



Improved Stability and Catalytic Efficiency of ω-Transaminase in Aqueous Mixture of Deep Eutectic Solvents
 ,
,
Abstract
1. Introduction
2. Results and Discussion
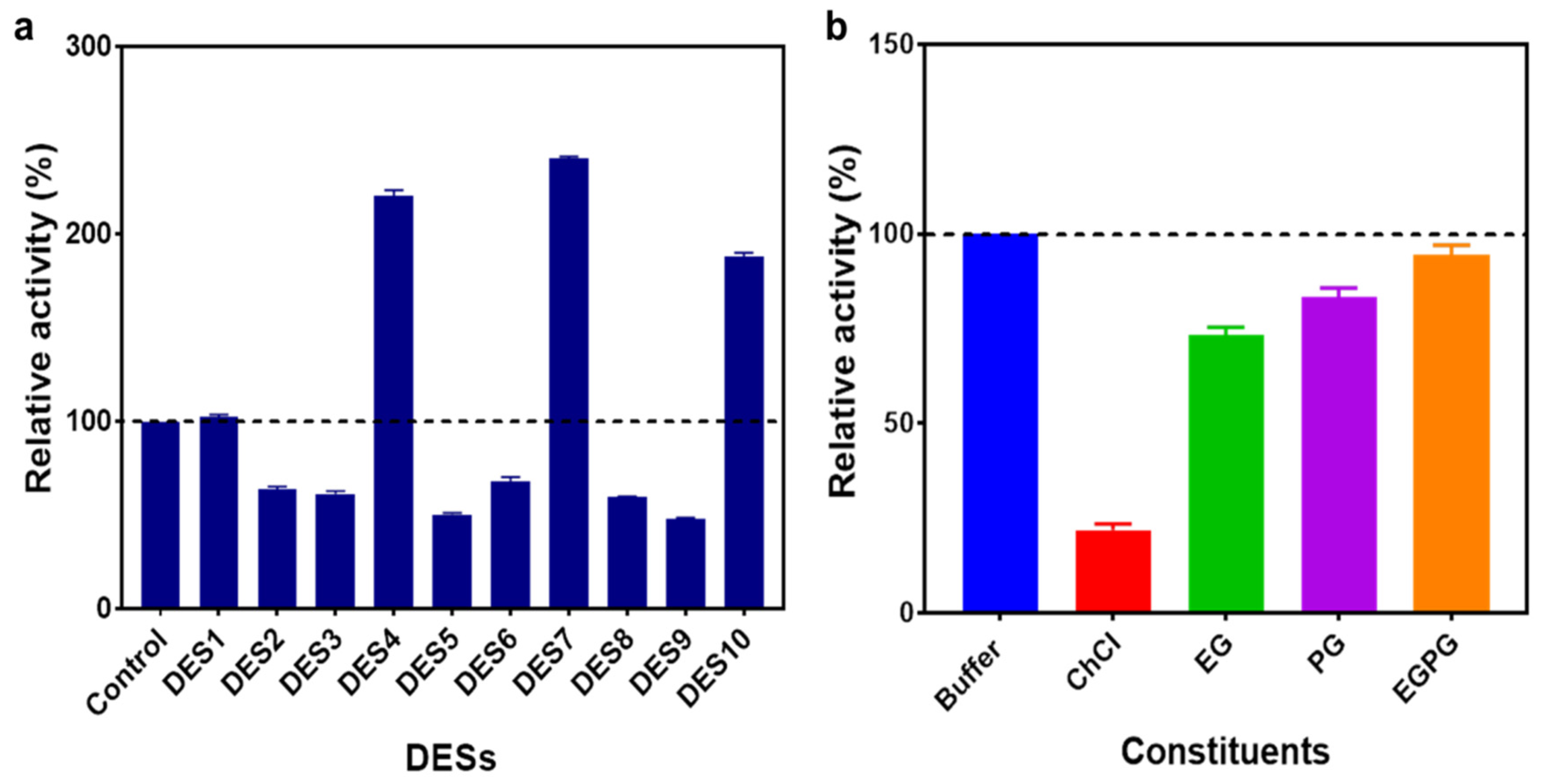
2.1. Screening of Various DESs in Enzyme-Catalysed Reaction
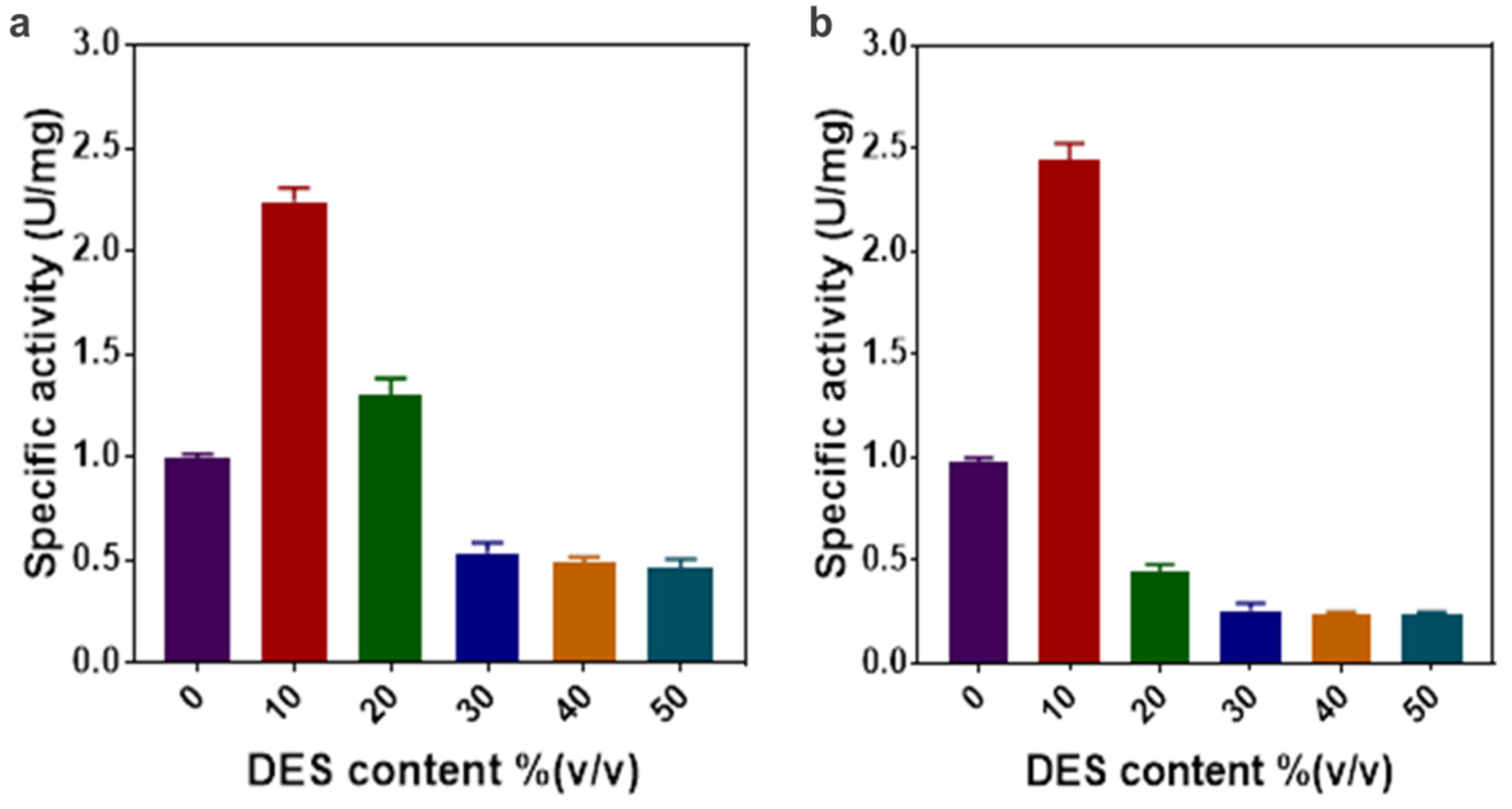
2.2. The Effect of DES Concentration on Enzyme Activity
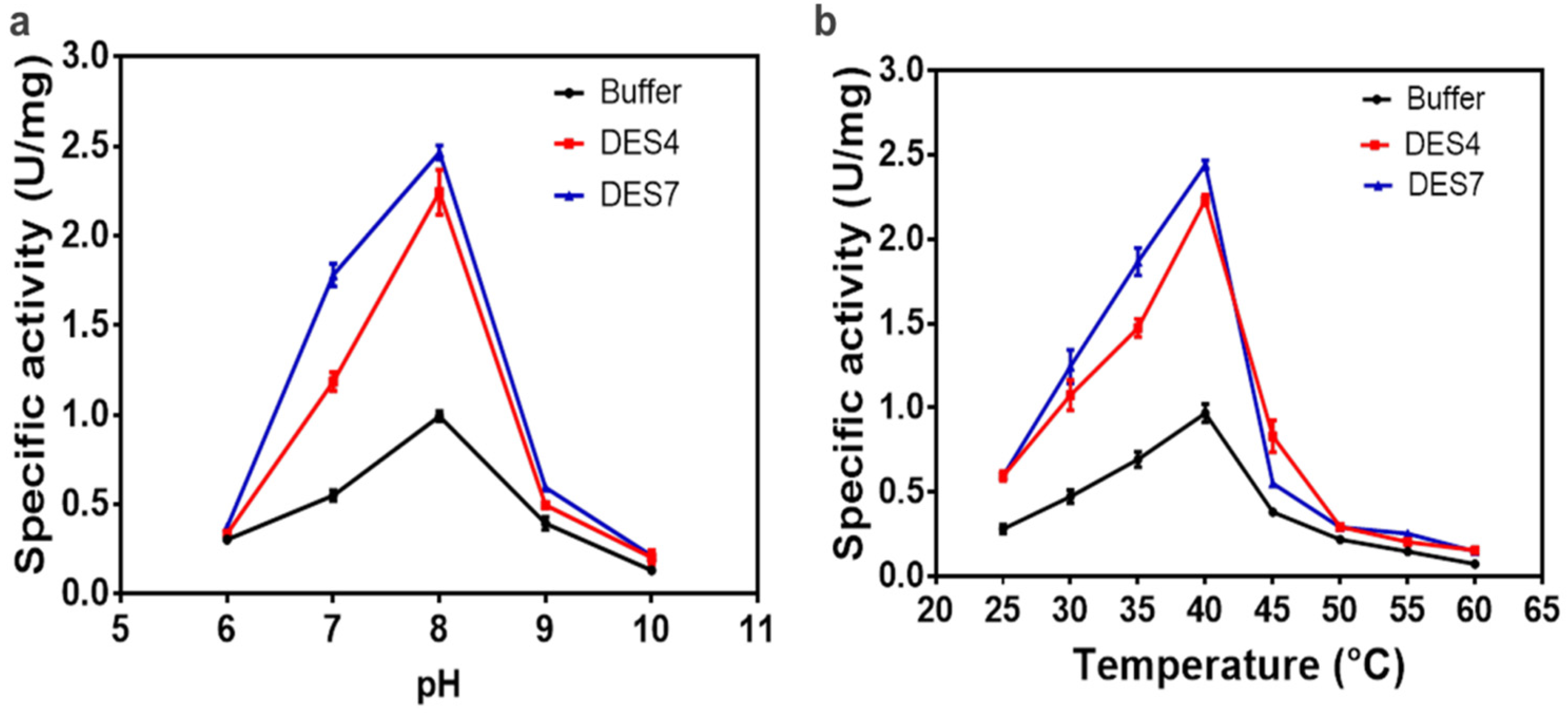
2.3. The Effect of pH and Temperature on Enzyme Activity
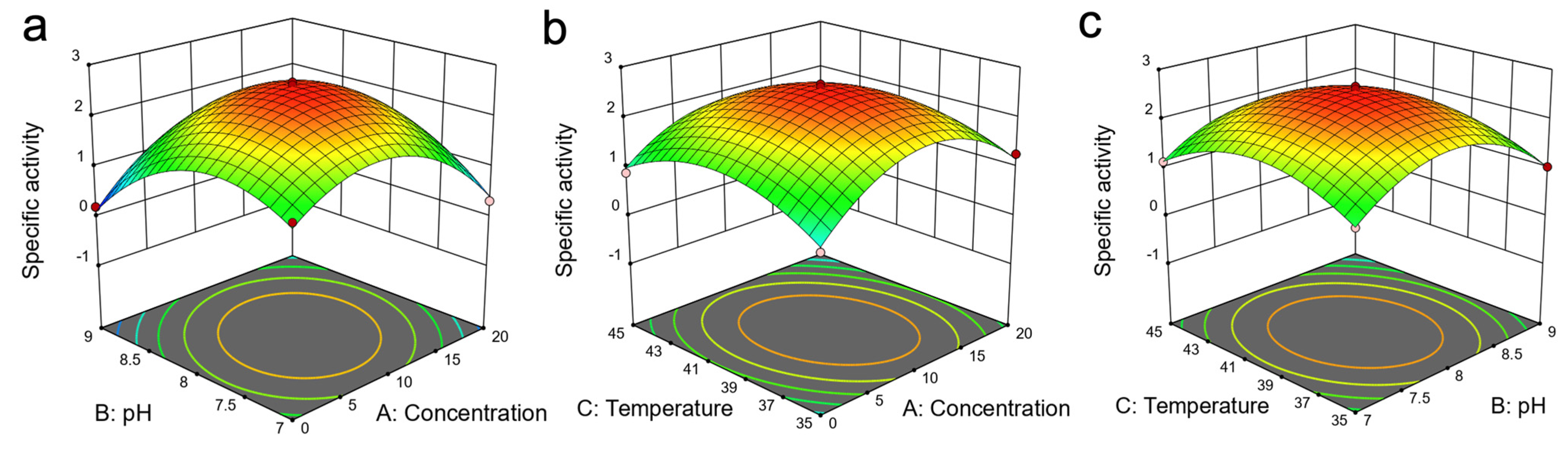
2.4. Optimisation of Reaction Conditions Using Response Surface Method
2.5. Determination of the Kinetic Parameters in the Presence of DESs
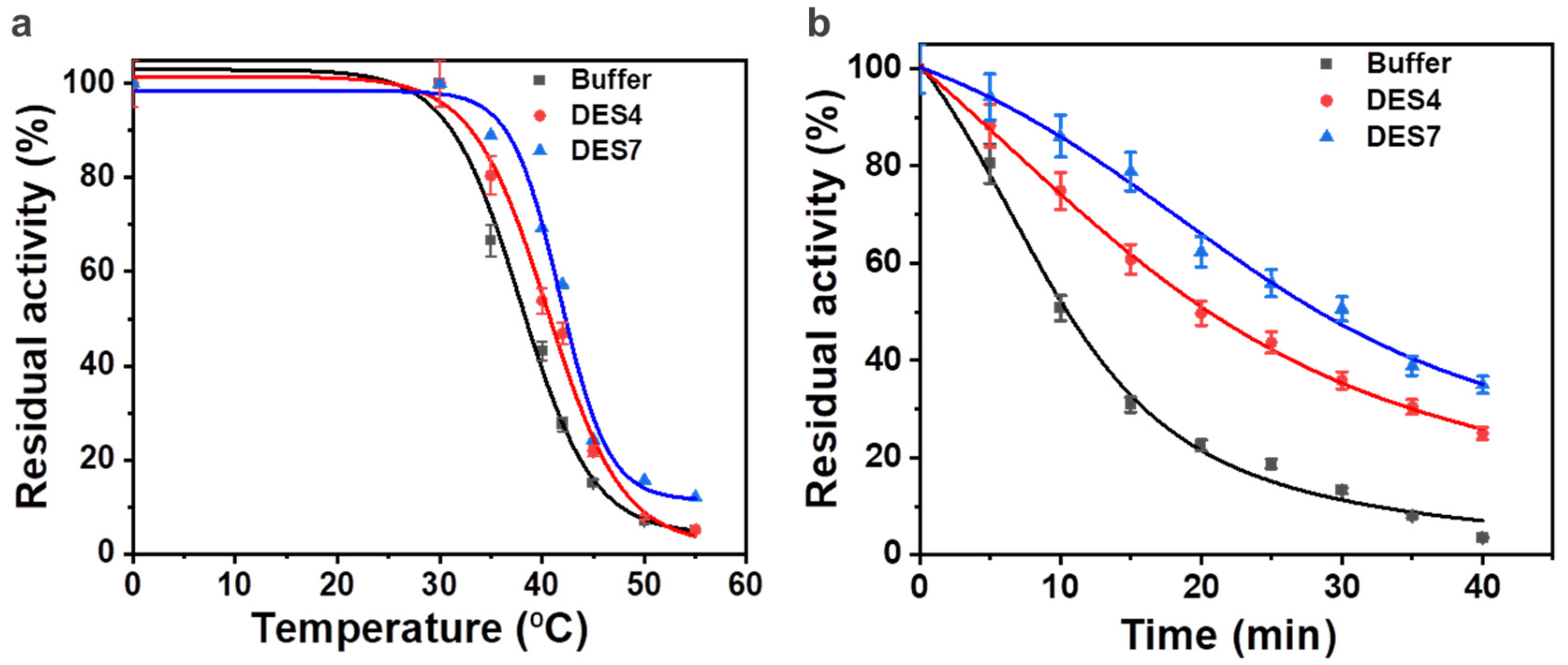
2.6. Stability of the ω-Transaminase in Aqueous Solutions of DES
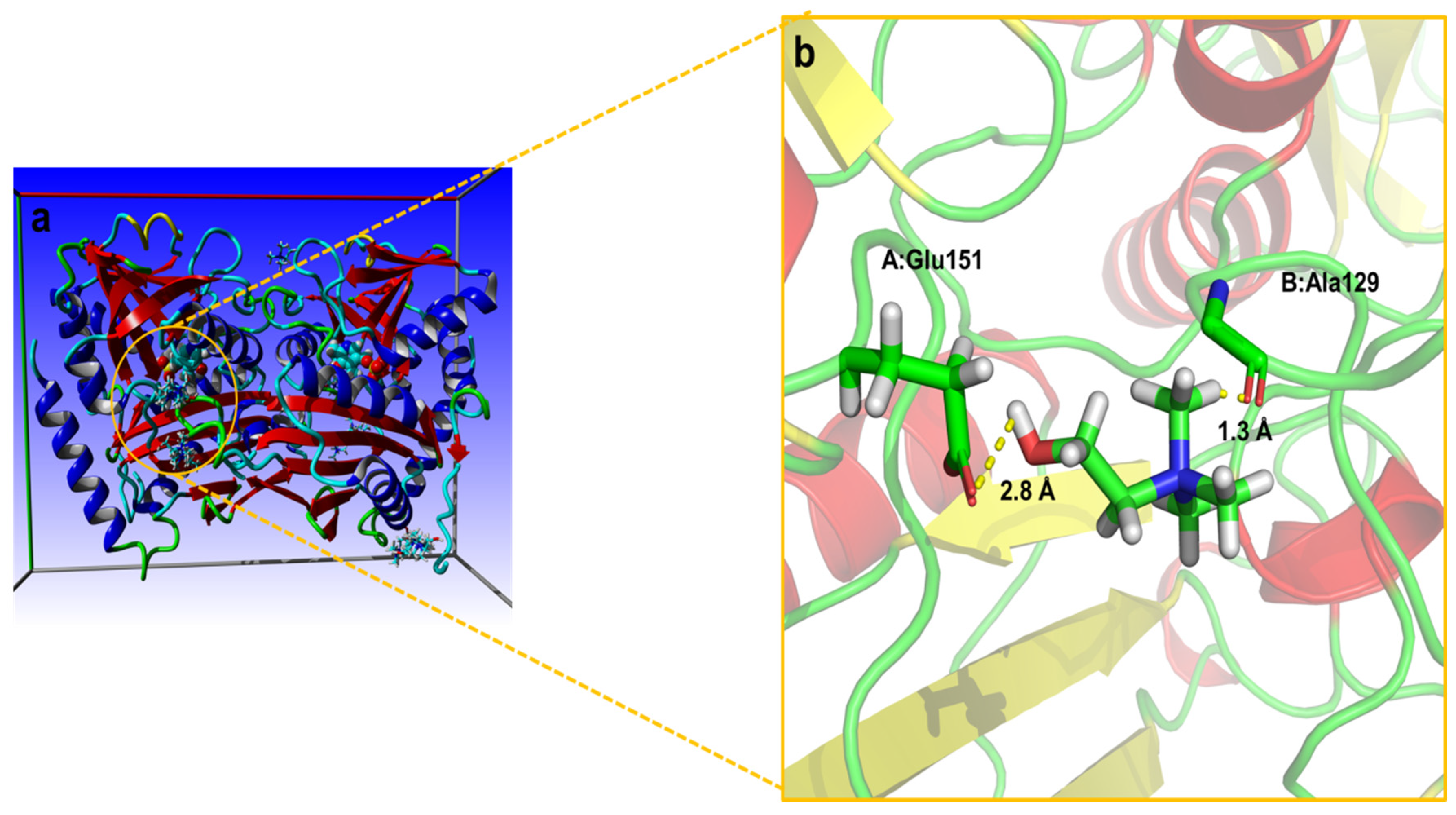
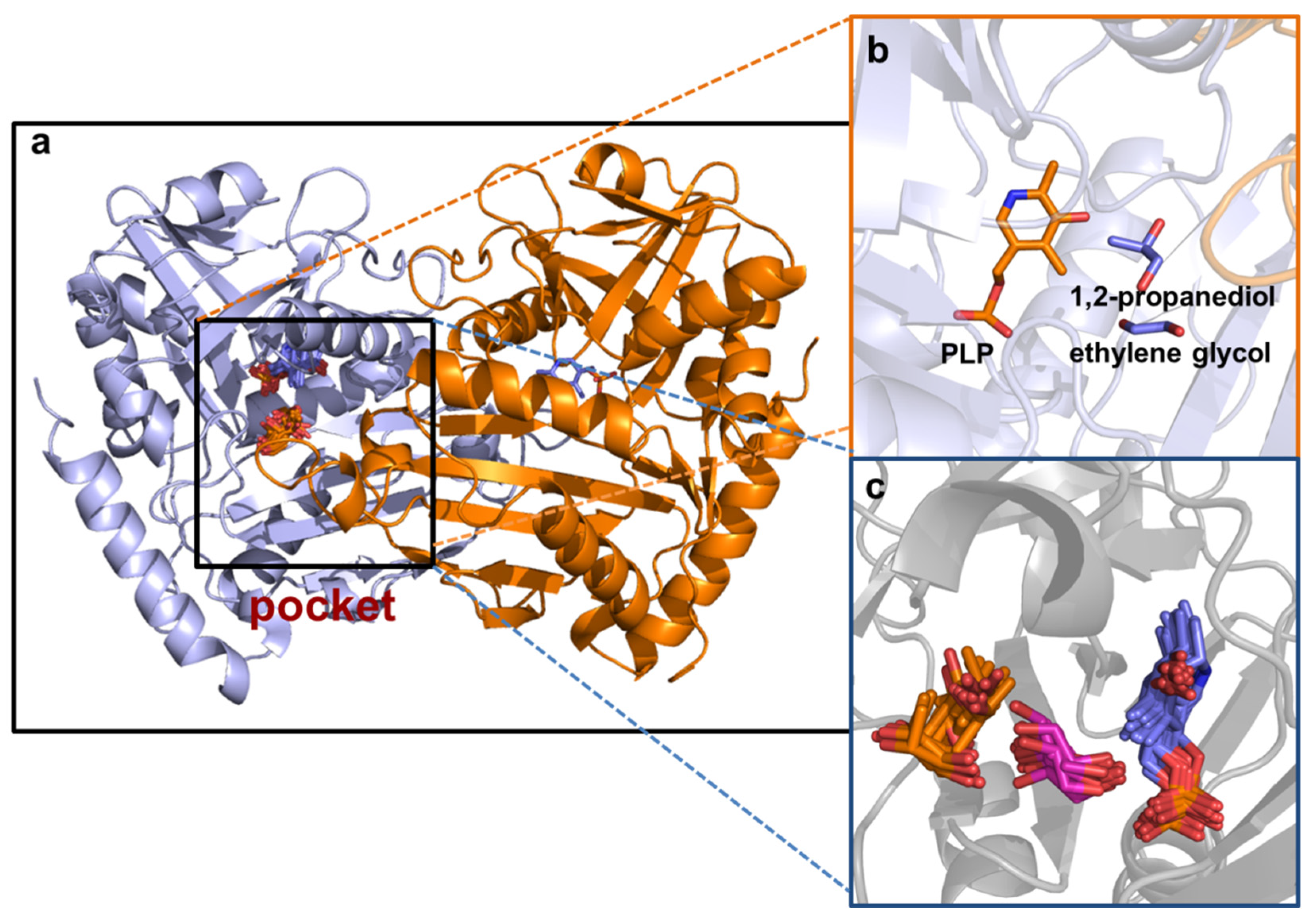
2.7. Molecular Docking Simulation
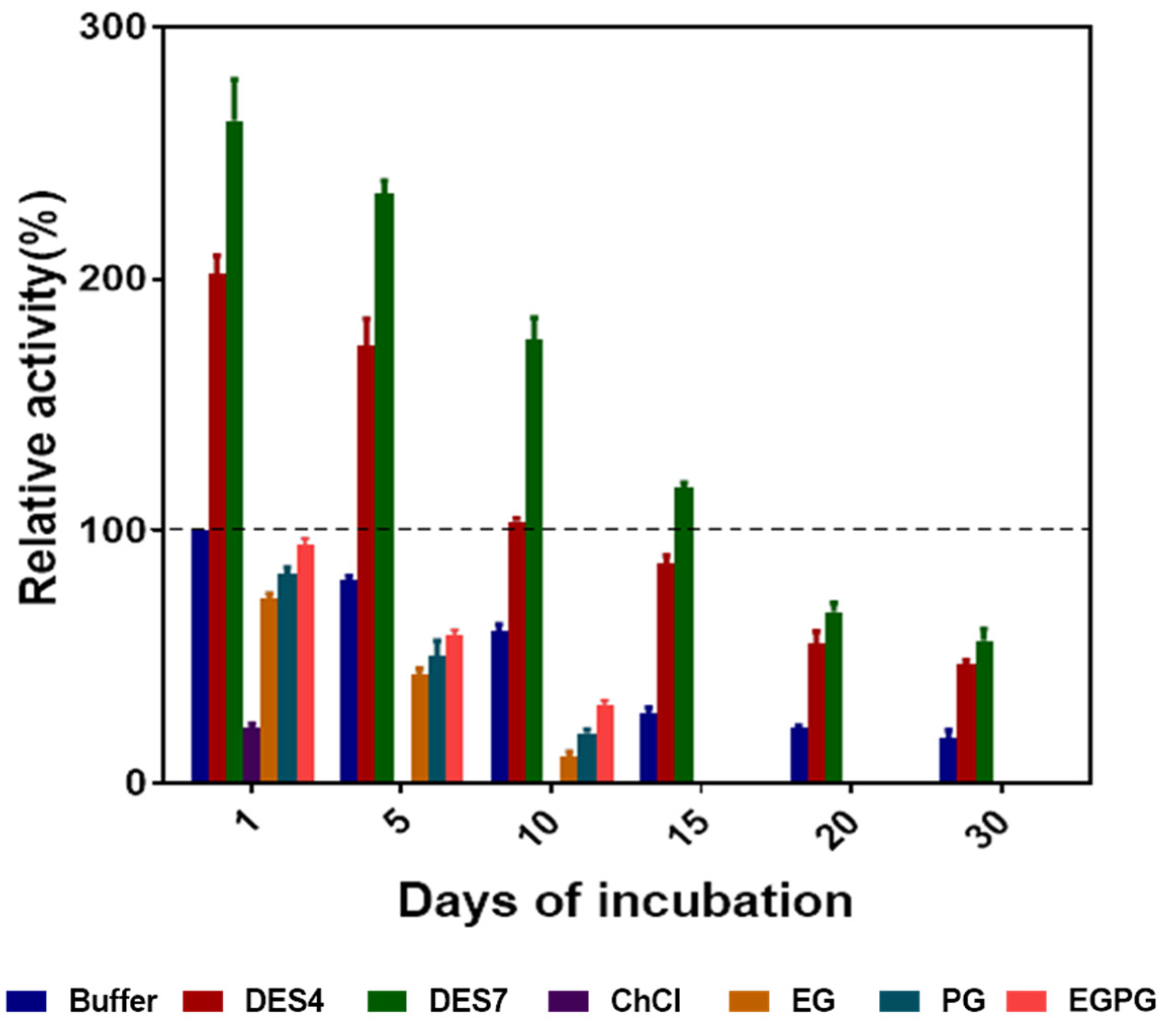
2.8. Storage Stability
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. DES Preparation
3.2.2. Induction and Purification of ω-Transaminase
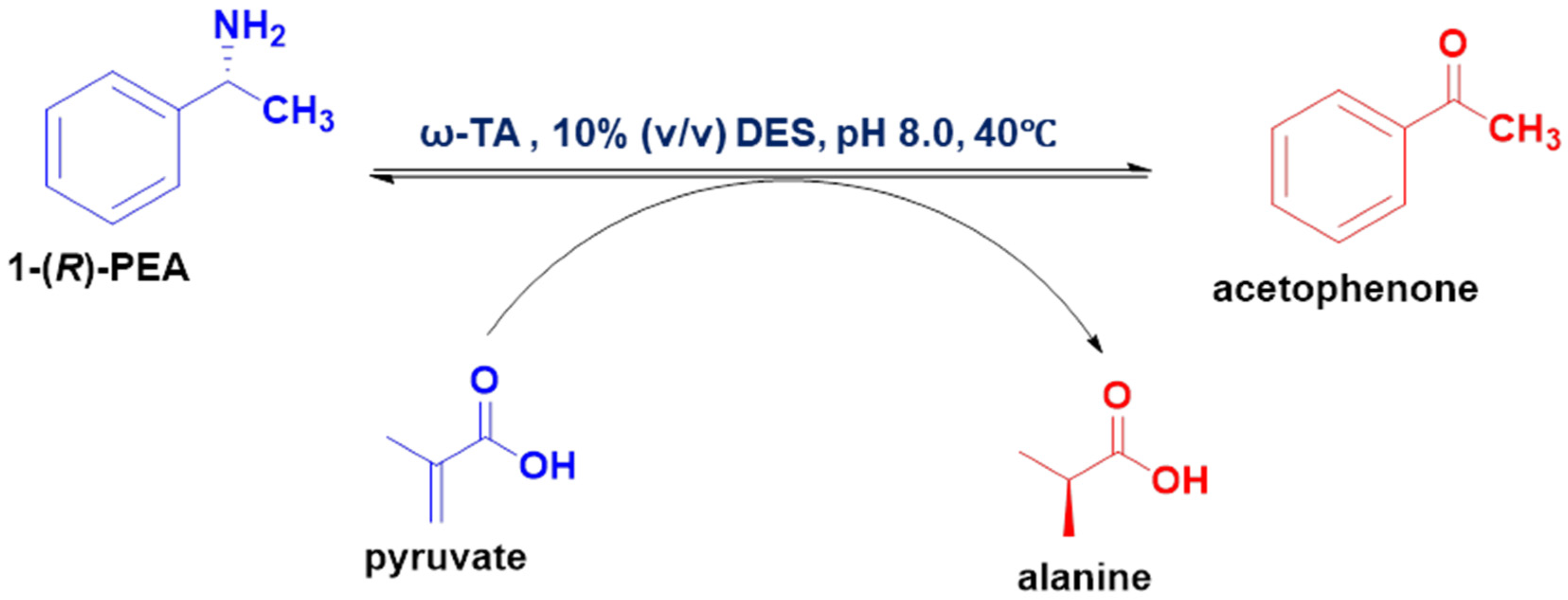
3.2.3. Screening of DESs in ω-Transaminase-Catalysed Reactions
3.2.4. Activity of ω-Transaminase in Various Aqueous Solutions of DES Concentrations
3.2.5. Factors Influencing ω-Transaminase Activity in the Presence of Aqueous Solutions of DES
3.2.6. Kinetics and Thermal Stability of ω-Transaminase
3.2.7. Storage Stability of ω-Transaminase in DES
3.2.8. Product Quantification by HPLC
3.2.9. Molecular Docking Simulation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gao, S.; Su, Y.; Zhao, L.; Li, G.; Zheng, G. Characterization of a (R)-selective amine transaminase from Fusarium oxysporum. Process. Biochem. 2017, 63, 130–136. [Google Scholar] [CrossRef]
- Malik, M.S.; Park, E.; Shin, J. Features and technical applications of omega-transaminases. Appl. Microbiol. Biotechnol. 2012, 94, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Kohls, H.; Steffen-Munsberg, F.; Höhne, M. Recent achievements in developing the biocatalytic toolbox for chiral amine synthesis. Curr. Opin. Chem. Biol. 2014, 19, 180–192. [Google Scholar] [CrossRef]
- Kelly, S.A.; Pohle, S.; Wharry, S.; Mix, S.; Allen, C.; Moody, T.; Gilmore, B. Application of omega-Transaminases in the Pharmaceutical Industry. Chem. Rev. 2018, 118, 349–367. [Google Scholar] [CrossRef]
- Ghislieri, D.; Turner, N.J. Biocatalytic Approaches to the Synthesis of Enantiomerically Pure Chiral Amines. Top. Catal. 2014, 57, 284–300. [Google Scholar] [CrossRef]
- Neto, W.; Schürmann, M.; Panella, L.; Vogel, A.; Woodley, J.M. Immobilisation of ω-transaminase for industrial application: Screening and characterisation of commercial ready to use enzyme carriers. J. Mol. Catal. B Enzym. 2015, 117, 54–61. [Google Scholar] [CrossRef]
- Kelly, S.A.; Mix, S.; Moody, T.S.; Gilmore, B.F. Transaminases for industrial biocatalysis: Novel enzyme discovery. Appl. Microbiol. Biotechnol. 2020, 104, 4781–4794. [Google Scholar] [CrossRef]
- Fesko, K.; Steiner, K.; Breinbauer, R.; Schwab, H.; Schürmann, M.; Strohmeier, G.A. Investigation of one-enzyme systems in the ω-transaminase-catalyzed synthesis of chiral amines. J. Mol. Catal. B Enzym. 2013, 96, 103–110. [Google Scholar] [CrossRef]
- Rueda, N.; dos Santos, J.C.S.; Ortiz, C.; Torres, R.; Barbosa, O.; Rodrigues, R.C.; Murcia, B.; Fernandez-Lafuente, R. Chemical Modification in the Design of Immobilized Enzyme Biocatalysts: Drawbacks and Opportunities. Chem. Rec. 2016, 16, 1436–1455. [Google Scholar] [CrossRef]
- Ni, J.; Li, Q.; Gong, L.; Liao, X.-L.; Zhang, Z.-J.; Ma, C.; He, Y. Highly Efficient Chemoenzymatic Cascade Catalysis of Biomass into Furfurylamine by a Heterogeneous Shrimp Shell-Based Chemocatalyst and an ω-Transaminase Biocatalyst in Deep Eutectic Solvent–Water. ACS Sustain. Chem. Eng. 2021, 9, 13084–13095. [Google Scholar] [CrossRef]
- Perna, F.M.; Vitale, P.; Capriati, V. Deep eutectic solvents and their applications as green solvents. Curr. Opin. Green Sustain. Chem. 2020, 21, 27–33. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Ríos-Lombardía, N.; Winkler, C.K.; Schmidt, S.; Morís, F.; Kroutil, W.; González-Sabín, J.; Kourist, R. Using Deep Eutectic Solvents to Overcome Limited Substrate Solubility in the Enzymatic Decarboxylation of Bio-Based Phenolic Acids. ACS Sustain. Chem. Eng. 2019, 7, 16364–16370. [Google Scholar] [CrossRef]
- Pätzold, M.; Siebenhaller, S.; Kara, S.; Liese, A.; Syldatk, C.; Holtmann, D. Deep eutectic solvents as efficient solvents in biocatalysis. Trends Biotechnol. 2019, 37, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Abbott, A.P.; Capper, G.; Davies, D.L.; Rasheed, R.K.; Tambyrajah, V. Novel solvent properties of choline chloride/urea mixtures. Chem. Commun. 2003, 39, 70–71. [Google Scholar] [CrossRef]
- Zhang, Q.; De Oliveira Vigier, K.; Royer, S.; Jérôme, F. Deep eutectic solvents: Syntheses, properties and applications. Chem. Soc. Rev. 2012, 41, 7108–7146. [Google Scholar] [CrossRef]
- Yang, T.-X.; Zhao, L.-Q.; Wang, J.; Song, G.-L.; Liu, H.-M.; Cheng, H.; Yang, Z. Improving Whole-Cell Biocatalysis by Addition of Deep Eutectic Solvents and Natural Deep Eutectic Solvents. ACS Sustain. Chem. Eng. 2017, 5, 5713–5722. [Google Scholar] [CrossRef]
- Durand, E.; Lecomte, J.; Baréa, B.; Piombo, G.; Dubreucq, E.; Villeneuve, P. Evaluation of deep eutectic solvents as new media for Candida antarctica B lipase catalyzed reactions. Process. Biochem. 2012, 47, 2081–2089. [Google Scholar] [CrossRef]
- Bubalo, M.C.; Tušek, A.; VinkoviĿ, M.; RadoševiĿ, K.; SrĿek, V.; RedovnikoviĿ, I. Cholinium-based deep eutectic solvents and ionic liquids for lipase-catalyzed synthesis of butyl acetate. J. Mol. Catal. B Enzy. 2015, 122, 188–198. [Google Scholar] [CrossRef]
- Cao, J.; Wu, R.; Zhu, F.; Dong, Q.; Su, E. How to improve the efficiency of biocatalysis in non-aqueous pure deep eutectic solvents: A case study on the lipase-catalyzed transesterification reaction. Biochem. Eng. J. 2022, 179, 108336. [Google Scholar] [CrossRef]
- Wu, B.-P.; Wen, Q.; Xu, H.; Yang, Z. Insights into the impact of deep eutectic solvents on horseradish peroxidase: Activity, stability and structure. J. Mol. Catal. B Enzym. 2014, 101, 101–107. [Google Scholar] [CrossRef]
- Peng, F.; Zhao, Y.; Li, F.-Z.; Zong, M.-H.; Lou, W.-Y. The effect of deep eutectic solvents on the asymmetric hydrolysis of styrene oxide by mung bean epoxide hydrolases. Bioresour. Bioprocess. 2018, 5, 5. [Google Scholar] [CrossRef]
- Yadav, N.; Bhakuni, K.; Bisht, M.; Bahadur, I.; Venkatesu, P. Expanding the potential role of deep eutectic solvents toward facilitating the structural and thermal stability of α-chymotrypsin. ACS Sustain. Chem. Eng. 2020, 8, 10151–10160. [Google Scholar] [CrossRef]
- Gunny, A.A.N.; Arbain, D.; Nashef, E.M.; Jamal, P. Applicability evaluation of Deep Eutectic Solvents–Cellulase system for lignocellulose hydrolysis. Bioresour. Technol. 2015, 181, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Juneidi, I.; Hayyan, M.; Hashim, M.A.; Hayyan, A. Pure and aqueous deep eutectic solvents for a lipase-catalysed hydrolysis reaction. Biochem. Eng. J. 2016, 117, 129–138. [Google Scholar] [CrossRef]
- Daneshjou, S.; Khodaverdian, S.; Dabirmanesh, B.; Rahimi, F.; Daneshjoo, S.; Ghazi, F.; Khajeh, K. Improvement of chondroitinases ABCI stability in natural deep eutectic solvents. J. Mol. Liq. 2017, 227, 21–25. [Google Scholar] [CrossRef]
- Rashid, S.N.; Hayyan, A.; Hayyan, M.; Hashim, M.A.; Elgharbawy, A.A.; Sani, F.S.; Basirun, W.J.; Lee, V.S.; Alias, Y.; Mohammed, A.K.; et al. Ternary glycerol-based deep eutectic solvents: Physicochemical properties and enzymatic activity. Chem. Eng. Res. Des. 2021, 169, 77–85. [Google Scholar] [CrossRef]
- Huang, Z.-L.; Wu, B.-P.; Wen, Q.; Yang, T.-X.; Yang, Z. Deep eutectic solvents can be viable enzyme activators and stabilizers. J. Chem. Technol. Biotechnol. 2014, 89, 1975–1981. [Google Scholar] [CrossRef]
- Oh, Y.; Park, S.; Yoo, E.; Jo, S.; Hong, J.; Kim, H.J.; Kim, K.J.; Oh, K.K.; Lee, S.H. Dihydrogen-bonding deep eutectic solvents as reaction media for lipase-catalyzed transesterification. Biochem. Eng. J. 2018, 142, 34–40. [Google Scholar] [CrossRef]
- Toledo, M.L.; Pereira, M.M.; Freire, M.G.; Silva, J.B.D.A.E.; Coutinho, J.A.P.; Tavares, A.P.M. Laccase Activation in Deep Eutectic Solvents. ACS Sustain. Chem. Eng. 2019, 7, 11806–11814. [Google Scholar] [CrossRef]
- Zhao, H.; Baker, G.A.; Holmes, S. Protease activation in glycerol-based deep eutectic solvents. J. Mol. Catal. B Enzym. 2011, 72, 163–167. [Google Scholar] [CrossRef]
- Gygli, G.; Xu, X.; Pleiss, J. Meta-analysis of viscosity of aqueous deep eutectic solvents and their components. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Cao, C.; Nian, B.; Li, Y.; Wu, S.; Liu, Y. Multiple Hydrogen-Bonding Interactions Enhance the Solubility of Starch in Natural Deep Eutectic Solvents: Molecule and Macroscopic Scale Insights. J. Agric. Food Chem. 2019, 67, 12366–12373. [Google Scholar] [CrossRef]
- Patrick, W.M.; Weisner, J.; Blackburn, J. Site-directed mutagenesis of Tyr354 in Geobacillus stearothermophilus alanine racemase identifies a role in controlling substrate specificity and a possible role in the evolution of antibiotic resistance. Chembiochem 2002, 3, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Sayer, C.; Bommer, M.; Isupov, M.; Ward, J.; Littlechild, J. Crystal structure and substrate specificity of the thermophilic serine:pyruvate aminotransferase from Sulfolobus solfataricus. Acta Cryst. D Biol. Cryst. 2012, 68 Pt 7, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, I.S.; Farias, A.B.; Horta, B.A.C.; Milagre, H.M.S.; de Souza, R.O.M.A.; Bornscheuer, U.T.; Milagre, C.D.F. Computer Modeling Explains the Structural Reasons for the Difference in Reactivity of Amine Transaminases Regarding Prochiral Methylketones. Int. J. Mol. Sci. 2022, 23, 777. [Google Scholar] [CrossRef]
- Cao, J.-R.; Fan, F.-F.; Lv, C.-J.; Wang, H.-P.; Li, Y.; Hu, S.; Zhao, W.-R.; Chen, H.-B.; Huang, J.; Mei, L.-H. Improving the Thermostability and Activity of Transaminase From Aspergillus terreus by Charge-Charge Interaction. Front. Chem. 2021, 9, 224. [Google Scholar] [CrossRef]
- Smitha, K.; Pradeep, B. Application of Box-Behnken Design for the Optimization of Culture Conditions for Novel Fibrinolytic Enzyme Production by Bacillus altitudinis S-CSR 0020. J. Pure Appl. Microbiol. 2017, 11, 1447–1456. [Google Scholar] [CrossRef]
- Cheng, Q.-B.; Zhang, L.-W. Highly Efficient Enzymatic Preparation of Daidzein in Deep Eutectic Solvents. Molecules 2017, 22, 186. [Google Scholar] [CrossRef]
- Rostkowski, M.; Olsson, M.; Sondergaard, C.; Jensen, J. Graphical analysis of pH-dependent properties of proteins predicted using PROPKA. BMC Struct. Biol. 2011, 11, 6. [Google Scholar] [CrossRef]
- Sun, Y.; Kollman, P.A. Hydrophobic Solvation of Methane and Nonbond Parameters of the TIP3P Water Model. J. Comput. Chem. 1995, 16, 1164–1169. [Google Scholar] [CrossRef]
- Case, D.A.; Cheatham, T.E., III; Darden, H.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, V.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Chan, H.S.; Hu, Z. Using PyMOL as a platform for computational drug design. WIREs Comput. Mol. Sci. 2017, 7, e1298. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DES Code | Composition | Molar Ratio |
|---|---|---|
| DES1 | Choline chloride:urea | 1:1 |
| DES2 | Choline chloride:urea | 1:2 |
| DES3 | Choline chloride:urea | 1:3 |
| DES4 | Choline chloride:ethylene glycol | 1:1 |
| DES5 | Choline chloride:glycerol | 1:1 |
| DES6 | Choline chloride:1,2-propanediol | 1:1:1 |
| DES7 | Choline chloride:ethylene glycol:1,2-propanediol | 1:1:1 |
| DES8 | Choline chloride:glycerol:1,2-propanediol | 1:1:1 |
| DES9 | Choline chloride:urea:ethylene glycol | 1:1:1 |
| DES10 | Choline chloride:ethylene glycol:glycerol | 1:1:1 |
| Medium | Kmpyruvate (mM) | kcatpyruvate (min−1) | kcat/Kmpyruvate (L (min·mmol)−1) | Km1-(R)-PEA (mM) | kcat1-(R)-PEA (min−1) | kcat/Km1-(R)-PEA (L·(min·mmol)−1) |
|---|---|---|---|---|---|---|
| Buffer | 0.64 ± 0.07 | 32.98 ± 0.03 | 51.17 | 0.65 ± 0.07 | 48.16 ± 0.05 | 74.09 |
| DES4 | 0.60 ± 0.06 | 40.91 ± 0.06 | 68.62 | 0.52 ± 0.04 | 95.48 ± 0.06 | 183.61 |
| DES7 | 0.51 ± 0.03 | 45.37 ± 0.02 | 89.84 | 0.48 ± 0.02 | 102.87 ± 0.04 | 214.93 |
| ChCl | 0.97 ± 0.13 | 8.22 ± 0.012 | 8.43 | 0.83 ± 0.09 | 11.08 ± 0.09 | 13.31 |
| EG | 0.75 ± 0.08 | 19.01 ± 0.02 | 25.35 | 0.73 ± 0.09 | 26.83 ± 0.03 | 36.59 |
| PG | 0.81 ± 0.08 | 24.11 ± 0.08 | 29.88 | 0.79 ± 0.07 | 39.31 ± 0.04 | 49.50 |
| EGPG | 0.69 ± 0.07 | 26.34 ± 0.02 | 38.00 | 0.67 ± 0.03 | 43.62 ± 0.04 | 65.06 |
| Solvent | T50 °C | t½ (min) |
|---|---|---|
| Buffer | 38.0 | 10.49 |
| DES4 | 40.8 | 20.57 |
| DES7 | 42.2 | 28.36 |
| HBD | Docking Energy (kcal mol−1) | Interaction | From | To | Distance (Å) |
|---|---|---|---|---|---|
| EG | −9.5 | Hydrogen bond | EG | Lys180 | 2.9 |
| Tyr60 | EG | 2.0 | |||
| Carbon hydrogen bond | Val62 | EG | 2.9 | ||
| EG | His55 | 2.2 | |||
| PG | −11.4 | Hydrogen bond | PG | His55 | 2.1 |
| Tyr60 | PG | 2.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Masuku, M.V.; Tao, Y.; Yang, J.; Kuang, Y.; Lyu, C.; Huang, J.; Yang, S. Improved Stability and Catalytic Efficiency of ω-Transaminase in Aqueous Mixture of Deep Eutectic Solvents. Molecules 2023, 28, 3895. https://doi.org/10.3390/molecules28093895
Wang H, Masuku MV, Tao Y, Yang J, Kuang Y, Lyu C, Huang J, Yang S. Improved Stability and Catalytic Efficiency of ω-Transaminase in Aqueous Mixture of Deep Eutectic Solvents. Molecules. 2023; 28(9):3895. https://doi.org/10.3390/molecules28093895
Chicago/Turabian StyleWang, Hongpeng, Mercy Vimbai Masuku, Yachen Tao, Jiayao Yang, Yi Kuang, Changjiang Lyu, Jun Huang, and Shengxiang Yang. 2023. "Improved Stability and Catalytic Efficiency of ω-Transaminase in Aqueous Mixture of Deep Eutectic Solvents" Molecules 28, no. 9: 3895. https://doi.org/10.3390/molecules28093895
APA StyleWang, H., Masuku, M. V., Tao, Y., Yang, J., Kuang, Y., Lyu, C., Huang, J., & Yang, S. (2023). Improved Stability and Catalytic Efficiency of ω-Transaminase in Aqueous Mixture of Deep Eutectic Solvents. Molecules, 28(9), 3895. https://doi.org/10.3390/molecules28093895