

Evaluating the Application Potential of a Recombinant Ganoderma Protein as Bioactive Ingredients in Cosmetics


Abstract
1. Introduction
2. Results and Discussion
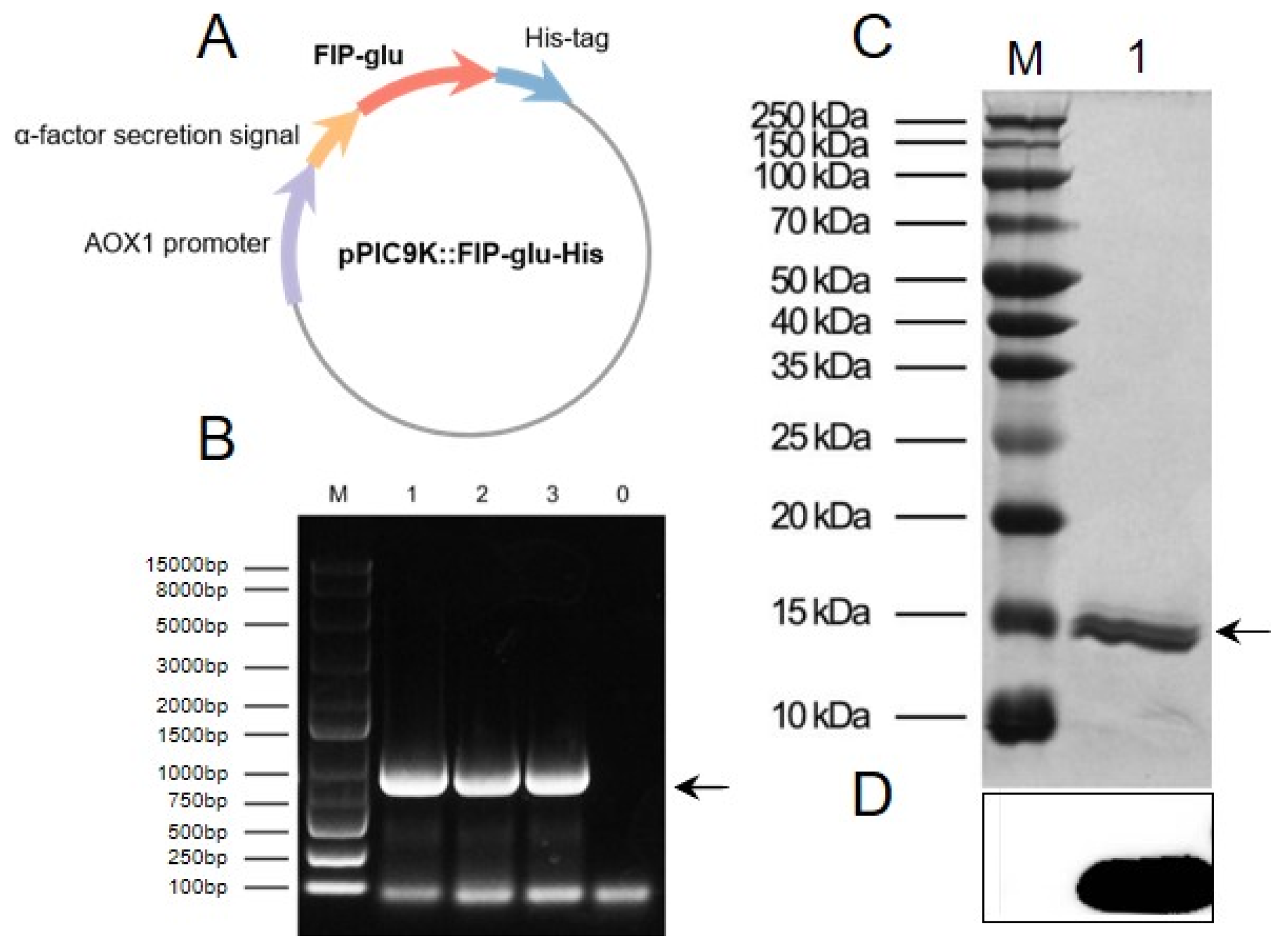
2.1. Preparation of rFIP-glu
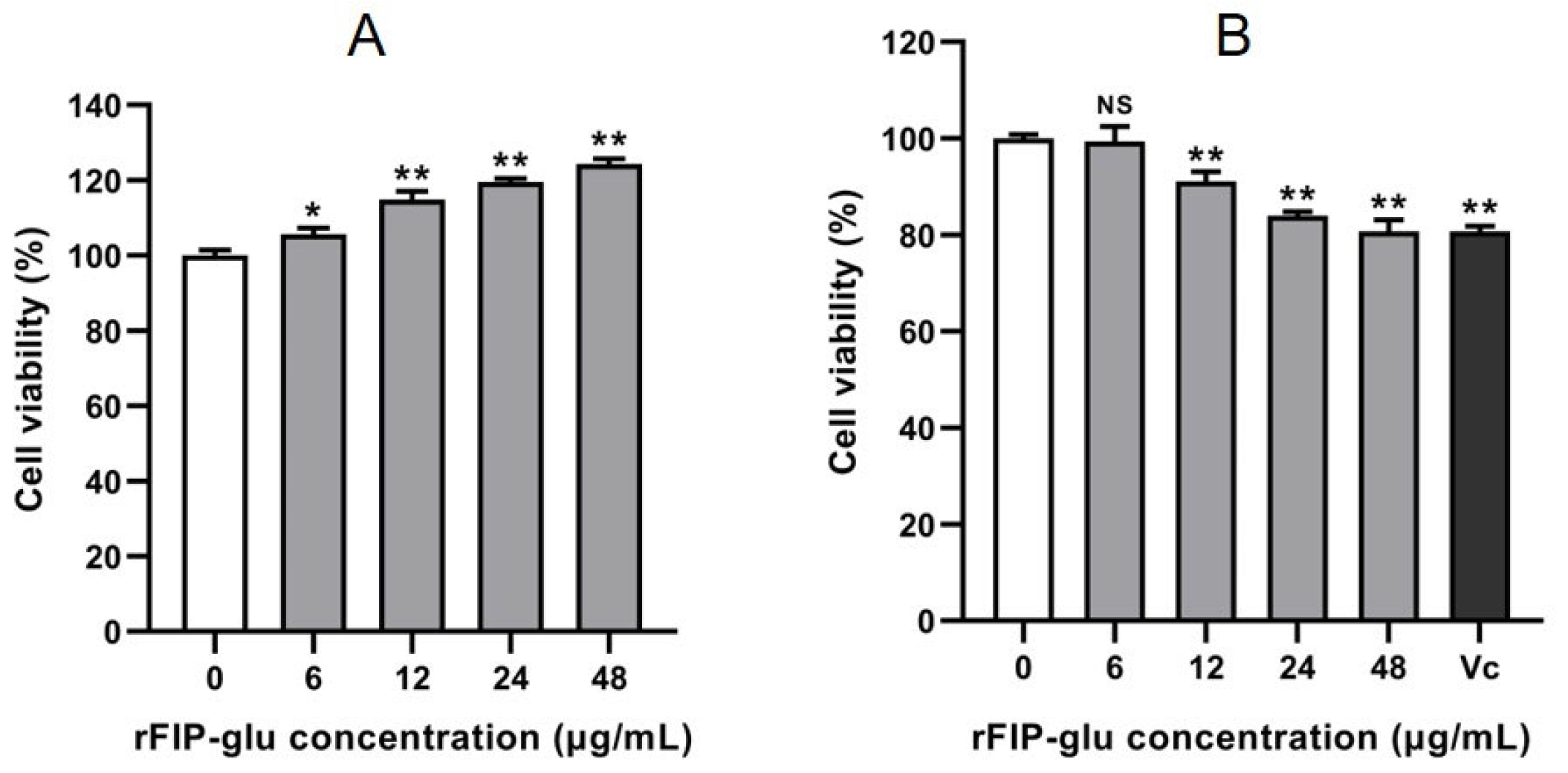
2.2. Cytotoxicity of rFIP-glu on HaCaT and B16 Cells
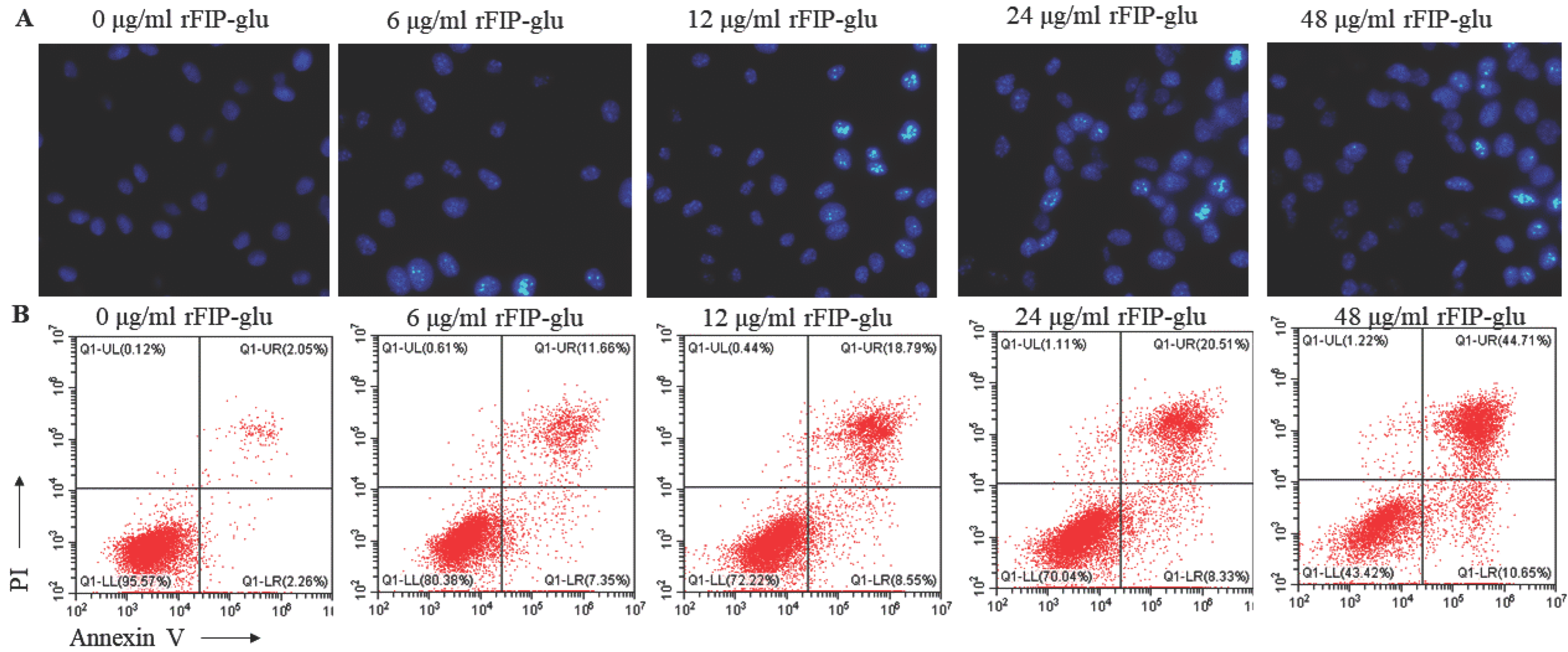
2.3. Cell Morphology and the Apoptosis of B16 Cells
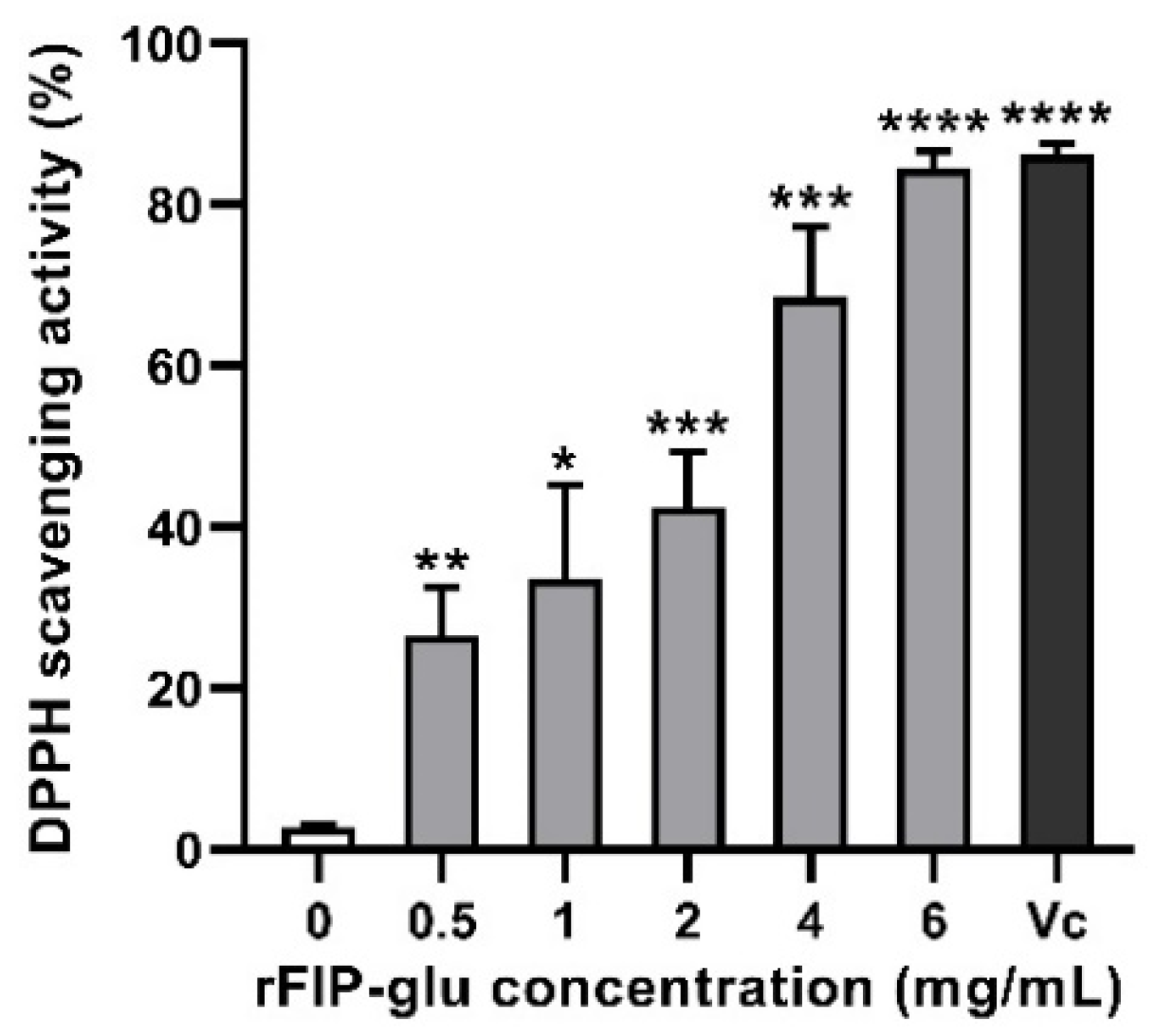
2.4. Antioxidant Activity
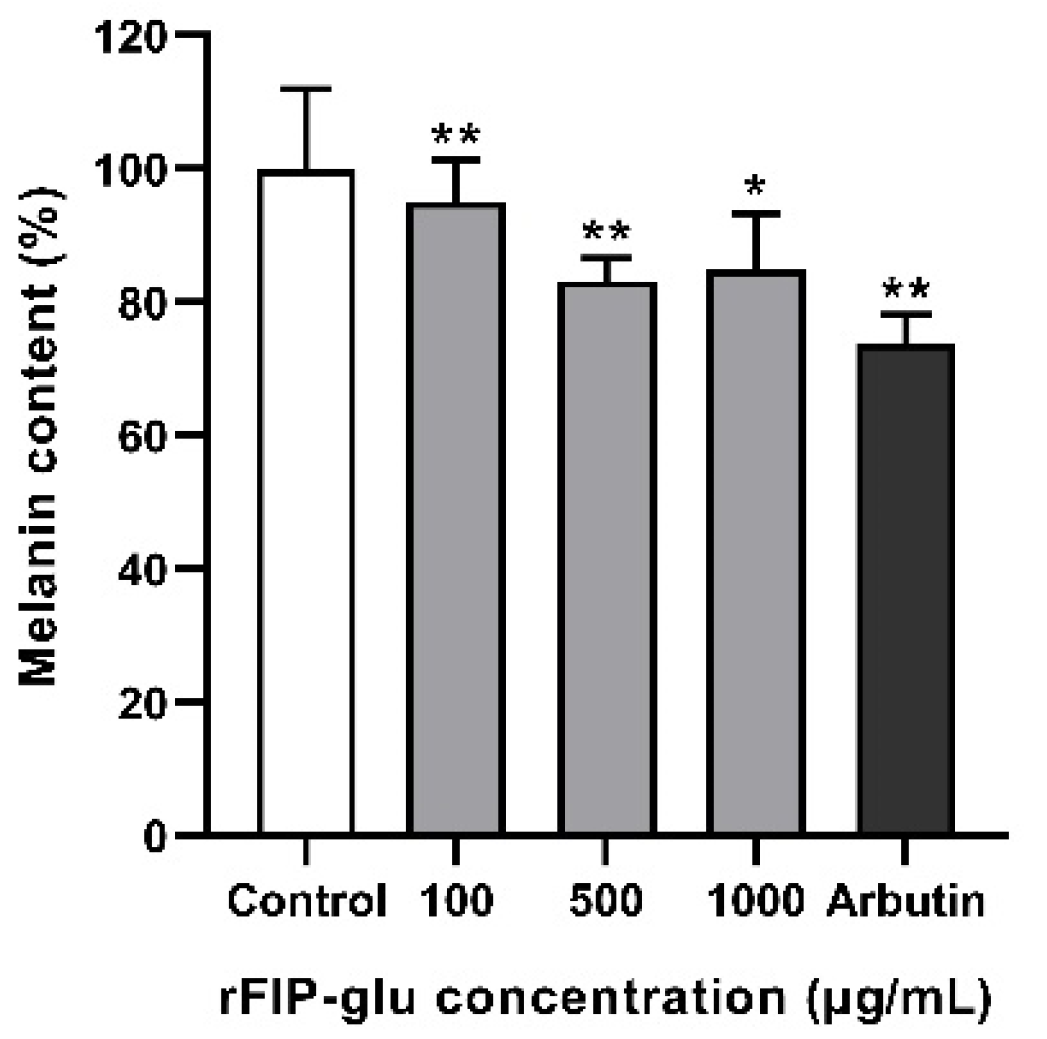
2.5. Melanin Synthesis
2.6. Tyrosinase Activity and mRNA Expression of Related Genes
3. Materials and Methods
3.1. Strain, Cells and Reagents
3.2. Preparation of rFIP-glu
3.3. Cell Culture
3.4. Cell Viability Assay
3.5. Hoechst 33258 Staining of B16 Cells
3.6. Flow Cytometry Analysis
3.7. Determination of DPPH Free Radical Scavenging Activity
3.8. Determination of Melanin Production
3.9. Determination of Tyrosinase Activity
3.10. RT-qPCR Analysis
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Zhou, X.-W.; Su, K.-Q.; Zhang, Y.-M. Applied modern biotechnology for cultivation of Ganoderma and development of their products. Appl. Microbiol. Biotechnol. 2012, 93, 941–963. [Google Scholar] [CrossRef] [PubMed]
- Kino, K.; Yamashita, A.; Yamaoka, K.; Watanabe, J.; Tanaka, S.; Ko, K.; Shimizu, K.; Tsunoo, H. Isolation and characterization of a new immunomodulatory protein, ling zhi-8 (LZ-8), from Ganoderma lucidium. J. Biol. Chem. 1989, 264, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Z.; Chang, Y.Z.; He, Z.M.; Chen, L.; Zhou, X.W. Immunomodulatory activity of Ganoderma lucidum immunomodulatory protein via PI3K/Akt and MAPK signaling pathways in RAW264. 7 cells. J. Cell. Physiol. 2019, 234, 23337–23348. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-D.; Mao, P.-W.; Shao, K.-D.; Bai, X.-H.; Zhou, X.-W. Ganoderma proteins and their potential applications in cosmetics. Appl. Microbiol. Biotechnol. 2019, 103, 9239–9250. [Google Scholar] [CrossRef] [PubMed]
- Arung, E.T.; Furuta, S.; Ishikawa, H.; Tanaka, H.; Shimizu, K. Melanin biosynthesis inhibitory and antioxidant activities of quercetin-3’-O-β-D-glucoside isolated from Allium cepa. Z. Für Nat. C 2011, 66, 209–214. [Google Scholar]
- Arung, E.T.; Syafrizal Pasedan, W.F.; Tandirogang, N.; Sukemi Allam, A.E.; Amen, Y.; Shimizu, K.; Ishikawa, H. Prenylated flavonoids as antioxidant and melanin inhibitors from stingless bee (Wallacetrigona incisa) propolis. Nat. Prod. Commun. 2020, 15, 1934578X20911272. [Google Scholar] [CrossRef]
- Baswan, S.M.; Leverett, J.; Pawelek, J. Clinical evaluation of the lightening effect of cytidine on hyperpigmented skin. J. Cosmet. Dermatol. 2019, 18, 278–285. [Google Scholar] [CrossRef]
- Pakdel, E.; Xie, W.; Wang, J.; Kashi, S.; Sharp, J.; Zhang, Q.; Varley, R.J.; Sun, L.; Wang, X. Superhydrophobic natural melanin-coated cotton with excellent UV protection and personal thermal management functionality. Chem. Eng. J. 2022, 433, 133688. [Google Scholar] [CrossRef]
- Liu, J.-K. Natural products in cosmetics. Nat. Prod. Bioprospecting 2022, 12, 40. [Google Scholar] [CrossRef]
- Juhasz, M.L.; Levin, M.K. The role of systemic treatments for skin lightening. J. Cosmet. Dermatol. 2018, 17, 1144–1157. [Google Scholar] [CrossRef]
- Li, Q.; Wang, X.; Chen, Y.; Lin, J.; Zhou, X. Cytokines expression induced by Ganoderma sinensis fungal immunomodulatory proteins (FIP-gsi) in mouse spleen cells. Appl. Biochem. Biotechnol. 2010, 162, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Mao, P.-W.; Li, L.-D.; Wang, Y.-L.; Bai, X.-H.; Zhou, X.-W. Optimization of the fermentation parameters for the production of Ganoderma lucidum immunomodulatory protein by Pichia pastoris. Prep. Biochem. Biotechnol. 2020, 50, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Stoyneva-Gärtner, M.; Uzunov, B.; Gärtner, G. Enigmatic microalgae from aeroterrestrial and extreme habitats in cosmetics: The potential of the untapped natural sources. Cosmetics 2020, 7, 27. [Google Scholar] [CrossRef]
- Levin, J.; Momin, S.B. How much do we really know about our favorite cosmeceutical ingredients? J. Clin. Aesthetic Dermatol. 2010, 3, 22. [Google Scholar]
- Thevanayagam, H.; Mohamed, S.M.; Chu, W.-L. Assessment of UVB-photoprotective and antioxidative activities of carrageenan in keratinocytes. J. Appl. Phycol. 2014, 26, 1813–1821. [Google Scholar] [CrossRef]
- Taofiq, O.; Rodrigues, F.; Barros, L.; Barreiro, M.F.; Ferreira, I.C.; Oliveira, M.B.P. Mushroom ethanolic extracts as cosmeceuticals ingredients: Safety and ex vivo skin permeation studies. Food Chem. Toxicol. 2019, 127, 228–236. [Google Scholar] [CrossRef]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tymińska, A. Skin melanocytes: Biology and development. Adv. Dermatol. Allergol. 2013, 30, 30–41. [Google Scholar] [CrossRef]
- Osborne, S.N.; Schmidt, M.A.; Derrick, K.; Harper, J.R. Epidermal micrografts produced via an automated and minimally invasive tool form at the dermal/epidermal junction and contain proliferative cells that secrete wound healing growth factors. Adv. Ski. Wound Care 2015, 28, 397. [Google Scholar] [CrossRef]
- Ali, A.; Gupta, J. Applications of stem cell therapy and adipose-derived stem cells for skin repair. Curr. Dermatol. Rep. 2022, 11, 120–130. [Google Scholar] [CrossRef]
- Merecz-Sadowska, A.; Sitarek, P.; Kowalczyk, T.; Zajdel, K.; Kucharska, E.; Zajdel, R. The modulation of melanogenesis in B16 cells upon treatment with plant extracts and isolated plant compounds. Molecules 2022, 27, 4360. [Google Scholar] [CrossRef]
- Jena, K.; Pandey, J.; Kumari, R.; Sinha, A.; Gupta, V.; Singh, G. Free radical scavenging potential of sericin obtained from various ecoraces of tasar cocoons and its cosmeceuticals implication. Int. J. Biol. Macromol. 2018, 120, 255–262. [Google Scholar] [CrossRef]
- Juang, L.J.; Gao, X.Y.; Mai, S.T.; Lee, C.H.; Lee, M.C.; Yao, C.L. Safety assessment, biological effects, and mechanisms of Myrica rubra fruit extract for anti-melanogenesis, anti-oxidation, and free radical scavenging abilities on melanoma cells. J. Cosmet. Dermatol. 2019, 18, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Shin, S.; Lee, J.-A.; Park, D.; Lee, J.; Jung, E. Inhibition of melanogenesis by Gaillardia aristata flower extract. BMC Complement. Altern. Med. 2015, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, F.; Hashizume, E.; Chan, G.P.; Kamimura, A. Skin-whitening and skin-condition-improving effects of topical oxidized glutathione: A double-blind and placebo-controlled clinical trial in healthy women. Clin. Cosmet. Investig. Dermatol. 2014, 7, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Kilala Tilaar, M.; Junardy, F.D.; Subroto, E.; Puspitosari, D. Safety and efficacy evaluation on combination of Lansium domesticum fruit extract and Hibiscus rosa-sinensis flower extract as lightening agent for cosmetic. Int. J. Pharm. Med. Biol. Sci 2018, 7, 67–70. [Google Scholar]
- Saeedi, M.; Khezri, K.; Seyed Zakaryaei, A.; Mohammadamini, H. A comprehensive review of the therapeutic potential of α-arbutin. Phytother. Res. 2021, 35, 4136–4154. [Google Scholar] [CrossRef] [PubMed]
- Wachamo, S.A.; Patel, M.H.; Varghese, P.K.; Dolinska, M.B.; Sergeev, Y.V. Characterization of Temperature-Dependent Kinetics of Oculocutaneous Albinism-Causing Mutants of Tyrosinase. Int. J. Mol. Sci. 2021, 22, 7771. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.-I.; Yun, J.-M.; Park, E.-J.; Kim, Y.-S.; Lee, Y.-M.; Lim, J.-H. Plumbagin suppresses α-MSH-induced melanogenesis in B16F10 mouse melanoma cells by inhibiting tyrosinase activity. Int. J. Mol. Sci. 2017, 18, 320. [Google Scholar] [CrossRef] [PubMed]
- Jeon, G.; Ro, H.-S.; Kim, G.-R.; Lee, H.-Y. Enhancement of Melanogenic Inhibitory Effects of the Leaf Skin Extracts of Aloe barbadensis Miller by the Fermentation Process. Fermentation 2022, 8, 580. [Google Scholar] [CrossRef]
- Boo, Y.C. Arbutin as a skin depigmenting agent with antimelanogenic and antioxidant properties. Antioxidants 2021, 10, 1129. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Guo, Y.; Zhang, Y.; Zhuang, Y. Antioxidant and anti-tyrosinase activities of phenolic extracts from rape bee pollen and inhibitory melanogenesis by cAMP/MITF/TYR pathway in B16 mouse melanoma cells. Front. Pharmacol. 2017, 8, 104. [Google Scholar] [CrossRef]
- Li, Q.-Z.; Chen, X.; Mao, P.-W.; Jin, M.-Y.; Wu, Q.; Zhou, X.-W. N-Glycosylated Ganoderma lucidum immunomodulatory protein improved anti-inflammatory activity via inhibition of the p38 MAPK pathway. Food Funct. 2021, 12, 3393–3404. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Q.-Z.; Zhou, X.-W. Immunostimulatory effects of the intracellular polysaccharides isolated from liquid culture of Ophiocordyceps sinensis (Ascomycetes) on RAW264. 7 cells via the MAPK and PI3K/Akt signaling pathways. J. Ethnopharmacol. 2021, 275, 114130. [Google Scholar] [CrossRef]
- Hong, J.-H.; Chen, H.-J.; Xiang, S.-J.; Cao, S.-W.; An, B.-C.; Ruan, S.-F.; Zhang, B.; Weng, L.-D.; Zhu, H.-X.; Liu, Q. Capsaicin reverses the inhibitory effect of licochalcone A/β-Arbutin on tyrosinase expression in b16 mouse melanoma cells. Pharmacogn. Mag. 2018, 14, 110. [Google Scholar]
- Teng, H.; Fan, X.; Lv, Q.; Zhang, Q.; Xiao, J.; Qian, Y.; Zheng, B.; Gao, H.; Gao, S.; Chen, L. Folium nelumbinis (Lotus leaf) volatile-rich fraction and its mechanisms of action against melanogenesis in B16 cells. Food Chem. 2020, 330, 127030. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Direction | Sequence (5′ → 3′) |
|---|---|---|
| TYR | Forward | CACCATGCTTTTGTGGACAG |
| TYR | Reverse | GGCTTCTGGGTAAACTTCCAA |
| TRYP-1 | Forward | CTGTGGATTATTGGGATGA |
| TRYP-1 | Reverse | GTGAGCCACCACTTTGAG |
| TRYP-2 | Forward | GCTGATTAGTCGGAACTCGA |
| TRYP-2 | Reverse | GCTGATTAGTCGGAACTCGA |
| MITF | Forward | CAAATGGCAAATACGTTACCCG |
| MITF | Reverse | CAAATGGCAAATACGTTACCCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.-J.; Liu, Y.; Yang, J.-Y.; Jin, M.-Y.; Mao, P.-W.; Zhou, X.-W. Evaluating the Application Potential of a Recombinant Ganoderma Protein as Bioactive Ingredients in Cosmetics. Molecules 2023, 28, 3272. https://doi.org/10.3390/molecules28073272
Guo Z-J, Liu Y, Yang J-Y, Jin M-Y, Mao P-W, Zhou X-W. Evaluating the Application Potential of a Recombinant Ganoderma Protein as Bioactive Ingredients in Cosmetics. Molecules. 2023; 28(7):3272. https://doi.org/10.3390/molecules28073272
Chicago/Turabian StyleGuo, Zhi-Jian, Yan Liu, Jia-Yi Yang, Meng-Yuan Jin, Pei-Wen Mao, and Xuan-Wei Zhou. 2023. "Evaluating the Application Potential of a Recombinant Ganoderma Protein as Bioactive Ingredients in Cosmetics" Molecules 28, no. 7: 3272. https://doi.org/10.3390/molecules28073272
APA StyleGuo, Z.-J., Liu, Y., Yang, J.-Y., Jin, M.-Y., Mao, P.-W., & Zhou, X.-W. (2023). Evaluating the Application Potential of a Recombinant Ganoderma Protein as Bioactive Ingredients in Cosmetics. Molecules, 28(7), 3272. https://doi.org/10.3390/molecules28073272