
The Synergy between Zinc and Antimicrobial Peptides: An Insight into Unique Bioinorganic Interactions
 , ,
, , 
Abstract
1. Introduction
2. Susceptibility of Microorganisms towards Zinc(II) Ions
3. Antimicrobial Peptides
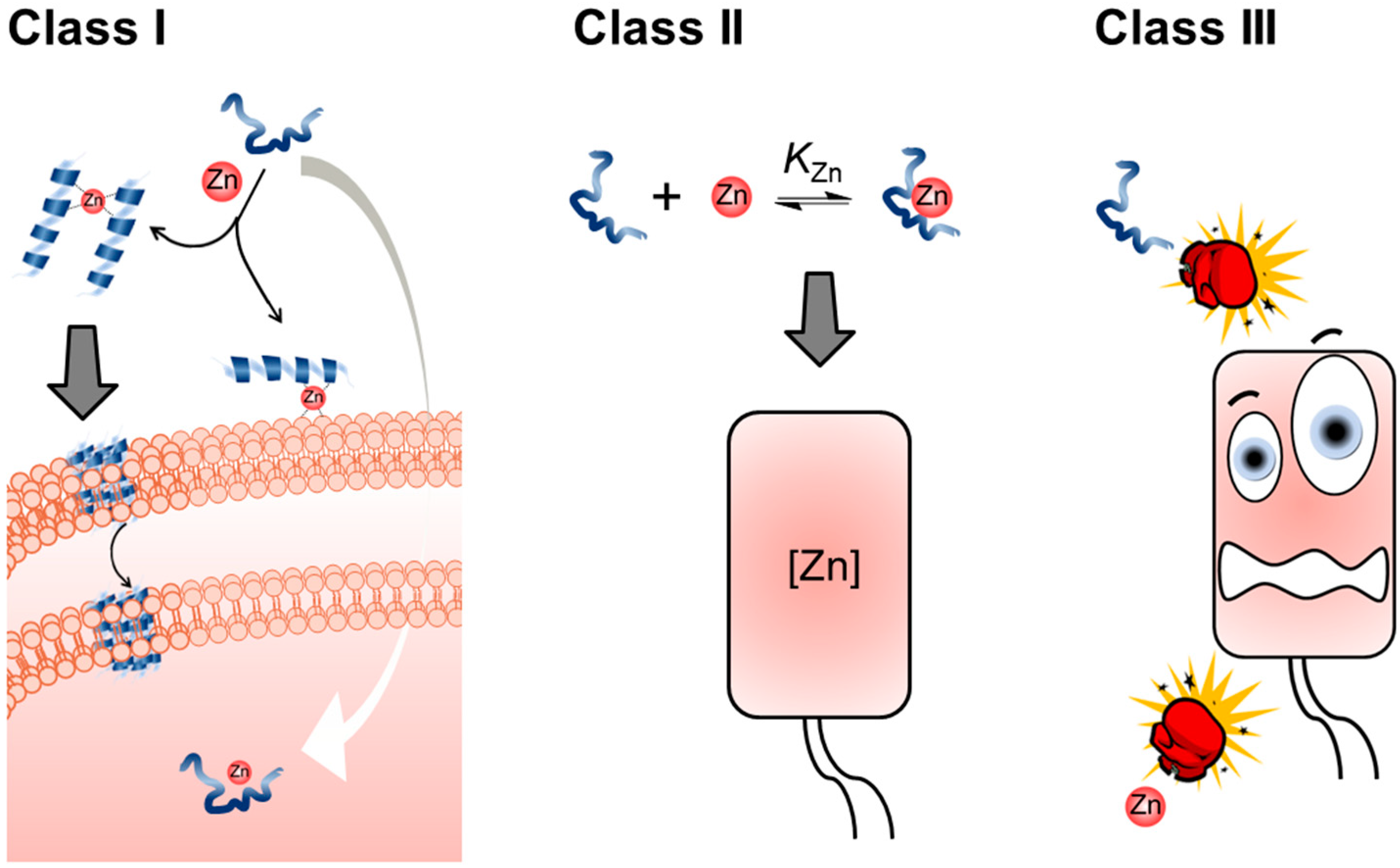
4. Nexus between Zn(II) and AMPs
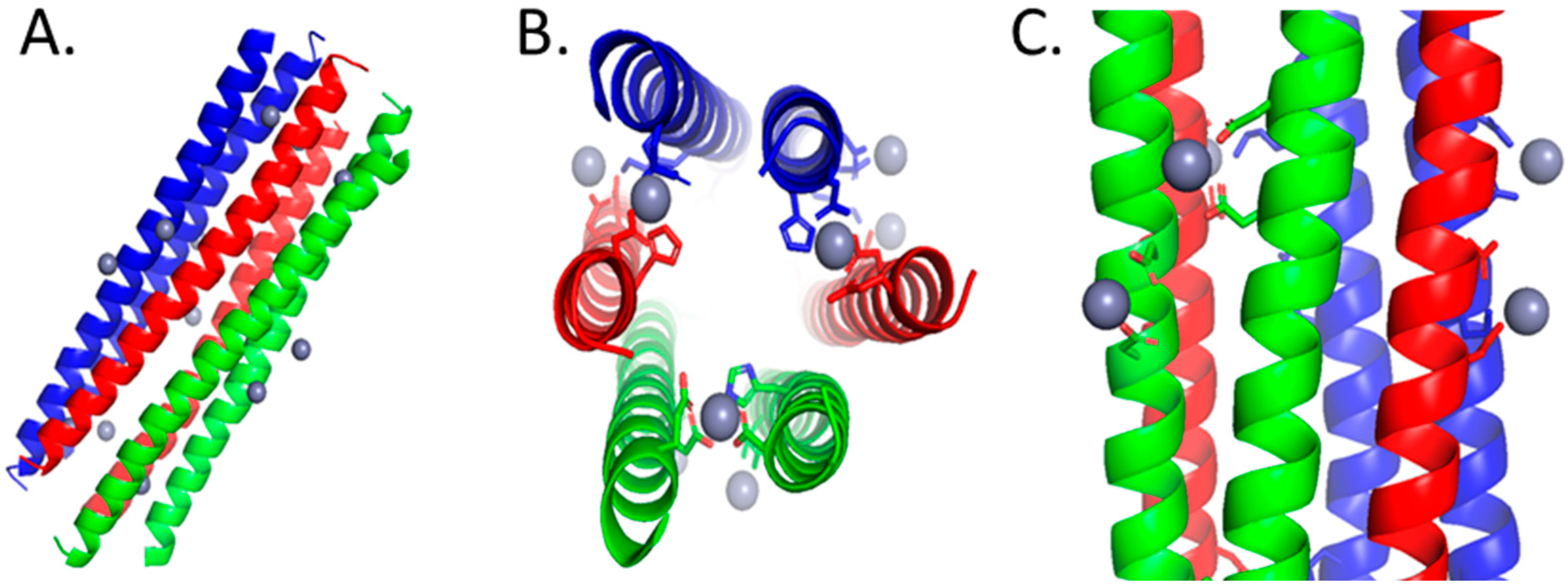
4.1. Dermcidin Derived Peptides
4.2. Clavanin A
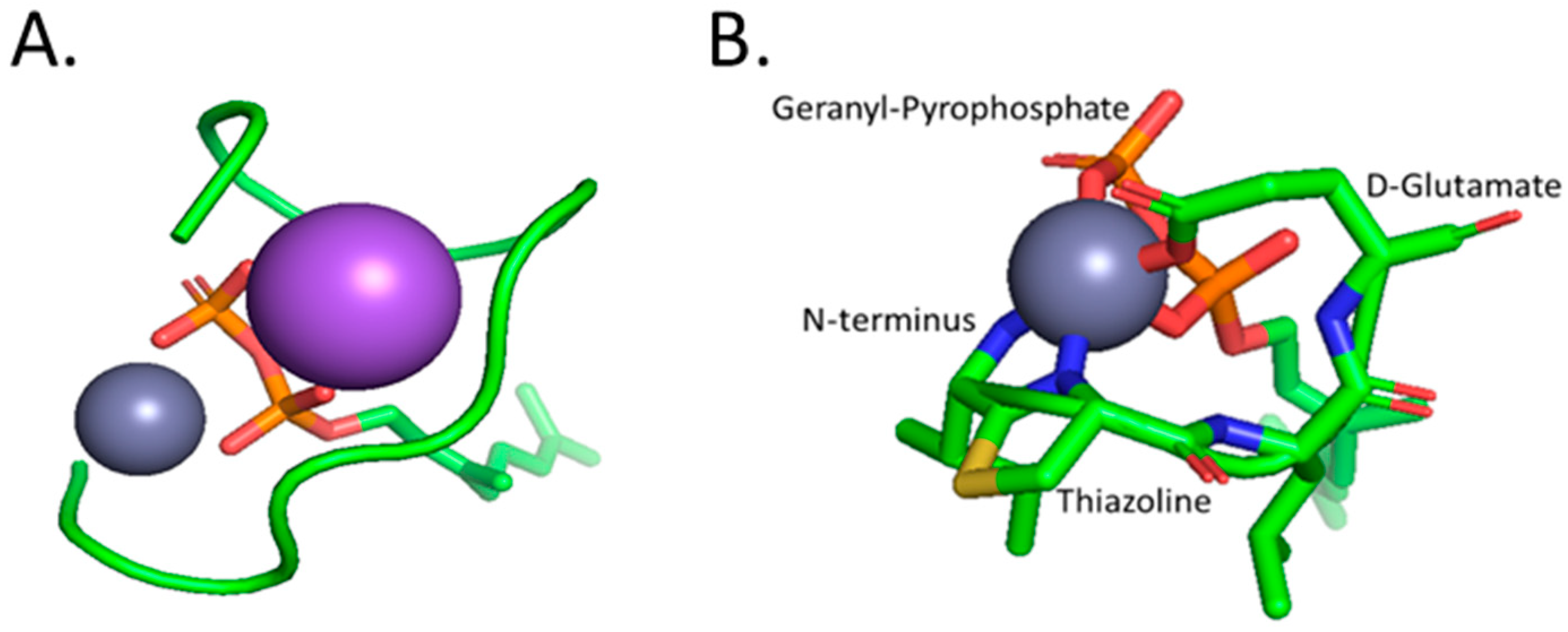
4.3. Bacitracin
4.4. Kappacin
4.5. Calcitermin
4.6. *ARVA Peptide
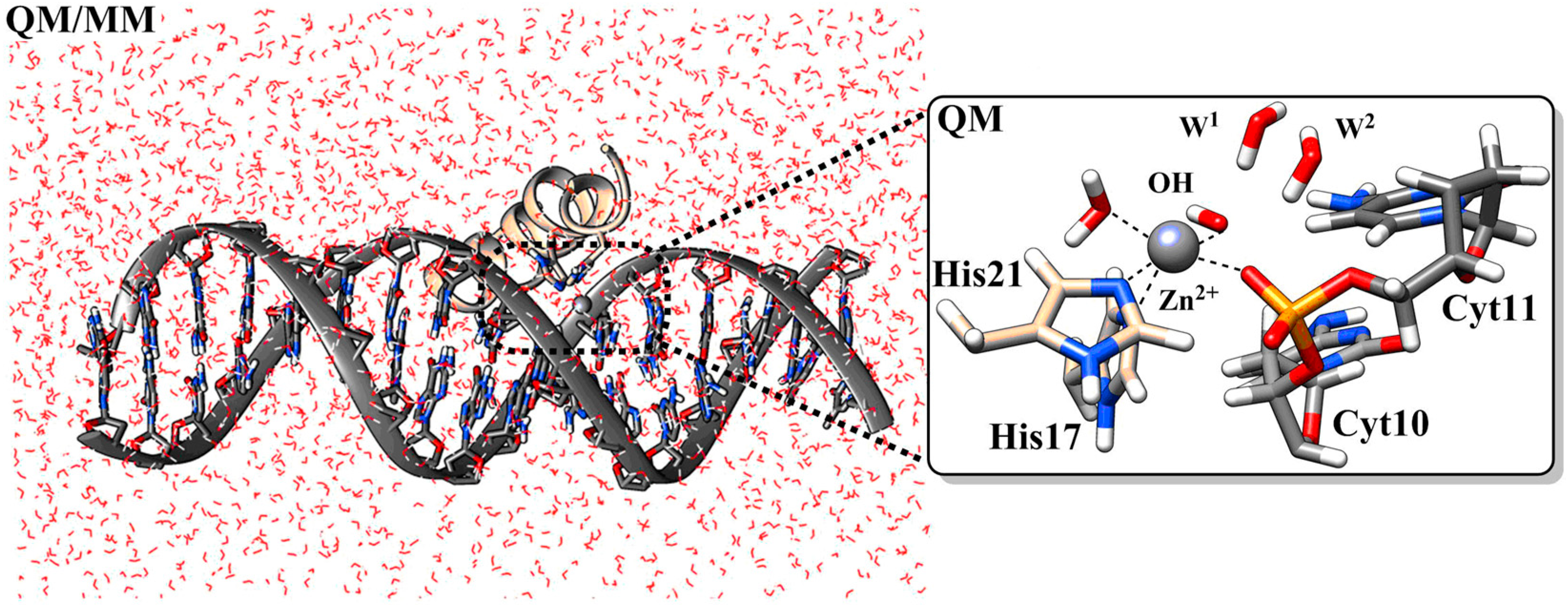
4.7. Histatins
4.8. Shepherin
4.9. Surfactant-Associated Anionic Peptides (SAAPs)
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Howell, M.; Wenc, A.; Donaghy, C.; Wasche, D.; Abissi, I.; Naing, M.; Pierce, S.; Angeles-Boza, A.M. Chapter Five—Exploring Synergy and Its Role in Antimicrobial Peptide Biology. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2022; Volume 663, pp. 99–130. [Google Scholar]
- Maret, W. The Redox Biology of Redox-Inert Zinc Ions. Free Radic. Biol. Med. 2019, 134, 311–326. [Google Scholar] [CrossRef]
- Kluska, K.; Adamczyk, J.; Krężel, A. Metal Binding Properties, Stability and Reactivity of Zinc Fingers. Coord. Chem. Rev. 2018, 367, 18–64. [Google Scholar] [CrossRef]
- Ong, C.Y.; Berking, O.; Walker, M.J.; McEwan, A.G. New Insights into the Role of Zinc Acquisition and Zinc Tolerance in Group A Streptococcal Infection. Infect. Immun. 2018, 86, e00048-18. [Google Scholar] [CrossRef] [PubMed]
- Djoko, K.Y.; Ong, C.Y.; Walker, M.J.; McEwan, A.G. The Role of Copper and Zinc Toxicity in Innate Immune Defense against Bacterial Pathogens. J. Biol. Chem. 2015, 290, 18954–18961. [Google Scholar] [CrossRef] [PubMed]
- Ibs, K.-H.; Rink, L. Zinc-Altered Immune Function. J. Nutr. 2003, 133, 1452S–1456S. [Google Scholar] [CrossRef] [PubMed]
- Milanino, R.; Marrella, M.; Gasperini, R.; Pasqualicchio, M.; Velo, G. Copper and Zinc Body Levels in Inflammation: An Overview of the Data Obtained from Animal and Human Studies. Agents Actions 1993, 39, 195–209. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, R.; Wei, X.; Lv, M.; Jiang, Z. Metalloimmunology: The Metal Ion-Controlled Immunity. In Advances in Immunology; Academic Press: Cambridge, MA, USA, 2020; Volume 145, pp. 187–241. [Google Scholar]
- Stafford, S.L.; Bokil, N.J.; Achard, M.E.S.; Kapetanovic, R.; Schembri, M.A.; McEwan, A.G.; Sweet, M.J. Metal Ions in Macrophage Antimicrobial Pathways: Emerging Roles for Zinc and Copper. Biosci. Rep. 2013, 33, e00049. [Google Scholar] [CrossRef]
- Ong, C.Y.; Gillen, C.M.; Barnett, T.C.; Walker, M.J.; McEwan, A.G. An Antimicrobial Role for Zinc in Innate Immune Defense Against Group A Streptococcus. J. Infect. Dis. 2014, 209, 1500–1508. [Google Scholar] [CrossRef]
- Wagner, D.; Maser, J.; Lai, B.; Cai, Z.; Barry, C.E.; Höner zu Bentrup, K.; Russell, D.G.; Bermudez, L.E. Elemental Analysis of Mycobacterium avium-, Mycobacterium tuberculosis-, and Mycobacterium smegmatis—Containing Phagosomes Indicates Pathogen-Induced Microenvironments within the Host Cell’s Endosomal System. J. Immunol. 2005, 174, 1491–1500. [Google Scholar] [CrossRef]
- Juliano, S.A.; Serafim, L.F.; Duay, S.S.; Heredia Chavez, M.; Sharma, G.; Rooney, M.; Comert, F.; Pierce, S.; Radulescu, A.; Cotten, M.L.; et al. A Potent Host Defense Peptide Triggers DNA Damage and Is Active against Multidrug-Resistant Gram-Negative Pathogens. ACS Infect. Dis. 2020, 6, 1250–1263. [Google Scholar] [CrossRef]
- Ning, C.; Wang, X.; Li, L.; Zhu, Y.; Li, M.; Yu, P.; Zhou, L.; Zhou, Z.; Chen, J.; Tan, G.; et al. Concentration Ranges of Antibacterial Cations for Showing the Highest Antibacterial Efficacy but the Least Cytotoxicity against Mammalian Cells: Implications for a New Antibacterial Mechanism. Chem. Res. Toxicol. 2015, 28, 1815–1822. [Google Scholar] [CrossRef]
- Choudhury, R.; Srivastava, S. Zinc Resistance Mechanisms in Bacteria. Curr. Sci. 2001, 81, 768–775. [Google Scholar]
- Braymer, J.J.; Giedroc, D.P. Recent Developments in Copper and Zinc Homeostasis in Bacterial Pathogens. Curr. Opin. Chem. Biol. 2014, 19, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Beard, S.; Hashim, R.; Membrillo-Hernandez, J.; Hughes, M.; Poole, R. Zinc(II) Tolerance in Escherichia coli K-12: Evidence That the ZntA Gene (O732) Encodes a Cation Transport ATPase. Mol. Microbiol. 2004, 25, 883–891. [Google Scholar] [CrossRef]
- Ong, C.Y.; Walker, M.J.; McEwan, A.G. Zinc Disrupts Central Carbon Metabolism and Capsule Biosynthesis in Streptococcus Pyogenes. Sci. Rep. 2015, 5, 10799. [Google Scholar] [CrossRef]
- Soares, L.W.; Bailão, A.M.; de Almeida Soares, C.M.; Bailão, M.G.S. Zinc at the Host–Fungus Interface: How to Uptake the Metal? J. Fungi 2020, 6, 305. [Google Scholar] [CrossRef]
- Jung, W.H. The Zinc Transport Systems and Their Regulation in Pathogenic Fungi. Mycobiology 2015, 43, 179–183. [Google Scholar] [CrossRef]
- Volkova, M.; Atamas, A.; Tsarenko, A.; Rogachev, A.; Guskov, A. Cation Transporters of Candida albicans—New Targets to Fight Candidiasis? Biomolecules 2021, 11, 584. [Google Scholar] [CrossRef] [PubMed]
- McDevitt, C.A.; Ogunniyi, A.D.; Valkov, E.; Lawrence, M.C.; Kobe, B.; McEwan, A.G.; Paton, J.C. A Molecular Mechanism for Bacterial Susceptibility to Zinc. PLoS Pathog. 2011, 7, e1002357. [Google Scholar] [CrossRef]
- Li, J.; Ren, X.; Fan, B.; Huang, Z.; Wang, W.; Zhou, H.; Lou, Z.; Ding, H.; Lyu, J.; Tan, G. Zinc Toxicity and Iron-Sulfur Cluster Biogenesis in Escherichia coli. Appl. Environ. Microbiol. 2019, 85, e01967-18. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.E.; Mason, Z.; Rodrigues, A.V.; Nuth, M.; Dizin, E.; Cowan, J.A.; Stemmler, T.L. Unique Roles of Iron and Zinc Binding to the Yeast Fe–S Cluster Scaffold Assembly Protein “Isu1”. Metallomics 2019, 11, 1820–1835. [Google Scholar] [CrossRef] [PubMed]
- Bunnell, B.E.; Escobar, J.F.; Bair, K.L.; Sutton, M.D.; Crane, J.K. Zinc Blocks SOS-Induced Antibiotic Resistance via Inhibition of RecA in Escherichia coli. PLoS ONE 2017, 12, e0178303. [Google Scholar] [CrossRef] [PubMed]
- Gutknecht, J. Cadmium and Thallous Ion Permeabilities through Lipid Bilayer Membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 1983, 735, 185–188. [Google Scholar] [CrossRef]
- Duay, S.S.; Sharma, G.; Prabhakar, R.; Angeles-Boza, A.M.; May, E.R. Molecular Dynamics Investigation into the Effect of Zinc(II) on the Structure and Membrane Interactions of the Antimicrobial Peptide Clavanin A. J. Phys. Chem. B 2019, 123, 3163–3176. [Google Scholar] [CrossRef] [PubMed]
- Kučerka, N.; Dushanov, E.; Kholmurodov, K.T.; Katsaras, J.; Uhríková, D. Calcium and Zinc Differentially Affect the Structure of Lipid Membranes. Langmuir 2017, 33, 3134–3141. [Google Scholar] [CrossRef]
- Portelinha, J.; Duay, S.S.; Yu, S.I.; Heilemann, K.; Libardo, M.D.J.; Juliano, S.A.; Klassen, J.L.; Angeles-Boza, A.M. Antimicrobial Peptides and Copper(II) Ions: Novel Therapeutic Opportunities. Chem. Rev. 2021, 121, 2648–2712. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.L.; Thompson, Z.; Cowan, J.A. Antimicrobial Metallopeptides. ACS Chem. Biol. 2018, 13, 844–853. [Google Scholar] [CrossRef]
- Wimley, W.C. Describing the Mechanism of Antimicrobial Peptide Action with the Interfacial Activity Model. ACS Chem. Biol. 2010, 5, 905–917. [Google Scholar] [CrossRef]
- Mercer, D.K.; Torres, M.D.T.; Duay, S.S.; Lovie, E.; Simpson, L.; von Köckritz-Blickwede, M.; de la Fuente-Nunez, C.; O’Neil, D.A.; Angeles-Boza, A.M. Antimicrobial Susceptibility Testing of Antimicrobial Peptides to Better Predict Efficacy. Front. Cell. Infect. Microbiol. 2020, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W.; Lehrer, R. Cationic Peptides: A New Source of Antibiotics. Trends Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Harris, F.; Dennison, S.; Phoenix, D. Anionic Antimicrobial Peptides from Eukaryotic Organisms and Their Mechanisms of Action. Curr. Chem. Biol. 2011, 5, 142–153. [Google Scholar] [CrossRef]
- Lehrer, R.; Ganz, T. Antimicrobial Peptides in Mammalian and Insect Host Defence. Curr. Opin. Immunol. 1999, 11, 22–27. [Google Scholar] [CrossRef]
- Libardo, M.D.; Angeles-Boza, A.M. Bioinorganic Chemistry of Antimicrobial and Host-Defense Peptides. Comments Inorg. Chem. 2014, 34, 42–58. [Google Scholar] [CrossRef]
- Tonk, M.; Vilcinskas, A. The Medical Potential of Antimicrobial Peptides from Insects. Curr. Top. Med. Chem. 2017, 17, 554–575. [Google Scholar] [CrossRef] [PubMed]
- Walkenhorst, W.F. Using Adjuvants and Environmental Factors to Modulate the Activity of Antimicrobial Peptides. Biochim. Biophys. Acta (BBA)-Biomembr. 2016, 1858, 926–935. [Google Scholar] [CrossRef]
- Guha, S.; Ghimire, J.; Wu, E.; Wimley, W.C. Mechanistic Landscape of Membrane-Permeabilizing Peptides. Chem. Rev. 2019, 119, 6040–6085. [Google Scholar] [CrossRef] [PubMed]
- Le, C.-F.; Fang, C.-M.; Sekaran, S.D. Intracellular Targeting Mechanisms by Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, e02340-16. [Google Scholar] [CrossRef] [PubMed]
- Madani, F.; Lindberg, S.; Langel, Ü.; Futaki, S.; Gräslund, A. Mechanisms of Cellular Uptake of Cell-Penetrating Peptides. J. Biophys. 2011, 2011, 414729. [Google Scholar] [CrossRef]
- Subbalakshmi, C.; Sitaram, N. Mechanism of Antimicrobial Action of Indolicidin. FEMS Microbiol. Lett. 1998, 160, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-H.; Shah, P.; Chen, Y.-W.; Chen, C.-S. Systematic Analysis of Intracellular-Targeting Antimicrobial Peptides, Bactenecin 7, Hybrid of Pleurocidin and Dermaseptin, Proline–Arginine- Rich Peptide, and Lactoferricin B, by Using Escherichia coli Proteome Microarrays. Mol. Cell. Proteom. 2016, 15, 1837–1847. [Google Scholar] [CrossRef]
- Nishikata, M.; Kanehira, T.; Oh, H.; Tani, H.; Tazaki, M.; Kuboki, Y. Salivary Histatin as an Inhibitor of a Protease Produced by the Oral Bacterium Bacteroides Gingivalis. Biochem. Biophys. Res. Commun. 1991, 174, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.X.; Gao, S.; Anand, K.S.; Franz, K.J. Zinc Binding Inhibits Cellular Uptake and Antifungal Activity of Histatin-5 in Candida albicans. ACS Infect. Dis. 2022, 8, 1920–1934. [Google Scholar] [CrossRef] [PubMed]
- Juliano, S.A.; Pierce, S.; deMayo, J.A.; Balunas, M.J.; Angeles-Boza, A.M. Exploration of the Innate Immune System of Styela clava: Zn2+ Binding Enhances the Antimicrobial Activity of the Tunicate Peptide Clavanin A. Biochemistry 2017, 56, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Eissa, A.; Amodeo, V.; Smith, C.R.; Diamandis, E.P. Kallikrein-Related Peptidase-8 (KLK8) Is an Active Serine Protease in Human Epidermis and Sweat and Is Involved in a Skin Barrier Proteolytic Cascade. J. Biol. Chem. 2011, 286, 687–706. [Google Scholar] [CrossRef] [PubMed]
- Schittek, B.; Hipfel, R.; Sauer, B.; Bauer, J.; Kalbacher, H.; Stevanovic, S.; Schirle, M.; Schroeder, K.; Blin, N.; Meier, F.; et al. Dermcidin: A Novel Human Antibiotic Peptide Secreted by Sweat Glands. Nat. Immunol. 2001, 2, 1133–1137. [Google Scholar] [CrossRef]
- Rieg, S.; Steffen, H.; Seeber, S.; Humeny, A.; Kalbacher, H.; Dietz, K.; Garbe, C.; Schittek, B. Deficiency of Dermcidin-Derived Antimicrobial Peptides in Sweat of Patients with Atopic Dermatitis Correlates with an Impaired Innate Defense of Human Skin In Vivo. J. Immunol. 2005, 174, 8003–8010. [Google Scholar] [CrossRef]
- Paulmann, M.; Arnold, T.; Linke, D.; Özdirekcan, S.; Kopp, A.; Gutsmann, T.; Kalbacher, H.; Wanke, I.; Schuenemann, V.J.; Habeck, M.; et al. Structure-Activity Analysis of the Dermcidin-Derived Peptide DCD-1L, an Anionic Antimicrobial Peptide Present in Human Sweat. J. Biol. Chem. 2012, 287, 8434–8443. [Google Scholar] [CrossRef]
- Steffen, H.; Rieg, S.; Wiedemann, I.; Kalbacher, H.; Deeg, M.; Sahl, H.-G.; Peschel, A.; Götz, F.; Garbe, C.; Schittek, B. Naturally Processed Dermcidin-Derived Peptides Do Not Permeabilize Bacterial Membranes and Kill Microorganisms Irrespective of Their Charge. Antimicrob. Agents Chemother. 2006, 50, 2608–2620. [Google Scholar] [CrossRef]
- Song, C.; Weichbrodt, C.; Salnikov, E.S.; Dynowski, M.; Forsberg, B.O.; Bechinger, B.; Steinem, C.; de Groot, B.L.; Zachariae, U.; Zeth, K. Crystal Structure and Functional Mechanism of a Human Antimicrobial Membrane Channel. Proc. Natl. Acad. Sci. USA 2013, 110, 4586–4591. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System; Schrodinger LLC: New York, NY, USA, 2010.
- Lee, I.H.; Cho, Y.; Lehrer, R.I. Effects of PH and Salinity on the Antimicrobial Properties of Clavanins. Infect. Immun. 1997, 65, 2898–2903. [Google Scholar] [CrossRef]
- Juliano, S.A. From Antimicrobial Activity to Zinc Binding: An In-Depth Analysis of the Tunicate Host Defense Peptide Clavanin A. Ph.D. Thesis, University of Connecticut, Storrs, CT, USA, 2020. [Google Scholar]
- van Kan, E.J.M.; van der Bent, A.; Demel, R.A.; de Kruijff, B. Membrane Activity of the Peptide Antibiotic Clavanin and the Importance of Its Glycine Residues. Biochemistry 2001, 40, 6398–6405. [Google Scholar] [CrossRef] [PubMed]
- van Kan, E.J.M.; Demel, R.A.; Breukink, E.; van der Bent, A.; de Kruijff, B. Clavanin Permeabilizes Target Membranes via Two Distinctly Different PH-Dependent Mechanisms. Biochemistry 2002, 41, 7529–7539. [Google Scholar] [CrossRef] [PubMed]
- Tay, W.M.; Epperson, J.D.; da Silva, G.F.Z.; Ming, L.-J. 1H NMR, Mechanism, and Mononuclear Oxidative Activity of the Antibiotic Metallopeptide Bacitracin: The Role of d-Glu-4, Interaction with Pyrophosphate Moiety, DNA Binding and Cleavage, and Bioactivity. J. Am. Chem. Soc. 2010, 132, 5652–5661. [Google Scholar] [CrossRef] [PubMed]
- Economou, N.J.; Cocklin, S.; Loll, P.J. High-Resolution Crystal Structure Reveals Molecular Details of Target Recognition by Bacitracin. Proc. Natl. Acad. Sci. USA 2013, 110, 14207–14212. [Google Scholar] [CrossRef]
- Tsuji, K.; Robertson, J.; Bach, J. Quantitative High-Pressure Liquid Chromatographic Analysis of Bacitracin, a Polypeptide Antibiotic. J. Chromatogr. A 1974, 99, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.-J.; Epperson, J.D. Metal Binding and Structure–Activity Relationship of the Metalloantibiotic Peptide Bacitracin. J. Inorg. Biochem. 2002, 91, 46–58. [Google Scholar] [CrossRef]
- Adler, R.H.; Snoke, J.E. Requirement of Divalent Metal Ions for Bacitracin Activity. J Bacteriol 1962, 83, 1315–1317. [Google Scholar] [CrossRef]
- Storm, D.R. Mechanism of Bacitractin Action: A Specific Lipid-Peptide Interaction. Ann. N. Y. Acad. Sci. 1974, 235, 387–398. [Google Scholar] [CrossRef]
- Storm, D.R.; Strominger, J.L. Complex Formation between Bacitracin Peptides and Isoprenyl Pyrophosphates. J. Biol. Chem. 1973, 248, 3940–3945. [Google Scholar] [CrossRef]
- Malkoski, M.; Dashper, S.G.; O’Brien-Simpson, N.M.; Talbo, G.H.; Macris, M.; Cross, K.J.; Reynolds, E.C. Kappacin, a Novel Antibacterial Peptide from Bovine Milk. Antimicrob. Agents Chemother. 2001, 45, 2309–2315. [Google Scholar] [CrossRef]
- Dashper, S.G.; O’Brien-Simpson, N.M.; Cross, K.J.; Paolini, R.A.; Hoffmann, B.; Catmull, D.V.; Malkoski, M.; Reynolds, E.C. Divalent Metal Cations Increase the Activity of the Antimicrobial Peptide Kappacin. Antimicrob. Agents Chemother. 2005, 49, 2322–2328. [Google Scholar] [CrossRef]
- Smith, M.; Edwards, P.; Palmano, K.; Creamer, L. Structural Features of Bovine Caseinomacropeptide A and B by 1H Nuclear Magnetic Resonance Spectroscopy. J. Dairy Res. 2002, 69, 85–94. [Google Scholar] [CrossRef]
- Cole, A.M.; Kim, Y.-H.; Tahk, S.; Hong, T.; Weis, P.; Waring, A.J.; Ganz, T. Calcitermin, a Novel Antimicrobial Peptide Isolated from Human Airway Secretions. FEBS Lett. 2001, 504, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Gottsch, J.D.; Eisinger, S.W.; Liu, S.H.; Scott, A.L. Calgranulin C Has Filariacidal and Filariastatic Activity. Infect. Immun. 1999, 67, 6631–6636. [Google Scholar] [CrossRef] [PubMed]
- Bellotti, D.; Toniolo, M.; Dudek, D.; Mikołajczyk, A.; Guerrini, R.; Matera-Witkiewicz, A.; Remelli, M.; Rowińska-Żyrek, M. Bioinorganic Chemistry of Calcitermin—he Picklock of Its Antimicrobial Activity. Dalton Trans. 2019, 48, 13740–13752. [Google Scholar] [CrossRef] [PubMed]
- Rathinakumar, R.; Walkenhorst, W.F.; Wimley, W.C. Broad-Spectrum Antimicrobial Peptides by Rational Combinatorial Design and High-Throughput Screening: The Importance of Interfacial Activity. J. Am. Chem. Soc. 2009, 131, 7609–7617. [Google Scholar] [CrossRef] [PubMed]
- Walkenhorst, W.F.; Sundrud, J.N.; Laviolette, J.M. Additivity and Synergy between an Antimicrobial Peptide and Inhibitory Ions. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 2234–2242. [Google Scholar] [CrossRef]
- Aoki, S.; Iwaida, K.; Hanamoto, N.; Shiro, M.; Kimura, E. Guanidine Is a Zn 2+ -Binding Ligand at Neutral PH in Aqueous Solution. J. Am. Chem. Soc. 2002, 124, 5256–5257. [Google Scholar] [CrossRef]
- Sabatini, L.; Azen, E. Histatins, a Family of Salivary Histidine-Rich Proteins, Are Encoded by at Least Two Loci (HIS1 and HIS2). Biochem. Biophys. Res. Commun. 1989, 160, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Melino, S.; Santone, C.; Di Nardo, P.; Sarkar, B. Histatins: Salivary Peptides with Copper(II)- and Zinc(II)-binding Motifs. FEBS Lett. 2013, 281, 657–672. [Google Scholar] [CrossRef]
- Oppenheim, F.G.; Xu, T.; McMillian, F.M.; Levitz, S.M.; Diamond, R.D.; Offner, G.D.; Troxler, R.F. Histatins, a Novel Family of Histidine-Rich Proteins in Human Parotid Secretion. Isolation, Characterization, Primary Structure, and Fungistatic Effects on Candida Albicans. J. Biol. Chem. 1988, 263, 7472–7477. [Google Scholar] [CrossRef] [PubMed]
- Castagnola, M.; Inzitari, R.; Rossetti, D.V.; Olmi, C.; Cabras, T.; Piras, V.; Nicolussi, P.; Sanna, M.T.; Pellegrini, M.; Giardina, B.; et al. A Cascade of 24 Histatins (Histatin 3 Fragments) in Human Saliva. J. Biol. Chem. 2004, 279, 41436–41443. [Google Scholar] [CrossRef] [PubMed]
- Puri, S.; Edgerton, M. How Does It Kill?: Understanding the Candidacidal Mechanism of Salivary Histatin 5. Eukaryot. Cell 2014, 13, 958–964. [Google Scholar] [CrossRef]
- Pan, L.; Zhang, X.; Gao, Q. Effects and Mechanisms of Histatins as Novel Skin Wound-Healing Agents. J. Tissue Viability 2021, 30, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Puri, S.; McCall, A.; Norris, H.L.; Russo, T.; Edgerton, M. Human Salivary Protein Histatin 5 Has Potent Bactericidal Activity against ESKAPE Pathogens. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Norris, H.L.; Kumar, R.; Ong, C.Y.; Xu, D.; Edgerton, M. Zinc Binding by Histatin 5 Promotes Fungicidal Membrane Disruption in C. albicans and C. glabrata. J. Fungi 2020, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Helmerhorst, E.J.; Reijnders, I.M.; van ’t Hof, W.; Simoons-Smit, I.; Veerman, E.C.I.; Amerongen, A.V.N. Amphotericin B- and Fluconazole-Resistant Candida spp., Aspergillus fumigatus, and Other Newly Emerging Pathogenic Fungi Are Susceptible to Basic Antifungal Peptides. Antimicrob. Agents Chemother. 1999, 43, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Fidel Jr., P.; Thunayyan, A.; Varlotta, S.; Meiller, T.; Jabra-Rizk, M.A. Impaired Histatin-5 Levels and Salivary Antimicrobial Activity against C.Albicans in HIV Infected Individuals. J. AIDS Clin. Res. 2013, 4, 1000193. [Google Scholar] [CrossRef]
- McCaslin, T.G.; Pagba, C.V.; Yohannan, J.; Barry, B.A. Specific Metallo-Protein Interactions and Antimicrobial Activity in Histatin-5, an Intrinsically Disordered Salivary Peptide. Sci. Rep. 2019, 9, 17303. [Google Scholar] [CrossRef]
- Cragnell, C.; Staby, L.; Lenton, S.; Kragelund, B.; Skepö, M. Dynamical Oligomerisation of Histidine Rich Intrinsically Disordered Proteins Is Regulated through Zinc-Histidine Interactions. Biomolecules 2019, 9, 168. [Google Scholar] [CrossRef]
- Kurut, A.; Henriques, J.; Forsman, J.; Skepo, M.; Lund, M. Role of Histidine for Charge Regulation of Unstructured Peptides at Interfaces and in Bulk. Proteins 2013, 82, 657–667. [Google Scholar] [CrossRef]
- Stewart, L.; Hong, Y.; Holmes, I.; Firth, S.; Bolton, J.; Santos, Y.; Cobb, S.; Jakubovics, N.; Djoko, K. The Role of Metal Binding in the Function of the Human Salivary Antimicrobial Peptide Histatin-5. bioRxiv 2022. preprint. [Google Scholar] [CrossRef]
- Norris, H.; Kumar, R.; Edgerton, M. A Novel Role for Histatin 5 in Combination with Zinc to Promote Commensalism in C. albicans Survivor Cells. Pathogens 2021, 10, 1609. [Google Scholar] [CrossRef] [PubMed]
- Flora, B.; Heloisa, G.; Helmerhorst, E.J.; Troxler, R.; Oppenheim, F.G. A New Method for the Isolation of Histatins 1, 3, and 5 from Parotid Secretion Using Zinc Precipitation. Protein Expr. Purif. 2001, 23, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Park, C.J.; Park, C.B.; Hong, S.-S.; Lee, H.-S.; Lee, S.Y.; Kim, S.C. Characterization and CDNA Cloning of Two Glycine- and Histidine-Rich Antimicrobial Peptides from the Roots of Shepherd’s Purse, Capsella Bursa-Pastoris. Plant Mol. Biol. 2000, 44, 187–197. [Google Scholar] [CrossRef]
- Remuzgo, C.; Oewel, T.S.; Daffre, S.; Lopes, T.R.S.; Dyszy, F.H.; Schreier, S.; Machado-Santelli, G.M.; Teresa Machini, M. Chemical Synthesis, Structure–Activity Relationship, and Properties of Shepherin I: A Fungicidal Peptide Enriched in Glycine-Glycine-Histidine Motifs. Amino Acids 2014, 46, 2573–2586. [Google Scholar] [CrossRef]
- Reichert, T. Análogos Sintéticos da Cheferina I: Interação com Íons Metálicos Divalentes e o Seu Efeito na Internalização Celular e Nas Atividades Anticandida e Candidacida. Ph.D. Thesis, University of São Paulo, Sao Paulo, Brazil, 2018. [Google Scholar]
- Libardo, M.D.J.; De La Fuente-Nuñez, C.; Anand, K.; Krishnamoorthy, G.; Kaiser, P.; Pringle, S.C.; Dietz, C.; Pierce, S.; Smith, M.B.; Barczak, A.; et al. Phagosomal Copper-Promoted Oxidative Attack on Intracellular Mycobacterium Tuberculosis. ACS Infect. Dis. 2018, 4, 1623–1634. [Google Scholar] [CrossRef]
- Brogden, K.A.; De Lucca, A.J.; Bland, J.; Elliott, S. Isolation of an Ovine Pulmonary Surfactant-Associated Anionic Peptide Bactericidal for Pasteurella Haemolytica. Proc. Natl. Acad. Sci. USA 1996, 93, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Ovine Pulmonary Surfactant Induces Killing of Pasteurella haemolytica, Escherichia coli, and Klebsiella pneumoniae by Normal Serum. Infect. Immun. 1992, 60, 5182–5189. [Google Scholar] [CrossRef]
- Miller, A.; Matera-Witkiewicz, A.; Mikołajczyk, A.; Wątły, J.; Wilcox, D.; Witkowska, D.; Rowińska-Żyrek, M. Zn-Enhanced Asp-Rich Antimicrobial Peptides: N-Terminal Coordination by Zn(II) and Cu(II), Which Distinguishes Cu(II) Binding to Different Peptides. Int. J. Mol. Sci. 2021, 22, 6971. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Structural Features in Membrane Mimetic Environments | Classification |
|---|---|---|---|
| DCD-1L | SSLLEKGLDGAKKAVGGLGKLGKDAVEDLESVGKGAVHDVKDVLDSVL | α-helical, Oligomerizes to trimers of Zn(II)-bridged dimers in presence of Zn(II) | Class I |
| Clavanin A | VFQFLGKIIHHVGNFVHGFSHVF-NH2 | α-helical | Class I |
| Bacitracin | ICLEI-cyclo(KOIFHDN) | Cyclic peptide, Mixture of dodecapeptides | Class I |
| Kappacin (33-53) | AVESTVATLEDSPEVIESPPE | Slight α-helix upon binding to divalent ion | Class I * |
| Calcitermin | VAIALKAAHYHTHKE | α-helical | Class I * |
| *ARVA peptide | RRGWALRLVLAY-NH2 | β-sheet | Class III |
| Histatin-5 | DSHAKRHHGYKRKFHEKHHSHRGY | Slightly α-helical, Forms oligomers upon interaction with Zn(II) | Class I and/or III |
| Shepherin I | GTGGHGGHGGHGGHGGHGGHGHGGGGHG | Slight β-sheet | Unclassified |
| Shepherin II | GYHGGHGGHGGGYNGGGGHGGHGGGYNGGGHHGGGGHG | Random coil | Unclassified |
| SAAP2 | GDDDDDD | Random coil | Unclassified |
| SAAP3 | DDDDDDD | Random coil | Unclassified |
| SAAP6 | GADDDDD | Random coil | Unclassified |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donaghy, C.; Javellana, J.G.; Hong, Y.-J.; Djoko, K.; Angeles-Boza, A.M. The Synergy between Zinc and Antimicrobial Peptides: An Insight into Unique Bioinorganic Interactions. Molecules 2023, 28, 2156. https://doi.org/10.3390/molecules28052156
Donaghy C, Javellana JG, Hong Y-J, Djoko K, Angeles-Boza AM. The Synergy between Zinc and Antimicrobial Peptides: An Insight into Unique Bioinorganic Interactions. Molecules. 2023; 28(5):2156. https://doi.org/10.3390/molecules28052156
Chicago/Turabian StyleDonaghy, Caroline, Jose Gabriel Javellana, Young-Jin Hong, Karrera Djoko, and Alfredo M. Angeles-Boza. 2023. "The Synergy between Zinc and Antimicrobial Peptides: An Insight into Unique Bioinorganic Interactions" Molecules 28, no. 5: 2156. https://doi.org/10.3390/molecules28052156
APA StyleDonaghy, C., Javellana, J. G., Hong, Y.-J., Djoko, K., & Angeles-Boza, A. M. (2023). The Synergy between Zinc and Antimicrobial Peptides: An Insight into Unique Bioinorganic Interactions. Molecules, 28(5), 2156. https://doi.org/10.3390/molecules28052156