

Relationship between Protein Digestibility and the Proteolysis of Legume Proteins during Seed Germination



Abstract
1. Introduction
2. Plant Proteins and Their Alterations during Germination
2.1. Changes in Crude Protein Content during Germination
2.2. Changes in Polypeptide Molecular Weight Distributions during Germination
2.3. Storage Protein Changes during Germination
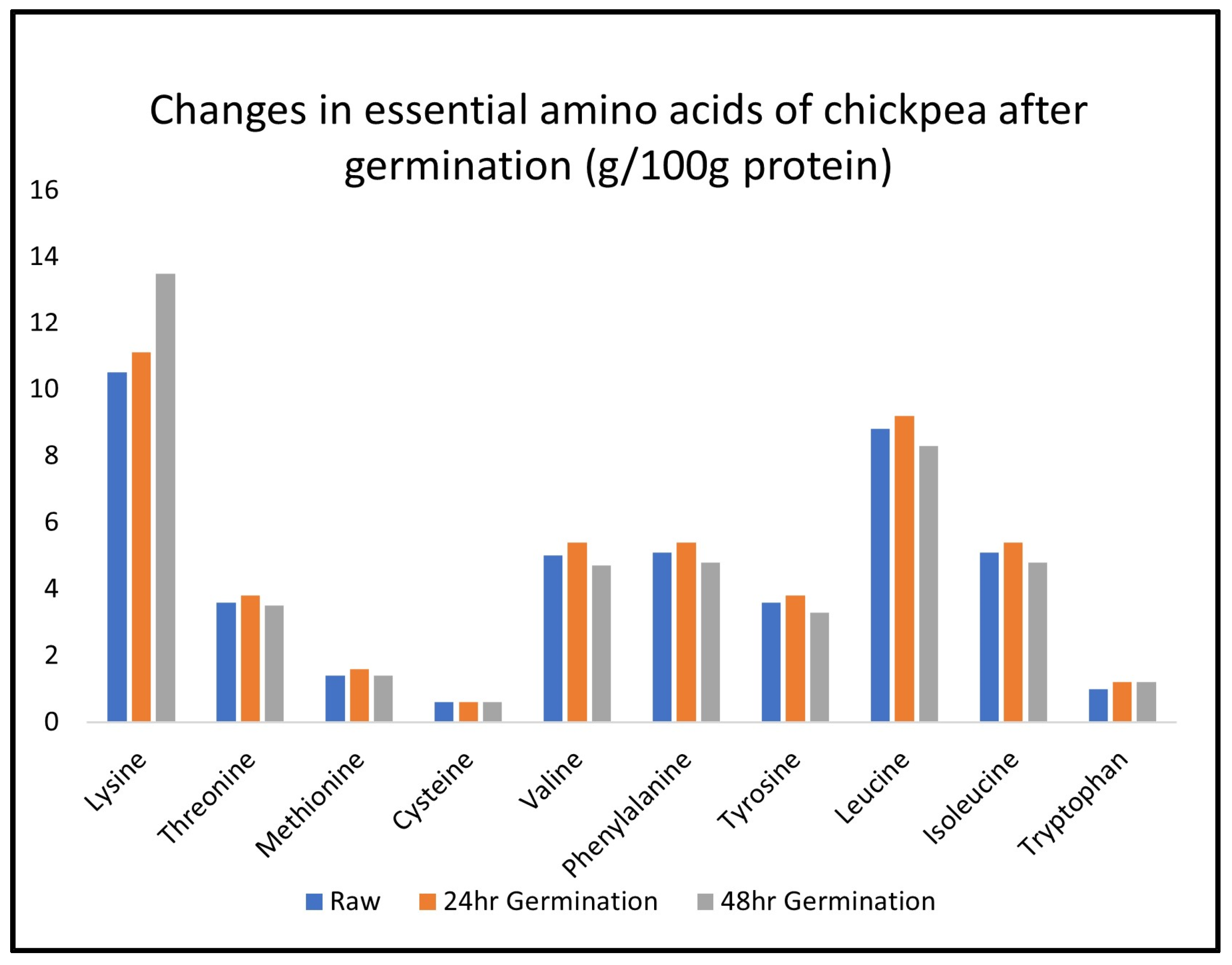
2.4. Changes in Free Amino Acids and Protein Amino Acids during Legume Germination
3. Changes in Protein Digestibility during Legume Germination
4. Changes in Proteases during Legume Germination
5. Changes in Protease Inhibitors during Legume Germination
6. Changes in Protein Allergens during Germination
7. Impact of Food Processing on Protein Digestibility
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nadathur, S.R.; Wanasundara, J.P.D.; Scanlin, L. Proteins in the Diet: Challenges in Feeding the Global Population. Sustainable Development Goals—Resource Centre. 2017. Available online: https://sdgresources.relx.com/research-book-chapters/sustainable-protein-sources-chapter-1-proteins-diet-challenges-feeding-global (accessed on 18 January 2023).
- Phillips, R.D. Starchy legumes in human nutrition, health and culture. Plant Foods Hum. Nutr. 1993, 44, 195–211. [Google Scholar] [CrossRef]
- Day, L. Proteins from land plants—Potential resources for human nutrition and food security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]
- Jones, O.G. Recent advances in the functionality of non-animal-sourced proteins contributing to their use in meat analogs. Curr. Opin. Food Sci. 2016, 7, 7–13. [Google Scholar] [CrossRef]
- Ahnen, R.T.; Jonnalagadda, S.S.; Slavin, J.L. Role of plant protein in nutrition, wellness, and health. Nutr. Rev. 2019, 77, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Gepts, P.; Beavis, W.D.; Brummer, E.C.; Shoemaker, R.C.; Stalker, H.T.; Weeden, N.F.; Young, N.D. Legumes as a Model Plant Family. Genomics for Food and Feed Report of the Cross-Legume Advances through Genomics Conference. Plant Physiol. 2005, 137, 1228–1235. [Google Scholar] [CrossRef]
- Barać, M.; Cabrilo, S.; Pešić, M.; Stanojević, S.; Pavlićević, M.; Maćej, O.; Ristić, N. Functional properties of pea (Pisum sativum, L.) protein isolates modified with chymosin. Int. J. Mol. Sci. 2011, 12, 8372–8387. [Google Scholar] [CrossRef] [PubMed]
- De Pace, C.; Delre, V.; Scarascia Mugnozza, G.T.; Maggini, F.; Cremonini, R.; Frediani, M.; Cionini, P.G. Legumin of Vicia faba major: Accumulation in developing cotyledons, purification, mRNA characterization and chromosomal location of coding genes. Theor. Appl. Genet. 1991, 83, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Joyce, B.; Fatemeh, Z.; Alison, P. Pulse proteins: Processing, characterization, functional properties and applications in food and feed. Food Res. Int. 2022, 43, 414–431. [Google Scholar] [CrossRef]
- Campos-Vega, R.; Loarca-Piña, G.; Oomah, B.D. Minor components of pulses and their potential impact on human health. Food Res. Int. 2010, 43, 461–482. [Google Scholar] [CrossRef]
- Uppal, V.; Bains, K. Effect of germination periods and hydrothermal treatments on in vitro protein and starch digestibility of germinated legumes. J. Food Sci. Technol. 2012, 49, 184–191. [Google Scholar] [CrossRef]
- Jha, U.C.; Nayyar, H.; Parida, S.K.; Deshmukh, R.; Von Wettberg, E.J.B.; Siddique, K.H.M. Ensuring Global Food Security by Improving Protein Content in Major Grain Legumes Using Breeding and ‘Omics’ Tools. Int. J. Mol. Sci. 2022, 23, 7710. [Google Scholar] [CrossRef]
- Maphosa, Y.; Jideani, V.A. The Role of Legumes in Human Nutrition. In Functional Food—Improve Health through Adequate Food; Hueda, M.C., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Bazzano, L.A.; He, J.; Ogden, L.G.; Loria, C.; Vupputuri, S.; Myers, L.; Whelton, P.K. Legume consumption and risk of coronary heart disease in US men and women: NHANES I Epidemiologic Follow-up Study. Arch. Intern Med. 2001, 161, 2573–2578. [Google Scholar] [CrossRef]
- Papandreou, C.; Becerra-Tomás, N.; Bulló, M.; Martínez-González, M.Á.; Corella, D.; Estruch, R.; Ros, E.; Arós, F.; Schroder, H.; Fitó, M.; et al. Legume consumption and risk of all-cause, cardiovascular, and cancer mortality in the PREDIMED study. Clin Nutr. 2019, 38, 348–356. [Google Scholar] [CrossRef]
- Kolonel, L.N.; Hankin, J.H.; Whittemore, A.S.; Wu, A.H.; Gallagher, R.P.; Wilkens, L.R.; John, E.M.; Howe, G.R.; Dreon, D.M.; West, D.W.; et al. Vegetables, fruits, legumes and prostate cancer: A multiethnic case-control study. Cancer Epidemiol. Biomark. Prev. 2000, 9, 795–804. [Google Scholar]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.L.; Chibbar, R.N. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Br. J. Nutr. 2012, 108, S11–S26. [Google Scholar] [CrossRef]
- Tomé, D. Digestibility issues of vegetable versus animal proteins: Protein and amino acid requirements--functional aspects. Food Nutr. Bull. 2013, 34, 272–274. [Google Scholar] [CrossRef] [PubMed]
- Bolter, C.; Jongsma, M.A. The adaptation of insects to plant protease inhibitors. J. Insect. Physiol. 1997, 43, 885–895. [Google Scholar] [CrossRef]
- Zhu-Salzman, K.; Zeng, R. Insect response to plant defensive protease inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252. [Google Scholar] [CrossRef]
- Carbonaro, M.; Maselli, P.; Nucara, A. Structural aspects of legume proteins and nutraceutical properties. Food Res. Int. 2015, 76, 19–30. [Google Scholar] [CrossRef]
- Deshpande, S.S.; Damodaran, S. Structure-Digestiblity Relationship of Legume7S Proteins. J. Food Sci. 1989, 54, 108–113. [Google Scholar] [CrossRef]
- Carbonaro, M.; Maselli, P.; Nucara, A. Relationship between digestibility and secondary structure of raw and thermally treated legume proteins: A Fourier transform infrared (FT-IR) spectroscopic study. Amino Acids 2012, 43, 911–921. [Google Scholar] [CrossRef]
- Müntz, K. Proteases and proteolytic cleavage of storage proteins in developing and germinating dicotyledonous seeds. J. Exp. Bot. 1996, 47, 605–622. [Google Scholar] [CrossRef]
- De Leo, F.; Volpicella, M.; Licciulli, F.; Liuni, S.; Gallerani, R.; Ceci, L.R. PLANT-PIs: A database for plant protease inhibitors and their genes. Nucleic Acids Res. 2002, 30, 347–348. [Google Scholar] [CrossRef]
- Mossé, J.; Baudet, J. Crude protein content and aminoacid composition of seeds: Variability and correlations. Plant Foods Hum. Nutr. 1983, 32, 225–245. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell. 1995, 7, 945–956. [Google Scholar] [CrossRef]
- Osborne, T.B. The Vegetable Proteins. Nature 1924, 114, 822. [Google Scholar] [CrossRef]
- Gemede, H.F.; Ratta, N. Antinutritional Factors in Plant Foods: Potential Health Benefits and Adverse Effects. Int. J. Nutr. Food Sci. 2014, 3, 284. [Google Scholar] [CrossRef]
- Periago, M.J.; Ros, G.; Martínez, C.; Rincón, F. Variations of non-protein nitrogen in six Spanish legumes according to the extraction method used. Food Res. Int. 1996, 29, 489–494. [Google Scholar] [CrossRef]
- Vidal-Valverde, C.; Frias, J.; Hernández, A.; Martín-Alvarez, P.; Sierra, I.; Rodríguez, C.; Blazquez, I.; Vicente, G. Assessment of nutritional compounds and antinutritional factors in pea (Pisum sativum) seeds. J. Sci. Food Agric. 2003, 83, 298–306. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Simsek, S.; Hall, C.; Rao, J.; Chen, B. Effect of germination on the chemical composition, thermal, pasting, and moisture sorption properties of flours from chickpea, lentil, and yellow pea. Food Chem. 2019, 295, 579–587. [Google Scholar] [CrossRef]
- Ferreira, C.D.; Bubolz, V.K.; da Silva, J.; Dittgen, C.L.; Ziegler, V.; De Oliveira Raphaelli, C.; de Oliveira, M. Changes in the chemical composition and bioactive compounds of chickpea (Cicer arietinum L.) fortified by germination. Lebensm. Wiss. Technol. 2019, 111, 363–369. [Google Scholar] [CrossRef]
- Dipnaik, K.; Bathere, D. Effect of soaking and sprouting on protein content and transaminase activity in pulses. Int. J. Res. Med. Sci. 2017, 5, 4271. [Google Scholar] [CrossRef][Green Version]
- Mansour, E.H. Biological and chemical evaluation of chick pea seed proteins as affected by germination, extraction and alpha-amylase treatment. Plant Foods Hum. Nutr. 1996, 49, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.W.; Zeb, A.; Mahmood, F.; Tariq, S.; Khattak, A.B.; Shah, H. Comparison of sprout quality characteristics of desi and kabuli type chickpea cultivars (Cicer arietinum L.). LWT 2007, 40, 937–945. [Google Scholar] [CrossRef]
- Kumar, Y.; Sharanagat, V.S.; Singh, L.; Mani, S. Effect of germination and roasting on the proximate composition, total phenolics, and functional properties of black chickpea (Cicer arietinum). Legume Sci. 2019, 2, e20. [Google Scholar] [CrossRef]
- Joshi, P.; Varma, K. Effect of germination and dehulling on the nutritive value of soybean. Nutr. Food Sci. 2016, 46, 595–603. [Google Scholar] [CrossRef]
- Kayembe, N.; Van Rensburg, C.J. Germination as a processing technique for soybeans in small-scale farming. S. Afr. J. Anim. Sci. 2013, 43, 7. [Google Scholar] [CrossRef]
- Kassegn, H.H.; Atsbha, T.W.; Weldeabezgi, L.T.; Yildiz, F. Effect of germination process on nutrients and phytochemicals contents of faba bean (Vicia faba L.) for weaning food preparation. Cogent Food Agric. 2018, 4, 1. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Gulewicz, P.; Frias, J.; Gulewicz, K.; Vidal-Valverde, C. Assessment of protein fractions of three cultivars of Pisum sativum L.: Effect of germination. Eur. Food Res. Technol. 2013, 226, 1465–1478. [Google Scholar] [CrossRef]
- Ul Eain Hyder Rizvi, Q.; Kumar, K.; Ahmed, N.; Yadav, A.N.; Chauhan, D.; Thakur, P.; Jan, S.; Sheikh, I. Influence of soaking and germination treatments on the nutritional, anti-nutritional, and bioactive composition of pigeon pea (Cajanus cajan L.). J. Appl. Biol. Biotechnol. 2022, 10, 317. [Google Scholar] [CrossRef]
- Mitharwal, S.; Chauhan, K. Impact of germination on the proximate composition, functional properties, and structural characteristics of black soybean (Glycine max L. Merr). J. Food Process. Preserv. 2022, 46, e17202. [Google Scholar] [CrossRef]
- Sofi, S.A.; Rafiq, S.; Singh, J.; Mir, S.A.; Sharma, S.; Bakshi, P.; McClements, D.J.; Khaneghah, A.M.; Dar, B. Impact of germination on structural, physicochemical, techno-functional, and digestion properties of desi chickpea (Cicer arietinum L.) flour. Food Chem. 2023, 405, 135011. [Google Scholar] [CrossRef] [PubMed]
- Chinma, C.E.; Abu, J.O.; Adedeji, O.E.; Aburime, L.C.; Joseph, D.G.; Agunloye, G.F.; Adebo, J.A.; Oyeyinka, S.A.; Njobeh, P.B.; Adebo, O.A. Nutritional composition, bioactivity, starch characteristics, thermal and microstructural properties of germinated pigeon pea flour. Food Biosci. 2023, 49, 101900. [Google Scholar] [CrossRef]
- Tarasevičienė, Ž.; Danilčenko, H.; Jarienė, E.; Paulauskienė, A.; Gajewski, M. Changes in some chemical components during germination of broccoli seeds. Not. Bot. Horti Agrobot. 2009, 37, 173–176. [Google Scholar]
- Moongngarm, A.; Saetung, N. Comparison of chemical compositions and bioactive compounds of germinated rough rice and brown rice. Food Chem. 2010, 122, 782–788. [Google Scholar] [CrossRef]
- Mamilla, R.; Mishra, V.K. Effect of germination on antioxidant and ACE inhibitory activities of legumes. LWT—Food Sci. Technol. 2017, 75, 51–58. [Google Scholar] [CrossRef]
- Temba, M.C.; Njobeh, P.B.; Adebo, O.A.; Olugbile, A.O.; Kayitesi, E. The role of compositing cereals with legumes to alleviate protein energy malnutrition in Africa. Int. J. Food Sci. Technol. 2016, 51, 543–554. [Google Scholar] [CrossRef]
- Rubio, L.A.; Pérez, A.; Ruiz, R.; Guzmán, M.Á.; Aranda-Olmedo, I.; Clemente, A. Characterization of pea (Pisum sativum) seed protein fractions. J. Sci. Food Agric. 2014, 94, 280–287. [Google Scholar] [CrossRef]
- Bennetau-Pelissero, C. Plant Proteins from Legumes. In Bioactive Molecules in Food; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Roy, F.; Boye, J.; Simpson, B. Bioactive proteins and peptides in pulse crops: Pea, chickpea and lentil. Food Res. Int. 2010, 43, 432–442. [Google Scholar] [CrossRef]
- Hall, C.; Hillen, C.; Robinson, J.G. Composition, Nutritional Value, and Health Benefits of Pulses. Cereal Chem. 2016, 94, 11–31. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Contreras, M.D.M.; Recio, I.; Alaiz, M.; Vioque, J. Identification and characterization of antioxidant peptides from chickpea protein hydrolysates. Food Chem. 2015, 180, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Portari, G.V.; Tavano, O.L.; da Silva, M.A.; Neves, V.A. Effect of chickpea (Cicer arietinum L.) germination on the major globulin content and in vitro digestibility. Soc. Bras. Ciência Tecnol. Aliment. 2005, 25, 29. [Google Scholar] [CrossRef]
- Villacrés, E.; Allauca, V.; Peralta, E.; Insuasti, G.; Álvarez, J.; Quelal, M. Germination, an Effective Process to Increase the Nutritional Value and Reduce Non-Nutritive Factors of Lupine Grain (Lupinus mu tabilis Sweet). Int. J. Food Sci. Nutr. Eng. 2015, 5, 163–168. [Google Scholar]
- Abd El-Moneim, A.M.R.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Protein Solubility, Digestibility and Fractionation after Germination of Sorghum Varieties. PLoS ONE 2012, 7, e31154. [Google Scholar] [CrossRef]
- Rozan, P.; Kuo, Y.H.; Lambein, F. Amino acids in seeds and seedlings of the genus Lens. Phytochemistry 2001, 58, 281–289. [Google Scholar] [CrossRef]
- Kuo, Y.-H.; Rozan, P.; Lambein, F.; Frias, J.; Vidal-Valverde, C. Effects of different germination conditions on the contents of free protein and non-protein amino acids of commercial legumes. Food Chem. 2004, 86, 537–545. [Google Scholar] [CrossRef]
- Fernandez, M.L.; Berry, J.W. Nutritional evaluation of chickpea and germinated chickpea flours. Plant Foods Hum. Nutr. 1988, 38, 127–134. [Google Scholar] [CrossRef]
- Sá, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Food processing for the improvement of plant proteins digestibility. Crit. Rev. Food Sci. Nutr. 2020, 60, 3367–3386. [Google Scholar] [CrossRef]
- Schaafsma, G. The Protein Digestibility-Corrected Amino Acid Score (PDCAAS)—A concept for describing protein quality in foods and food ingredients: A critical review. J. AOAC Int. 2005, 88, 988–994. [Google Scholar] [CrossRef]
- Büchmann, N.B. In vitro digestibility of protein from barley and other cereals. J. Sci. Food Agric. 1979, 30, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Picariello, G.; Miralles, B.; Mamone, G.; Sánchez-Rivera, L.; Recio, I.; Addeo, F.; Ferranti, P. Role of intestinal brush border peptidases in the simulated digestion of milk proteins. Mol. Nutr. Food Res. 2015, 59, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Nitride, C.; Vegarud, G.E.; Comi, I.; Devold, T.G.; Røseth, A.; Marti, A.; Iametti, S.; Mamone, G.; Picariello, G.; Alfieri, F.; et al. Effect of sprouting on the proteome of chickpea flour and on its digestibility by ex vivo gastro-duodenal digestion complemented with jejunal brush border membrane enzymes. Food Res. Int. 2022, 154, 111012. [Google Scholar] [CrossRef]
- Chitra, U.; Singh, U.; Rao, P.V. Phytic acid, in vitro protein digestibility, dietary fiber, and minerals of pulses as influenced by processing methods. Plant Foods Hum. Nutr. 1996, 49, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Ghavidel, R.; Prakash, J. The impact of germination and dehulling on nutrients, antinutrients, in vitro iron and calcium bioavailability and in vitro starch and protein digestibility of some legume seeds. LWT 2007, 40, 1292–1299. [Google Scholar] [CrossRef]
- Kalpanadevi, V.; Mohan, V.R. Effect of processing on antinutrients and in vitro protein digestibility of the underutilized legume Vigna unguiculata (L.) Walp subsp. unguiculata. LWT Food Sci. Technol. 2013, 51, 455–461. [Google Scholar] [CrossRef]
- Shimelis, E.A.; Rakshit, S.K. Effect of processing on antinutrients and in vitro protein digestibility of kidney bean (Phaseolus vulgaris L.) varieties grown in East Africa. Food Chem. 2007, 11, 43–49. [Google Scholar] [CrossRef]
- Setia, R.; Dai, Z.; Nickerson, M.T.; Sopiwnyk, E.; Malcolmson, L.; Ai, Y. Impacts of short-term germination on the chemical compositions, technological characteristics and nutritional quality of yellow pea and faba bean flours. Food Res Int. 2019, 122, 263–272. [Google Scholar] [CrossRef]
- Muñoz-Llandes, C.B.; Guzmán-Ortiz f, A.; Román-Gutiérrez, A.D.; Palma-Rodríguez, H.M.; Castro-Rosas, J.; Hernández-Sánchez, H.; Zamora-Natera, J.F.; Vargas-Torres, A. Effect of Germination Time on Protein Subunits of Lupinus angustifolius L. And Its Influence on Functional Properties and Protein Digestibility. Food Sci. Technol 2022, 42, 821. [Google Scholar] [CrossRef]
- Hu, M.; Du, X.; Liu, G.; Zhang, S.; Wu, H.; Li, Y. Germination improves the functional properties of soybean and enhances soymilk quality. Int. J. Food Sci. Technol. 2021, 57, 3892–3902. [Google Scholar] [CrossRef]
- Onyango, C.; Ochanda, S.O.; Mwasaru, M.; Ochieng, J.K.; Mathooko, F.; Kinyuru, J. Effects of Malting and Fermentation on Anti-Nutrient Reduction and Protein Digestibility of Red Sorghum, White Sorghum and Pearl Millet. J. Food Res. 2013, 2, 41. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Bateman, A. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2012, 40, D343–D350. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J. Evolutionary families of peptidases. Biochem. J. 1993, 290 Pt 1, 205–218. [Google Scholar] [CrossRef]
- Ghavidel, R.; Prakash, J.; Davoodi, M. Assessment of enzymatic changes in some legume seeds during germination. Agro Food Ind. Hi Tech. 2011, 22, 45–47. [Google Scholar]
- Chrispeels, M.J.; Boulter, D. Control of storage protein metabolism in the cotyledons of germinating mung beans: Role of endopeptidase. Plant Physiol. 1975, 55, 1031–1037. [Google Scholar] [CrossRef]
- Srikanth, S.; Chen, Z. Plant Protease Inhibitors in Therapeutics-Focus on Cancer Therapy. Front Pharmacol. 2016, 7, 470. [Google Scholar] [CrossRef]
- Hellinger, R.; Gruber, C.W. Peptide-based protease inhibitors from plants. Drug Discov. Today 2019, 24, 1877–1889. [Google Scholar] [CrossRef] [PubMed]
- Rustgi, S.; Boex-Fontvieille, E.; Reinbothe, C.; von Wettstein, D.; Reinbothe, S. The complex world of plant protease inhibitors: Insights into a Kunitz-type cysteine protease inhibitor of Arabidopsis thaliana. Commun. Integr. Biol. 2017, 11, e1368599. [Google Scholar] [CrossRef]
- Lawrence, P.K.; Koundal, K.R. Plant protease inhibitors in control of phytophagous insects. Electron. J. Biotechnol. 2002, 5, 5–6. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Park, S.-C.; Hwang, I.; Cheong, H.; Nah, J.-W.; Hahm, K.-S.; Park, Y. Protease Inhibitors from Plants with Antimicrobial Activity. Int. J. Mol. Sci. 2009, 10, 2860–2872. [Google Scholar] [CrossRef] [PubMed]
- Magee, P.J.; Owusu-Apenten, R.; McCann, M.J.; Gill, C.I.; Rowland, I.R. Chickpea (Cicer arietinum) and other plant-derived protease inhibitor concentrates inhibit breast and prostate cancer cell proliferation in vitro. Nutr. Cancer 2012, 64, 741–748. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process. Nutr. 2020, 2, 6. [Google Scholar] [CrossRef]
- Caccialupi, P.; Ceci, L.R.; Siciliano, R.A.; Pignone, D.; Clemente, A.; Sonnante, G. Bowman-Birk inhibitors in lentil: Heterologous expression, functional characterisation and anti-proliferative properties in human colon cancer cells. Food Chem. 2010, 120, 1058–1066. [Google Scholar] [CrossRef]
- Clemente, A.; Gee, J.M.; Johnson, I.T.; Mackenzie, D.A.; Domoney, C. Pea (Pisum sativum L.) protease inhibitors from the Bowman-Birk class influence the growth of human colorectal adenocarcinoma HT29 cells in vitro. J. Agric. Food Chem. 2005, 53, 8979–8986. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sehgal, S. Effect of domestic processing, cooking and germination on the trypsin inhibitor activity and tannin content of faba bean (Vicia faba). Plant Foods Hum. Nutr. 1992, 42, 127–133. [Google Scholar] [CrossRef]
- Frias, J.; Diaz-Pollan, C.; Hedley, C.L.; Vidal-Valverde, C. Evolution of Trypsin Inhibitor Activity during Germination of Lentils. J. Agric. Food Chem. 1995, 43, 2231–2234. [Google Scholar] [CrossRef]
- Pusztai, A. Metabolism of trypsin-inhibitory proteins in the germinating seeds of kidney bean (Phaseolus vulgaris). Planta 1972, 107, 121–129. [Google Scholar] [CrossRef]
- Nielsen, S.; Liener, I. Effect of Germination on Trypsin Inhibitor and Hemagglutinating Activities in Phaseolus vulgaris. J. Food Sci. 1988, 53, 298–299. [Google Scholar] [CrossRef]
- Gupta, Y.P. Anti-nutritional and toxic factors in food legumes: A review. Plant Foods Hum. Nutr. 1987, 37, 201–228. [Google Scholar] [CrossRef]
- Holland, C.; Ryden, P.; Edwards, C.H.; Grundy, M.M.-L. Plant Cell Walls: Impact on Nutrient Bioaccessibility and Digestibility. Foods 2020, 9, 201. [Google Scholar] [CrossRef]
- Dallas, D.C.; Sanctuary, M.R.; Qu, Y.; Khajavi, S.H.; Van Zandt, A.E.; Dyandra, M.; Frese, S.A.; Barile, D.; German, J.B. Personalizing protein nourishment. Crit. Rev. Food Sci. Nutr. 2017, 57, 3313–3331. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.; Mosenthin, R.; Zimmermann, B.; Greiner, R.; Roth, S. Distribution of phytase activity, total phosphorus and phytate phosphorus in legume seeds, cereals and cereal by-products as influenced by harvest year and cultivar. Anim. Feed Sci. Technol. 2007, 133, 320–334. [Google Scholar] [CrossRef]
- Liener, I.E. Implications of antinutritional components in soybean foods. Crit. Rev. Food Sci. Nutr. 1994, 34, 31–67. [Google Scholar] [CrossRef] [PubMed]
- el-Adawy, T.A. Nutritional composition and antinutritional factors of chickpeas (Cicer arietinum L.) undergoing different cooking methods and germination. Plant Foods Hum Nutr. 2002, 57, 83–97. [Google Scholar] [CrossRef]
- Pal, R.S.; Bhartiya, A.; ArunKumar, R.; Kant, L.; Aditya, J.P.; Bisht, J.K. Impact of dehulling and germination on nutrients, antinutrients, and antioxidant properties in horsegram. J. Food Sci. Technol. 2016, 53, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Aguirre, A.; Marzo, F. Effects of extrusion and traditional processing methods on antinutrients and in vitro digestibility of protein and starch in faba and kidney beans. Food Chem. 2000, 68, 159–165. [Google Scholar] [CrossRef]
- Wu, X.; Mei, T.; Zhu, Y.; Duan, H.; Ramaswamy, H.S.; Bai, W.; Wang, C. The influence of high pressure processing and germination on anti-nutrients contents, in vitro amino acid release and mineral digestibility of soybeans. J. Food Compos. Anal. 2023, 115, 104953. [Google Scholar] [CrossRef]
- Verma, A.K.; Kumar, S.; Das, M.; Dwivedi, P.D. A comprehensive review of legume allergy. Clin. Rev. Allergy Immunol. 2013, 45, 30–46. [Google Scholar] [CrossRef]
- Maria John, K.M.; Khan, F.; Luthria, D.L.; Garrett, W.; Natarajan, S. Proteomic analysis of anti-nutritional factors (ANF’s) in soybean seeds as affected by environmental and genetic factors. Food Chem. 2017, 218, 321–329. [Google Scholar] [CrossRef]
- Hugh, S.; Liam, O.; Burks, A.; Marshall, P.; Gideon, L.; Cezmi, A. Mechanisms of food allergy. J. Allergy Clin. Immunol. 2018, 141, 11–19. [Google Scholar] [CrossRef]
- Astwood, J.D.; Leach, J.N.; Fuchs, R.L. Stability of food allergens to digestion in vitro. Nat Biotechnol. 1996, 14, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Troszyńska, A.; Szymkiewicz, A.; Wołejszo, A. The effects of germination on the sensory quality and immunoreactive properties of pea (Pisum sativum L.) and soybean (Glycine max). J. Food Qual. 2007, 30, 1083–1100. [Google Scholar] [CrossRef]
- Wu, Y.M.; Guan, R.X.; Liu, Z.X.; Li, R.Z.; Chang, R.Z.; Qiu, L.J. Synthesis and degradation of the major allergens in developing and germinating soybean seed. J. Integr. Plant Biol. 2012, 54, 4–14. [Google Scholar] [CrossRef]
- Boye, J.; Wijesinha-Bettoni, R.; Burlingame, B. Protein quality evaluation twenty years after the introduction of the protein digestibility corrected amino acid score method. Br. J. Nutr. 2012, 108 (Suppl. 2), S183–S211. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; del Refugio Rocha-Pizaña, M.; García-Lara, S.; López-Castillo, L.M.; Serna-Saldívar, S.O. Effect of thermal processing and reducing agents on trypsin inhibitor activity and functional properties of soybean and chickpea protein concentrates. LWT 2018, 98, 629–634. [Google Scholar] [CrossRef]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of Fermentation in Improving Nutritional Quality of Soybean Meal—A Review. Asian-Australas. J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.J.; Lee, C.H.; Kim, S.W. Aspergillus oryzae GB-107 fermentation improves nutritional quality of food soybeans and feed soybean meals. J. Med. Food. 2004, 7, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.S.; Frias, J.; Martinez-Villaluenga, C.; Vidal-Valdeverde, C.; de Mejia, E.G. Immunoreactivity reduction of soybean meal by fermentation, effect on amino acid composition and antigenicity of commercial soy products. Food Chem. 2008, 108, 571–581. [Google Scholar] [CrossRef]
- Pi, X.; Wan, Y.; Yang, Y.; Li, R.; Wu, X.; Xie, M.; Li, X.; Fu, G. Research progress in peanut allergens and their allergenicity reduction. Trends Food Sci. Technol. 2019, 93, 212–220. [Google Scholar] [CrossRef]
- Ketnawa, S.; Ogawa, Y. Evaluation of protein digestibility of fermented soybeans and changes in biochemical characteristics of digested fractions. J. Funct. Foods 2019, 52, 640–647. [Google Scholar] [CrossRef]
- Ojokoh, A.; Yimin, W. Effect of Fermentation on Chemical Composition and Nutritional Quality of Extruded and Fermented Soya Products. Int. J. Food Eng. 2011, 7, 1857. [Google Scholar] [CrossRef]
- Shewry, P.R.; Casey, R. (Eds.) Seed Proteins; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Avezum, L.; Rondet, E.; Mestres, C.; Achir, N.; Madode, Y.; Gibert, O.; Lefevre, C.; Hemery, Y.; Verdeil, J.-L.; Rajjou, L. Improving the nutritional quality of pulses via germination. Food Rev. Int. 2022, 1–34. [Google Scholar] [CrossRef]
- Grasso, N.; Lynch, N.L.; Arendt, E.K.; O’Mahony, J.A. Chickpea protein ingredients: A review of composition, functionality, and applications. Compr. Rev. Food Sci. Food Saf. 2022, 21, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Smolikova, G.; Gorbach, D.; Lukasheva, E.; Mavropolo-Stolyarenko, G.; Bilova, T.; Soboleva, A.; Tsarev, A.; Romanovskaya, E.; Podolskaya, E.; Zhukov, V.; et al. Bringing New Methods to the Seed Proteomics Platform: Challenges and Perspectives. Int. J. Mol. Sci. 2020, 21, 9162. [Google Scholar] [CrossRef] [PubMed]
- Farrokhi, N.; Whitelegge, J.P.; Brusslan, J.A. Plant peptides and peptidomics. Plant Biotechnol. J. 2008, 6, 105–134. [Google Scholar] [CrossRef] [PubMed]
- Bessada, S.M.; Barreira, J.C.; Oliveira, M.B.P. Pulses and food security: Dietary protein, digestibility, bioactive and functional properties. Trends Food Sci. Technol. 2019, 93, 53–68. [Google Scholar] [CrossRef]
- Hemalatha, S.; Platel, K.; Srinivasan, K. Influence of germination and fermentation on bioaccessibility of zinc and iron from food grains. Eur. J. Clin. Nutr. 2006, 61, 342–348. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sprout Species | Pre-Germination | Post-Germination | References |
|---|---|---|---|
| Chickpea | 24.4% | 27.7% | Xu et al. (2019) [33] |
| 18.4% | 24.6% | Ferreira et al. (2019) [34] | |
| 32 ± 1.8% * | 48 ± 0.5% * | Dipnaik and Bathere (2017) [35] | |
| 22.3% | 24.1% | Mansour (1987) [36] | |
| ~20% | 23.9% | Khalil et al. (2007) [37] | |
| 20.3% | 23.6% | Uppal et al. (2012) [11] | |
| Chickpea desi | 14.8 ± 0.6% | 15.9 ± 0.4% | Kumar et al. (2019) [38] |
| Chickpea desi | ~21% | 24.1% | Khalil et al. (2007) [37] |
| Mungbean | 22.5 ± 0.9% | 36 ± 0.5% | Dipnaik and Bathere (2017) [35] |
| 22.3% | 24.9% | Uppal et al. (2012) [11] | |
| Cowpea | 30 ± 1.07% | 40 ± 0.5% | Dipnaik and Bathere (2017) [35] |
| 22.5% | 24.9% | Uppal et al. (2012) [11] | |
| Moth bean | 30 ± 1.0% | 40 ± 12.3% | Dipnaik and Bathere (2017) [35] |
| Soybean | 40.2 ± 0.3% | 46.3 ± 0.4% | Joshi and Varma (2016) [39] |
| 39.1% | 45.1% | Kayembe et al. (2013) [40] | |
| Faba bean | 26.4% | 30.6% | Kassegn et al. (2018) [41] |
| Pea | |||
| var ucero | 25.4 ± 0.1% | 27.0 ± 0.1% | Martinez-Villaluenga et al. (2008) [42] |
| var ramrod | 21.1 ± 0.0% | 22.7 ± 0.1% | Martınez-Villaluenga et al. (2008) [42] |
| var agra | 22.9 ± 0.1% | 22.7 ± 0.1% | Martınez-Villaluenga et al. (2008) [42] |
| Black gram | 20 ± 1.5% | 36 ± 1.54% | Dipnaik and Bathere (2017) [35] |
| Pigeon Pea | 19.53 ± 0.02% | 22.54 ± 0.02% | Rizvi et al. (2022) [43] |
| Black soybean flour | 39.46 ± 0.08% | 43.30 ± 0.05% | Mitharwal and Chauhan (2022) [44] |
| Chickpea flour | 21.9 ± 0.2% | 24.0 ± 0.2% | Sofi et al. (2023) [45] |
| Pigeon pea flour | 22.71 ± 0.15% | 26.72 ± 0.11% | Chinma et al. (2022) [46] |
| (Broccoli) | 26.1% | 29.8% | Taraseviciene et al. (2009) [47] |
| (Brown Rice) | 6.9 ± 0.0% | 8.9 ± 0.2% | Moongngarm et al. (2010) [48] |
| Sprout Species | Pre- Germination | Post- Germination | References |
|---|---|---|---|
| Chickpea (major globulin) | 45.85% | 37.08% | Portari et al. (2005) [56] |
| Pea (albumin and globulin) | Martinez-Villaluenga et al. (2008) [42] | ||
| var ucero | 28.95% | 24.9% | |
| var ramrod | 26.71% | 22.69% | |
| var agra | 26.67% | 25.03% | |
| Lupin | 12.81% | 15.7% | Villacrés et al. (2015) [57] |
| Sweet lupin (albumin and globulin) | Gulewicz et al. (2008) [42] | ||
| Lupinus luteus cv. 4486 | 36.89% | 39.45% | |
| Lupinus luteus cv. 4492 | 39.63% | 35.91% | |
| Lupinus angustifolius cv.troll | 34.9% | 35.2% | |
| Lupinus angustifolius cv.zapato | 35.4% | 29.48% | |
| (Sorghum) | 25% | 28% | Afify et al. (2012) [58] |
| Free Amino Acids | Pre- Germination | Post- Germination | All Amino Acids | Pre- Germination | Post- Germination |
|---|---|---|---|---|---|
| Arg | 0.1 | 0.94 | Arg | 10.61 | 12.11 |
| His | 0.22 | 0.68 | His | 8.74 | 10.79 |
| Ile | 0 | 2.06 | Ile | 6.26 | 11.44 |
| Leu | 0 | 2.05 | Leu | 10.64 | 17.1 |
| Lys | 0 | 0.93 | Lys | 4.54 | 16.99 |
| Met | 0 | 0.26 | Met | 1.49 | 1.02 |
| Cys | 0 | 0 | Cys | 0.4 | 0 |
| Phe | 0 | 2.32 | Phe | 6.7 | 11.56 |
| Tyr | 0 | 1.1 | Tyr | 6.34 | 7.7 |
| Pro | 0.17 | 3.24 | Pro | 11.11 | 10.84 |
| Ser | 0 | 2.64 | Ser | 11.38 | 15.54 |
| Thr | 0.03 | 1.18 | Thr | 5.57 | 7.14 |
| Val | 0 | 2.83 | Val | 8.54 | 13.23 |
| Trp | 0 | 0.52 | Trp | 0 | 0 |
| Ala | 0.45 | 3.21 | Ala | 20.42 | 36.76 |
| Asp | 0.18 | 0.32 | ASX * | 10.96 | 41.39 |
| Asn | 0.51 | 18.96 | |||
| Glu | 0.48 | 3.15 | GLX * | 26.55 | 34.12 |
| Gln | 0 | 1 | |||
| Gly | 0.05 | 1.23 | Gly | 9.77 | 10.77 |
| Total | 2.19 | 48.62 | 160.02 | 258.50 |
| Amino Acids | Beans (Phaseolus Vulgaris) | Lentils (Lens Culinaris) | Pea (Pisum Sativum) | |||
|---|---|---|---|---|---|---|
| Germination | Pre- | Post- | Pre- | Post- | Pre- | Post- |
| Alanine | 2.93 | 4.4 | 0.5 | 0.8 | 0.25 | 2.58 |
| Arginine | 13.2 | 2.95 | 0.6 | 1.2 | 3.4 | 3.8 |
| Asparagine | 5.9 | 8.0 | 0.88 | 28.7 | 0.8 | 23.0 |
| Aspartic acid | 4.0 | 2.5 | 0.70 | 1.27 | 1.6 | 5.4 |
| Glutamic acid | 11.2 | 4.09 | 1.34 | 3.93 | 2.0 | 3.5 |
| Glutamine | 0.0 | 1.27 | 0.0 | 1.06 | 0.0 | 2.38 |
| Glycine | 0.50 | 0.09 | 0.08 | 0.35 | 0.07 | 0.30 |
| Histidine | 0.45 | 0 | 0.04 | 0.80 | 0.14 | 0.0 |
| Isoleucine | 0.8 | 0.8 | 0.0 | 0.99 | 0.0 | 0.58 |
| Leucine | 0.6 | 1.2 | 0.0 | 0.61 | 0.01 | 0.52 |
| Lysine | 0.11 | 0.53 | 0.0 | 1.05 | 0.08 | 0.75 |
| Methionine | 0.0 | 0.382 | 0.0 | 0.0 | 0.0 | 0.0 |
| Phenylalanine | 1.0 | 0.8 | 0.0 | 1.04 | 0.15 | 1.16 |
| Proline | 0.8 | 0.8 | 0.23 | 2.91 | 0.53 | 2.23 |
| Serine | 0.1 | 2.0 | 0.0 | 2.43 | 0.02 | 1.51 |
| Threonine | 0.2 | 1.0 | 0.042 | 2.12 | 0.0 | 0.36 |
| Tryptophan | 0.68 | 0.33 | 0.0 | 0.27 | 0.10 | 0.50 |
| Tyrosine | 4.0 | 0.33 | 0 | 0.64 | 0.06 | 0.52 |
| Valine | 2.0 | 1.8 | 0.11 | 2.42 | 0.0 | 1.78 |
| Total | 48.47 | 33.27 | 4.52 | 52.59 | 9.2 | 50.86 |
| Sprout Species | Pre-Germination | Post-Germination | References |
|---|---|---|---|
| Chickpea | 67.7% 75.4% 64.2 ± 1.8% | 79.0% 86.5% 73.4 ± 0.7% | Uppal et al. (2012) [11] Chitra et al. (1996) [67] Ghavidel et al. (2007) [68] |
| Chickpea flour Mungbean | 83.8 ± 2.8% 66.4% 70.9% | 88.5 ± 3.2% 83.0% 82.7% | Sofi et al. (2023) [45] Uppal et al. (2012) [11] Chitra et al. (1996) [67] |
| Cowpea | 73.3% 71.2 ± 0.1% | 85.7% 73.5 ± 0.4% | Uppal et al. (2012) [11] Kalpanadevi et al. (2013) [69] |
| Soybean | 63.3% | 73.6% | Chitra et al. (1996) [67] |
| Pigeon pea | 69.1% | 85.1% | Chitra et al. (1996) [67] |
| Pigeon pea flour Kidney bean | 72.30 ± 0.24% 80.6 ± 0.02% | 82.66 ± 0.17% 87.1 ± 0.03% | Chinma et al. (2022) [46] Shimelis et al. (2006) [70] |
| Yellow pea | 78.6 ± 0.1% | 79.9 ± 0.1% | Setia et al. (2019) [71] |
| Fava bean | 78.0 ± 0.2% | 80.4 ± 0.1% | Setia et al. (2019) [71] |
| Lentil | 65.6 ± 1.1% | 64.2 ± 1.8% | Ghavidel et al. (2007) [68] |
| Pigeon Pea Lupin Green gram | 68% 73.0 ± 4.87% 61.0 ± 1.0% | 88% 74.3 ± 1.89% 72.7 ± 0.8% | Rizvi et (2022) [43] Munoz-Landes et al. (2022) [72] Ghavidel et al. (2007) [68] |
| Soy milk (Sorghum) | 80% 51% | 85% 65% | Hu et al. (2022) [73] Afify et al. (2012) [58] |
| (Red sorghum) | 48% | 68.1% | Onyango et al. (2013) [74] |
| (Pearl millet) | 21.5% | 34.5% | Onyango et al. (2013) [74] |
| Sprout Species | Pre-Germination | Post-Germination | References |
|---|---|---|---|
| Chickpea | 11.9 | 7.86 | El-Adawy (2002) [97] |
| Mungbean | 16.5 | 12.8 | El-Adawy et al. (2003) [97] |
| Pea | 10.8 | 8.6 | El-Adawy et al. (2003) [97] |
| Lentil | 33.3 | 27.3 | El-Adawy et al. (2003) [97] |
| Horsegram | 11.5 | 8.4 | Pal et al. (2013) [98] |
| Kidney bean | Shimelis et al. [74] | ||
| Roba variety | 4.5 | 3.8 | |
| Awash variety | 20.8 | 17.3 | |
| Beshbesh variety | 29.2 | 24.5 | |
| French bean | 3.1 | 2.2 | Alonso et al. (1999) [99] |
| Fava bean | 4.4 | 3.3 | Alonso et al. (1999) [99] |
| Soybean | 275 mg/g | 225 mg/g | Wu et al. (2023) [100] |
| (Sorghum) | |||
| Hamra variety | 31.6 | 19.9 | Osman et al. (2013) [92] |
| Allergic Protein Family | Characteristics |
|---|---|
| Prolamin superfamily | Largest family of plant food allergens, low molecular weight, sulfur-rich, glycosylated, includes 2S storage proteins from legumes, non-specific lipid transfer proteins, protease inhibitors |
| Cupin superfamily | Consists of two conserved consensus sequence motifs, β barrel structural domain, seed storage proteins of soybeans and peanuts |
| Pathogenesis-related proteins | Comprised of 14 different unrelated protein families, small size, stable in acidic conditions, Increased synthesis during environmental and pathogen stresses |
| Profilins | Small 12–15 kDa MW, highly conserved sequences, cytoplasmic immunological cross-reactivity with pollens |
| Vicilins | Part of the globulin family, anti-fungal, anti-microbial activity |
| Glycilins | Hexamer, 300–400 kDa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bera, I.; O’Sullivan, M.; Flynn, D.; Shields, D.C. Relationship between Protein Digestibility and the Proteolysis of Legume Proteins during Seed Germination. Molecules 2023, 28, 3204. https://doi.org/10.3390/molecules28073204
Bera I, O’Sullivan M, Flynn D, Shields DC. Relationship between Protein Digestibility and the Proteolysis of Legume Proteins during Seed Germination. Molecules. 2023; 28(7):3204. https://doi.org/10.3390/molecules28073204
Chicago/Turabian StyleBera, Indrani, Michael O’Sullivan, Darragh Flynn, and Denis C. Shields. 2023. "Relationship between Protein Digestibility and the Proteolysis of Legume Proteins during Seed Germination" Molecules 28, no. 7: 3204. https://doi.org/10.3390/molecules28073204
APA StyleBera, I., O’Sullivan, M., Flynn, D., & Shields, D. C. (2023). Relationship between Protein Digestibility and the Proteolysis of Legume Proteins during Seed Germination. Molecules, 28(7), 3204. https://doi.org/10.3390/molecules28073204