
Naproxen-Derived New Compound Inhibits the NF-κB, MAPK and PI3K/Akt Signaling Pathways Synergistically with Resveratrol in RAW264.7 Cells
 , , ,
, , ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
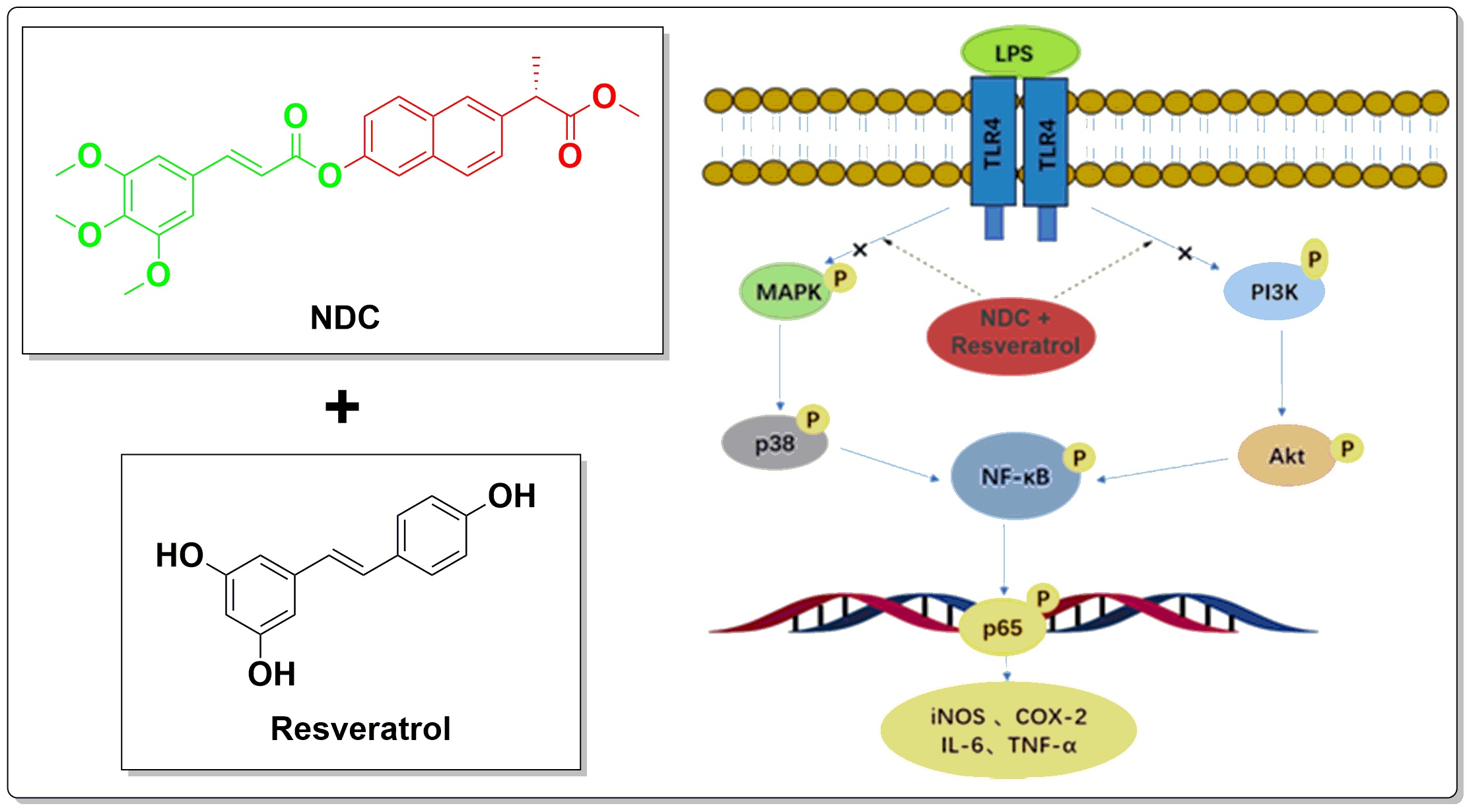
2.1. NDC Design
2.2. Effect of NDC and Resveratrol on the RAW264.7 Cell Viability
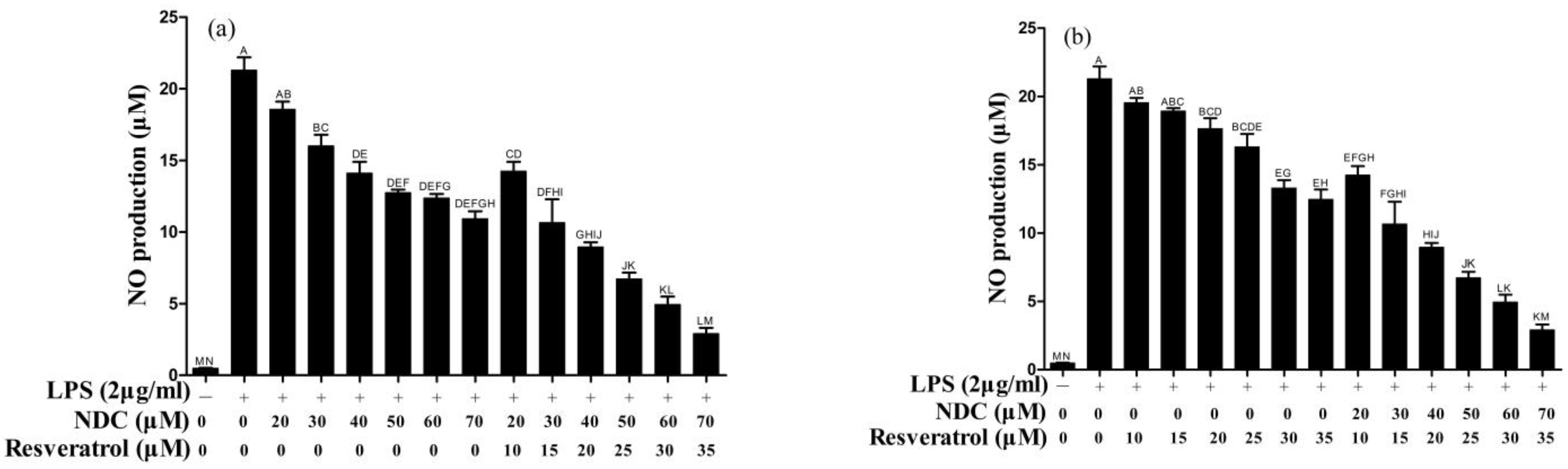
2.3. Effects of NDC and Resveratrol on LPS-Induced NO Production
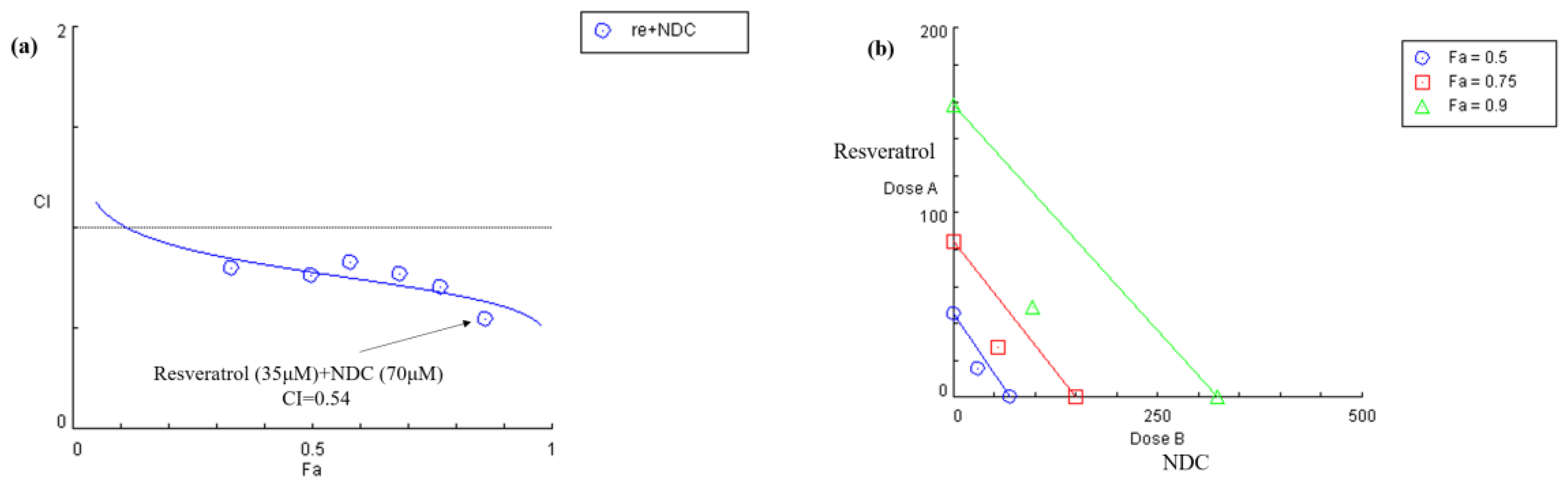
2.4. Synergistic Effect Analysis
2.5. Effects of NDC and Resveratrol on LPS-Induced IL-6 and TNF-α
2.6. Effects of NDC and Resveratrol on LPS-Induced ROS
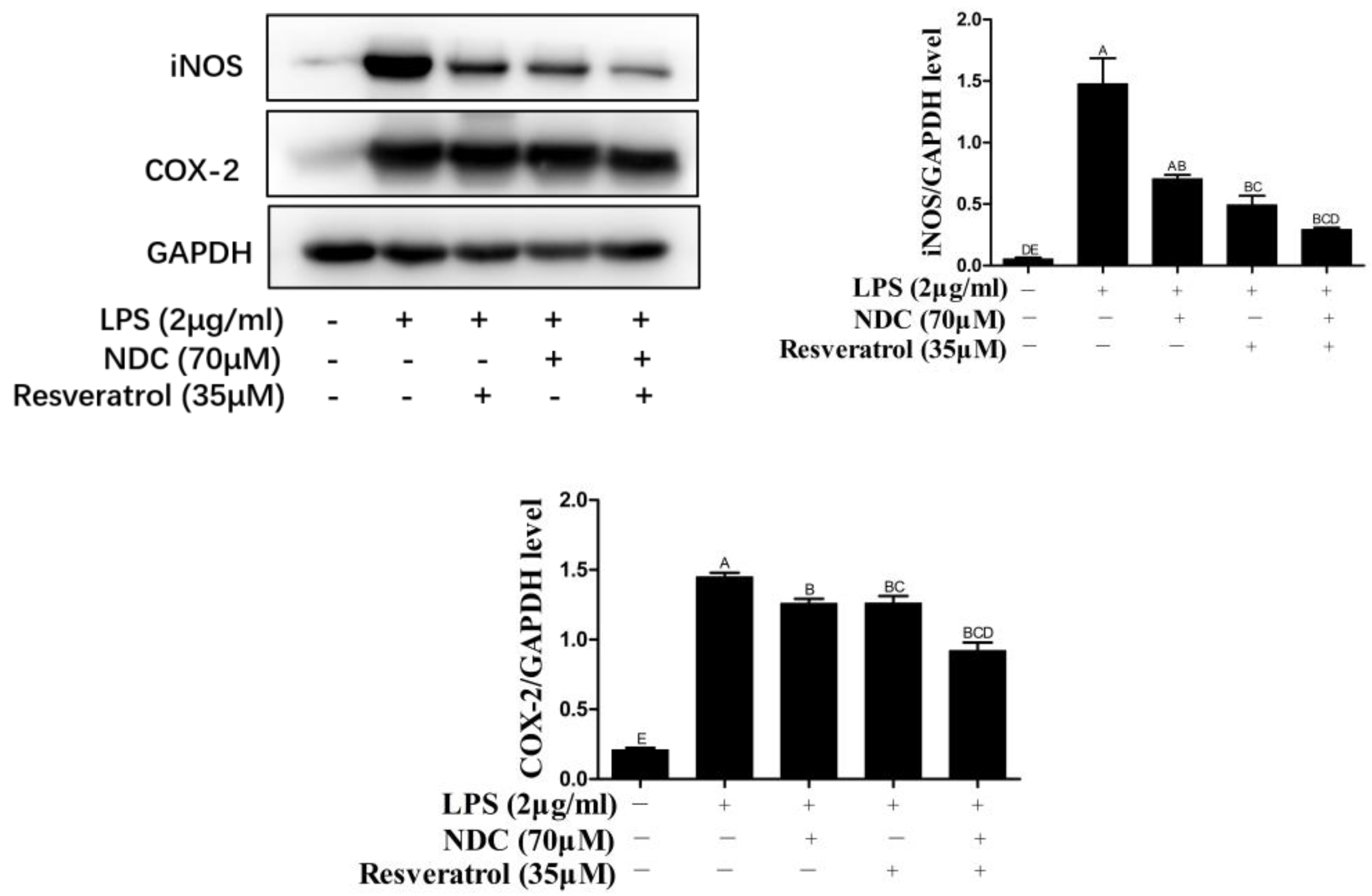
2.7. Effects of NDC and Resveratrol on iNOS and COX-2 Protein Expressions
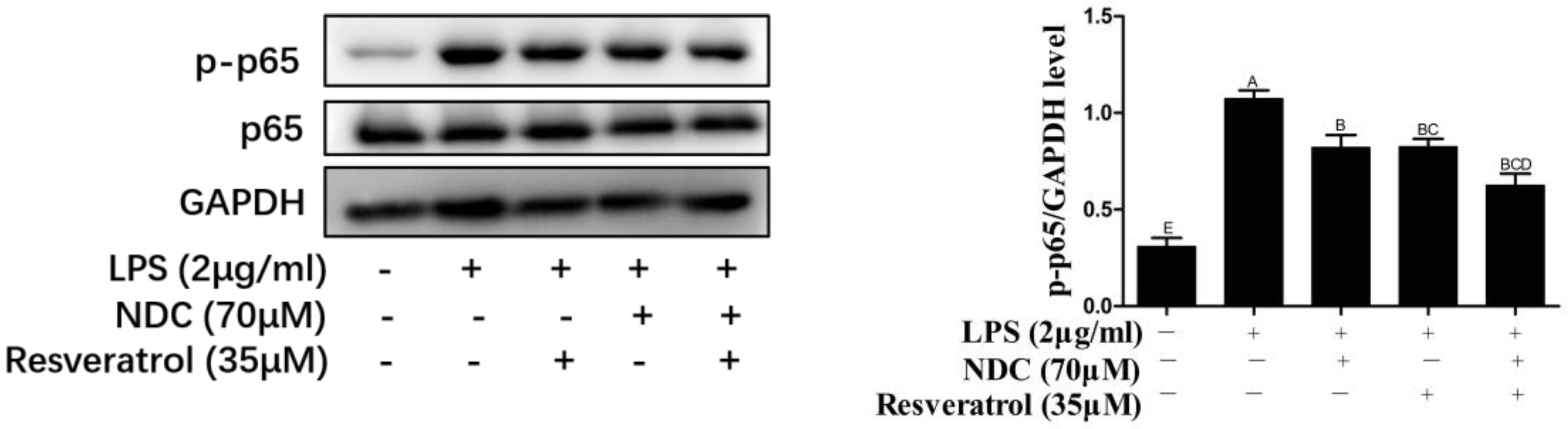
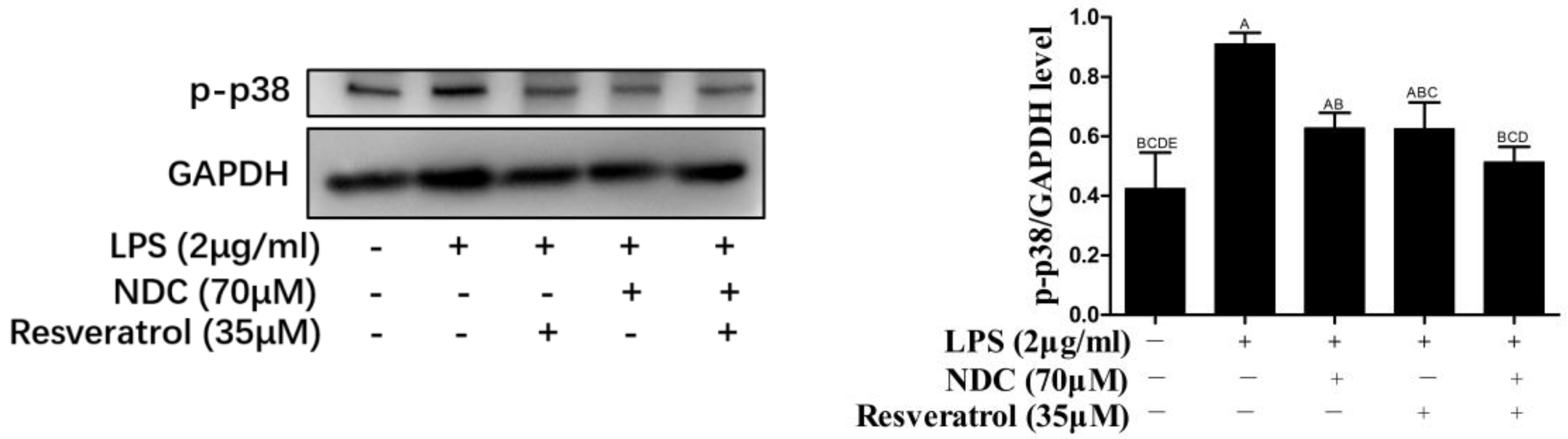
2.8. Effects of NDC and Resveratrol on the Activation of Side NF-κB, MAPK and PI3K/Akt Pathway Proteins in LPS-Induced RAW264.7 Cells
3. Materials and Methods
3.1. Materials
3.2. Synthesis of NDC
3.3. Cell Culture
3.4. Cell Viability Assay
3.5. Measurement of NO in RAW264.7 Cells
3.6. Synergistic Effect Analysis
3.7. ELISA Assay
3.8. Measurement of ROS
3.9. Protein Extraction and Western Blot Analysis
3.10. Data and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability Statement
References
- Gund, M.; Gaikwad, P.; Borhade, N.; Burhan, A.; Desai, D.C.; Sharma, A.; Dhiman, M.; Patil, M.; Sheikh, J.; Thakre, G.; et al. Gastric-sparing nitric oxide-releasable ‘true’prodrugs of aspirin and naproxen. Bioorg. Med. Chem. Lett. 2014, 24, 5587–5592. [Google Scholar] [CrossRef] [PubMed]
- Saleh-e-In, M.M.; Van Staden, J. Ethnobotany, phytochemistry and pharmacology of Arctotis arctotoides (Lf) O. Hoffm.: A review. J. Ethnopharmacol. 2018, 220, 294–320. [Google Scholar] [CrossRef]
- Ma, X.; Okyere, S.K.; Hu, L.; Wen, J.; Ren, Z.; Deng, J.; Hu, Y. Anti-inflammatory activity and mechanism of cryptochlorogenic acid from ageratina adenophora. Nutrients 2022, 14, 439. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Invest. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Shapiro, H.; Lutaty, A.; Ariel, A. Macrophages, meta-inflammation, and immuno-metabolism. Sci. World J. 2011, 11, 2509–2529. [Google Scholar] [CrossRef]
- Hoeve, M.A.; Nash, A.A.; Jackson, D.; Randall, R.E.; Dransfield, I. Influenza virus A infection of human monocyte and macrophage subpopulations reveals increased susceptibility associated with cell differentiation. PLoS ONE 2012, 7, e29443. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, L.; Xiao, M.; Wang, C.; Qin, Z. FSP1+ fibroblasts promote skin carcinogenesis by maintaining MCP-1-mediated macrophage infiltration and chronic inflammation. Am. J. Pathol. 2011, 178, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Lv, R.; Bao, Q.; Li, Y. Regulation of M1-type and M2-type macrophage polarization in RAW264. 7 cells by Galectin-9. Mol. Med. Rep. 2017, 16, 9111–9119. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, Z.; Zhou, X.; Zhao, Z.; Zhao, R.; Xu, X.; Kong, X.; Ren, J.; Yao, X.; Wen, Q.; et al. Microglia and macrophage exhibit attenuated inflammatory response and ferroptosis resistance after RSL3 stimulation via increasing Nrf2 expression. J. Neuroinflammation 2021, 18, 249. [Google Scholar] [CrossRef] [PubMed]
- Nissen, S.E.; Yeomans, N.D.; Solomon, D.H.; Lüscher, T.F.; Libby, P.; Husni, M.E.; Graham, D.Y.; Borer, J.S.; Wisniewski, L.M.; Wolski, K.E.; et al. Cardiovascular safety of celecoxib, naproxen, or ibuprofen for arthritis. N. Engl. J. Med. 2016, 375, 2519–2529. [Google Scholar] [CrossRef] [PubMed]
- Arfè, A.; Scotti, L.; Varas-Lorenzo, C.; Nicotra, F.; Zambon, A.; Kollhorst, B.; Schink, T.; Garbe, E.; Herings, R.; Straatman, H.; et al. Non-steroidal anti-inflammatory drugs and risk of heart failure in four European countries: Nested case-control study. BMJ 2016, 354, i4857. [Google Scholar] [CrossRef] [PubMed]
- Bally, M.; Dendukuri, N.; Rich, B.; Nadeau, L.; Helin-Salmivaara, A.; Garbe, E.; Brophy, J. Risk of acute myocardial infarction with NSAIDs in real world use: Bayesian meta-analysis of individual patient data. BMJ 2017, 357, j1909. [Google Scholar] [CrossRef] [PubMed]
- Szeto, C.-C.; Sugano, K.; Wang, J.-G.; Fujimoto, K.; Whittle, S.; Modi, G.K.; Chen, C.-H.; Park, J.-B.; Tam, L.-S.; Vareesangthip, K.; et al. Non-steroidal anti-inflammatory drug (NSAID) therapy in patients with hypertension, cardiovascular, renal or gastrointestinal comorbidities: Joint APAGE/APLAR/APSDE/APSH/APSN/PoA recommendations. Gut 2020, 69, 617–629. [Google Scholar] [CrossRef]
- Bjarnason, I.; Scarpignato, C.; Holmgren, E.; Olszewski, M.; Rainsford, K.D.; Lanas, A. Mechanisms of damage to the gastrointestinal tract from nonsteroidal anti-inflammatory drugs. Gastroenterology 2018, 154, 500–514. [Google Scholar] [CrossRef]
- Mohiuddin, G.; Khan, K.M.; Salar, U.; Kanwal; Lodhi, M.A.; Wadood, A.; Riaz, M.; Perveen, S. Biology-oriented drug synthesis (BIODS), in vitro urease inhibitory activity, and in silico study of S-naproxen derivatives. Bioorg. Chem. 2018, 83, 29–46. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Yue, Y.; Zhang, K.; Chen, Q.; Wang, H.; Lu, Y.; Huang, M.-T.; Zheng, X.; Du, Z. Synthesis and biological evaluation of curcumin derivatives containing NSAIDs for their anti-inflammatory activity. Bioorg. Med. Chem. Lett. 2015, 25, 3044–3051. [Google Scholar] [CrossRef]
- Liu, W.; Yue, Y.; Li, Y.; Zheng, X.; Zhang, K.; Du, Z. Inspired by magnolol: Design of NSAID-based compounds with excellent anti-inflammatory effects. MedChemComm 2015, 6, 2129–2139. [Google Scholar] [CrossRef]
- Pawełczyk, A.; Olender, D.; Sowa-Kasprzak, K.; Zaprutko, L. Hybrid compounds strategy in the synthesis of oleanolic acid skeleton-NSAID derivatives. Molecules 2016, 21, 420. [Google Scholar] [CrossRef]
- Yue, Y.; Liu, W.; Zhou, W.; Chen, M.; Huang, B.; Zhang, L.; Wang, Z.; He, Y.; Zhang, K.; Zheng, X.; et al. Synergistic inhibitory effects of naproxen in combination with magnolol on TPA-induced skin inflammation in mice. RSC Adv. 2016, 6, 38092–38099. [Google Scholar] [CrossRef]
- Kowalski, J.; Samojedny, A.; Paul, M.; Pietsz, G.; Wilczok, T. Effect of apigenin, kaempferol and resveratrol on the expression of interleukin-1beta and tumor necrosis factor-alpha genes in J774.2 macrophages. Pharmacol. Rep. 2005, 57, 390–394. [Google Scholar]
- Huang, S.; Liu, W.; Li, Y.; Zhang, K.; Zheng, X.; Wu, H.; Tang, G. Design, synthesis, and activity study of cinnamic acid derivatives as potent antineuroinflammatory agents. ACS Chem. Neurosci. 2021, 12, 419–429. [Google Scholar] [CrossRef]
- Adeniji, A.; Uddin, M.J.; Zang, T.; Tamae, D.; Wangtrakuldee, P.; Marnett, L.J.; Penning, T.M. Discovery of (R)-2-(6-methoxynaphthalen-2-yl) butanoic acid as a potent and selective aldo-keto reductase 1C3 inhibitor. J. Med. Chem. 2016, 59, 7431–7444. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, D.-L.; Xie, L.-N.; Ma, Y.-R.; Wu, P.-P.; Li, C.; Liu, W.-F.; Zhang, K.; Zhou, R.-P.; Xu, X.-T.; et al. Synergistic anti-inflammatory effects of silibinin and thymol combination on LPS-induced RAW264. 7 cells by inhibition of NF-κB and MAPK activation. Phytomedicine 2020, 1, 78. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-mediated cellular signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef]
- Song, S.B.; Tung, N.H.; Quang, T.H.; Ngan, N.T.T.; Kim, K.E.; Kim, Y.H. Inhibition of TNF-α-mediated NF-κB transcriptional activity in HepG2 cells by dammarane-type saponins from Panax ginseng leaves. J. Ginseng. Res. 2012, 36, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Tsai, P.S.; Huang, C.J. Bupivacaine inhibits COX-2 expression, PGE2, and cytokine production in endotoxin-activated macrophages. Acta Anaesthesiol. Scand. 2008, 52, 530–535. [Google Scholar] [CrossRef]
- Xu, M.; Cheng, Z.; Ding, Z.; Wang, Y.; Guo, Q.; Huang, C. Resveratrol enhances IL-4 receptor-mediated anti-inflammatory effects in spinal cord and attenuates neuropathic pain following sciatic nerve injury. Mol. Pain 2018, 14, 1744806918767549. [Google Scholar] [CrossRef]
- Doyle, S.L.; O’Neill, L.A. Toll-like receptors: From the discovery of NFκB to new insights into transcriptional regulations in innate immunity. Biochem. Pharmacol. 2006, 72, 1102–1113. [Google Scholar] [CrossRef]
- Zhang, Y.; Cardell, L.O.; Edvinsson, L.; Xu, C.B. MAPK/NF-κB-dependent upregulation of kinin receptors mediates airway hyperreactivity: A new perspective for the treatment. Pharmacol. Res. 2013, 71, 9–18. [Google Scholar] [CrossRef]
- Li, S.T.; Dai, Q.; Zhang, S.X.; Liu, Y.J.; Yu, Q.Q.; Tan, F.; Lu, S.-H.; Wang, Q.; Chen, J.-W.; Huang, H.-Q.; et al. Ulinastatin attenuates LPS-induced inflammation in mouse macrophage RAW264.7 cells by inhibiting the JNK/NF-κB signaling pathway and activating the PI3K/Akt/Nrf2 pathway. Acta Pharmacol. Sin. 2018, 39, 1294–1304. [Google Scholar] [CrossRef]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method synergy quantification method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, Y.; You, Z.; Yao, M.; Cao, Y.; Xue, X.; Chen, M.; Wu, R.; Gan, L.; Li, D.; Wu, P.; et al. Naproxen-Derived New Compound Inhibits the NF-κB, MAPK and PI3K/Akt Signaling Pathways Synergistically with Resveratrol in RAW264.7 Cells. Molecules 2023, 28, 3395. https://doi.org/10.3390/molecules28083395
Ou Y, You Z, Yao M, Cao Y, Xue X, Chen M, Wu R, Gan L, Li D, Wu P, et al. Naproxen-Derived New Compound Inhibits the NF-κB, MAPK and PI3K/Akt Signaling Pathways Synergistically with Resveratrol in RAW264.7 Cells. Molecules. 2023; 28(8):3395. https://doi.org/10.3390/molecules28083395
Chicago/Turabian StyleOu, Yi, Zonglin You, Min Yao, Yingfan Cao, Xiu Xue, Min Chen, Rihui Wu, Lishe Gan, Dongli Li, Panpan Wu, and et al. 2023. "Naproxen-Derived New Compound Inhibits the NF-κB, MAPK and PI3K/Akt Signaling Pathways Synergistically with Resveratrol in RAW264.7 Cells" Molecules 28, no. 8: 3395. https://doi.org/10.3390/molecules28083395
APA StyleOu, Y., You, Z., Yao, M., Cao, Y., Xue, X., Chen, M., Wu, R., Gan, L., Li, D., Wu, P., Xu, X., Wong, W., Wong, V. K. W., Liu, W., Ye, J., & Jin, J. (2023). Naproxen-Derived New Compound Inhibits the NF-κB, MAPK and PI3K/Akt Signaling Pathways Synergistically with Resveratrol in RAW264.7 Cells. Molecules, 28(8), 3395. https://doi.org/10.3390/molecules28083395