
Proteomic Study of Broiler Plasma Supplemented with Different Levels of Copper and Manganese from Different Sources
 ,
,  , and
, and
Abstract
1. Introduction
2. Results
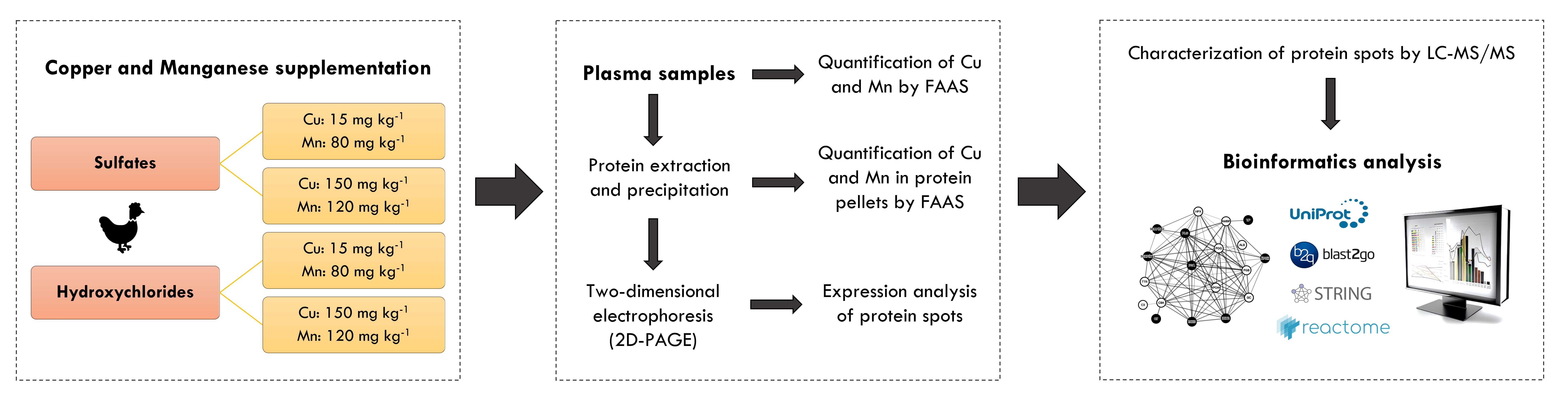
2.1. Fractionation of Broiler Plasma Proteome
2.2. Copper and Manganese Determinations
2.3. Expression Analysis of Protein Spot
3. Discussion
3.1. Cu and Mn Concentrations in Plasma and Protein Pellet Samples
3.2. Regulation of Plasma Proteins
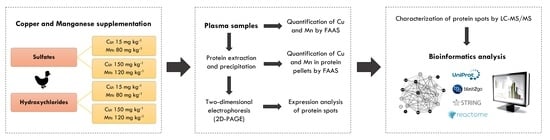
4. Materials and Methods
4.1. Birds, Experimental Groups and Sample Collection
4.2. Extraction, Precipitation, and Determination of Total Protein
4.3. Two-Dimensional Polyacrylamide Gel Electrophoresis (2D-PAGE) and Gel Analysis
4.4. Characterization of Protein Spot
4.5. Bioinformatics Analysis
4.6. Quantification of Copper and Manganese in Protein Pellets and Plasma Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, Y.; Chen, Z.; Lu, L.; Zhang, L.; Liu, T.; Luo, X.; Liao, X. Determination of Dietary Copper Requirement by the Monoamine Oxidase Activity in Kidney of Broilers from 1 to 21 Days of Age. Anim. Nutr. 2022, 8, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.B.; Kuang, Y.G.; Ma, Z.X.; Liu, Y.G. The Effect of Feeding Broiler with Inorganic, Organic, and Coated Trace Minerals on Performance, Economics, and Retention of Copper and Zinc. J. Appl. Poult. Res. 2020, 29, 1084–1090. [Google Scholar] [CrossRef]
- Brugger, D.; Windisch, W.M. Environmental Responsibilities of Livestock Feeding Using Trace Mineral Supplements. Anim. Nutr. 2015, 1, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Sun, X.; Lu, L.; Lin, X.; Liao, X.; Zhang, L.; Wang, R.; Luo, X. Effect of in Ovo Manganese Injection on the Embryonic Development, Antioxidation, Hatchability, and Performances of Offspring Broilers under Normal and High Temperatures. Poult. Sci. 2022, 101, 101936. [Google Scholar] [CrossRef] [PubMed]
- Horn, D.; Barrientos, A. Mitochondrial Copper Metabolism and Delivery to Cytochrome c Oxidase. IUBMB Life 2008, 60, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.; Gao, L.; Xie, C.; Xiang, Y.; Huang, Y.; Zhang, Y.; Wu, X. Manganese Methionine Hydroxy Analog Chelated Affects Growth Performance, Trace Element Deposition and Expression of Related Transporters of Broilers. Anim. Nutr. 2021, 7, 481–487. [Google Scholar] [CrossRef]
- Underwood, E.J.; Suttle, N.F. The Mineral Nutrition of Livestock, 3rd ed.; CABI Publishing: New York, NY, USA, 1999; ISBN 0851991289. [Google Scholar]
- Bao, Y.M.; Choct, M. Trace Mineral Nutrition for Broiler Chickens and Prospects of Application of Organically Complexed Trace Minerals: A Review. Anim. Prod. Sci. 2009, 49, 269–282. [Google Scholar] [CrossRef]
- Lu, L.; Wang, R.L.; Zhang, Z.J.; Steward, F.A.; Luo, X.; Liu, B. Effect of Dietary Supplementation with Copper Sulfate or Tribasic Copper Chloride on the Growth Performance, Liver Copper Concentrations of Broilers Fed in Floor Pens, and Stabilities of Vitamin e and Phytase in Feeds. Biol. Trace Elem. Res. 2010, 138, 181–189. [Google Scholar] [CrossRef]
- Miles, R.D.; Keefe, S.F.O.; Henry, P.R.; Ammerman, C.B.; Luo, X.G. The Effect of Dietary Supplementation with Copper Sulfate or Tribasic Copper Chloride on Broiler Performance, Relative Copper Bioavailability, and Dietary Prooxidant Activity. Poult. Sci. 1998, 77, 416–425. [Google Scholar] [CrossRef]
- Olukosi, O.A.; Van Kuijk, S.J.A.; Han, Y. Sulfate and Hydroxychloride Trace Minerals in Poultry Diets—Comparative Effects on Egg Production and Quality in Laying Hens, and Growth Performance and Oxidative Stress Response in Broilers. Poult. Sci. 2019, 98, 4961–4971. [Google Scholar] [CrossRef]
- Kim, J.W.; Kil, D.Y. Determination of Relative Bioavailability of Copper in Tribasic Copper Chloride to Copper in Copper Sulfate for Broiler Chickens Based on Liver and Feather Copper Concentrations. Anim. Feed Sci. Technol. 2015, 210, 138–143. [Google Scholar] [CrossRef]
- M’Sadeq, S.A.; Wu, S.B.; Choct, M.; Swick, R.A. Influence of Trace Mineral Sources on Broiler Performance, Lymphoid Organ Weights, Apparent Digestibility, and Bone Mineralization. Poult. Sci. 2018, 97, 3176–3182. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Morgan, N.; Roberts, J.R.; Swick, R.A.; Toghyani, M. Copper Hydroxychloride Is More Efficacious than Copper Sulfate in Improving Broiler Chicken’s Growth Performance, Both at Nutritional and Growth-Promoting Levels. Poult. Sci. 2020, 99, 6964–6973. [Google Scholar] [CrossRef] [PubMed]
- Olukosi, O.A.; Van Kuijk, S.; Han, Y. Copper and Zinc Sources and Levels of Zinc Inclusion Influence Growth Performance, Tissue Trace Mineral Content, and Carcass Yield of Broiler Chickens. Poult. Sci. 2018, 97, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Geng, S.; Yuan, T.; Liu, Y.; Zhang, Y.; Di, Y.; Li, J.; Zhang, L. Effects of Manganese Hydroxychloride on Growth Performance, Antioxidant Capacity, Tibia Parameters and Manganese Deposition of Broilers. Animals 2021, 11, 3470. [Google Scholar] [CrossRef] [PubMed]
- Ruas, F.A.D.; Barboza, N.R.; Castro-Borges, W.; Guerra-Sá, R. Manganese Alters Expression of Proteins Involved in the Oxidative Stress of Meyerozyma Guilliermondii. J. Proteom. 2019, 196, 173–188. [Google Scholar] [CrossRef]
- Jasek, A.; Coufal, C.D.; Parr, T.M.; Lee, J.T. Evaluation of Increasing Manganese Hydroxychloride Level on Male Broiler Growth Performance and Tibia Strength. J. Appl. Poult. Res. 2019, 28, 1039–1047. [Google Scholar] [CrossRef]
- Adele, B.O.; Idama, C.; Ige, A.O.; Odetola, A.O.; Emediong, I.E.; Adewoye, E.O. Alterations in Plasma and Erythrocyte Membrane Fatty Acid Composition Following Exposure to Toxic Copper Level Affect Membrane Deformability and Fluidity in Female Wistar Rats. J. Trace Elem. Med. Biol. 2023, 80, 127316. [Google Scholar] [CrossRef]
- Adele, B.O.; Ojo, G.T.; Ige, A.O.; Odetola, A.O.; Emediong, I.E.; Adewoye, E.O. Toxic Copper Level Increases Erythrocyte Glycolytic Rate, Glutathione Production and Alters Electrolyte Balance in Male Wistar Rats. J. Trace Elem. Med. Biol. 2023, 79, 127231. [Google Scholar] [CrossRef]
- Lei, C.; Huo, Y.; Ma, F.; Liao, J.; Hu, Z.; Han, Q.; Li, Y.; Pan, J.; Hu, L.; Guo, J.; et al. Long-Term Copper Exposure Caused Hepatocytes Autophagy in Broiler via MiR-455-3p-OXSR1 Axis. Chem. Biol. Interact. 2023, 369, 110256. [Google Scholar] [CrossRef]
- Liao, J.; Yang, F.; Yu, W.; Qiao, N.; Zhang, H.; Han, Q.; Hu, L.; Li, Y.; Guo, J.; Pan, J.; et al. Copper Induces Energy Metabolic Dysfunction and AMPK-MTOR Pathway-Mediated Autophagy in Kidney of Broiler Chickens. Ecotoxicol. Environ. Saf. 2020, 206, 111366. [Google Scholar] [CrossRef]
- Fasano, M.; Curry, S.; Terreno, E.; Galliano, M.; Fanali, G.; Narciso, P.; Notari, S.; Ascenzi, P. The Extraordinary Ligand Binding Properties of Human Serum Albumin. IUBMB Life 2005, 57, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Siotto, M.; Squitti, R. Copper Imbalance in Alzheimer’s Disease: Overview of the Exchangeable Copper Component in Plasma and the Intriguing Role Albumin Plays. Coord. Chem. Rev. 2018, 371, 86–95. [Google Scholar] [CrossRef]
- Kaplan, I.V.; Attaelmannan, M.; Levinson, S.S. Fibrinogen Is an Antioxidant That Protects β-Lipoproteins at Physiological Concentrations in a Cell Free System. Atherosclerosis 2001, 158, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Mañucat-Tan, N.; Zeineddine Abdallah, R.; Kaur, H.; Saviane, D.; Wilson, M.R.; Wyatt, A.R. Hypochlorite-Induced Aggregation of Fibrinogen Underlies a Novel Antioxidant Role in Blood Plasma. Redox Biol. 2021, 40, 101847. [Google Scholar] [CrossRef] [PubMed]
- Hahl, P.; Hunt, R.; Bjes, E.S.; Skaff, A.; Keightley, A.; Smith, A. Identification of Oxidative Modifications of Hemopexin and Their Predicted Physiological Relevance. J. Biol. Chem. 2017, 292, 13658–13671. [Google Scholar] [CrossRef]
- Kelly-Robinson, G.A.; Reihill, J.A.; Lundy, F.T.; McGarvey, L.P.; Lockhart, J.C.; Litherland, G.J.; Thornbury, K.D.; Martin, S.L. The Serpin Superfamily and Their Role in the Regulation and Dysfunction of Serine Protease Activity in Copd and Other Chronic Lung Diseases. Int. J. Mol. Sci. 2021, 22, 6351. [Google Scholar] [CrossRef]
- Adair, J.E.; Stober, V.; Sobhany, M.; Zhuo, L.; Roberts, J.D.; Negishi, M.; Kimata, K.; Garantziotis, S. Inter-α-Trypsin Inhibitor Promotes Bronchial Epithelial Repair after Injury through Vitronectin Binding. J. Biol. Chem. 2009, 284, 16922–16930. [Google Scholar] [CrossRef]
- Chen, X.; Nakada, S.; Donahue, J.E.; Chen, R.H.; Tucker, R.; Qiu, J.; Lim, Y.P.; Stopa, E.G.; Stonestreet, B.S. Neuroprotective Effects of Inter-Alpha Inhibitor Proteins after Hypoxic-Ischemic Brain Injury in Neonatal Rats. Exp. Neurol. 2019, 317, 244–259. [Google Scholar] [CrossRef]
- Garantziotis, S.; Hollingsworth, J.W.; Ghanayem, R.B.; Timberlake, S.; Zhuo, L.; Kimata, K.; Schwartz, D.A. Inter-Trypsin Inhibitor Attenuates Complement Activation and Complement-Induced Lung Injury 1. J. Immunol. 2007, 179, 4187–4192. [Google Scholar] [CrossRef]
- Rostagno, H.S.; Albino, L.F.T.; Hannas, M.I.; Donzele, J.L.; Sakomura, N.K.; Alysson, F.G.P.; Saraiva, A.; Teixeira, M.L.; Rodrigues, P.B.; de Oliveira, R.F.; et al. Tabelas Brasileiras Para Aves e Suínos, 4th ed.; Rostagno, H.S., Ed.; Departamento de Zootecnia—UFV: Viçosa, Brazil, 2017; ISBN 9788581791203. [Google Scholar]
- Doumas, B.T.; Bayse, D.D.; Carter, R.J.; Peters, J.R.; Schaffer, R.A. Candidate Reference Method for Determination of Total Protein in Serum. I. Development and Validation. II. Tests for Transferability. Clin. Chem. 1981, 27, 1651–1654. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.S.; De Magalh, C.S.; Aur, M.; Arruda, Z. Trends in Metal-Binding and Metalloprotein Analysis. Talanta 2006, 69, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Braga, C.P.; Vieira, J.C.S.; Grove, R.A.; Boone, C.H.T.; Leite, A.D.L.; Buzalaf, M.A.R.; Fernandes, A.A.H.; Adamec, J.; Padilha, P.D.M. A Proteomic Approach to Identify Metalloproteins and Metal-Binding Proteins in Liver from Diabetic Rats. Int. J. Biol. Macromol. 2017, 96, 817–832. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cavecci, B.; de Lima, P.M.; Vieira, J.C.S.; Braga, C.P.; de Queiroz, J.V.; Bittarello, A.C.; de Padilha, P.M. Use of Ultrasonic Extraction in Determining Apparent Digestibility in Fish Feed. J. Food Meas. Charact. 2015, 9, 599–603. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Cu Concentration | Mn Concentration |
|---|---|---|
| Plasma (mg L−1) | ||
| S15-80 | 2.65 ± 0.07 c | 5.25 ± 0.07 d |
| S150-120 | 23.40 ± 0.28 a | 16.85 ± 0.07 b |
| H15-80 | 5.75 ± 0.07 b | 7.85 ± 0.07 c |
| H150-120 | 20.70 ± 0.14 a | 20.45 ± 0.07 a |
| Protein pellet (mg kg−1) | ||
| S15-80 | 1.67 ± 0.12 c | 3.15 ± 0.09 d |
| S150-120 | 37.57 ± 0.97 a | 25.33 ± 0.75 b |
| H15-80 | 3.57 ± 0.42 b | 4.30 ± 0.10 c |
| H150-120 | 40.13 ± 0.61 a | 32.70 ± 0.82 a |
| Spot ID | Accession | Protein | “Fold-Change” | |||
|---|---|---|---|---|---|---|
| H15-80 versus S15-80 | H150-120 versus S150-120 | H150-120 versus H15-80 | S150-120 versus S15-80 | |||
| 3 | R9PXM5 | Immunoglobulin-lambda-like polypeptide 1 | −1.362/+1.362 | |||
| A0A3Q2UDC8 | Ig-like domain-containing protein | −1.362/+1.362 | ||||
| 13 | O93601 | Apolipoprotein AIV | −1.325/+1.325 | |||
| 25 | E1C7T1 | Serpin peptidase inhibitor_ clade A (alpha-1 antiproteinase_ antitrypsin)_ member 1 | +1.424/−1.424 | |||
| E1BV78 | Fibrinogen gamma chain | +1.424/−1.424 | ||||
| 26 | F1P4V1 | Fibrinogen alpha chain | +2.438/−2.438 | |||
| 28 | F1NUL9 | Fibrinogen beta chain | −1.478/+1.478 | |||
| 31 | Q90WR3 | Hemopexin (Fragment) | −1.740/+1.740 | |||
| 38 | F1NWX6 | Plasminogen | −1.625/+1.625 | −1.647/+1.647 | ||
| Q7LZF3 | Plasmin | −1.625/+1.625 | −1.647/+1.647 | |||
| 44 | F1NUL9 | Fibrinogen beta chain | +1.919/−1.919 | −1.339/+1.339 | −1.753/+1.753 | |
| Q9W6F5 | Vitamin D-binding protein | +1.919/−1.919 | −1.339/+1.339 | −1.753/+1.753 | ||
| 48 | Q90633 | Complement C3 | −1.328/+1.328 | |||
| F1DQG4 | Complement component 7 | −1.328/+1.328 | ||||
| A6N9E0 | Complement component 3d (Fragment) | −1.328/+1.328 | ||||
| 50 | A0A3Q2U3V9 | Beta-microseminoprotein-like | −1.626/+1.626 | |||
| Q7LZS1 | 12K serum protein_ beta-2-m cross-reactive (Fragment) | −1.626/+1.626 | ||||
| 54 | A0A1D5NXA6 | Inter-alpha-trypsin inhibitor heavy chain 3 | +1.373/−1.373 | |||
| B3VE14 | Inter-alpha inhibitor heavy chain 2 | +1.373/−1.373 | ||||
| A0A1D5PU00 | Protein AMBP | +1.373/−1.373 | ||||
| 56 | A0A3Q2UFG5 | Ig-like domain-containing protein | −1.426/+1.426 | |||
| A2N881 | VH1 protein | −1.426/+1.426 | ||||
| A0A1D5NW68 | Albumin | −1.426/+1.426 | ||||
| F1NJU5 | Complement C8 alpha chain | −1.426/+1.426 | ||||
| 59 | A0A1I7Q422 | Transthyretin | −1.349/+1.349 | |||
| 64 | A0A1D5PW77 | C-reactive protein_ pentraxin-related | +2.030/−2.030 | |||
| Q2EJU6 | Pentraxin | +2.030/−2.030 | ||||
| 67 | F1NW43 | Pyruvate kinase | +1.802/−1.802 | |||
| F1NUL9 | Fibrinogen beta chain | +1.802/−1.802 | ||||
| 68 | F1NUL9 | Fibrinogen beta chain | +2.262/−2.262 | |||
| F1NW43 | Pyruvate kinase | +2.262/−2.262 | ||||
| A0A1D5PNU2 | Beta-2-glycoprotein 1 | +2.262/−2.262 | ||||
| F1P4V1 | Fibrinogen alpha chain | +2.262/−2.262 | ||||
| 70 | A0A3Q2UFG5 | Ig-like domain-containing protein | +2.414/−2.414 | |||
| A0A1D5NW68 | Albumin | +2.414/−2.414 | ||||
| A2N881 | VH1 protein | +2.414/−2.414 | ||||
| F1ND07 | Proteasome activator subunit 4 | +2.414/−2.414 | ||||
| 71 | A0A3Q2UFG5 | Ig-like domain-containing protein | +5.128/−5.128 | |||
| A0A1D5NW68 | Albumin | +5.128/−5.128 | ||||
| A2N881 | VH1 protein | +5.128/−5.128 | ||||
| 72 | Q4ADJ7 | Ovotransferrin | −1.394/+1.394 | |||
| 73 | A0A3Q2UFG5 | Ig-like domain-containing protein | −1.439/+1.439 | |||
| A2N881 | VH1 protein | −1.439/+1.439 | ||||
| 75 | F1NWX6 | Plasminogen | +1.741/−1.741 | |||
| Q7LZF3 | Plasmin | +1.741/−1.741 | ||||
| 79 | A0A3Q2U3V9 | Beta-microseminoprotein-like | +1.382/−1.382 | |||
| Q7LZS1 | 12K serum protein_ beta-2-m cross-reactive (Fragment) | +1.382/−1.382 | ||||
| 98 | A0A1D5NW68 | Albumin | +2.293/−2.293 | |||
| 100 | A0A1L1RIW5 | Keratin 8 | −2.011/+2.011 | |||
| 104 | A0A1D5PNU2 | Beta-2-glycoprotein 1 | −1.507/+1.507 | |||
| F1NUL9 | Fibrinogen beta chain | −1.507/+1.507 | ||||
| F1P4V1 | Fibrinogen alpha chain | −1.507/+1.507 | ||||
| F1NW43 | Pyruvate kinase | −1.507/+1.507 | ||||
| A0A1L1RIW5 | Keratin 8 | −1.507/+1.507 | ||||
| 105 | E1BV78 | Fibrinogen gamma chain | −1.909/+1.909 | |||
| A0A1D5NW68 | Albumin | −1.909/+1.909 | ||||
| F1ND07 | Proteasome activator subunit 4 | −1.909/+1.909 | ||||
| 106 | Q98TD1 | PIT 54 | −1.308/+1.308 | |||
| A0A3Q2UF48 | Peptidase S1 domain-containing protein | −1.308/+1.308 | ||||
| E1BV78 | Fibrinogen gamma chain | −1.308/+1.308 | ||||
| A0A1D5NW68 | Albumin | −1.308/+1.308 | ||||
| 107 | A0A3Q2UFG5 | Ig-like domain-containing protein | −2.109/+2.109 | |||
| 108 | F1NAR5 | Serpin family F member 2 | −1.703/+1.703 | |||
| A0A1L1RIW5 | Keratin 8 | −1.703/+1.703 | ||||
| 111 | F1NK40 | Alpha-2-macroglobulin-like 4 | −1.425/+1.425 | |||
| E1BV78 | Fibrinogen gamma chain | −1.425/+1.425 | ||||
| H1AC38 | A2M_recep domain-containing protein (Fragment) | −1.425/+1.425 | ||||
| 112 | Q90WR3 | Hemopexin (Fragment) | −2.550/+2.550 | |||
| A0A1D5PEU7 | Vanin 1 | −2.550/+2.550 | ||||
| Q5ZHM4 | CN hydrolase domain-containing protein | −2.550/+2.550 | ||||
| Experimental Groups | Cu (mg kg−1) | Mn (mg kg−1) | |
|---|---|---|---|
| Sulfates | S15-80 | 15 | 80 |
| S150-120 | 150 | 120 | |
| Hydroxychlorides | H15-80 | 15 | 80 |
| H150-120 | 150 | 120 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, R.A.; Assunção, A.S.d.A.; Vieira, J.C.S.; Rocha, L.C.; Urayama, P.M.G.; Buzalaf, M.A.R.; Sartori, J.R.; Padilha, P.d.M. Proteomic Study of Broiler Plasma Supplemented with Different Levels of Copper and Manganese from Different Sources. Molecules 2023, 28, 8155. https://doi.org/10.3390/molecules28248155
Martins RA, Assunção ASdA, Vieira JCS, Rocha LC, Urayama PMG, Buzalaf MAR, Sartori JR, Padilha PdM. Proteomic Study of Broiler Plasma Supplemented with Different Levels of Copper and Manganese from Different Sources. Molecules. 2023; 28(24):8155. https://doi.org/10.3390/molecules28248155
Chicago/Turabian StyleMartins, Renata Aparecida, Andrey Sávio de Almeida Assunção, José Cavalcante Souza Vieira, Leone Campos Rocha, Priscila Michelin Groff Urayama, Marília Afonso Rabelo Buzalaf, José Roberto Sartori, and Pedro de Magalhães Padilha. 2023. "Proteomic Study of Broiler Plasma Supplemented with Different Levels of Copper and Manganese from Different Sources" Molecules 28, no. 24: 8155. https://doi.org/10.3390/molecules28248155
APA StyleMartins, R. A., Assunção, A. S. d. A., Vieira, J. C. S., Rocha, L. C., Urayama, P. M. G., Buzalaf, M. A. R., Sartori, J. R., & Padilha, P. d. M. (2023). Proteomic Study of Broiler Plasma Supplemented with Different Levels of Copper and Manganese from Different Sources. Molecules, 28(24), 8155. https://doi.org/10.3390/molecules28248155