

Immunomodulatory Activity and Its Mechanisms of Two Polysaccharides from Poria cocos
Abstract
:1. Introduction
2. Results
2.1. Analysis of the Glycosidic Linkages of Polysaccharides
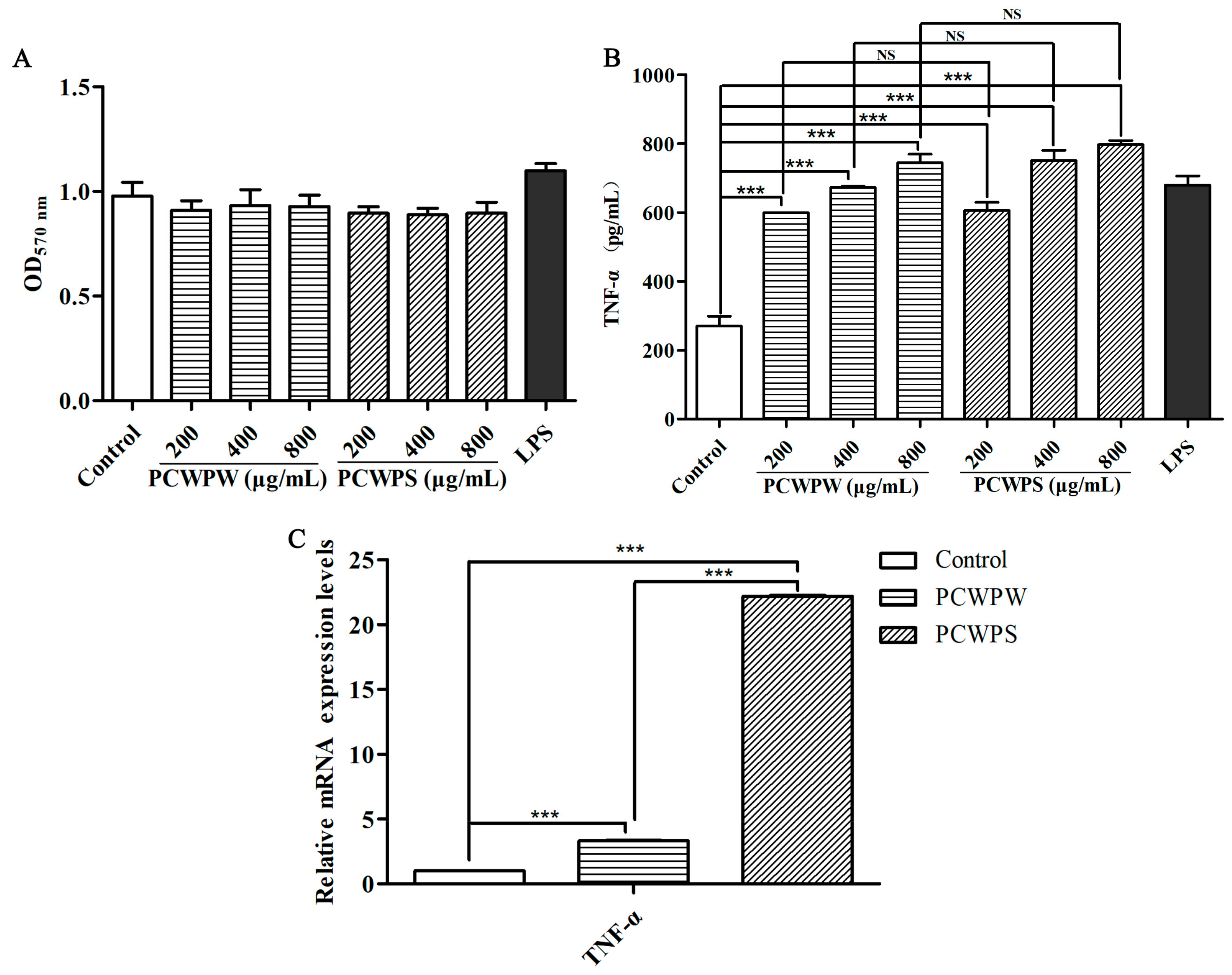
2.2. Effects of PCWPW and PCWPS on the TNF-α Production and mRNA Expressions in RAW264.7 Cells
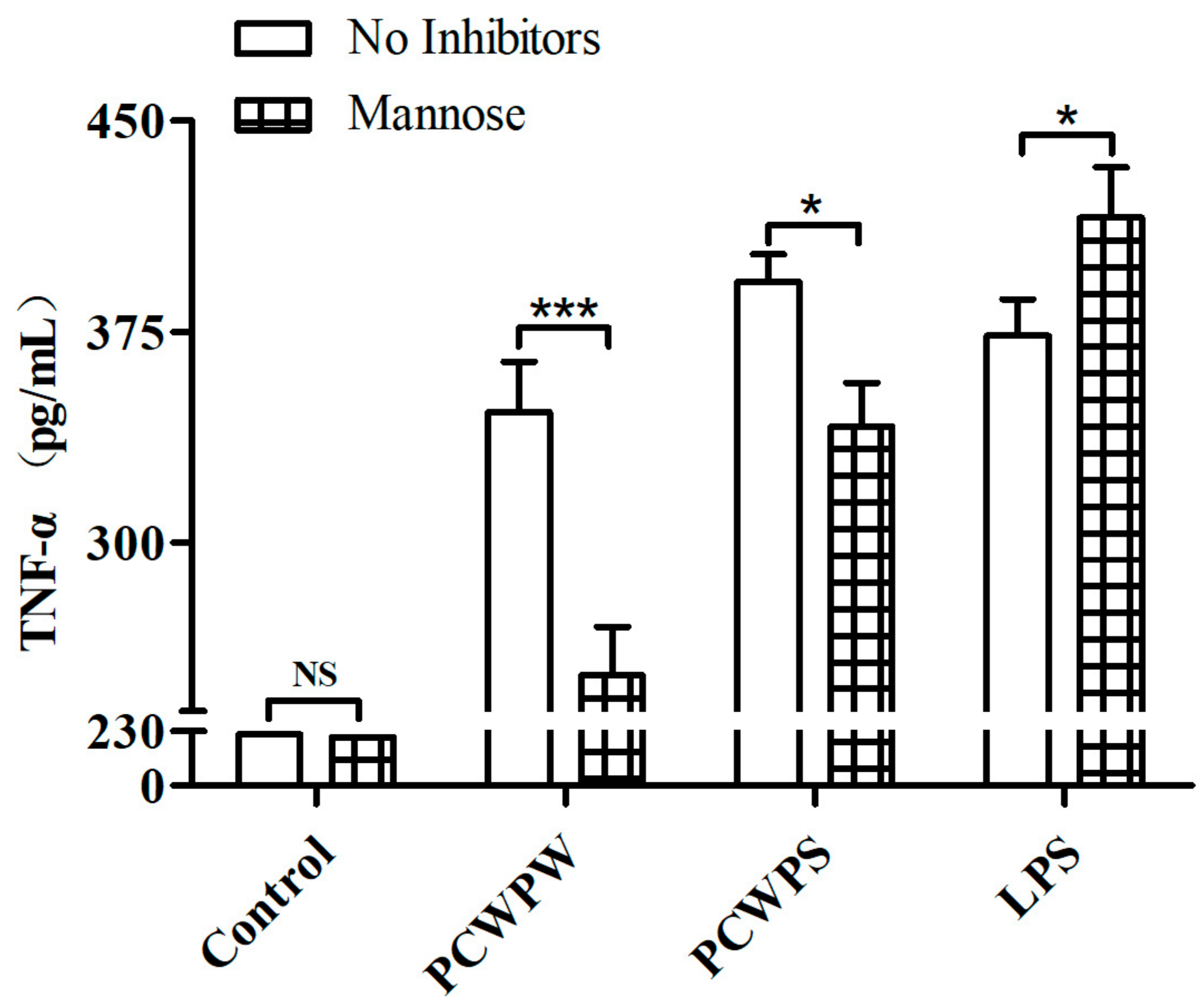
2.3. PCWPW and PCWPS Activated Macrophages by Binding to Mannose Receptor (MR)
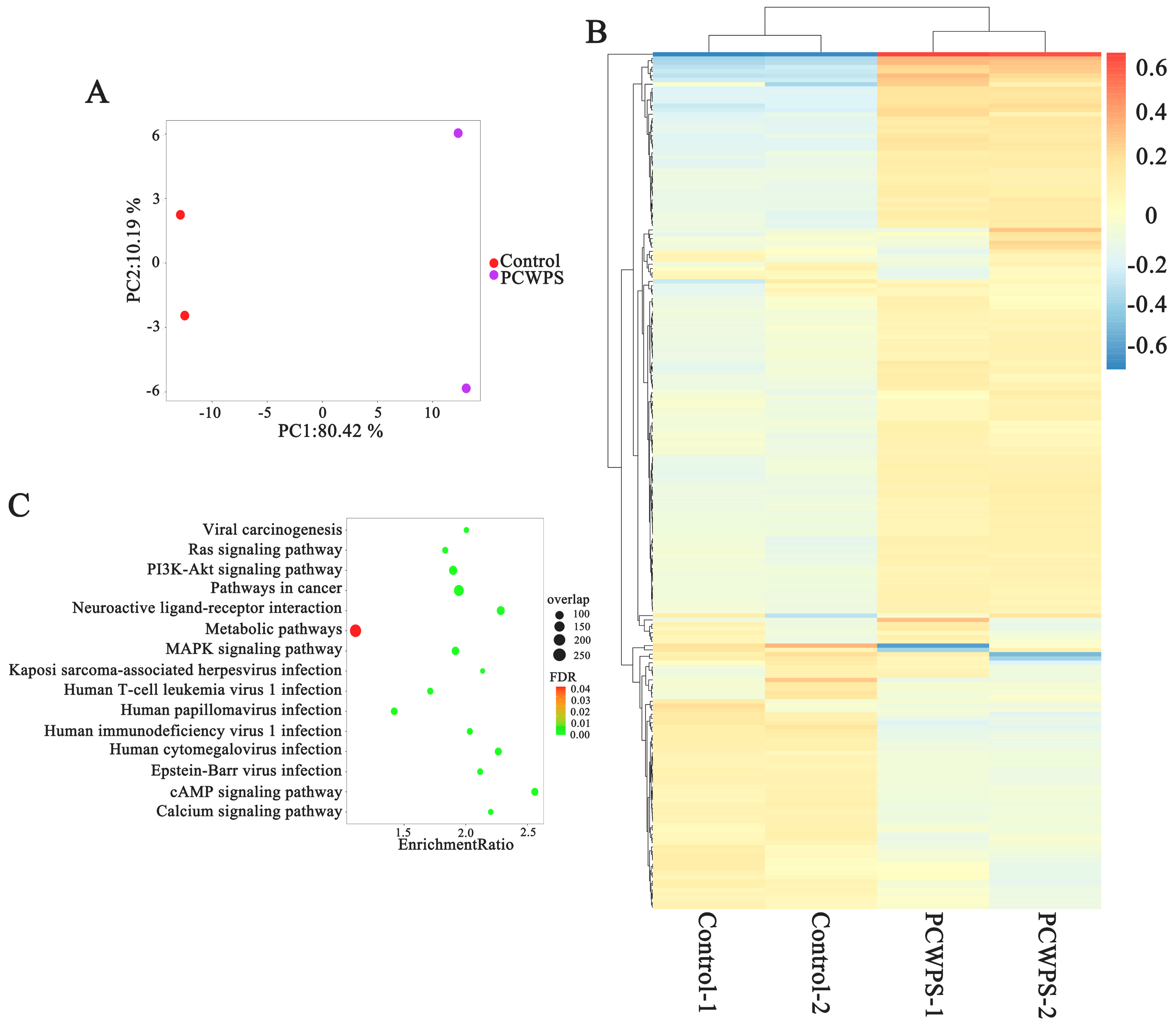
2.4. MAPK Pathway Activation Was Responsible for the Release of Proinflammatory Cytokines
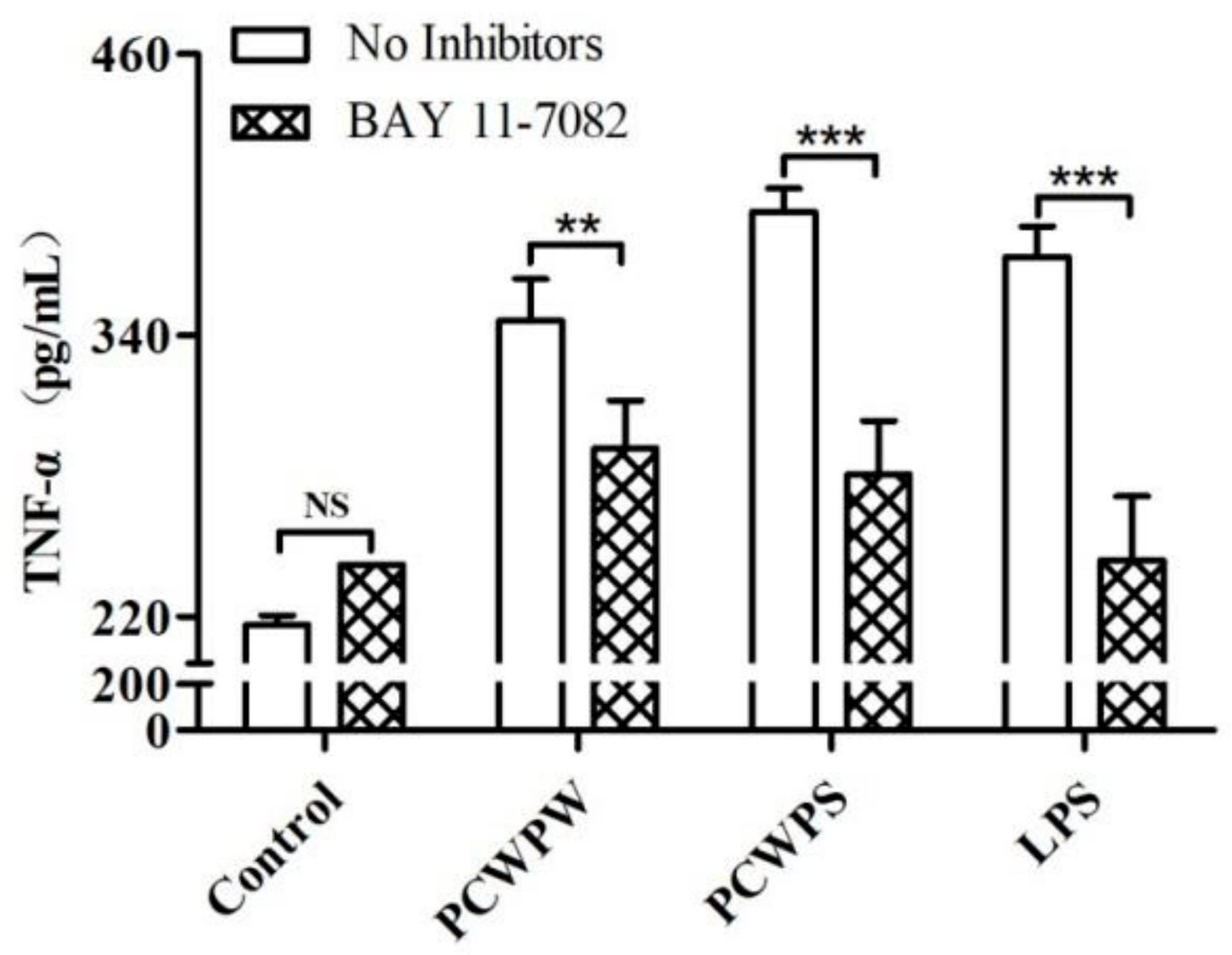
2.5. NF-κB Mediated the Immune Activity of PCWPW and PCWPS
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Methylation Analysis
4.3. Determine the Secretion of TNF-α Induced by PCWPW and PCWPS
4.4. Investigation of Mannose Receptor
4.5. Extraction, Preparation, and Sequencing of RNA-Seq Libraries
4.6. Target Prediction Based on Differential Gene Expression Profiles
4.7. Analysis of Transcription Levels of Differentially Expressed Genes
4.8. Cellula Pathway Inhibitor Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, W.J.; Wang, S.; Kang, C.Z.; Lv, C.G.; Guo, L.P. Pharmacodynamic material basis of traditional Chinese medicine based on biomacromolecules: A review. Plant Methods 2020, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.A.R.; Coimbra, M.A. The antioxidant activity of polysaccharides: A structure-function relationship overview. Carbohydr. Polym. 2023, 314, 120965. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Q.; Zhu, Y.; Li, Y.; Mei, S.; Luo, H.; Wu, K. Structural characterization of a mannoglucan polysaccharide from Dendrobium huoshanense and evaluation of its osteogenesis promotion activities. Int. J. Biol. Macromol. 2022, 211, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Guo, Y.; Huang, Q.; Shi, X.; Liu, Q.; Wang, F.; Liu, Q.; Yu, K.; Wang, Z. GPP(composition of Ganoderma lucidum polysaccharides and Polyporus umbellatus polysaccharides) protects against DSS-induced murine colitis by enhancing immune function and regulating intestinal flora. Food Sci. Hum. Wellness 2022, 11, 795–805. [Google Scholar] [CrossRef]
- Duan, Y.; Huang, J.; Sun, M.; Jiang, Y.; Wang, S.; Wang, L.; Yu, N.; Peng, D.; Wang, Y.; Chen, W.; et al. Poria cocos polysaccharide improves intestinal barrier function and maintains intestinal homeostasis in mice. Int. J. Biol. Macromol. 2023, 249, 125953. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.H.; Wang, L.; Chen, W.D.; Duan, Y.T.; Sun, M.J.; Huang, J.J.; Peng, D.Y.; Yu, N.J.; Wang, Y.Y.; Zhang, Y. Poria cocos polysaccharide prevents alcohol-induced hepatic injury and inflammation by repressing oxidative stress and gut leakiness. Front. Nutr. 2022, 9, 963598. [Google Scholar] [CrossRef]
- Li, W.; Yu, J.; Zhao, J.; Xiao, X.; Li, W.; Zang, L.; Yu, J.; Liu, H.; Niu, X. Poria cocos polysaccharides reduces high-fat diet-induced arteriosclerosis in ApoE-/- mice by inhibiting inflammation. Phytother. Res. 2021, 35, 2220–2229. [Google Scholar] [CrossRef]
- Afroz, R.; Tanvir, E.M.; Tania, M.; Fu, J.; Kamal, M.A.; Khan, M.A. LPS/TLR4 Pathways in Breast Cancer: Insights into Cell Signalling. Curr. Med. Chem. 2022, 29, 2274–2289. [Google Scholar] [CrossRef]
- Shin, M.S.; Song, J.H.; Choi, P.; Lee, J.H.; Kim, S.Y.; Shin, K.S.; Ham, J.; Kang, K.S. Stimulation of innate immune function by Panax ginseng after heat processing. J. Agr. Food Chem. 2018, 66, 4652–4659. [Google Scholar] [CrossRef]
- Zeng, P.; Li, J.; Chen, Y.; Zhang, L. The structures and biological functions of polysaccharides from traditional Chinese herbs. Prog. Mol. Biol. Transl. Sci. 2019, 163, 423–444. [Google Scholar] [CrossRef]
- Du, J.; Li, J.; Zhu, J.; Huang, C.; Bi, S.; Song, L.; Hu, X.; Yu, R. Structural characterization and immunomodulatory activity of a novel polysaccharide from Ficus carica. Food Funct. 2018, 9, 3930–3943. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Q.Z.; Li, L.D.; Zhou, X.W. Immunostimulatory effects of the intracellular polysaccharides isolated from liquid culture of Ophiocordyceps sinensis (Ascomycetes) on RAW264.7 cells via the MAPK and PI3K/Akt signaling pathways. J. Ethnopharmacol. 2021, 275, 114130. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zuo, Z.T.; Wang, Y.Z. The Traditional usages, chemical components and pharmacological activities of Wolfiporia cocos: A Review. Am. J. Chin. Med. 2022, 50, 389–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, L.; Li, P.; Zhao, J.; Duan, J. Antidepressant and immunosuppressive activities of two polysaccharides from Poria cocos (Schw.) Wolf. Int. J. Biol. Macromol. 2018, 120, 1696–1704. [Google Scholar] [CrossRef]
- Holbrook, J.; Lara-Reyna, S.; Jarosz-Griffiths, H.; McDermott, M. Tumour necrosis factor signalling in health and disease. F1000Research 2019, 8, F1000, Faculty Rev-1111. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Mi, Y.; Duan, Z.; Ma, P.; Fan, D. Structural characterization and immunomodulatory activity of a polysaccharide from Eurotium cristatum. Int. J. Biol. Macromol. 2020, 162, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ji, T.; Wang, L.; Qu, Y.; Wang, X.; Wang, W.; Lv, M.; Wang, Y.; Li, X.; Jiang, P. Exploration of the mechanism by which Huangqi Guizhi Wuwu decoction inhibits Lps-induced inflammation by regulating macrophage polarization based on network pharmacology. BMC Complement. Med. Ther. 2023, 23, 8. [Google Scholar] [CrossRef]
- Li, Y.; Jin, T.; Liu, N.; Wang, J.; Qin, Z.; Yin, S.; Zhang, Y.; Fu, Z.; Wu, Y.; Wang, Y.; et al. A short peptide exerts neuroprotective effects on cerebral ischemia-reperfusion injury by reducing inflammation via the miR-6328/IKKβ/NF-κB axis. J. NeuroInflamm. 2023, 20, 53. [Google Scholar] [CrossRef]
- Duan, G.L.; Yu, X.B. Isolation, purification, characterization, and antioxidant activity of low-molecular-weight polysaccharides from Sparassis latifolia. Int. J. Biol. Macromol. 2019, 137, 1112–1120. [Google Scholar] [CrossRef]
- Zavadinack, M.; de Lima Bellan, D.; da Rocha Bertage, J.L.; da Silva Milhorini, S.; da Silva Trindade, E.; Simas, F.F.; Sassaki, G.L.; Cordeiro, L.M.C.; Iacomini, M. An α-D-galactan and a β-D-glucan from the mushroom Amanita muscaria: Structural characterization and antitumor activity against melanoma. Carbohydr. Polym. 2021, 274, 118647. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, H.; Wei, K.; Zhang, T.; Che, Y.; Nguyễn, A.D.; Pandita, S.; Wan, X.; Cui, X.; Zhou, B.; et al. Structure of a new glycyrrhiza polysaccharide and its immunomodulatory activity. Front. Immunol. 2022, 13, 1007186. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yin, X.; Zhang, D.; Lu, J.; Wang, X. Isolation, Structural characterization and macrophage activation activity of an acidic polysaccharide from Raspberry Pulp. Molecules 2022, 27, 1674. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, S.; Huang, P.; Zhang, H.; Yu, C. Development of a prognostic model based on an immunogenomic landscape analysis of medulloblastoma. Biosci. Rep. 2021, 41, BSR20202907. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.A.-O.; Chang, S.; Liu, J.; Li, H.; Xu, P.; Wang, P.; Wang, X.; Zhao, M.; Zhao, B.; Wang, L.; et al. Physicochemical properties, antioxidant and antidiabetic activities of polysaccharides from Quinoa (Chenopodium quinoa Willd.) seeds. Molecules 2020, 25, 3840. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.D.S.; Sabry, D.A.; Silva, C.H.F.; Gomes, D.L.; Santana-Filho, A.P.; Sassaki, G.L.; Rocha, H.A.O. Immunostimulatory effect of sulfated galactans from the green seaweed Caulerpa cupressoides var. flabellata. Mar. Drugs. 2020, 18, 234. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Huang, G. Extraction, structure, and activity of polysaccharide from Radix astragali. Biomed. Pharmacother. 2022, 150, 113015. [Google Scholar] [CrossRef]
- Li, K.; Cao, Y.-X.; Jiao, S.-M.; Du, G.-H.; Du, Y.-G.; Qin, X.-M. Structural characterization and immune activity screening of polysaccharides with different molecular weights from Astragali Radix. Front. Pharmacol. 2020, 11, 582091. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, N.; Sun, C.; Sun, D.; Wang, Y. Polysaccharides from Polygonatum sibiricum Delar. ex Redoute induce an immune response in the RAW264.7 cell line via an NF-κB/MAPK pathway. RSC Adv. 2019, 9, 17988–17994. [Google Scholar] [CrossRef]
- Wang, H.; Ma, C.; Sun-Waterhouse, D.; Wang, J.; Waterhouse, G.I.N.; Kang, W. Immunoregulatory polysaccharides from Apocynum venetum L. flowers stimulate phagocytosis and cytokine expression via activating the NF-κB/MAPK signaling pathways in RAW264.7 cells. Food Sci. Hum. Wellness 2022, 11, 806–814. [Google Scholar] [CrossRef]
- Qin, L.; Chen, S.; Xie, L.; Yu, Q.; Chen, Y.; Shen, M.; Xie, J. Mechanisms of RAW264.7 macrophages immunomodulation mediated by polysaccharide from mung bean skin based on RNA-seq analysis. Food Res. Int. 2022, 154, 111017. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Lin, R.; Hou, X.; Wu, J.; Zhao, W.; Ma, H.; Fan, Z.; Li, S.; Zhu, Y.; Zhang, D. Immunomodulatory mechanism of a purified polysaccharide isolated from Isaria cicadae Miquel on RAW264.7 cells via activating TLR4-MAPK-NF-κB signaling pathway. Int. J. Biol. Macromol. 2020, 164, 4329–4338. [Google Scholar] [CrossRef] [PubMed]
- Talapphet, N.; Palanisamy, S.; Li, C.; Ma, N.; Prabhu, N.M.; You, S. Polysaccharide extracted from Taraxacum platycarpum root exerts immunomodulatory activity via MAPK and NF-κB pathways in RAW264.7 cells. J. Ethnopharmacol. 2021, 281, 114519. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, D.; Wang, H.; Wan, X. Protective effect of Poria cocos polysaccharides on fecal peritonitis-induced sepsis in mice through inhibition of oxidative stress, inflammation, apoptosis, and reduction of Treg cells. Front. Microbiol. 2022, 13, 887949. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.K.; Kim, Y.E.; Kim, A.; Jung, J.; Son, M.J. The herbal drug, Bu-Zhong-Yi-Qi-Tang, for the treatment of atopic dermatitis: Protocol for a systematic review. Medicine 2019, 98, e13938. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, H.; Wang, S.; Xiang, Z.; Kong, H.; Xue, Q.; He, M.; Yu, X.; Li, Y.; Sun, D.; et al. A review on the advances in the extraction methods and structure elucidation of Poria cocos polysaccharide and its pharmacological activities and drug carrier applications. Int. J. Biol. Macromol. 2022, 217, 536–551. [Google Scholar] [CrossRef] [PubMed]
- Oladele, A.K.; Duodu, K.G.; Emmambux, N.M. Hydrolysis and antioxidant activity of starch modified with phenolic extracts from grape pomace and sorghum bran under alkaline conditions. Carbohydr. Polym. 2020, 240, 116291. [Google Scholar] [CrossRef]
- Wen, L.; Sheng, Z.; Wang, J.; Jiang, Y.; Yang, B. Structure of water-soluble polysaccharides in spore of Ganoderma lucidum and their anti-inflammatory activity. Food Chem. 2022, 373, 131374. [Google Scholar] [CrossRef]
- Feng, H.; Feng, Y.; Lin, L.; Wu, D.; Liu, Q.; Li, H.; Zhang, X.; Li, S.; Tang, F.; Liu, Z.; et al. Mannose receptor-mediated carbon nanotubes as an antigen delivery system to enhance immune response both in vitro and in vivo. Int. J. Mol. Sci. 2022, 23, 4239. [Google Scholar] [CrossRef]
- Li, Z.; Spoelstra, N.S.; Sikora, M.A.-O.; Sams, S.B.; Elias, A.; Richer, J.A.-O.; Lee, A.A.-O.X.; Oesterreich, S.A.-O. Mutual exclusivity of ESR1 and TP53 mutations in endocrine resistant metastatic breast cancer. NPJ Breast Cancer 2022, 8, 62. [Google Scholar] [CrossRef]
- Gonther, S.; Bachmann, M.; Goren, I.; Huard, A.; Weigert, A.A.-O.; Köhl, J.A.-O.; Mühl, H.A.-O. 3′mRNA sequencing reveals pro-regenerative properties of c5ar1 during resolution of murine acetaminophen-induced liver injury. NPJ Regen. Med. 2022, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Hu, Y.; Zhan, L.; He, J.; Lu, J.; Gao, C.; Du, W.; Yue, A.; Zhao, J.; Zhang, W. A natural glucan from Black Bean inhibits cancer cell proliferation via PI3K-Akt and MAPK Pathway. Molecules 2023, 28, 1971. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, T.; Liu, Y.; Chen, T.; Zhang, Q.; Li, W.; Zhou, H.; Zhang, Y.; Zhang, Z.A.-O. Molecular insights into salinity responsiveness in contrasting genotypes of rice at the seedling stage. Int. J. Mol. Sci. 2022, 23, 1624. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.; Lin, Y.; Zhang, Z.; Qiu, L.; Ding, W.; Gao, Q.; Xue, J.; Li, Y.; He, H. The transcriptomic profile of Spodoptera frugiperda differs in response to a novel insecticide, cyproflanilide, compared to chlorantraniliprole and avermectin. BMC Genom. 2023, 24, 3. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Linkage | Mass Fragments (m/z) | Molar Ratios (%) | |
|---|---|---|---|
| PCWPW | PCWPS | ||
| Araf-1 | 43, 71, 87, 101, 117, 129, 145, 161 | - | 0.2 |
| Fucp-1-3 | 43, 89, 101, 117, 131, 159, 173, 233 | 0.4 | 0.85 |
| Glcp-1 | 43, 71, 87, 101, 117, 129, 145, 161, 205 | 5.07 | 3.99 |
| Araf-1-5 | 43, 71, 87, 99, 101, 117, 129, 161, 189 | 0.78 | 1.85 |
| Glcp-1-3 | 43, 71, 87, 99, 101, 117, 129, 161, 173, 233 | 78.39 | 72.78 |
| Glcp-1-4 | 43, 71, 87, 99, 101, 113, 117, 129, 131, 161, 173, 233 | 3.24 | 1.49 |
| Glcp-1-6 | 43, 71, 87, 99, 101, 117, 129, 161, 173, 189, 233 | 3.67 | 2.38 |
| Galp-1-6 | 43, 71, 87, 99, 101, 117, 129, 161, 173, 189, 233 | 1.81 | 4.61 |
| Glcp-1-3-6 | 43, 87, 99, 101, 117, 129, 139, 159, 173, 189, 233 | 5.57 | 4.72 |
| Manp-1-2-6 | 43, 87, 99, 113, 129, 173, 189, 233 | 1.07 | 3.65 |
| Manp-1-2 | 43,87,129,161,189 | - | 1.97 |
| Galp-1-3-6 | 43,87,117,129,159,189,233 | - | 1.51 |
| Primer Name | Sequences |
|---|---|
| actin-β | Forward: 5′-TCACCCACACTGTGCCCATCTACGA-3′ |
| Reverse: 5′-GGATGCCACAGGATTCCATACCCA-3′ | |
| TNF-α | Forward: 5′-ATAGCTCCCAGAAAAGCAAGC-3′ |
| Reverse: 5′-CACCCCGAAGTTCAGTAGACA-3′ | |
| JNK | Forward: 5′-ATTGAACAGCTCGGAACACC-3′ |
| Reverse: 5′-GAGTCAGCTGGGAAAAGCAC-3′ | |
| RRAS2 | Forward: 5′-AATGCCCTCCTTCACCAG-3′ |
| Reverse: 5′-GCAGCCTTTCTTGTCTTTT-3′ | |
| ERK | Forward: 5′-TGACCTCAAGCCTTCCAACC-3′ |
| Reverse: 5′-ATCTGGATCTGCAACACGGG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; He, J.; Zheng, D.; Zhao, P.; Wang, Y.; Zhao, J.; Li, P. Immunomodulatory Activity and Its Mechanisms of Two Polysaccharides from Poria cocos. Molecules 2024, 29, 50. https://doi.org/10.3390/molecules29010050
Zhang W, He J, Zheng D, Zhao P, Wang Y, Zhao J, Li P. Immunomodulatory Activity and Its Mechanisms of Two Polysaccharides from Poria cocos. Molecules. 2024; 29(1):50. https://doi.org/10.3390/molecules29010050
Chicago/Turabian StyleZhang, Wuxia, Jiaqi He, Danping Zheng, Panpan Zhao, Yingdong Wang, Jinzhong Zhao, and Peng Li. 2024. "Immunomodulatory Activity and Its Mechanisms of Two Polysaccharides from Poria cocos" Molecules 29, no. 1: 50. https://doi.org/10.3390/molecules29010050
APA StyleZhang, W., He, J., Zheng, D., Zhao, P., Wang, Y., Zhao, J., & Li, P. (2024). Immunomodulatory Activity and Its Mechanisms of Two Polysaccharides from Poria cocos. Molecules, 29(1), 50. https://doi.org/10.3390/molecules29010050