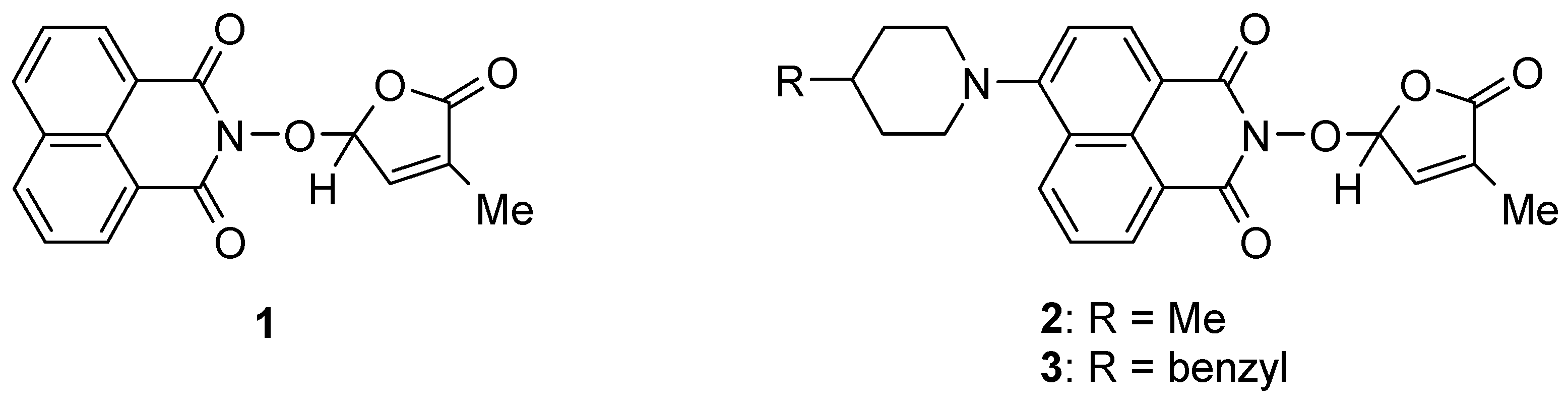
Continuing our research on bioactive compounds [54,67,68,69], we have been interested in developing new bioactive and fluorescent SL mimics suitable for bio-imaging studies in fungi and/or plant cells. Considering the potential exploitation of SL mimics as active ingredients of the plant biostimulants, due to their direct effect on plant tolerance to abiotic stress, and especially to drought [31,70,71] and/or mediators of plant interaction with beneficial and/or detrimental rhizosphere organisms for plants, as a component of next-generation plant biostimulants [72], we developed new fluorescent SL mimics derived from the 1,8-naphthalimide fluorophore molecule. The 1,8-Naphthalinimide ring system is the central component of different fluorescent compounds with potential applications in various fields [73,74,75,76,77,78,79]. We already reported an SL mimic bearing the 1,8-naphthalimide system, namely 2-(4-methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-benzo[de]isoquinoline-1,3-dione 1 (Figure 2), and its spectroscopic and biological characteristics [54,80].
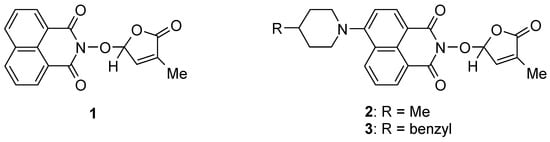
Figure 2.
SL mimics derived from 1,8-naphthalimide.
We reported other, new fluorescent SL-mimics, with the amino-substituted 1,8-naphthalimide ring system, namely 2-(4-methyl-5-oxo-2,5-dihydrofuran-2-yloxy)-6-(4-methylpiperidin-1-yl)-benzo[de]isoquinoline-1,3-dione 2 and 2-(4-methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-6-(4-benzylpiperidin-1-yl)-benzo[de]isoquinoline-1,3-dione 3 (Figure 1) [81]. They have been prepared via successive reactions of 4-chloro-1,8-naphthalic anhydride 4 with 4-methylpiperidine and 4-benzylpiperidine, respectively; reactions of the resulting 4-amino-1,8-naphthalic anhydrides with hydroxylamine hydrochloride to give the fluorescent 2-hydroxy-6-amino-1,8-naphthalimide intermediates [79]; and condensations of 2-hydroxy-6-amino-1,8-naphthalimide intermediates with 5-bromo-3-methyl-5H-furan-2-one 6 to access fluorescent SL mimics 2 and 3, respectively [81].
Interesting biological and fluorescent properties of the previously reported SL mimics bearing a 1,8-naphthalimide system [54,80,81] prompted us to prepare new, potentially bioactive and fluorescent SL mimics aiming to develop SL-mimics-based agricultural inputs that modulate plant interactions with fungal pathogens. Thus, two other new SL mimics containing halo-substituted 1,8-naphthalimide molecules connected through an ether link to a bioactive furan-2-one unit have been prepared and fully characterized. They have been used in an alternative synthetic procedure towards the previously reported SL mimics 2 and 3, respectively. The biologic properties of both the growth inhibition of phytopathogenic fungi and the effects on the hyphal branches have been investigated for all these SL mimics.
2. Results and Discussion
2.1. Synthesis of 1,8-Naphthalimide-Derived Strigolactone Mimics
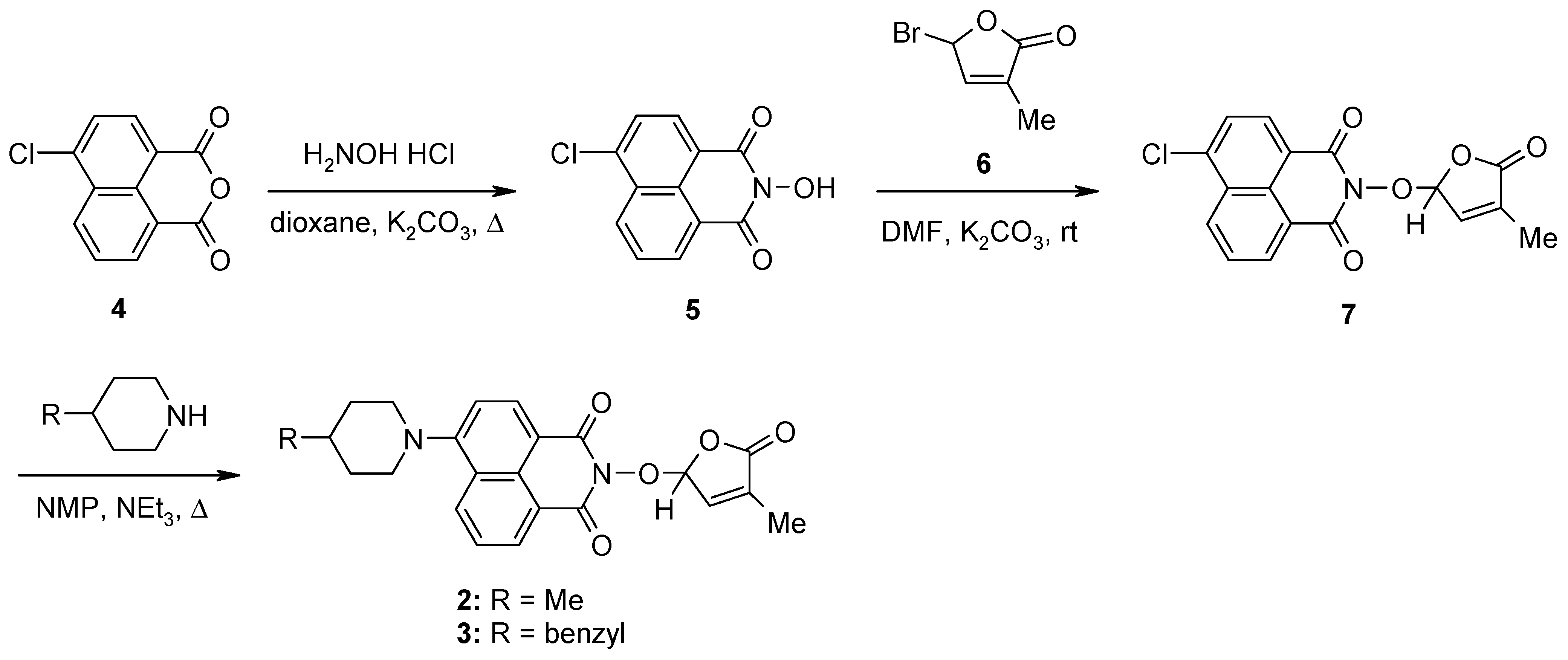
The synthesis of the potentially bioactive and fluorescent SL mimics started from commercially available 4-chloro-1,8-naphthalic anhydride 4. The reactions of 4 with hydroxylamine hydrochloride in dioxane led to 2-hydroxy-6-chloro-1,8-naphthalimide intermediate 5 [79]. The condensation reaction of the resulting 2-hydroxy-6-chloro-1,8-naphthalimide intermediate 5 with common intermediate 5-bromo-3-methyl-5H-furan-2-one 6 carried out in DMF, using anhydrous potassium carbonate as the basic catalyst, gave a new SL mimic, namely 6-chloro-2-(4-methyl-5-oxo-2,5-dihydrofuran-2-yloxy)-benzo[de] isoquinoline-1,3-dione 7 (Scheme 1). The common intermediate in the SL mimic syntheses, i.e., 5-bromo-3-methylfuran-2-one 6, was prepared by brominating the commercially technical product 3-methyl-5H-furan-2-one with N-bromosuccinimide (NBS), in the presence of a radical initiator [82]. Based on the new SL mimic 7, we developed a new synthetic route towards previously reported putative fluorescent SL mimics 2 and 3 [81]. The two putative fluorescent SL mimics 2 and 3 were easily prepared by the coupling reactions of SL mimic 7 with cyclic secondary amines 4-methylpiperidine and 4-benzylpiperidine, respectively. These reactions took place in N-methylpyrrolidone (NMP), in the presence of triethylamine, at the reflux temperature of the reaction mixture. In this way, we developed a new synthetic protocol which starts from commercially available 4-chloro-1,8-naphthalic anhydride 4 and gave easier access to 1,8-naphthalimide-based SL mimics 7, 2, and 3 (Scheme 1).
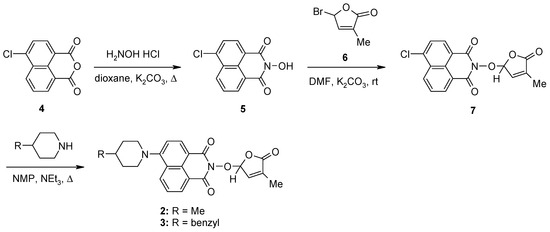
Scheme 1.
The synthetic procedure for SL mimics 7, 2, and 3.
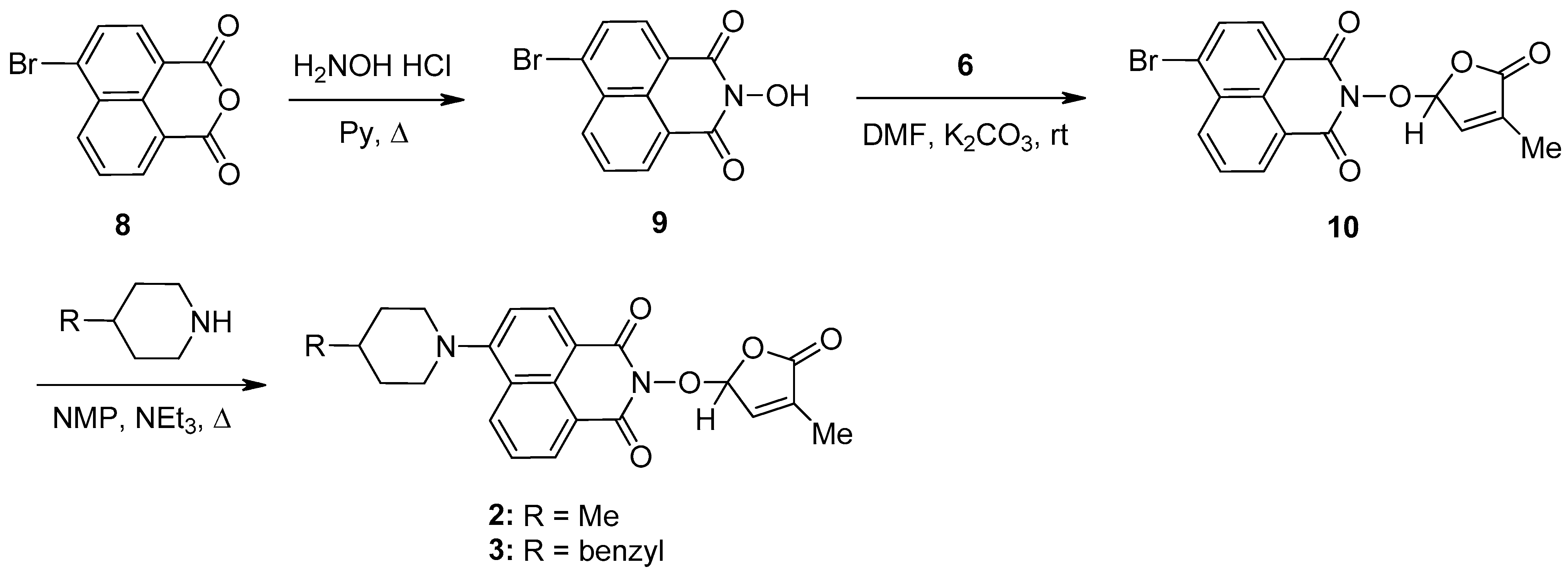
A new potentially bioactive SL mimic, namely 6-bromo-2-(4-methyl-5-oxo-2,5-dihydrofuran-2-yloxy)-benzo[de] isoquinoline-1,3-dione 10 (SL-27), was also easily prepared via the reaction of commercially available 4-bromo-1,8-naphthalic anhydride 8 with hydroxylamine hydrochloride in pyridine [83] to obtain 2-hydroxy-6-bromo-1,8-naphthalimide intermediate 9, followed by the coupling reaction of intermediate 9 with 5-bromo-3-methylfuran-2-one intermediate 6 (Scheme 2). The yields in 2-hydroxy-6-halo-1,8-naphthalimide intermediate 8 and 9, respectively, were similar (about 90%). The two fluorescent SL mimics 2 and 3 were again easily prepared by the coupling reactions of SL mimic 10 with 4-methylpiperidine and 4-benzylpiperidine, respectively, in N-methylpyrrolidone at reflux temperature, in the presence of triethylamine. In this way, starting from commercially 4-bromo-1,8-naphthalic anhydrides 8, we gained access to 1,8-naphthalimide-based SL mimics 10, 2, and 3. The final yields in SL mimics were similar irrespective of whether the synthetic procedure started from 4-chloro- or 4-bromo-1,8-naphthalic anhydride.
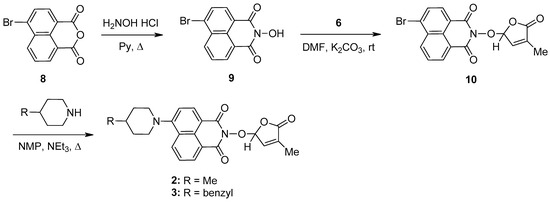
Scheme 2.
The synthetic procedure for SL mimics 10, 2, and 3.
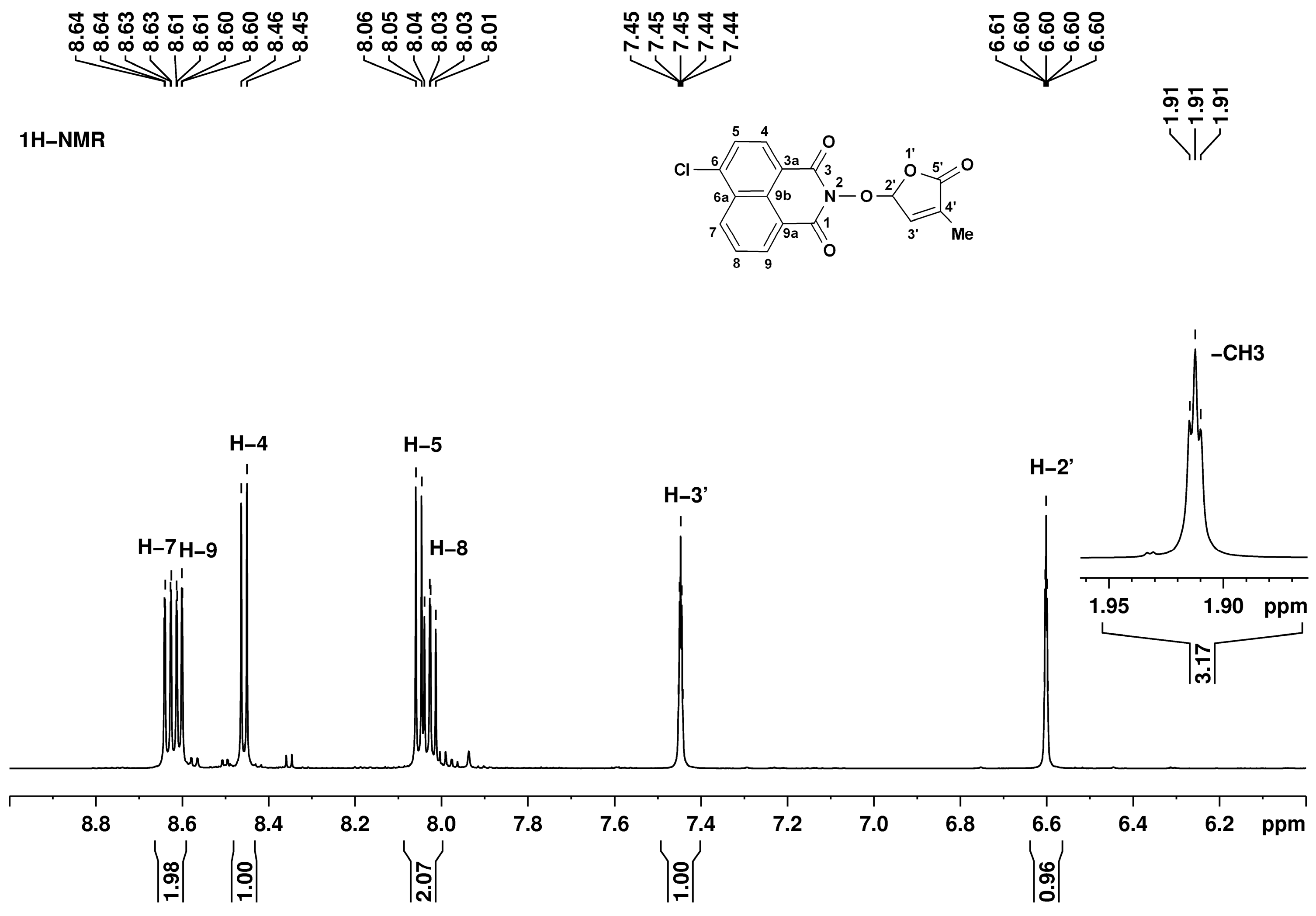
The structures of all synthesized compounds were proven by 1H, 13C, and 15N NMR spectroscopy. The 1,8-naphthalimide and 3-methylfuran-2-one fragments have individual resonance signals, readily assigned based on the bond correlations from bidimensional homo- and heteronuclear experiments. For example, in the 1H NMR spectrum corresponding to SL mimic 7, presented in Figure 3, there are five signals in the low-field region (above 8.00 ppm) assigned to the naphthalimide fragment as follows: 8.03 ppm doublet of doublets H-8, 8.05 ppm doublet H-5, 8.45 ppm doublet H-4, 8.61 ppm doublet of doublets H-9, and 8.63 ppm doublet of doublets H-7. The other fragment, 3-methylfuran-2-one, has three characteristic signals at 1.91 (triplet, CH3), 6.60 (quintet, H-2′), and 7.45 ppm (quintet, H-3′).
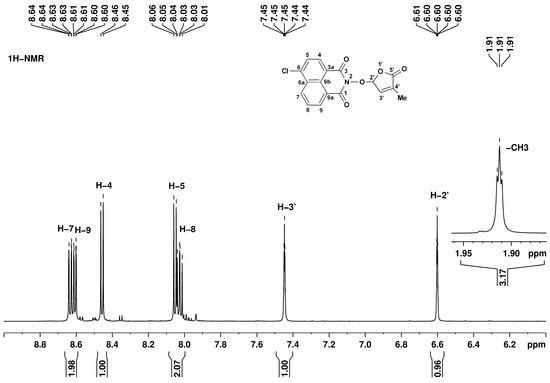
Figure 3.
1H NMR spectrum corresponding to SL mimic 7, recorded in DMSO-d6, at 600 MHz. Signal assignments are annotated on the figure.
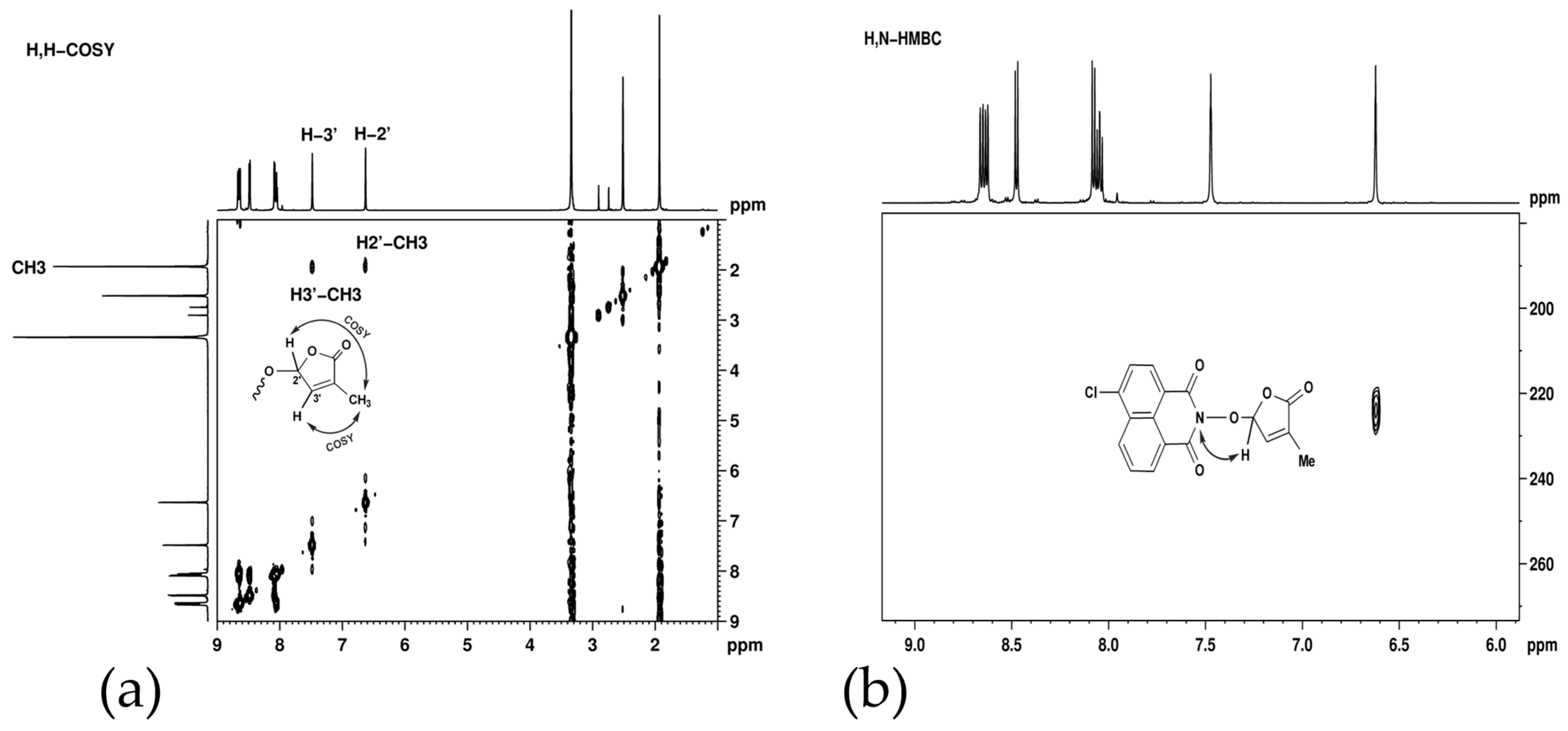
Long-range proton–proton couplings, over four and five bonds, were observed in H,H-COSY (Figure 4a) between methyl protons and the two furan cycle protons. These couplings, facilitated by the double bond, are responsible for the unexpected signal shapes: triplet for CH3 and quintet for the two CH groups.
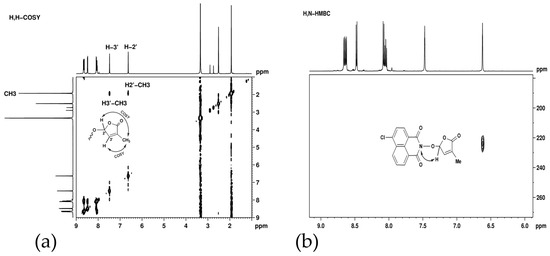
Figure 4.
(a) H,H-COSY spectrum of SL mimic 7 showing long-range correlations between methyl and furan cycle protons; (b) H,N-HMBC spectrum corresponding to SL mimic 7 showing a correlation signal between the nitrogen atom from 224 ppm and the furan cycle proton from 6.60 ppm.
The successful formation of the naphthalimide intermediate and its consequent condensation with 5-bromo-3-methyl-5H-furan-2-one were demonstrated from long-range proton–nitrogen interactions—Figure 4. In the H,N-HMBC spectrum corresponding to SL mimic 7 (Figure 4b), a correlation signal was observed between the nitrogen atom from 224 ppm and the furan cycle proton from 6.60 ppm.
The exact molecular weights were confirmed by high-resolution mass spectrometry and the consistency of the molecular formulas was assessed by comparison of the experimental and simulated isotopic patterns. The proton and carbon NMR assignments and coupling constants are presented in the experimental section. All NMR spectra together with the experimental and simulated MS isotopic patterns are presented as Supplementary Materials (Figures S1–S12).
The electronic absorption spectra of the 1,8-naphthalimide-based strigolactone mimics 7 and 10 in solvents with different polarities such as toluene (PhMe, ε = 2.38), dichloromethane (DCM, ε = 9.08), and N,N-dimethylformamide (DMF, ε = 38.25) were investigated. The spectral parameters for these compounds, namely absorption maxima (λmax), emission maxima (λem), and Stokes shifts (Δν), are listed in Table 1. In Figures S13 and S14, the electronic absorption and, respectively, emission spectra in dichloromethane are presented for SL mimics 7 and 10 (see Supplementary Materials).

Table 1.
Spectral and photophysical characteristics of SL mimics 7 and 10.
SL mimics 1, 7, and 10 present absorption bands in the range of 300–370 nm in DCM (Table 1) with maxima centered around 335 nm (1), 343.5 nm (7), and 345 nm (10), respectively, arising from a π-π* electron transition of the 1,8-naphthalimide ring [75,76,77,78,79,80,81]. Expectably, the introduction of halogen groups in the C-6 position of the naphthalimide ring (SL mimics 7 and 10) determines a red shift in the absorption maximum as compared to the unsubstituted SL mimic 1. The solvent polarity has a small effect on the absorption maxima of SL mimics 7 and 10 (Table 1). The SL mimics containing the piperidine substituent at the 1,8-naphthalimide unit in the C-6 position (SL mimics 2 and 3) display the longest-wavelength absorption in the visible range without a clear vibrational structure [81]. Changing the solvent polarity from nonpolar toluene to polar DMF, a red shift in the position of the absorption band of 11 nm was found and this red shift was 15 nm in DCM for 2.
The fluorescence characteristics of these SL mimics are displayed in Table 1. In DMF solutions, SL mimics 7 and 10 show an intense violet emission at about 400 nm. From Table 1, it is noticed that the influence of the solvent polarity on the fluorescence maxima is more pronounced than that on the electronic absorption spectra. The Stokes shift (Δν) reveals the difference in the structure and properties of fluorophores between the ground and the first excited state, estimated according to Equation (1):
The values Δν for SL mimics under study are in the 3200–9500 cm−1 range in the three solvents, which are in agreement with the values reported for other naphthalimide derivatives [75,78,81]. For SL mimics 7 and 10, a hypsochromic shift in emission maxima was induced when the solvent was changed from toluene to DMF. This blue shift can indicate that a decrease in the electron density can occur in the excited state relating to the ground state.
The photophysical parameters of these derivatives such as quantum yield and fluorescence lifetimes are listed in Table 1 and Table 2. It is observed that the values of the fluorescence quantum yield depend significantly on the solvent nature.
Table 2.
Fluorescence lifetimes of SL mimics 7 and 10.
SL mimic 7 presents moderately high values of quantum yield in toluene (5.49%) and DCM (6.82%), whereas in DMF, a very small value was obtained (0.01%). Replacing chlorine in C-6 of the 1,8-naphthalimide ring with bromine determined an evident decrease in the quantum yield value for SL mimic 10 in toluene (0.65%) and DCM (1.57%), but a slight increase in DMF (0.89%).
The emission profiles were fitted to one, two, or three exponential curves, depending on the derivative structure and solvent. The fluorescence decay traces of naphthalimide derivatives 1, 7, and 10 are displayed in Figures S15 and S16 (see Supplementary Materials). From the fitted decay parameters, the decay profile of 7 exhibited two lifetime components in toluene with a predominance of the short lifetime around 0.94 ns and an amplitude of 82.14%, whereas in DMF, a monoexponential decay was noticed with a very short lifetime. The bromine derivative 10 presented a biexponential decay in DMF, leading to emission lifetimes of 0.33 ns (53.86%) and 4.99 ns (46.14%), respectively, whereas in DCM, a triexponential decay was observed (Table 2). The increase in solvent polarity leads to a decrease in the fluorescence lifetime in DMF, which was correlated with the changes in the emission quantum yield. It must be mentioned that the highest values of the fluorescence quantum yield were obtained for 1,8-naphthalimide derivatives with a piperidine substituent at the C-6 position of the naphthalimide ring (SL mimics 2 and 3) in nonpolar solvents (see Supplementary Material, Tables S1 and S2) [81]. Consequently, it would be suitable to investigate their temporal distribution in planta or in fungal organisms or to study the activity of SL receptors in vitro [84,85,86,87] after the hydrolysis of the bioactive 3-methyl-5H-furan-2-one unit.
2.2. Biological Activity of Synthesized Compounds
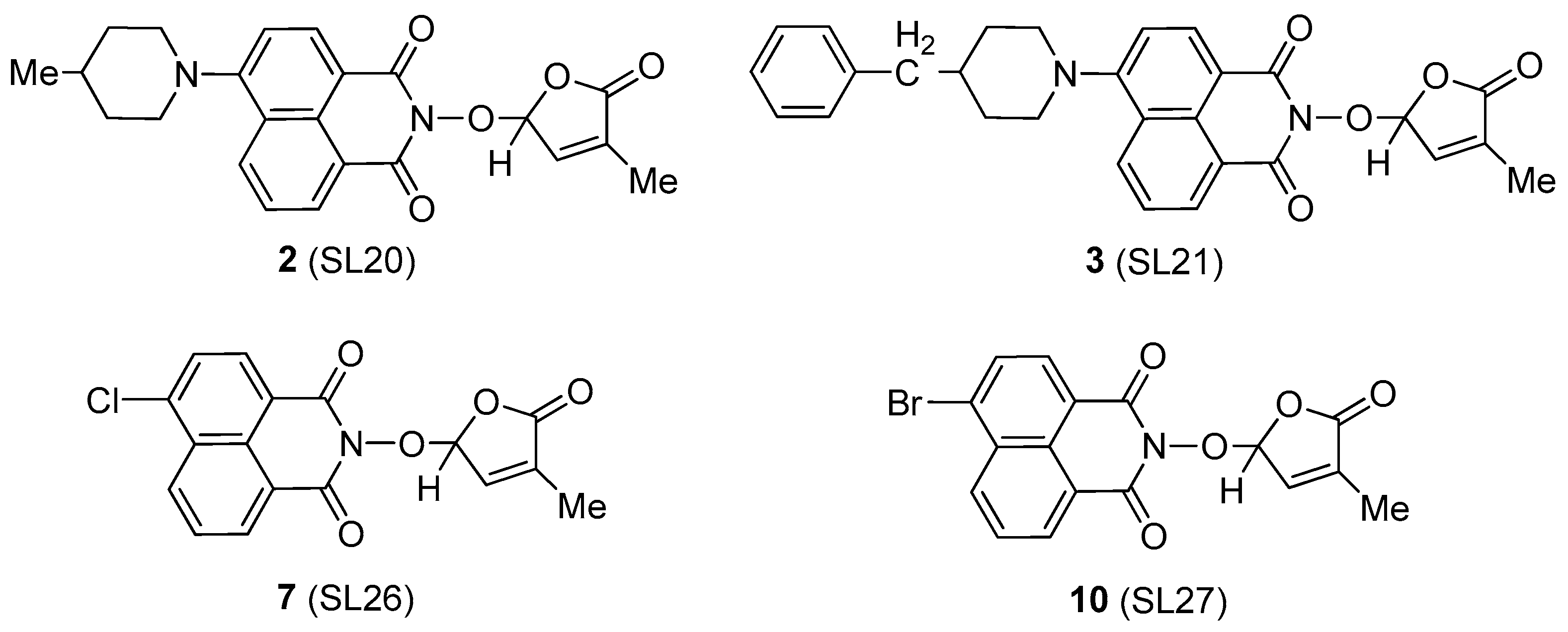
The influence of the 6-substituted 1,8-naphthalimide-derived SL mimics 2, 3, 7, and 10 (Scheme 3) on radial fungal growth and hyphal branching was investigated.
Scheme 3.
The investigated strigolactone mimics.
For presenting the biological activity, we shall continue to use the final given names for the synthesized SL mimics, i.e., SL20 for mimic 2, SL21 for mimic 3, SL26 for mimic 7, and SL27 for mimic 10. These given names are essential for reporting further tests with these molecules. We monitored the SL dose dependence by testing three SL concentrations: 5 × 10−6 M (C1), 10−5 M (C2), and 5 × 10−5 M (C3). The reference compound was GR24, an SL analog with a proven effect on the branching of plant pathogenic fungi [19,88]. This SL analog was developed from strigol as a basic blueprint and includes a tricyclic lactone system (A, B, and C rings) connected through an enol-ether bond to a furan-2-one ring (D) [38]. The effects were tested on the following fungal strains: Fusarium graminearum DSM 4527, Rhizoctonia solani DSM 22842, Sclerotinia sclerotium DSM 1946, and Colletotrichum acutatum CBS 113008.
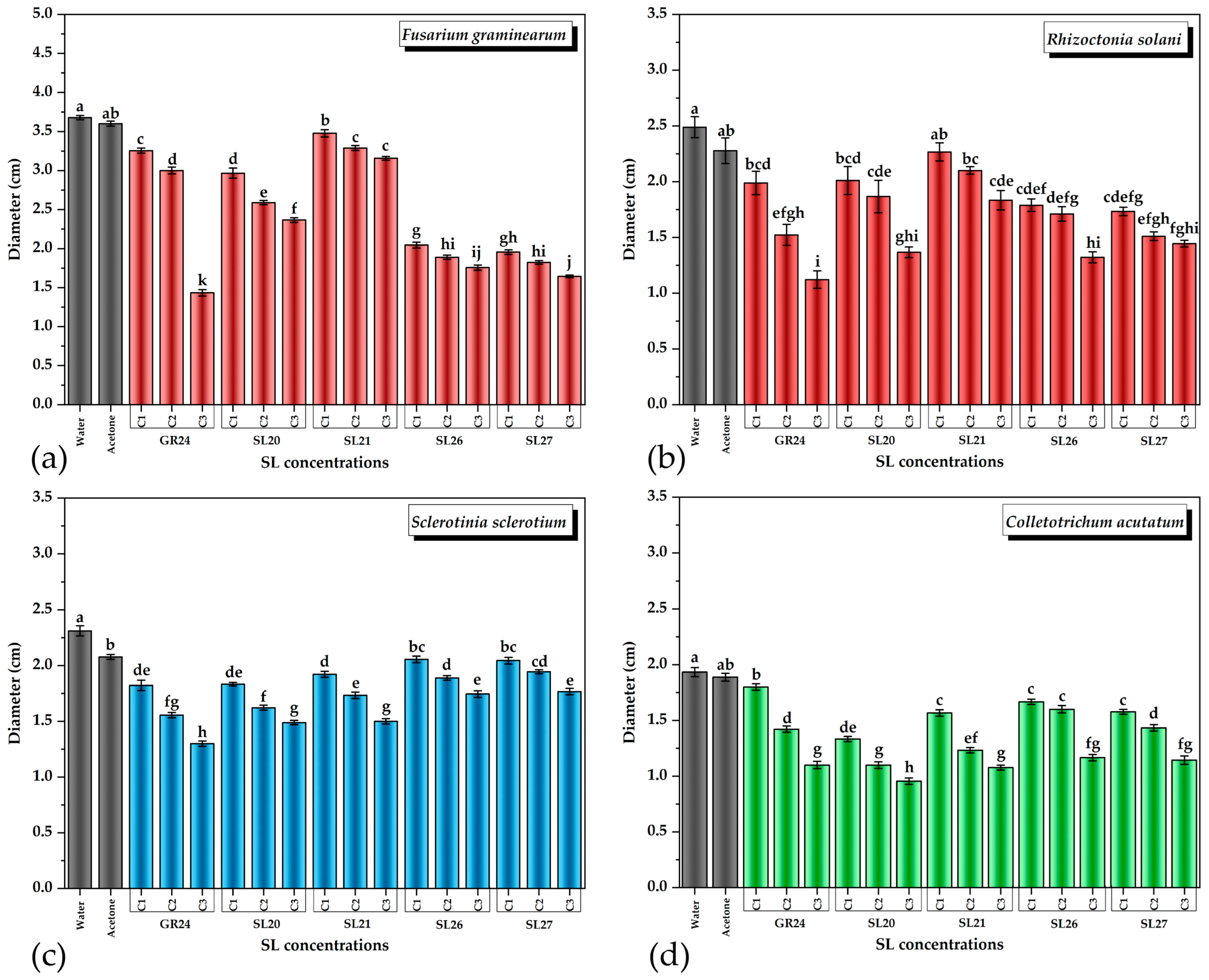
Based on the experimental results, it was observed that both GR24 and the new SL mimics had a statistically significant inhibitory effect on the growth of all four phytopathogenic fungi that were tested (Figure 5).
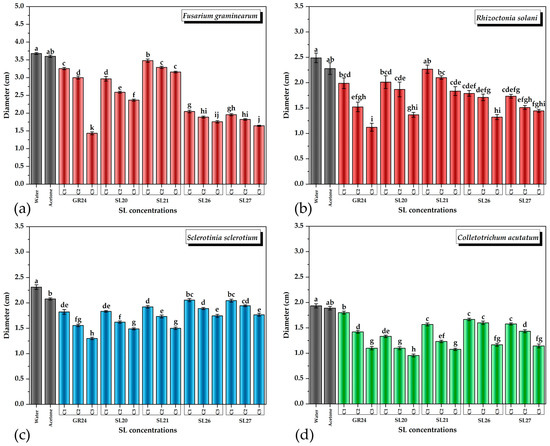
Figure 5.
The effect of SL mimics on the growth of phytopathogens expressed as colony diameter on agar plates: (a) Fusarium graminearum DSM 4527; (b) Rhizoctonia solani DSM 22842; (c) Sclerotinia sclerotium DSM 1946; (d) Colletotrichum acutatum CBS 113008; C1 = 5 × 10−6 M; C2 = 10−5 M; C3 = 5 × 10−5 M. The bars represent the standard error. Significant differences are shown by different letters at p < 0.05 (n = 3).
As can be observed in Figure 5, even at the lowest concentration tested of 5 × 10−6 M (C1), the inhibitory effect was noticeable. The inhibitory effects on fungal colonies were dependent on both the fungal strain and the SL mimic tested. The compounds showed a dose-dependent suppression of the fungal strains. All SL mimics inhibited F. graminearum compared with the acetone control. Except SL21, all the other SL mimics had a stronger inhibitory effect than GR24, with SL27 being the strongest inhibitor. In the case of R. solani, all SL mimics inhibited the fungal growth except SL21 at C1. The inhibition by the SL mimics was comparable to GR24, except SL21, which had a lower effect. The effects of SL20 and SL21 on S. sclerotium were similar to those of GR24, but SL26 and SL27 had slightly lower effects than GR24. In the case of C. acutatum, all SL mimics had a similar or even higher inhibitory effect than GR24 on fungal growth.
Taken together, all the newly synthesized SL mimics had a similar or even higher effect on the phytopathogens tested, especially at the lowest compound concentration applied. The effects depend on both the strain and the SL mimic. The SL mimics differ by the chemical group in the C-6 position of the naphthalimide ring, and it is intriguing that this can influence the effects observed, in a strain-dependent manner. Future work should aim to bring light to the mechanistic aspects.
We next investigated the effects of the SL mimics on the hyphal branching of phytopathogens, in comparison with GR24. Figures S17–S20 show the typical hyphae development of F. graminearum, R. solani, S. sclerotium, and C. acutatum, respectively, from our experimental treatments. In all cases, the number of branched hyphae was recorded along the length of 2000 µm of the primary hyphae, starting from the end of the youngest hyphal edge. Since many primary hyphae were not straight, in these cases, the total distance consisted of several shorter segments along the length of the linear hyphae.
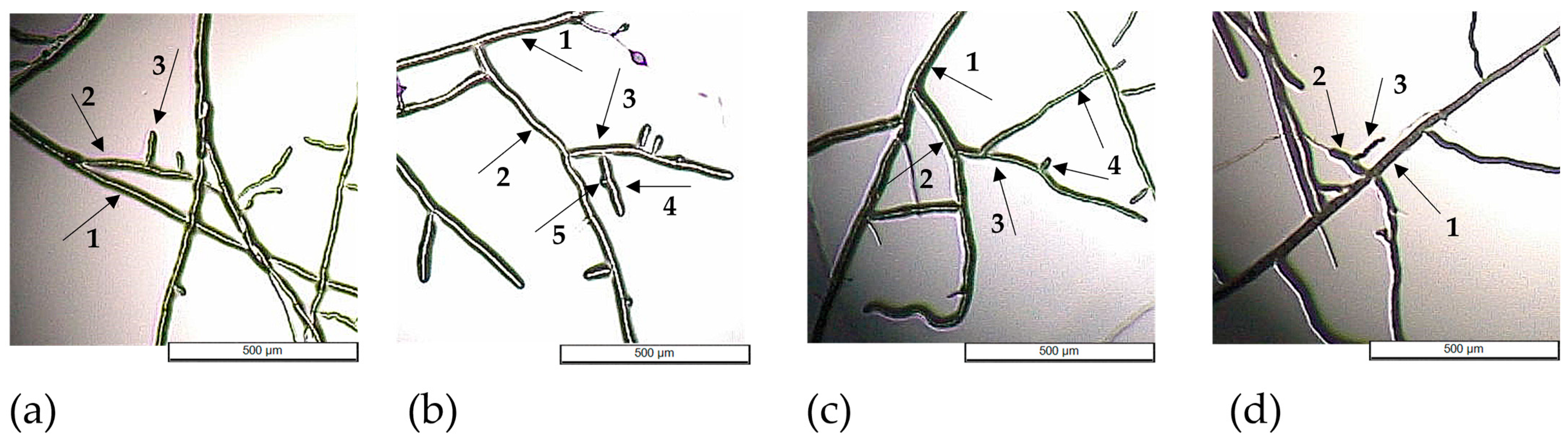
In Figure 6, the pattern of hyphal branching and analysis for each of the four fungi show hyphal branching up to the 5th order, but most of the variants displayed maximum 4th-order branching.
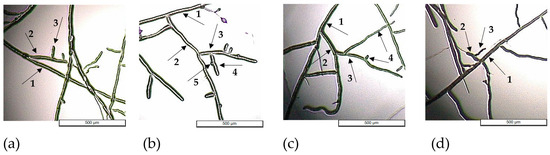
Figure 6.
Hyphal branching of Fusarium graminearum DSM 4527 (a), Rhizoctonia solani DSM 22842 (b), Sclerotinia sclerotium DSM 1946 (c), and Colletotrichum acutatum CBS 113008 (d) exposed to GR24 at concentration C3 (5 × 10−5 M); the arrows indicate the following: 1—primary branch; 2—2nd-order branch; 3—3rd-order branch; 4—4th-order branch; 5—5th-order branch.
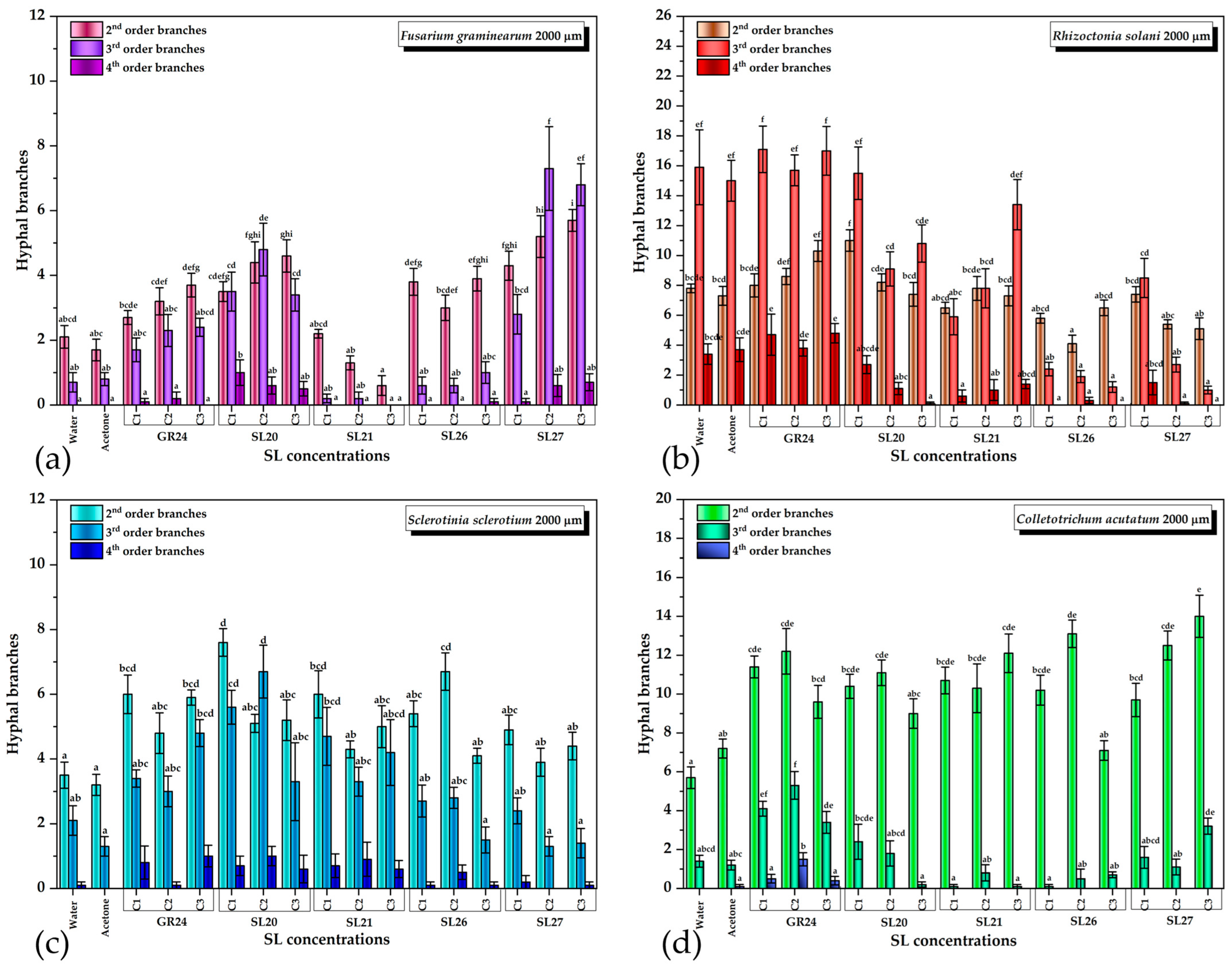
The quantification of each type of branch for all four phytopathogens and variants is shown in Figure 7. For F. graminearum, the vast majority of the treatments with SL mimics were significantly different from the control in the case of the second-order branches except SL21 at the lowest concentration C1. Most treatments induced a higher number of second-order branches, except SL21 C2 and C3, which slightly inhibited the number of branches compared with the control. GR24, SL20, and SL27 stimulated the third-order branches, whereas SL21 inhibited and SL26 slightly stimulated them only at the highest concentration C3. In the case of the fourth-order branches, only SL20 and SL27 induced a statistically significant appearance compared with the control (Figure 7a).
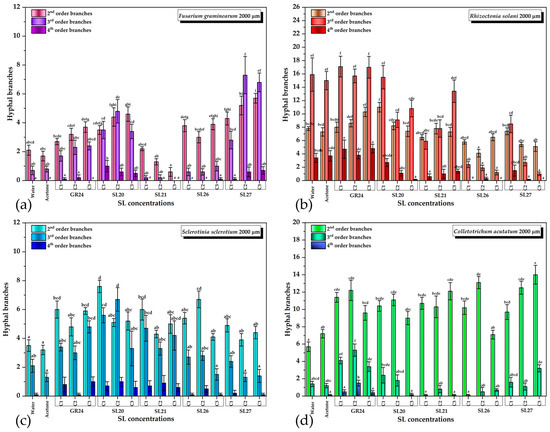
Figure 7.
The effect of GR24 and SL mimics on the number of hyphal branches of phytopathogens: (a) Fusarium graminearum DSM 4527; (b) Rhizoctonia solani DSM 22842; (c) Sclerotinia sclerotium DSM 1946; (d) Colletotrichum acutatum CBS 113008; C1 = 5×10−6 M; C2 = 10−5 M; C3 = 5 × 10−5 M. The bars represent the standard error. Significant differences are shown by different letters at p < 0.05 (n = 30).
It is worth mentioning that the dependence on the SL dose was specific for each SL mimic: whereas the GR24 and SL27 stimulatory and SL21 inhibitory effects increased with the concentration (SL27 showed signs of saturation at C2 and C3), SL26 did not show a concentration dependence. The most intriguing dependence was that of SL20, which had a heterogeneous behavior: the 2nd-order branching increased with the concentration, the 3rd-order branching had a maximum at C2, and the 4th-order branching decreased with the concentration (Figure 7a). This suggests that, in the case of SL20, the higher the branching order, the lower the optimum SL mimic concentration for maximum branching that is needed. This aspect, as well as the inhibitory effect observed for SL21, deserves further investigation.
For R. solani, the second-order branching was stimulated only by GR24 in a dose-dependent manner and by SL20 at the lowest concentration C1, after which the C2 and C3 concentrations had control-like effects. SL21 did not have a significant effect, whereas SL26 and SL27 slightly inhibited the second-order branching. The 3rd- and 4th-order branching was stimulated only by GR24. SL20 at C1 had no effect and all the other variants inhibited the third-order branching. All SL mimics inhibited the fourth-order branching. We underline the observation that R. solani had a significant number of 3rd- and 4th-order branching in the control, much higher than the other strains (Figure 6b). It is not clear whether the inhibitory effects of the SL mimics are related to this aspect, and this needs more in-depth studies.
Almost all SL variants stimulated the 2nd- and 3rd-order branching of S. sclerotium. SL26 C3 and SL27 C2 and C3 did not have a stimulatory effect on the third-order branching. In general, it was observed that lower SL concentrations had a slightly higher branching effect, with the SL20 effect being higher than the GR24 effect. The fourth-order branching was absent in the control with acetone and appeared when GR24 or SL mimics were applied, but the difference was not statistically significant (Figure 6c).
For C. acutatum, all treatments, except SL26 C3, induced significantly higher second-order branching than the control. In the case of the third-order branches, only the treatments with GR24, SL20 C1, C2, and SL27 C3 had a stimulatory effect. The other treatments inhibited or, in a few cases, did not have any effect. The fourth-order branches were slightly stimulated only by GR24 (Figure 6d).
As in the case of fungal growth inhibition, the effects on the hyphal branches is strain- and SL-mimic-dependent. Moreover, whereas the concentration dependence of fungal growth inhibition is homogeneous, the number of hyphal branches varies in a hormetic manner and the effects depend on the hyphal order as well. The observation that the chemical group in the C-6 position of the naphthalimide ring influences this pattern is in accordance with the relatively recent paradigm shift by which it was proposed that the signal transduction induced by the SL receptor is triggered by the conformational changes in the complex with the intact SL, and not by the butenolide molecule released upon hydrolysis [89]. This variation in structure could induce a variation in the receptor–SL mimic affinity as well as variations in the conformational changes, which could explain the heterogeneity in the biological outcome observed. Recently, a fungal homolog from Cryphonectria parasitica, of the plant DWARF14 (D14) SL receptor, was reported and characterized [90]. This receptor is able to hydrolyze GR24 and this enzymatic activity requires the known catalytic triad Ser/His/Asp. The knockout of this gene was reported to reduce the fungus response to SLs. We believe that the specificity of the specie–SL mimic pair is probably based on the differences in the binding site of the fungal receptor among species. Each SL will have to be taken separately and investigated in-depth in order to understand the differences observed and optimize the desired response, which we plan to do in the future. From the analysis and comparison of all data, the most promising candidate for future fluorescence studies seems to be SL20: it behaves similarly to GR24 for all the strains tested and it has the highest fluorescence quantum yield and a reasonable lifetime even in the more polar solvent, DMF, important for biological imaging applications where a polar environment is involved.
3. Materials and Methods
3.1. Materials
The starting materials 4-chloro-1,8-naphthalic anhydride 4 and 4-bromo-1,8-naphthalic anhydride 8 were commercial products (Sigma-Aldrich, St. Louis, MO, USA). The intermediate compounds, 6-chloro-2-hydroxy-benzo[de]isoquinoline-1,3-dione 5 [65] and 6-bromo-2-hydroxy-benzo[de]isoquinoline-1,3-dione 9, respectively [69], were prepared according to the reported procedures. The common intermediate, 5-bromo-3-methyl-5H-furan-2-one 6, was obtained in good yield by the bromination of 3-methyl-5H-furan-2-one with N-bromosuccinimide in CCl4 in the presence of small amounts of benzoyl peroxide or azobisisobutyronitrile [69]. All reagents and solvents, including the solvents of spectrophotometric degree utilized in this work, were commercial products (Sigma-Aldrich, St. Louis, MO, USA) and were used without purification. The fungal strains tested were represented by C. acutatum CBS 113008 (Culture collection of fungi and yeasts, Westerdijk Fungal Biodiversity Institute, Utrecht, The Netherland), F. graminearum DSM 4527, R. solani DSM 22842, and S. sclerotium DSM 1946 (DSMZ, Braunschweig, Germany). For SL mimic solubilization and strain culture, the following commercial reagents were used: GR24 racemic (Strigolab, Turin, Italy), acetone (Chimreactiv, Bucharest, Romania), potato dextrose agar, and agar (Scharlau, Barcelona, Spain).
3.2. Structural and Physical–Chemical Characterization of the Synthesized Compounds
A Boetus apparatus was used to determine the melting points of the synthetic strigolactone mimics. These melting points are uncorrected. For recording the FTIR spectra in KBr pellets, the Nicolet Impact 410 spectrometer (Thermo Scientific, Waltham, MA, USA) was used. We used a Bruker Avance Neo spectrometer for the NMR analyses (Bruker Biospin, Ettlingen, Germany), operating at 600.1, 150.9, and 60.8 MHz for 1H, 13C, and 15N, respectively. The 1D and 2D spectra were recorded on a 5 mm multinuclear inverse detection z-gradient probe. Chemical shifts are reported in δ units (ppm) and reference the residual solvent signals (1H at 2.51 ppm and 13C at 39.4 ppm). The 15N chemical shifts reference liquid ammonia (0.0 ppm) using nitromethane (380.2 ppm) as the external standard. The 2D NMR homo- and heteronuclear correlations were used to make the unambiguous 1D NMR signal assignments. H,H COSY, H,C HSQC, and H,C HMBC experiments were recorded using standard pulse sequences in the version with z-gradients, as delivered by Bruker with TopSpin 4.0.8 spectrometer control and processing software. The 15N chemical shifts were obtained as projections from the 2D indirectly detected H,N HMBC spectra, employing a standard pulse sequence in the version with z-gradients as delivered by Bruker (TopSpin 4.0.8).
High-resolution MS spectra recorded on a Bruker Maxis II QTOF spectrometer (Bruker Daltonics, Bremen, Germany), with electrospray ionization (ESI) in the positive mode, were used to obtain the exact molecular weights and isotopic patterns.
Electronic absorption spectra were performed with a Specord 210PLUS UV-Vis spectrometer (Analytik Jena, Jena, Germany) in 10 mm path length cuvettes.
The fluorescence spectra were recorded on a Perkin Elmer LS55 luminescence spectrometer (Perkin Elmer, Inc., Waltham, MA, USA) using quartz cells of 10 mm optical path length. Time-correlated single-photon-counting measurements were performed using an FLS980 fluorospectrometer (Edinburgh Instruments, Livingston, UK), with 10 mm quartz cuvettes. A nanosecond diode laser centered at 375 nm was used as the excitation source. The fluorescence quantum yield (Φ) was determined in dilute solutions (A < 0.1) at the excitation wavelength corresponding to the maximum of the absorption band. Fluorescence decay measurements were carried out at the emission maxima. The decay data were fitted to the multiexponential model (1) to obtain the fluorescence lifetimes. In Equation (2), I(t) denotes the emission intensity at a time t, and ai and τi represent the pre-exponential factor and the decay time of component i, respectively. The decay times and pre-exponential factors were estimated by minimizing the reduced chi-square χ2 value and by the plots of the weighted residuals versus channel number. All curve fittings possess χ2 values below 1.1 and a symmetrical distribution of the residuals.
3.3. Synthesis of 6-Chloro-2-(4-methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-benzo[de]isoquinoline-1,3-dione (7) SL-26 or 6-Bromo-2-(4-methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-benzo[de]isoquinoline-1,3-dione (10) SL-27
An amount of 15 mL of DMF K2CO3 (2.76 g, 20 mmol) was added to a solution of 6-chloro-2-hydroxy-benzo[de]isoquinoline-1,3-dione 5 (2.475 g, 10 mmol) or 6-bromo-2-hydroxy-benzo[de]isoquinoline-1,3-dione 9 (2.921, 10 mmol), followed by the slow addition of crude 5-bromo-3-methyl-5H-furan-2-one 6 (2.21 g, 12.5 mmol) under stirring at room temperature. The reaction mixture was stirred at room temperature for 24 h. Next, the reaction mixture was poured into 100 mL of water and extracted with 3 × 100 mL of CHCl3; the combined extracts were washed with an equal volume of water and dried on anhydrous Na2SO4. The solvent was partly removed under reduced pressure and the solid formed was filtered and recrystallized.
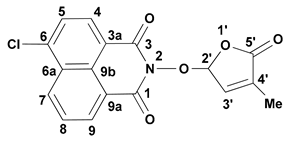
- 6-Chloro-2-(4-methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-benzo[de]isoquinoline-1,3-dione (7) SL-26. Light beige crystals, m.p. 215–217 °C (CHCl3/MeOH). Yield 67% (2.3 g). Anal. Calcd. for C17H10ClNO5 (343.73): C, 59.40; H, 2.93; N, 4.07%. Found: C, 58.57; H, 3.02; N, 3.95%. FTIR (KBr, νmax): 2922, 1777, 1727, 1684, 1589, 1367, 1234, 1192, 1090, 1029 cm−1. 1H NMR (600 MHz, DMSO-d6), δ (ppm): 1.93 (t, J = 1.4 Hz, 3H, CH3), 6.62 (quintet, J = 1.4 Hz, 1H, H-2′), 7.45 (quintet, J = 1.4 Hz, 1H, H-3′), 8.05 (dd, J = 8.3, 7.4 Hz, 1H, H-8), 8.08 (d, J = 7.9 Hz, 1H, H-5), 8.45 (d, J = 7.9 Hz, 1H, H-4), 8.63 (dd, J = 7.3, 1.0 Hz, 1H, H-9), 8.66 (dd, J = 8.4, 1.0 Hz, 1H, H-7). 13C NMR (150.9 MHz, DMSO-d6), δ (ppm): 10.2 (CH3), 104.0 (CH-2′), 121.8 (C-3a), 123.1 (C-9a), 127.9 (CH-5 and C-9b), 128.6 (C-6a), 128.8 (CH-8), 130.7 (CH-7), 131.2 (CH-4), 132.0 (CH-9), 134.3 (C-4′), 138.1 (C-6), 141.7 (CH-3′), 159.7 (CO-3), 159.9 (CO-1), 170.9 (CO-5′). 15N NMR (60.8 MHz, DMSO-d6), δ (ppm): 223.9 (N-2). HRMS-ESI (m/z): [M + Na]+ for C17H10ClNNaO5, calcd. 366.0140, found 366.0150.
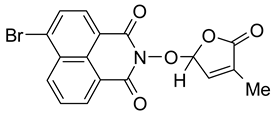
- 6-Bromo-2-(4-methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-benzo[de]isoquinoline-1,3-dione (10). SL-27. Light beige crystals, m.p. 238–241 °C (MeNO2). Yield 65% (2.5 g). Anal. Calcd. for C17H10BrNO5 (388.18): C, 52.60; H, 2.60; N, 3.61%. Found: C, 51.89; H, 2.70; N, 3.51%. ATR-FTIR (solid, νmax): 1775, 1721, 1679, 1579, 1359, 1336, 1229, 1094, 1028 cm−1. 1H NMR (600 MHz, DMSO-d6), δ (ppm): 1.93 (bs, 3H, CH3), 6.62 (bs, 1H, H-2′), 7.48 (bs, 1H, H-3′), 8.03 (t, J = 7.8 Hz, 1H, H-8), 8.26 (d, J = 7.8 Hz, 1H, H-5), 8.37 (d, J = 7.8 Hz, 1H, H-4), 8.59 (d, J = 8.5, 1H, H-9), 8.62 (d, J = 7.2 Hz, 1H, H-7). 13C NMR (150.9 MHz, DMSO-d6), δ (ppm): 10.2 (CH3), 104.1 (CH-2′), 122.4 (C-3a), 123.1 (C-9a), 127.8 (C-9b), 129.0 (CH-8), 129.9 (C-6), 130.0 (C-6a), 131.4 (CH-4), 131.5 (CH-5), 132.0 (CH-7), 133.3 (CH-9), 134.3 (C-4′), 141.8 (CH-3′), 159.9 (CO-3), 160.0 (CO-1), 170.9 (CO-5′). 15N NMR (60.8 MHz, DMSO-d6), δ (ppm): 223.0 (N-2). HRMS-ESI (m/z): [M + Na]+ for C17H10BrNO5, calcd. 409.9640, found 409.9655.
3.4. New Synthetic Procedure for SL Mimics 2 and 3
An amount of 30 mL of N-methylpyrrolidone, 4-methylpiperidine, and 4-benzylpyperidine (12 mmol), respectively, or 2.1 mL (15 mmol) of triethylamine were added to a suspension of SL mimic 7 (2.06 g, 6 mmol) or SL mimic 10 (2.33 g, 6 mmol), and the reaction mixture was heated at reflux temperature for 24 h. The reaction mixture was poured in 100 mL of water and extracted with 3 × 150 mL of CHCl3, and the combined extracts were washed with an equal volume of water and dried on anhydrous Na2SO4. The solvent was distilled off, and the solid was filtered and crystallized to obtain SL mimics 2 and 3, respectively.
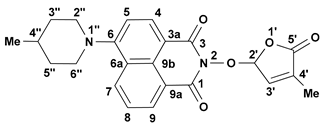
- 2-(4-Methyl-5-oxo-2,5-dihydrofuran-2-yloxy)-6-(4-methylpiperidin-1-yl)-benzo[de]isoquinoline-1,3-dione (2): SL-20. Orange crystals, m.p. 198–200 °C (CHCl3). Yield: 60% (1.45 g). Anal. Calcd. for C23H22N2O5 (406.44): C, 67.97; H, 5.46; N, 6.89%. Found: C, 68.10; H, 5.54; N, 5.93%. FTIR (KBr, νmax): 3432, 2916, 1784, 1715, 1676, 1584, 1457, 1363, 1229, 1193, 1088 cm−1. 1H NMR (600 MHz, DMSO-d6), δ (ppm): 1.04 (d, J = 6.4 Hz, 3H, CH3-4″), 1.52 (dd, J = 22.6, 11.2 Hz, 1H, CH2-3″A), 1.63–1.65 (m, 1H, CH-4″), 1.82 (d, J = 11.9 Hz, 1H, CH2-3″B), 1.92 (bs, 3H, CH3-4′), 2.93 (t, J = 11.9 Hz, 1H, CH2-2″A), 3.56 (d, J = 11.9 Hz, 1H, CH2-2″B), 6.60 (bs, 1H, H-2′), 7.34 (d, J = 8.2 Hz, 1H, H-5), 7.47 (bs, 1H, H-3′), 7.83 (t, J = 7.8 Hz, 1H, H-8), 8.40 (d, J = 8.1 Hz, 1H, H-4), 8.44 (d, J = 8.4 Hz, 1H, H-7), 8.50 (d, J = 7.1 Hz, 1H, H-9). 13C NMR (150.9 MHz, DMSO-d6), δ (ppm): 10.2 (CH3-4′), 21.7 (CH3-4″), 30.2 (CH-4″), 33.9 (CH2-3″), 53.2 and 53.3 (CH2-2″ and CH2-6″), 104.0 (CH-2′), 114.6 (C-3a), 115.1 (CH-5), 122.7 (C-9a), 125.5 (C-6a), 125.9 (CH-8), 128.7 (C-9b), 131.1 (CH-9), 131.4 (CH-7), 132.8 (CH-4), 134.2 (C-4′), 141.9 (CH-3′), 157.2 (C-6), 159.9 (CO-3), 160.4 (CO-1), 171.0 (CO-5′). 15N NMR (60.8 MHz, DMSO-d6), δ (ppm): 73.6 (N-1″), 223.6 (N-2). HRMS-ESI (m/z): [M + Na]+ for C23H22N2NaO5, calcd. 429.1421, found 429.1425 [67].
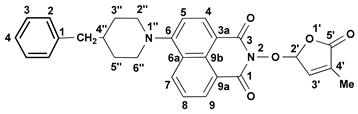
- 2-(4-Methyl-5-oxo-2,5-dihydro-furan-2-yloxy)-6-(4-benzyl-piperidin-1-yl)-benzo[de]isoquinoline-1,3-dione (3): SL-21. Orange crystals, m.p. 182–184 °C (CHCl3). Yield: 58% (1.67 g). Anal. Calcd. for C29H26N2O5 (482.54): C, 72.19; H, 5.43; N, 5.81%. Found: C, 72.06; H, 5.33; N, 5.93%. FTIR (KBr, νmax): 2911, 2794, 1784, 1718, 1684, 1584, 1452, 1364, 1235, 1177, 1026 cm−1. 1H NMR (600 MHz, DMSO-d6), δ (ppm): 1.58 (dd, J = 22.6, 11.2 Hz, 1H, CH2-3″A), 1.77 (d, J = 11.3 Hz, 2H, CH2-3″B and CH-4″), 1.91 (bs, 3H, CH3-4′), 2.65 (d, J = 6.5 Hz, 2H, CH2-4″), 2.87 (t, J = 12.1 Hz, 1H, CH2-2″A), 3.56 (d, J = 11.6 Hz, 1H, CH2-2″B), 6.60 (bs, 1H, H-2′), 7.21 (t, J = 7.2 Hz, 1H, H-4Ph), 7.24 (d, J = 7.3 Hz, 2H, H-2Ph), 7.31 (d, J = 7.2 Hz, 1H, H-5), 7.32 (t, J = 8.0 Hz, 2H, H-3Ph), 7.46 (bs, 1H, H-3′), 7.82 (t, J = 7.9 Hz, 1H, H-8), 8.39 (d, J = 8.2 Hz, 1H, H-4), 8.42 (d, J = 8.3 Hz, 1H, H-7), 8.49 (d, J = 7.2 Hz, 1H, H-9). 13C NMR (150.9 MHz, DMSO-d6), δ (ppm): 10.3 (CH3-4′), 31.8 (CH2-3″), 37.3 (CH-4″), 42.3 (CH2-4″), 53.2 and 53.3 (CH2-2″ and CH2-6″), 104.0 (CH-2′), 114.7 (C-3a), 115.1 (CH-5), 122.7 (C-9a), 125.5 (C-6a), 125.9 (CH-4Ph), 126.0 (CH-8), 128.2 (CH-3Ph), 128.7 (C-9b), 129.1 (CH-2Ph), 131.2 (CH-9), 131.4 (CH-7), 132.8 (CH-4), 134.2 (C-4′), 140.2 (C-1Ph), 141.9 (CH-3′), 157.1 (C-6), 160.0 (CO-3), 160.5 (CO-1), 171.1 (CO-5′). 15N NMR (60.8 MHz, DMSO-d6), δ (ppm): 74.2 (N-1″), 223.4 (N-2). HRMS-ESI (m/z): [M + Na]+ for C29H26N2NaO5, calcd. 505.1734, found 505.1738 [67].
3.5. Fungal Strain Cultivation and Experimental Design
The fungal strains were first inoculated and grown on potato dextrose agar (PDA) medium at 28 °C for 5 days. Stock solutions of SL mimics at a concentration of 10 mM were prepared by dissolving 1.5 mg of GR24, 2 mg of SL20, 2.5 mg of SL21, 3.43 mg of SL26, and 3.88 mg of SL27 in 500 µL of acetone. GR24 and SL mimics were tested by dissolving them in water agar 1.8%, with the final medium being poured into Petri dishes. The final concentrations of GR24 and SL mimics dissolved into water agar were 5 × 10−6 M (C1), 10−5 M (C2), and 5 × 10−5 M (C3). The acetone concentration in the final solution was 0.5% (v/v). Water agar of 1.8% and water agar embedded with 0.5% (v/v) acetone were used as controls, with GR24 as a chemical reference structure. The inoculated Petri dishes were kept for 3 days at 28 °C in an incubator MIR-154-PE (PHCBi Panasonic, Osaka, Japan). The developed colonies were macroscopically investigated by measuring the diameter. The images were collected on day 3 of shooting by examination in brightfield mode using a Leica DM 1000 LED microscope (Leica Microsystems, Wetzlar, Germany) equipped with an ICC50W digital camera. The number of hyphal branches of different orders was determined on each primary branch, from the second to the maximum, starting from the apical end of the youngest hyphae. The selected distance for hyphae counting was 2000 µm, where the maximum hyphae order could be observed for all variants. The observations focused on the number and the arrangement of the hyphae. Statistical analysis was performed using IBM® SPSS® Statistics version 26 (IBM, Armonk, NY, USA). For the statistical analysis, ten hyphae from three replicates for each variant were used. One-way ANOVA was used to determine whether there are significant differences between test groups.
4. Conclusions
Two new affordable bioactive SL mimics 7 and 10 containing a halo-substituted 1,8-naphthalimide ring connected through an ether link to a bioactive furan-2-one unit have been obtained and fully characterized. They have been further used in alternative synthetic routes towards the previously reported SL mimics 2 and 3. All investigated compounds (2, 3, 7, and 10) present fluorescence properties and interesting biological activity. Particularly, SL mimics 2 and 3, bearing piperidine-substituents on the 1,8-naphthalimide ring, exhibit emission maxima in DCM at 522 nm and 520 nm, respectively; high Stokes shifts; and convenient quantum fluorescence yields. SL mimics 2, 3, 7, and 10 showed biological activity similar to GR24 on selected phytopathogen strains. The specific behavior depends on the strain, SL mimic characteristics, and concentration, as well as on hyphal order. These characteristics make them, SL mimic 2 (SL20) in particular, convenient for sustainable agriculture and bio-imaging applications. The specific behavior depends on the strain, SL mimic characteristics, and concentration, as well as hyphal order. The fluorescence after the hydrolysis of the bioactive D-ring is preserved. Therefore, it could be further used to investigate the temporal distribution of the fluorescent bioactiphore probe in the plant or fungal cell.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/molecules29102283/s1. Figure S1. 1H NMR spectrum corresponding to SL mimic 7, recorded in DMSO-d6, at 600 MHz. Figure S2. 13C NMR spectrum corresponding to SL mimic 7, recorded in DMSO-d6, at 150 MHz. Figure S3. H,C HSQC spectrum corresponding to SL mimic 7 (SL26), recorded in DMSO-d6, at 600 MHz. Figure S4. H,C HMBC spectrum corresponding to SL mimic 7, recorded in DMSO-d6, at 600 MHz. Figure S5. 1H NMR spectrum corresponding to SL mimic 10, recorded in DMSO-d6, at 600 MHz. Figure S6. 13C NMR spectrum corresponding to SL mimic 10, recorded in DMSO-d6, at 150 MHz. Figure S7. H,N HMBC spectrum corresponding to SL mimic 10, recorded in DMSO-d6, at 60.8 MHz. Figure S8. H,H COSY spectrum corresponding to SL mimic 10, recorded in DMSO-d6, at 600 MHz. Figure S9. H,C HSQC spectrum corresponding to SL mimic 10, recorded in DMSO-d6, at 600 MHz. Figure S10. H,C HMBC spectrum corresponding to SL mimic 10, recorded in DMSO-d6, at 600 MHz. Figure S11. HRMS-ESI spectrum with isotopic pattern in positive mode for SL mimic 7. Upper trace (red)—simulated pattern; lower trace (black)—experimental spectrum for C17H10BrNO5 [M + Na]+. HRMS-ESI (m/z): [M + Na]+ for C17H10ClNO5, calcd. 366.0145, found 366.0150. Figure S12. HRMS-ESI spectrum with isotopic pattern in positive mode for SL mimic 10. Upper trace (red)—simulated pattern; lower trace (black)—experimental spectrum for C17H10BrNO5 [M + Na]+. HRMS-ESI (m/z): [M + Na]+ for C17H10BrNO5, calcd. 409.9640, found 409.9655. Figure S13. Absorption spectra of SL mimics 1, 7, and 10 in dichloromethane. Figure S14. Emission spectra of SL mimics 1, 7, and 10 in dichloromethane. Figure S15. The fluorescence decay traces of SL mimics 1 and 7 in toluene. Figure S16. The fluorescence decay traces of SL mimics 1, 7, and 10 in DMF. Figure S17. The aspect of hyphal branching of Fusarium graminearum in (a) control water agar, (b) acetone, (c) GR24 C1, (d) GR24 C2, (e) GR24 C3, (f) SL20 C1, (g) SL20 C2, (h) SL20 C3, (i) SL21 C1, (j) SL21 C2, (k) SL21 C3, (l) SL26 C1, (m) SL26 C2, (n) SL26 C3, (o) SL27 C1, (p) SL27 C2, and (q) SL27 C3 recorded on 2000 µm beginning from the end of the youngest hyphal edge. Figure S18. The aspect of hyphal branching of Rhizoctonia solani in (a) control water agar, (b) acetone, (c) GR24 C1, (d) GR24 C2, (e) GR24 C3, (f) SL20 C1, (g) SL20 C2, (h) SL20 C3, (i) SL21 C1, (j) SL21 C2, (k) SL21 C3, (l) SL26 C1, (m) SL26 C2, (n) SL26 C3, (o) SL27 C1, (p) SL27 C2, and (q) SL27 C3 recorded on 2000 µm beginning from the end of the youngest hyphal edge. Figure S19. The aspect of hyphal branching of Sclerotinia sclerotium in (a) control water agar, (b) acetone, (c) GR24 C1, (d) GR24 C2, (e) GR24 C3, (f) SL20 C1, (g) SL20 C2, (h) SL20 C3, (i) SL21 C1, (j) SL21 C2, (k) SL21 C3, (l) SL26 C1, (m) SL26 C2, (n) SL26 C3, (o) SL27 C1, (p) SL27 C2, and (q) SL27 C3 recorded on 2000 µm beginning from the end of the youngest hyphal edge. Figure S20. The aspect of hyphal branching of Colletotrichum acutatum in (a) control water agar, (b) acetone, (c) GR24 C1, (d) GR24 C2, (e) GR24 C3, (f) SL20 C1, (g) SL20 C2, (h) SL20 C3, (i) SL21 C1, (j) SL21 C2, (k) SL21 C3, (l) SL26 C1, (m) SL26 C2, (n) SL26 C3, (o) SL27 C1, (p) SL27 C2, and (q) SL27 C3 recorded on 2000 µm beginning from the end of the youngest hyphal edge. Table S1. Spectral and photophysical characteristics of the SL mimics 1, 2 and 3 measured in different organic solvents. Table S2. Fluorescence lifetimes of the SL mimics 1, 2 and 3.
Author Contributions
Conceptualization, F.O. and F.G.; methodology, F.G., F.D. and E.G.; validation, F.G., D.C.-A. and C.D.; formal analysis, I.-A.B. and A.N.; investigation, I.-A.B., A.N., F.G., F.D., E.G., A.A. and R.T.; resources, F.O. and E.G.; data curation, F.G., D.C.-A. and C.D.; writing—original draft preparation, I.-A.B., A.N., F.G., E.G. and A.A.; writing—review and editing, D.C.-A., F.O. and C.D.; visualization, D.C.-A. and F.O.; supervision, F.O. and C.D.; project administration, F.O.; funding acquisition, F.O. All authors have read and agreed to the published version of the manuscript.
Funding
The research leading to these results has received funding from NO Grants 2014–2021, under Project RO-NO-2019-540, contract no. 14/2020.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are included in the main manuscript and Supplementary Materials.
Conflicts of Interest
Author Florentina Georgescu was employed by the company Enpro Soctech Com. Author Emilian Georgescu was employed by the company Research Center Oltchim. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Waters, M.T.; Gutjahr, C.; Bennett, T.; Nelson, D.C. Strigolactone Signaling and Evolution. Annu. Rev. Plant Biol. 2017, 68, 291–322. [Google Scholar] [CrossRef] [PubMed]
- Burger, M.; Chory, J. The Many Models of Strigolactone Signaling. Trends Plant Sci. 2020, 25, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Smith, S.M.; Huang, J. Origins of strigolactone and karrikin signaling in plants. Trends Plant Sci. 2022, 27, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Proust, H.; Hoffmann, B.; Xie, X.; Yoneyama, K.; Schaefer, D.G.; Yoneyama, K.; Nogué, F.; Rameau, C. Strigolactones regulate protonema branching and act as a quorum sensing-like signal in the moss Physcomitrella patens. Development 2011, 138, 1531–1539. [Google Scholar] [CrossRef]
- Wheeldon, C.D.; Hamon-Josse, M.; Lund, H.; Yoneyama, K.; Bennett, T. Environmental strigolactone drives early growth responses to neighboring plants and soil volume in pea. Curr. Biol. 2022, 32, 3593–3600.e3. [Google Scholar] [CrossRef]
- Akiyama, K.; Matsuzaki, K.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef]
- Akiyama, K.; Ogasawara, S.; Ito, S.; Hayashi, H. Structural requirements of strigolactones for hyphal branching in AM fungi. Plant Cell Physiol. 2010, 51, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.J.; Roux, C.; Lopez-Raez, J.A.; Becard, G. Rhizosphere communication of plants, parasitic plants and AM fungi. Trends Plant Sci. 2007, 12, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.J.; Fernández-Aparicio, M.; Castellanos-Morales, V.; García-Garrido, J.M.; Ocampo, J.A.; Delgado, M.J.; Vierheilig, H. First indications for the involvement of strigolactones on nodule formation in alfalfa (Medicago sativa). Soil Biol. Biochem. 2010, 42, 383–385. [Google Scholar] [CrossRef]
- Foo, E.; Davies, N.W. Strigolactones promote nodulation in pea. Planta 2011, 234, 1073–1081. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Rehman, N.U.; Yu, S.; Zhou, Y.; Haq, B.u.; Wang, J.; Li, P.; Zeng, Z.; Zhao, J. GmMAX2–D14 and–KAI interaction-mediated SL and KAR signaling play essential roles in soybean root nodulation. Plant J. 2020, 101, 334–351. [Google Scholar] [CrossRef]
- Carvalhais, L.C.; Rincon-Florez, V.A.; Brewer, P.B.; Beveridge, C.A.; Dennis, P.G.; Schenk, P.M. The ability of plants to produce strigolactones affects rhizosphere community composition of fungi but not bacteria. Rhizosphere 2019, 9, 18–26. [Google Scholar] [CrossRef]
- Liu, F.; Rice, J.H.; Lopes, V.; Grewal, P.; Lebeis, S.L.; Hewezi, T.; Staton, M.E. Overexpression of strigolactone-associated genes exerts fine-tuning selection on soybean rhizosphere bacterial and fungal microbiome. Phytobiomes J. 2020, 4, 239–251. [Google Scholar] [CrossRef]
- Kim, B.; Westerhuis, J.A.; Smilde, A.K.; Flokova, K.; Suleiman, A.K.A.; Kuramae, E.E.; Bouwmeester, H.J.; Zancarini, A. Effect of strigolactones on recruitment of the rice root-associated microbiome. FEMS Microbiol. Ecol. 2022, 98, fiac010. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.E.; Whichard, L.P.; Turner, B.; Wall, M.E.; Egley, G.H. Germination of Witchweed (Striga lutea Lour.): Isolation and Properties of a Potent Stimulant. Science 1966, 154, 1189–1190. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, K.; Xie, X.; Yoneyama, K.; Takeuchi, Y. Strigolactones: Structures and biological activities. Pest Manag. Sci. 2009, 65, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Yoneyama, K.; Yoneyama, K. The strigolactone story. Annu. Rev. Phytopathol. 2010, 48, 93–117. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Hayashi, H. Strigolactones: Chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. 2006, 97, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Dor, E.; Joel, D.M.; Kapulnik, Y.; Koltai, H.; Hershenhorn, J. The synthetic strigolactone GR24 influences the growth pattern of phytopathogenic fungi. Planta 2011, 234, 419–427. [Google Scholar] [CrossRef]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pages, V.; Dun, E.A.; Pillot, J.P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.C.; et al. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Kapulnik, Y.; Resnick, N.; Mayzlish-Gati, E.; Kaplan, Y.; Wininger, S.; Hershenhorn, J.; Koltai, H. Strigolactones interact with ethylene and auxin in regulating root-hair elongation in Arabidopsis. J. Exp. Bot. 2011, 62, 2915–2924. [Google Scholar] [CrossRef] [PubMed]
- Koltai, H. Strigolactones are regulators of root development. New Phytol. 2011, 190, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Kapulnik, Y.; Delaux, P.M.; Resnick, N.; Mayzlish-Gati, E.; Wininger, S.; Bhattacharya, C.; Sejalon-Delmas, N.; Combier, J.P.; Becard, G.; Belausov, E.; et al. Strigolactones affect lateral root formation and root-hair elongation in Arabidopsis. Planta 2011, 233, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Boyer, F.D.; de Saint Germain, A.; Pillot, J.P.; Pouvreau, J.B.; Chen, V.X.; Ramos, S.; Stevenin, A.; Simier, P.; Delavault, P.; Beau, J.M.; et al. Structure-activity relationship studies of strigolactone-related molecules for branching inhibition in garden pea: Molecule design for shoot branching. Plant Physiol. 2012, 159, 1524–1544. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, B.; Pospisil, T. Structure and activity of strigolactones: New plant hormones with a rich future. Mol. Plant 2013, 6, 38–62. [Google Scholar] [CrossRef] [PubMed]
- Boyer, F.-D.; de Saint Germain, A.; Pouvreau, J.-B.; Clavé, G.; Pillot, J.-P.; Roux, A.; Rasmussen, A.; Depuydt, S.; Lauressergues, D.; dit Frey, N.F. New strigolactone analogs as plant hormones with low activities in the rhizosphere. Mol. Plant 2014, 7, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, B.; Pospisil, T.; Cavar Zeljkovic, S. Strigolactones: New plant hormones in action. Planta 2016, 243, 1311–1326. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.J.; Fonne-Pfister, R.; Screpanti, C.; De Mesmaeker, A. Strigolactones: Plant Hormones with Promising Features. Angew. Chem. Int. Ed. Engl. 2019, 58, 12778–12786. [Google Scholar] [CrossRef]
- Rameau, C.; Goormachtig, S.; Cardinale, F.; Bennett, T.; Cubas, P. Strigolactones as plant hormones. In Strigolactones—Biology and Applications; Springer: Berlin/Heidelberg, Germany, 2019; pp. 47–87. [Google Scholar]
- Kleman, J.; Matusova, R. Strigolactones: Current research progress in the response of plants to abiotic stress. Biologia 2022, 78, 307–318. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Li, W.; Nguyen, K.H.; Fujita, M.; Tran, L.P. Strigolactones in plant adaptation to abiotic stresses: An emerging avenue of plant research. Plant Cell Environ. 2018, 41, 2227–2243. [Google Scholar] [CrossRef] [PubMed]
- Saeed, W.; Naseem, S.; Ali, Z. Strigolactones biosynthesis and their role in abiotic stress resilience in plants: A critical review. Front. Plant Sci. 2017, 8, 279971. [Google Scholar] [CrossRef] [PubMed]
- Al-Babili, S.; Bouwmeester, H.J. Strigolactones, a novel carotenoid-derived plant hormone. Annu. Rev. Plant Biol. 2015, 66, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, K.; Xie, X.; Yoneyama, K.; Kisugi, T.; Nomura, T.; Nakatani, Y.; Akiyama, K.; McErlean, C.S. Which are the major players, canonical or non-canonical strigolactones? J. Exp. Bot. 2018, 69, 2231–2239. [Google Scholar] [CrossRef] [PubMed]
- Guercio, A.M.; Palayam, M.; Shabek, N. Strigolactones: Diversity, perception, and hydrolysis. Phytochem. Rev. 2023, 22, 339–359. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, B.; Mwakaboko, A.S.; Reizelman, A.; Anilkumar, G.; Sethumadhavan, D. Structure and function of natural and synthetic signalling molecules in parasitic weed germination. Pest Manag. Sci. 2009, 65, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.W.; Gowada, G.; Hassanali, A.; Knox, J.; Monaco, S.; Razavi, Z.; Rosebery, G. The preparation of synthetic analogues of strigol. J. Chem. Soc. Perkin Trans. 1 1981, 1734–1743. [Google Scholar] [CrossRef]
- Mangnus, E.M.; Zwanenburg, B. Synthesis, structural characterization, and biological evaluation of all four enantiomers of strigol Analog GR7. J. Agric. Food Chem. 1992, 40, 697–700. [Google Scholar] [CrossRef]
- Mangnus, E.M.; Van Vliet, L.A.; Vandenput, D.A.; Zwanenburg, B. Structural modifications of strigol analogs. Influence of the B and C rings on the bioactivity of the germination stimulant GR24. J. Agric. Food Chem. 1992, 40, 1222–1229. [Google Scholar] [CrossRef]
- Mangnus, E.M.; Dommerholt, F.J.; De Jong, R.L.; Zwanenburg, B. Improved synthesis of strigol analog GR24 and evaluation of the biological activity of its diastereomers. J. Agric. Food Chem. 1992, 40, 1230–1235. [Google Scholar] [CrossRef]
- Nefkens, G.H.; Thuring, J.W.J.; Beenakkers, M.F.; Zwanenburg, B. Synthesis of a phthaloylglycine-derived strigol analogue and its germination stimulatory activity toward seeds of the parasitic weeds Striga hermonthica and Orobanche crenata. J. Agric. Food Chem. 1997, 45, 2273–2277. [Google Scholar] [CrossRef]
- Mwakaboko, A.S.; Zwanenburg, B. Single step synthesis of strigolactone analogues from cyclic keto enols, germination stimulants for seeds of parasitic weeds. Bioorg. Med. Chem. 2011, 19, 5006–5011. [Google Scholar] [CrossRef] [PubMed]
- Mwakaboko, A.S.; Zwanenburg, B. Strigolactone analogs derived from ketones using a working model for germination stimulants as a blueprint. Plant Cell Physiol. 2011, 52, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Lachia, M.; Jung, P.M.; De Mesmaeker, A. A novel approach toward the synthesis of strigolactones through intramolecular [2+2] cycloaddition of ketenes and ketene-iminiums to olefins. Application to the asymmetric synthesis of GR-24. Tetrahedron Lett. 2012, 53, 4514–4517. [Google Scholar] [CrossRef]
- Zwanenburg, B.; Mwakaboko, A.S. Strigolactone analogues and mimics derived from phthalimide, saccharine, p-tolylmalondialdehyde, benzoic and salicylic acid as scaffolds. Biorg. Med. Chem. 2011, 19, 7394–7400. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, B.; Nayak, S.K.; Charnikhova, T.V.; Bouwmeester, H.J. New strigolactone mimics: Structure–activity relationship and mode of action as germinating stimulants for parasitic weeds. Bioorg. Med. Chem. Lett. 2013, 23, 5182–5186. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, M.; Soudek, P.; Vanek, T. Triazolide Strigolactone Mimics Influence Root Development in Arabidopsis. J. Nat. Prod. 2017, 80, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, A.; Heugebaert, T.; Matthys, C.; Van Deun, R.; Boyer, F.D.; Goormachtig, S.; Stevens, C.; Geelen, D. A fluorescent alternative to the synthetic strigolactone GR24. Mol. Plant 2013, 6, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, C.; Bonfante, P.; Deagostino, A.; Kapulnik, Y.; Larini, P.; Occhiato, E.G.; Prandi, C.; Venturello, P. A new class of conjugated strigolactone analogues with fluorescent properties: synthesis and biological activity. Org. Biomol. Chem. 2009, 7, 3413–3420. [Google Scholar] [CrossRef] [PubMed]
- Fornier, S.D.; de Saint Germain, A.; Retailleau, P.; Pillot, J.-P.; Taulera, Q.; Andna, L.; Miesch, L.; Rochange, S.; Pouvreau, J.-B.; Boyer, F.-D. Noncanonical Strigolactone Analogues Highlight Selectivity for Stimulating Germination in Two Phelipanche ramosa Populations. J. Nat. Prod. 2022, 85, 1976–1992. [Google Scholar] [CrossRef]
- Fukui, K.; Ito, S.; Asami, T. Selective Mimics of Strigolactone Actions and Their Potential Use for Controlling Damage Caused by Root Parasitic Weeds. Mol. Plant 2013, 6, 88–99. [Google Scholar] [CrossRef]
- Cala, A.; Ghooray, K.; Fernández-Aparicio, M.; Molinillo, J.M.; Galindo, J.C.; Rubiales, D.; Macías, F.A. Phthalimide-derived strigolactone mimics as germinating agents for seeds of parasitic weeds. Pest Manag. Sci. 2016, 72, 2069–2081. [Google Scholar] [CrossRef] [PubMed]
- Oancea, F.; Georgescu, E.; Matusova, R.; Georgescu, F.; Nicolescu, A.; Raut, I.; Jecu, M.-L.; Vladulescu, M.-C.; Vladulescu, L.; Deleanu, C. New strigolactone mimics as exogenous signals for rhizosphere organisms. Molecules 2017, 22, 961. [Google Scholar] [CrossRef] [PubMed]
- de Saint Germain, A.; Retailleau, P.; Norsikian, S.; Servajean, V.; Pelissier, F.; Steinmetz, V.; Pillot, J.-P.; Rochange, S.; Pouvreau, J.-B.; Boyer, F.-D. Contalactone, a contaminant formed during chemical synthesis of the strigolactone reference GR24 is also a strigolactone mimic. Phytochemistry 2019, 168, 112112. [Google Scholar] [CrossRef] [PubMed]
- Dieckmann, M.C.; Dakas, P.-Y.; De Mesmaeker, A. Synthetic Access to Noncanonical Strigolactones: Syntheses of Carlactonic Acid and Methyl Carlactonoate. J. Org. Chem. 2018, 83, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, B.; Blanco-Ania, D. Strigolactones: New plant hormones in the spotlight. J. Exp. Bot. 2018, 69, 2205–2218. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-W.; Xi, Z. Strigolactone agonists/antagonists for agricultural applications: New opportunities. Adv. Agrochem. 2022, 1, 61–72. [Google Scholar] [CrossRef]
- Yao, R.; Ming, Z.; Yan, L.; Li, S.; Wang, F.; Ma, S.; Yu, C.; Yang, M.; Chen, L.; Chen, L. DWARF14 is a non-canonical hormone receptor for strigolactone. Nature 2016, 536, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Conn, C.E.; Bythell-Douglas, R.; Neumann, D.; Yoshida, S.; Whittington, B.; Westwood, J.H.; Shirasu, K.; Bond, C.S.; Dyer, K.A.; Nelson, D.C. PLANT EVOLUTION. Convergent evolution of strigolactone perception enabled host detection in parasitic plants. Science 2015, 349, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Wang, F.; Ming, Z.; Du, X.; Chen, L.; Wang, Y.; Zhang, W.; Deng, H.; Xie, D. ShHTL7 is a non-canonical receptor for strigolactones in root parasitic weeds. Cell Res. 2017, 27, 838–841. [Google Scholar] [CrossRef]
- Prandi, C.; Occhiato, E.G.; Tabasso, S.; Bonfante, P.; Novero, M.; Scarpi, D.; Bova, M.E.; Miletto, I. New potent fluorescent analogues of strigolactones: Synthesis and biological activity in parasitic weed germination and fungal branching. Eur. J. Org. Chem. 2011, 2011, 3781–3793. [Google Scholar] [CrossRef]
- Prandi, C.; Rosso, H.; Lace, B.; Occhiato, E.G.; Oppedisano, A.; Tabasso, S.; Alberto, G.; Blangetti, M. Strigolactone analogs as molecular probes in chasing the (SLs) receptor/s: Design and synthesis of fluorescent labeled molecules. Mol. Plant 2013, 6, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Goossens, H.; Heugebaert, T.S.; Dereli, B.; Van Overtveldt, M.; Karahan, O.; Dogan, I.; Waroquier, M.; Van Speybroeck, V.; Aviyente, V.; Catak, S. Elucidating the structural isomerism of fluorescent strigolactone analogue CISA-1. Eur. J. Org. Chem. 2015, 2015, 1211–1217. [Google Scholar] [CrossRef]
- Lace, B.; Prandi, C. Shaping small bioactive molecules to untangle their biological function: A focus on fluorescent plant hormones. Mol. Plant 2016, 9, 1099–1118. [Google Scholar] [CrossRef] [PubMed]
- Parisotto, S.; Lace, B.; Artuso, E.; Lombardi, C.; Deagostino, A.; Scudu, R.; Garino, C.; Medana, C.; Prandi, C. Heck functionalization of an asymmetric aza-BODIPY core: Synthesis of far-red infrared probes for bioimaging applications. Org. Biomol. Chem. 2017, 15, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Oancea, A.; Georgescu, E.; Georgescu, F.; Nicolescu, A.; Oprita, E.I.; Tudora, C.; Vladulescu, L.; Vladulescu, M.-C.; Oancea, F.; Deleanu, C. Isoxazole derivatives as new nitric oxide elicitors in plants. Beilstein J. Org. Chem. 2017, 13, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, E.; Oancea, A.; Georgescu, F.; Nicolescu, A.; Oprita, E.I.; Vladulescu, L.; Vladulescu, M.-C.; Oancea, F.; Shova, S.; Deleanu, C. Schiff bases containing a furoxan moiety as potential nitric oxide donors in plant tissues. PLoS ONE 2018, 13, e0198121. [Google Scholar] [CrossRef] [PubMed]
- Bala, I.; Trica, B.; Georgescu, F.; Georgescu, E.; Constantinescu-Aruxandei, D.; Shaposhnikov, S.; Oancea, F. The effect of a strigolactone mimic on growth and colony morphology in phytopathogenic fungi. AgroLife Sci. J. 2021, 10, 36–47. [Google Scholar] [CrossRef]
- Tariq, A.; Ullah, I.; Sardans, J.; Graciano, C.; Mussarat, S.; Ullah, A.; Zeng, F.; Wang, W.; Al-Bakre, D.A.; Ahmed, Z.; et al. Strigolactones can be a potential tool to fight environmental stresses in arid lands. Environ. Res. 2023, 229, 115966. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.; Xu, K.; Li, X.; Liu, J.; Altaf, M.A.; Fu, H.; Lu, X.; Cheng, S.; Wang, Z. Exogenous strigolactone enhanced the drought tolerance of pepper (Capsicum chinense) by mitigating oxidative damage and altering the antioxidant mechanism. Plant Cell Rep. 2024, 43, 106. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 426696. [Google Scholar] [CrossRef]
- Banerjee, S.; Veale, E.B.; Phelan, C.M.; Murphy, S.A.; Tocci, G.M.; Gillespie, L.J.; Frimannsson, D.O.; Kelly, J.M.; Gunnlaugsson, T. Recent advances in the development of 1,8-naphthalimide based DNA targeting binders, anticancer and fluorescent cellular imaging agents. Chem. Soc. Rev. 2013, 42, 1601–1618. [Google Scholar] [CrossRef] [PubMed]
- Bekere, L.; Gachet, D.; Lokshin, V.; Marine, W.; Khodorkovsky, V. Synthesis and spectroscopic properties of 4-amino-1,8-naphthalimide derivatives involving the carboxylic group: A new molecular probe for ZnO nanoparticles with unusual fluorescence features. Beilstein J. Org. Chem. 2013, 9, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Marinova, N.V.; Georgiev, N.I.; Bojinov, V.B. Facile synthesis, sensor activity and logic behaviour of 4-aryloxy substituted 1, 8-naphthalimide. J. Photochem. Photobiol. A 2013, 254, 54–61. [Google Scholar] [CrossRef]
- Saini, A.; Thomas, K.J.; Huang, Y.-J.; Ho, K.-C. Synthesis and characterization of naphthalimide-based dyes for dye sensitized solar cells. J. Mater. Sci. Mater. Electron. 2018, 29, 16565–16580. [Google Scholar] [CrossRef]
- Ulla, H.; Garudachari, B.; Satyanarayan, M.; Umesh, G.; Isloor, A. Blue organic light emitting materials: Synthesis and characterization of novel 1, 8-naphthalimide derivatives. Opt. Mater. 2014, 36, 704–711. [Google Scholar] [CrossRef]
- Medel, S.; Bosch, P.; Grabchev, I.; de la Torre, M.C.; Ramírez, P. Click chemistry to fluorescent hyperbranched polymeric sensors. 2. Synthesis, spectroscopic and cation-sensing properties of new green fluorescent 1, 8-naphthalimides. Eur. Polym. J. 2016, 74, 241–255. [Google Scholar] [CrossRef]
- Nicolescu, A.; Airinei, A.; Georgescu, E.; Georgescu, F.; Tigoianu, R.; Oancea, F.; Deleanu, C. Synthesis, photophysical properties and solvatochromic analysis of some naphthalene-1, 8-dicarboxylic acid derivatives. J. Mol. Liq. 2020, 303, 112626. [Google Scholar] [CrossRef]
- Bala, I.; Airinei, A.; Georgescu, E.; Oancea, F.; Georgescu, F.; Nicolescu, A.; Tigoianu, R.; Deleanu, C. Photophysical and biological properties of a strigolactone mimic derived from 1, 8-naphthalic anhydride. Rev. Roum. Chim. 2022, 67, 51–62. [Google Scholar]
- Tigoianu, R.; Airinei, A.; Georgescu, E.; Nicolescu, A.; Georgescu, F.; Isac, D.L.; Deleanu, C.; Oancea, F. Synthesis and solvent dependent fluorescence of some piperidine-substituted naphthalimide derivatives and consequences for water sensing. Int. J. Mol. Sci. 2022, 23, 2760. [Google Scholar] [CrossRef]
- MacAlpine, G.A.; Raphael, R.A.; Shaw, A.; Taylor, A.W.; Wild, H.-J. Synthesis of the germination stimulant (±)-strigol. J. Chem. Soc. Perkin Trans. 1 1976, 410–416. [Google Scholar] [CrossRef]
- Sugamoto, K.; Matsushita, Y.i.; Kameda, Y.h.; Suzuki, M.; Matsui, T. Microwave-assisted synthesis of N-hydroxyphthalimide derivatives. Synth. Commun. 2005, 35, 67–70. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Yoshimura, M.; Sato, Y.; Kuwata, K.; Toh, S.; Holbrook-Smith, D.; Zhang, H.; McCourt, P.; Itami, K.; Kinoshita, T.; et al. PARASITIC PLANTS. Probing strigolactone receptors in Striga hermonthica with fluorescence. Science 2015, 349, 864–868. [Google Scholar] [CrossRef] [PubMed]
- de Saint Germain, A.; Clavé, G.; Boyer, F.-D. Synthesis of profluorescent strigolactone probes for biochemical studies. In Strigolactones: Methods in Molecular Biology; Prandi, C., Cardinale, F., Eds.; Humana: New York, NY, USA, 2021; Volume 2309, pp. 219–231. [Google Scholar]
- Wang, D.-W.; Yu, S.-Y.; Pang, Z.-L.; Ma, D.-J.; Liang, L.; Wang, X.; Wei, T.; Yang, H.-Z.; Ma, Y.-Q.; Xi, Z. Discovery of a broad-spectrum fluorogenic agonist for strigolactone receptors through a computational approach. J. Agric. Food Chem. 2021, 69, 10486–10495. [Google Scholar] [CrossRef] [PubMed]
- de Saint Germain, A.; Clave, G.; Schouveiler, P.; Pillot, J.P.; Singh, A.V.; Chevalier, A.; Daignan Fornier, S.; Guillory, A.; Bonhomme, S.; Rameau, C.; et al. Expansion of the Strigolactone Profluorescent Probes Repertory: The Right Probe for the Right Application. Front. Plant Sci. 2022, 13, 887347. [Google Scholar] [CrossRef] [PubMed]
- Belmondo, S.; Marschall, R.; Tudzynski, P.; López Ráez, J.A.; Artuso, E.; Prandi, C.; Lanfranco, L. Identification of genes involved in fungal responses to strigolactones using mutants from fungal pathogens. Curr. Genet. 2017, 63, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Seto, Y.; Yasui, R.; Kameoka, H.; Tamiru, M.; Cao, M.; Terauchi, R.; Sakurada, A.; Hirano, R.; Kisugi, T.; Hanada, A.; et al. Strigolactone perception and deactivation by a hydrolase receptor DWARF14. Nat. Commun. 2019, 10, 191. [Google Scholar] [CrossRef]
- Fiorilli, V.; Forgia, M.; de Saint Germain, A.; D’Arrigo, G.; Cornu, D.; Le Bris, P.; Al-Babili, S.; Cardinale, F.; Prandi, C.; Spyrakis, F.; et al. A structural homologue of the plant receptor D14 mediates responses to strigolactones in the fungal phytopathogen Cryphonectria parasitica. New Phytol. 2022, 234, 1003–1017. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).