
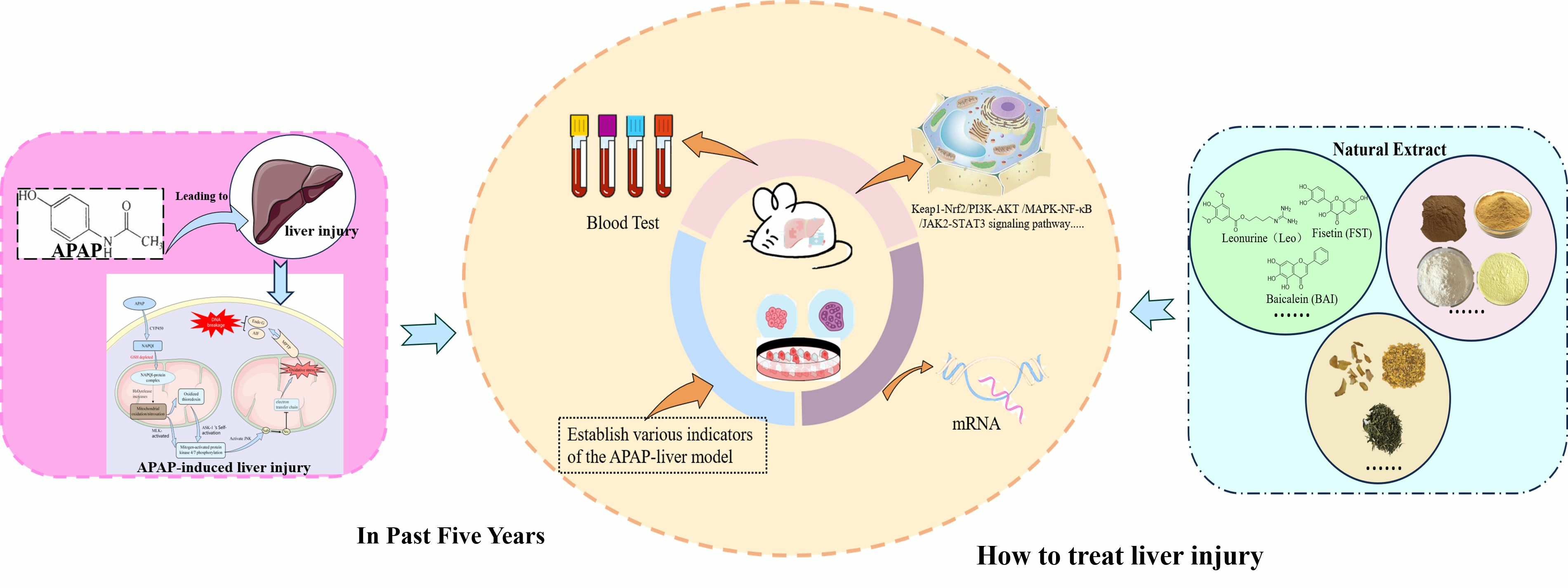
Natural Products for Acetaminophen-Induced Acute Liver Injury: A Review

 , and
, and
Abstract
:
1. Introduction
2. Potential Mechanisms of APAP-Induced Acute Liver Injury
3. Models of APAP-Induced Acute Liver Injury
3.1. In Vivo Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex, Strain, Age/Body, Weight | Dose of APAP | Pre Administration or Treatment | Administration Frequency | Administration | Experimental Index | No. Reference |
|---|---|---|---|---|---|---|
| Male C57BL/6J mice 8–10 weeks | 300, 500 mg/kg | Fasted overnight | Once | i.p. | Protein (Pro-CRAMP, CRAMP, CD11b, CYP2E1, JNK, p-JNK, Cyclin D1), Serum Index (ALT), Histology, Tissues (ROS, GSH), Cell quantification (Ki67 cells, neutrophils and macrophages) | Zhai, et al., 2023 [28] |
| Male C57BL/6J mice 6–8 weeks | 300, 500 mg/kg | Fasted for 15–17 h | Once | i.p. | Serum Index (ALT, AST, TNF-α, IL-6, MCP-1, mtDNA), Histology, Tissues (MDA, Caspase-3), Protein (P2RX1, BCL-2, BCL-X, STING, p-STING, TBK1, p-TBK1, P65, p-P65), RNA (P2RX1, TNF-α, IL-6, MCP-1) | Yu, et al., 2023 [29] |
| Male C57BL/6J mice 6–8 weeks | 250, 450, 550 mg/kg | Fasted for 16 h | Once | i.p. | Serum Index (ALT, AST, cfDNA, HMGB1, TNF-α, MCP-1), Histology, Cell quantification (leukocytes, neutrophils), Tissues (MDA, GSH-Px), RNA (Tnf, Il1β, Mmp3, Sphk1, Alox12, and Nqo1, TNF-α, TGF-β, 3-NT) | Sun, et al., 2023 [30] |
| Male C57BL/6N mice 4–8 weeks | 300, 600 mg/kg | Fasted overnight | Once | i.p. | Serum Index (ALT, AST, LDH, HMGB1), Protein (NEDD4-1, P-JNK, JNK, Bax, CYPD, VDAC1, COX IV, AIF, Endo G, Cyt C), RNA (NEDD4-1, Vdac1), Histology, Gene sets, Tissues (ROS, ATP) | Zhu, et al., 2023 [31] |
| Male C57BL/6N mice 6–8 weeks 18–22 g | 250 mg/kg | Fasted for 12 h but free water | Once | i.p. | Serum Index (ALT, AST), Tissues (GSH), mRNA (cGAS, STING, IFN-β1), Protein (ST2) | Wang, et al., 2023 [32] |
| Male C57BL/6 mice 5–7 weeks 18–22 g | 500 mg/kg | Fasted overnight | Once | i.p. | Histology (mitochondria), Protein (γH2AX, p-JNK, JNK, p-Src, Src, p-ATM, ATM, γH2AX, H2AX, p21), Serum Index (ALT, AST, GSH), Tissues (ATP, ROS) | Cen, et al., 2023 [33] |
| Male C57BL/6 mice 8 weeks | 300 mg/kg | - | Once | p.o. for 24 h | Plasma Index (ALT, AST, TNF-α, IL-6, MCP-1, MCP-3, mtDNA), Tissues (SOD, CAT, GSSG, GSH, GSH-Px, MDA, NAPQI, ROS, Fe2+), DA, β-galactosidase, β-glucosidase, LDH, Histology, Lactobacillus species, Cell quantification (cell death, neutrophils, macrophages), Protein (CYP2E1, CYP1A2, PCNA, p-ASK, p-MKK4, p-JNK, GPX4, xCT), mRNA (Ptgs2, Fdps) | Zeng, et al., 2023 [34] |
| Male C57BL/6J mice 8 weeks | 200, 400, 600 mg/kg | Fasted for 16 h but free water | Once | i.p. | Plasma Index (ALT, AST, miR122), Histology, mRNA (Cytochrome P450, HNF1AOS1), Enzyme Activities (CYP3A11, 1A2, 2B10, 2C29, 2E1), Sedation time | Bao, et al., 2022 [35] |
| Male C57BL/6 mice 8–9 weeks 18–22g | 300 mg/kg | Fasted overnight | Once | i.p. | Plasma Index (ALT, AST, ALB, TBIL), Histology, Tissues (MPO), mRNA (CCR 2, CCR 5, CXCL 9, CXCR 2, IL-1β, IL-6, TNF-α, CAT, GSH-PX, T-SOD), Protein (p-JAK2, JAK2, p-STAT3, STAT3, BAX, BCL-2, p-p65, p65) | He, et al., 2022 [36] |
| Male C57BL/6J mice 8–10 weeks | 300 mg/kg | Fasted overnight | Once | i.p. | Serum Index (ALT, MANF), Macrophages, Neutrophils, Histology, mRNA (Tnfa, Il1b, Il6, Ifna, Il10), Protein (p-AKT, AKT, p-JNK, JNK, p-p38, p-38, p-ERK, ERK, p-SAPK, SAPK), RNA (Gpnmb, Axl, Cd36, Cd5l, Macro, Mertk, Cd81, Trem2, CD36, CD64, THBS1, SIRPA, MSR1) | Hou, et al., 2022 [37] |
| C57BL/6J mice 6–8 weeks | 300, 750 mg/kg | Fasted for 12 h | Once | i.p. | Serum Index (ALT, AST, LDH, CK18-M30, CK18-M65, EGR1), Histology, Tissues (GSH, TG), mRNA (Egr1, Acaa2), Protein (Egr1, CYP2E1, HSP90) | Lei, et al., 2022 [38] |
| Male C57BL/6N mice 18–28 g | 250 mg/kg | Fasted for 12 h | Once | i.p. | Serum Index (ALT, AST, GSH, TNF-α, IL-6, IL-10), Histology, Protein (IL-33, IL-6, IL-1β, AMPKα, p-AMPKα, PI3K, p-PI3K, MEK, ERK, p-ERK, Akt, p-Akt, Beclin-1, LC3I, LC3II), mRNA (IL-33, ST2, Cyp1A2, Cyp2E1, TNF-α, IL-6, IL-10, iNOS, IL-12, Arg-1, IL-10) | Wang, et al., 2021 [39] |
| Male C57BL/6J mice 2–3 months | 500 mg/kg | - | Once | Feed or i.p. | Serum Index (ALT, VWF), Histology, Protein (p62, LC3II, CYP2E1, APAP-AD, p-p62, GCLC, GCLM, p-JNK, T-JNK, S6, p-S6, 4EBP1, p-4EBP1, Cyclin D1, PCNA, P21), mRNA (p62/Sqstm1, Atg8/LC3B, Gclc, Gclm), Tissues (GSH, APAP-Cys) | Qian, et al., 2021 [40] |
| Male and female C57BL/6J mice 8–12 weeks | 210 mg/kg (male) 325 mg/kg (female) | Fasted overnight | Once | i.p. | Serum Index (ALT, Chi3l1), Histology, Protein (CYP2E1, Chi3l1, CD44, His, NAPQI), Tissues (GSH), Mϕs, Platelet | Shan, et al., 2021 [41] |
| Male C57BL/6 mice 10 weeks | 300 mg/kg | Fasted for 24 h | Once | i.g. | Serum Index (ALT, AST, IL-1β, IL-18, LDH), Histology, Protein (PRX3, NLRP3, GSDMD, Caspase-1, Cleaved Caspase-1, IL-1β, IL-18, PRX5, PRX6), mRNA (PRX3) | Wang, et al., 2021 [42] |
| Male C57BL/6 mice 8–12 weeks | 250 mg/kg | Fasted overnight for 12 h | Once | i.p. | Plasma Index (ALT, sPD-L1, AST, TNF-α, IL-6, IL-10), Histology, Cell quantification (Macrophages, KCs, E. coli), Protein (PD-1, PD-L1), mRNA (Pdl1) | Triantafyllou, et al., 2021 [43] |
| Male C57BL/6 mice 6–8 weeks 19–20 g | 400 mg/kg | Free food and water | Once | i.p. | Histology, Serum Index (ALT, AST), Protein (Total SIRT6, Nuclear SIRT6, SIRT1-7) | Zhou, et al., 2021 [44] |
| Male C57BL/6 mice | 350 mg/kg | - | Once | i.p. | Histology, Serum Index (ALT, AST), Tissues (GSH, MDA, ROS), Oil Red O, Protein (SIRT1, GPX4, NRF2, HO-1) | Wang, et al., 2021 [45] |
| Male Kunming mice 20–25 g | 400 mg/kg | Fasted overnight | Once | i.p. | Serum Index (ALT, AST, IL-1β, TNF-α, IL-6), Tissues (SOD, CAT, GSH, MDA), Histology, Protein (Nrf2, NQO1, HO-1, p-p38, p38, p-p65, p65, iNOS, Bcl-2, Bax, Caspase-3, Caspase-9) | Wang, et al., 2021 [46] |
| Male and female C57BL/6J-Tg mice 3 months | 500 mg/kg | - | Once | i.p. | Serum Index (ALT, AST), Histology, Mitochondrial function assay, Lysophosphatidylcholine, iPLA2, Protein (PRDX6, PRDX6-SO3, JNK, pJNK, Bax, Bcl-2) | Lee, et al., 2020 [47] |
| Male C57BL/6 J mice 8–10 weeks | 200, 250, 300, 700 mg/kg | Fasted overnight for 12 h | Once | i.p. | Serum Index (ALT, AST, TNF-α, IL-1β, IL-6), Histology, Protein (SPHK1, SPHK2, IRE1α, STAT1, p65, CYP2E1, CHOP, p-IRE1α, IRE1α, PERK, p-PERK, p-elF2α, elF2α, ATF4, ATF6, p-JNK, JNK, p-ASK1, ASK1, p-GSK3β, GSK3β, p38, TRAF2), Tissues (GSH), Cell quantification (neutrophils, macrophages), mRNA (TNF-α, IL-1β, IL-6, Ccl2, Ccl3, Cxcl1, Cxcl2) | Li, et al., 2020 [48] |
| Male C57BL/6J mice 6–8 weeks | 300 mg/kg | Fasted overnight, but free water | Once | i.p. | Serum Index (ALT), Protein (Nqo1, AKR1C, Gstα3, Gstm1, Gstm5, Nrf2, p-JNK, JNK), mRNA (NQO1, AKR1C), Histology | Chen, et al., 2020 [49] |
| Male and female mice 8–10 weeks | 210 mg/kg (male) 325 mg/kg (female) | Fasted overnight for 18 h | Once | i.p. | Histology, Serum Index (ALT, IL-6, TGF-β, IL-13), Tissues (GSH, TNF-α, IL-1β, IL-4, IL-10, TGF-β, IL-13), Cell quantification (macrophages, neutrophils), Protein (HIF-2α, CYP2E1), mRNA (PAI-1, ADM, VEGF, IL-6, IL-1β, IL-4, IL-10, MCP-1, IFN-γ, TNF-α) | Gao, et al., 2020 [50] |
| Male mice 8–10 weeks | 300, 500 mg/kg | Fasted overnight | Once | i.p. | Histology, Serum Index (ALT, AST, CCL5), mRNA (Ccl5, CD206, Ym1, Arg1, iNOS, IL-1β, TNF-α), Tissues (GSH), Protein (PCNA, CD206, Ym1, Arg1, p-Erk1/2, Erk1/2, p-JNK, JNK, p-NF-κB, NF-κB, IκBα, p-IκB, CCR1, CCR5) | Li, et al., 2020 [51] |
| Male mice 8–10 weeks | 300, 500 mg/kg, 750 mg/kg (No fasted) | Fasted for 15–17 h | Once | i.p. | Histology, Serum Index (ALT, AST, TNF-α, IL-6), Protein (OPN, JNK, p-JNK, CYP2E1), Tissues (MDA), Ketone, mtDNA, Cell quantification (neutrophils) | Wen, et al., 2019 [52] |
| Male C57BL/6J mice 8–12 weeks | 500 mg/kg | Free food and water | Once | i.p. | Serum Index (ALT), Histology, Tissues (GSH, APAP-protein adducts), mitochondria bioenergetics, Protein (Ub, p62, Mfn1, Tom20) | Wang, et al., 2019 [53] |
| C57BL/6J mice | 250, 500 mg/kg | Fasted overnight | Once | i.p. | Serum Index (ALT, AST), Protein (p-JNK, JNK, PUMA, Endo G, AIF, Cyt c, Drp1, Bax, tBid, Bim, Bcl-XL, p53), mRNA (PUMA), Tissues (GSH, Caspase-3, Caspase-7), Histology | Chen, et al., 2019 [54] |
3.2. In Vitro Models
| Type | Dose of APAP | Pre Administration or Treatment | Administration Frequency | Medication Processing Time | Experimental Index | No. Reference |
|---|---|---|---|---|---|---|
| Primary hepatocytes | 5 mM | Starve for 12 h | Once | For 6 h | Mitochondria (ROS, membrane potential), Cell quantification (TUNEL-positive primary hepatocytes, CD11b-positive cells, MPO-positive cells) | Yu, et al., 2023 [29] |
| Primary hepatocytes | 10 mM | Once | For 12 h | Protein (NEDD4-1, VDAC1), LDH, HMGB1, mtROS | Zhu, et al., 2023 [31] | |
| HepaRG cells | 10 mM | Gene knockout | Once | For 12 h | Protein (Beclin-1, LC3I/LC3II, STING, IRF3, p-IRF3, IB: Flag, IB: HA, Myc-ST2, Flag, HA, HA-STING, Flag-TBK1, ATG5), mRNA (IFN-β1, ISG54, ISG56) | Wang, et al., 2023 [32] |
| AML-12 hepatocytes | 10 mM | Incubated | Once | For 24 h | Protein (JNK, p-JNK, p-ATM, ATM, γH2AX, H2AX), ROS | Cen, et al., 2023 [33] |
| AML-12 hepatocytes and primary hepatocytes | 5 mM | Incubated | Once | For 24 h | Cell viability, ROS, GSH, GSH/GSSG, mRNA (Fdps), Protein (GPX4, xCT, p-AKT, p-GSK3β, Nrf2), Fe2+ | Zeng, et al., 2023 [34] |
| Hepatocytes | 10 mM | Incubated | Once | For 6 h | Cell viability, ROS, Cell death evaluation, Protein (p-JAK2, JAK2, p-STAT3, STAT3, BAX, BCL-2), | He, et al., 2022 [36] |
| Primary hepatocytes | 20 mM | Incubated | Once | - | Cell viability, Seahorse XFe96 metabolic flux, FFA, mRNA (Acaa2), Genome-wide analysis, Protein (CYP2E1), ND-1, FFAs, OCR, Luciferase activity assay (Acaa2 N-terminal promoter, truncated N-terminal promoters, WT, mutant N-2 promoters) | Lei, et al., 2022 [38] |
| AML-12 cells | 10 mM | Incubated | Once | For 6 h | ||
| Primary hepatocytes | 10 mM | Incubated | Once | For 12 h | Protein (PRX3, NLRP3, GSDMD, Caspase-1, Cleaved Caspase-1, IL-1β, IL-18, PRX5, PRX6), Mitochondrial ROS, LDH | Wang, et al., 2021 [42] |
| AML-12 cells | 5 mM | Transfection | Once | For 48 h | Cell viability, ROS, Cell proliferation, LDH, ROS, GSH, SOD, Protein (CCNA1, CCND1, CDK4, total NRF2, nuclear NRF2, pNRF2), mRNA (Nrf2, Ho-1, Gstα, Gstμ) | Zhou, et al., 2021 [44] |
| L-02 cells | 10 mM | Incubate | Once | For 24 h | ROS, Protein (SIRT1, GPX4, NRF2, HO-1, Histone-H3) | Wang, et al., 2021 [45] |
| HepG2 cells | 60 μM | Incubated | Once | - | Cell survival rate, Apoptosis rate, Serum Index (MAD, GSH, SOD), mRNA (Bcl-2, Bax, Caspase-3), Protein (Bcl-2, Bax, Caspase-3) | Zhao, et al., 2020 [57] |
| Huh7 cell | 10 mM | Transfection | Once | For 24 h | Cell survival assay, Lysophosphatidylcholine, Protein (PRDX6), iPLA2, LPC | Lee, et al., 2020 [47] |
| AML-12 cells, primary hepatocytes | 10 mM | Incubated | Once | For 4–8 h | Cell death determination, ROS, TNF-α, IL-1β, IL-6, Protein (p-JNK, JNK, Bcl-2, Bax, VDAC1, p-ASK1, ASK1, p-GSK3β, GSK3β, p-TRAF2, TRAF2, IκBα, p-IκB, p65, HDAC4, HDAC5, HDAC7) | Li, et al., 2020 [48] |
4. Natural Product for APAP-Induced Acute Liver Injury
4.1. The Physiological Function of Natural Products
4.1.1. Oxidative Stress


4.1.2. Metabolic Pathways

4.1.3. Immunomodulation

4.2. Compounds
4.2.1. Nrf2-Related Pathways

4.2.2. NF-κB-Related Pathways


4.2.3. MAPK-Related Pathways


4.2.4. PI3K/Akt-Related Pathways

4.3. Natural Product Extracts

| Phytochemicals | Source | Dose, Route, and Duration of Administration | Model | APAP Usage | Findings and Key Mechanisms | Reference |
|---|---|---|---|---|---|---|
| 70% Ethanol Extract | Mulberry leaf, Morus alba L. | 400 mg/kg, p.o. for 28 days | Male Sprague–Dawley rats | 500 mg/kg, p.o. | Decreased CASP3, CYP2E1 immunoexpression, and antioxidant and anti-apoptotic properties. | Fadil, et al., 2023 [160] |
| 70% Ethanol Extract | Olive leaf | 400 mg/kg, p.o. for 28 days | ||||
| 80% Methanolic Extract | A. Americana | 100, 200, and 400 mg/kg, p.o. for 7 days | Male Wistar albino rats | 2 mg/kg, p.o. single dose | Mechanism not explored. | Ayenew, et al., 2023 [161] |
| Aqueous Extract | Curcumin, Curcuma aromatica Salisb | 1, 10, 25, 50, 100, 200, and 400 μg/mL for the Primary Cell | C57BL/6 mice Hepatocytes | 2 mM | Increased the protein expression of Sirt1 and HO-1 vand increased the mRNA expression of these genes in liver tissue. | Kim, et al., 2023 [162] |
| 20, 100 mg/kg, i.p. once daily for 1 week | Male C57BL/6 mice | 300 mg/kg, i.p. single dose | ||||
| 80% Ethanol Extract | Polygoni Multiflori (Thunb.) Nakai, Polygoni Multiflori Radix | 100, 200 mg/kg, p.o. for 21 days | Male Kunming mice | 400 mg/kg, p.o. single dose | Upregulating antioxidant enzymes and repressing lipid peroxidation. | Wang, et al., 2023 [163] |
| Aqueous Extract | Smilax china L. | 300, 600 mg/kg, p.o. for 14 days | Male BALB/c mice | 400 mg/kg, p.o. single dose | Activated the Nrf2-ARE signaling pathway and inhibited oxidative stress. | Wang, et al., 2022 [164] |
| Petroleum and Ethanol Extract | Broussonetia papyrifera | 100, 200, and 400 mg/kg, i.g. daily for 14 days | Male Kunming mice | 200 mg/kg, i.g. single dose | Remodeling of intestinal flora, activation of Nrf2 pathway, inhibition of apoptosis. | Xu, et al., 2022 [165] |
| Aqueous Extract | Amblygonocarpus andongensis | 125, 250, and 500 mg/kg, p.o. daily for 8 days | Male Wistar rats | 1 g/kg, p.o. single dose | Inhibited oxidative stress and promoted oxygen radical scavenging. | Baponwa, et al., 2022 [166] |
| Ethanol Extract | Ficus exasperata | 150, 250, and 500 mg/kg i.p. for 5 days | Albino rats | 250 mg/kg, i.p. single dose | Mechanism not explored. | Adetuyi, et al., 2022 [167] |
| 80% Methanol Extracts | C. africana Lam, Boraginaceae | 100, 200, or 400 mg/kg, i.g. daily for ten days | Rats | 2 g/kg, p.o. single dose | Anti-inflammatory and antioxidant activities of the plant. | Geresu, et al., 2022 [168] |
| Aqueous Extract | ||||||
| Aqueous Extract | Raphanus sativus L.var niger, Radish | 500, 1000 mg/kg, p.o. once daily for 4 weeks | Male BALB/c mice | 500 mg/kg, i.p. single dose | Regulating antioxidant and anti-apoptotic signaling factors. Increased the expression of Nrf-2 and HO-1, and regulated the Bax and Bcl-2. | Hwang, et al., 2022 [153] |
| 65% Methanol Extract | Galium aparine L. plants, Yogurt herb | Rats that drank tap water containing 250, 500 mg/kg for five days | Wistar albino rats | 1500 mg/kg, p.o single dose | Improving the serum levels of liver enzymes and liver histology changes. | Sahin, et al., 2022 [169] |
| 80% Aqueous Methanol Extract | Melaleuca rugulosa (Link) Craven | 250, 500, and 1000 mg/kg, p.o. for 7 days, once a day | Sprague–Dawley male rats | 3 g/kg, p.o. single dose | Oxidative stress-mediated activation of the JNK pathway in liver tissue. | Elsayed, et al., 2022 [170] |
| Aqueous Methanol Extract | Paspalidium flavidum, Watercrown grass | 250 and 500 mg/kg, p.o. once a day for 7 days | Wistar rats (both male and female) | 200 mg/kg, i.p. single dose, | hepatoprotective and gastroprotective, antioxidant properties. | Ismail, et al., 2022 [171] |
| 20% Methanolic Extract | Piper chaba Hunter, Chui Jhal | 250 and 500 mg/kg, p.o. once a day for 15 days | Male Sprague–Dawley rats | 2 g/kg, p.o. single dose | Enhancing antioxidant defense and accelerating APAP harmless metabolism, inhibiting hepatotoxicity-mediated NAPQI formation. | Sarkar, et al., 2022 [154] |
| 70% Methanolic Extract | Iris Florentina | 250 and 500 mg/kg p.o. once a day for 7 days | Male albino rats | 2 g/kg, p.o. single dose | Mechanism not explored. | Nawaz, et al., 2022 [172] |
| Aqueous Simmondsin-rich Extract | Simmondsia chinensis, Jojoba | 0.6 mg/kg, p.o. until the 19th day | Male Swiss rats | 2 g/kg, b.w | Inhibition of NF-kB pathway-based hepatocyte apoptosis and inflammatory stress. | Feki, et al., 2022 [173] |
| Simmondsin-Hydrolyzed Extract | ||||||
| 80% Methanol Extracts | Taraxacum mongolicum Hand.-Mazz, Dandelion | 250, 500, 1000 g/120 mL, by oral gavage for 7 days | Kunming mice | 350 mg/kg, i.p. single dose | Activating the Nrf-2/HO-1 pathway and inhibition of the intrinsic apoptosis pathway. | Zheng, et al., 2022 [159] |
| Root and Leaf Water Extracts | 1 and 2 mM for Primary cells | Male SD rats | 1 g/kg, p.o., once a day, five times a week for two weeks for rats | Activated the Nrf-2 pathway to reduce the level of oxidative stress. Inhibited pro-inflammatory cytokines. Attenuated the activation of the JNK pathway and reduced the expression of Bcl-2/Bax. | Wang, et al., 2022 [157] | |
| 80% Ethanol Extract | 100, 200 and 400 mg/kg orally for 7 consecutive days | Kunming mice | 350 mg/kg, i.p. single dose | Inhibited the occurrence of oxidative stress and apoptosis by suppressing MAPK and NF-κB pathways. | Ren, et al., 2020 [158] | |
| 0.5% Citric Acid in 80% Ethanol Extract | Cleistocalyx nervosum var. paniala, Ma-kiang | 100 and 300 mg/kg, p.o. for 7 days, once a day. | Female Wistar rats | 3000 mg/kg, p.o. single dose | Inducted antioxidant enzymes and detoxified enzymes leading to the restoration of GSH and a reduction in oxidative stress. | Chariyakornkul, et al., 2022 [174] |
| Acetone Extract | Drynaria quercifolia (L.) | 1, 5 mg/kg, i.g. for 21 days, once a day. | Male Swiss albino mice | 500 mg/kg, i.g. single dose | Inhibited the NF-κB-signaling pathway and activation of antioxidant response through increased expression of Nrf2. | Chatterjee, et al., 2022 [175] |
| 70% Ethanol Extract | Chrysanthemum morifolium Ramat., Chrysanthemi Flos | 110, 220 and 440 mg/kg, i.g. for eight days | Male Sprague–Dawley rats | 800 mg/kg, i.g. single dose | Inhibit the excessive oxidative stress via GSK3β–Nrf2 pathway and reduce apoptosis through PI3K–Akt pathway. | Zhou, et al., 2021 [176] |
| Absolute Methanol Extract | Pandanus odoratissimus Linn., Pandanaceae | 300, 600, 900 mg/kg, p.o. for 14 days. | Male Sprague–Dawley rats | 3 g/kg, p.o. a single dose | Mechanism not explored. | Sinaga, et al., 2021 [177] |
| Aqueous extract | Medicago denticulata | 100, 200, and 300 mg/kg, p.o. continued for 3 weeks | Rabbits | 200 mg/kg, p.o. for 3 weeks | Lipid peroxidation. | Ahmad, et al., 2021 [178] |
| Ethanol Extract | Orostachys fimbriata (Turcz.) A. Berger | 50, 100, or 200 mg/kg p.o. once a day for 7 days | Male C57BL/6 mice | 300 mg/kg, i.p. single dose | Reduced CYPs via PXR inhibition and restoration of hepatic GSH content. | Zhou, et al., 2021 [179] |
| 50% Ethanol Extract | Centella asiatica (L.) Urb., Centella Asiatica | 50, 100, and 200 mg/kg/days, p.o. for 7 days | Male BALB/c strain mice | 200 mg/kg, p.o. single dose | Antioxidant capacity and reduced expression of the inflammatory gene CYP2E1 transcripts. | Park, et al., 2021 [180] |
| Total Flavonoids | Sedum sarmentosum Bunge | 50, 100, 200 mg/kg, no indication of the number of days of injection and method of administration | Male ICR mice | No concentration indicated | Inhibited the expression of Nrf2 and ARE proteins in the liver tissue. | Jiang, et al., 2021 [181] |
| Methanol Extract | Acampe ochracea (Lindl.) Hochr. | 200, 100, and 50 mg/kg orally once daily for 14 consecutive days | Male Swiss albino rat | 2 g/kg, oral, once | The VEGF signaling pathway to recover the injured hepatocytes. | Ahmed, et al., 2021 [182] |
| Aqueous extracts. | Date palm seeds; Phoenix dactylifera L. | 200 and 400 mg/kg i.g. for six days | Male Wistar rats | 1.5 g/kg, oral, once | Potent antioxidant capacity, membrane stabilizing effect and inhibit CYP, decreased the formation of NAPQI. | Bouhlali, et al., 2021 [183] |
| 30% Ethanolic Extract | wheat, Triticum aestivum L. | 100, 200 mg/kg, p.o. for six days | Male C57BL/6 mice | 300 mg/kg, i.p. once | Mechanism not explored. | Lim, et al., 2021 [184] |
| Aqueous Extract | Dill shoots, A. graveolens L. | 100, 200 mg/kg, p.o. daily for eight consecutive days | Male Wistar rats | 2.5 g/kg, p.o. single dose, on the 9th day. | Increase in the level of GSH and TAC, and normalization of liver enzymes and inflammatory mediators. | Rasheed, et al., 2021 [185] |
| Triterpenoid-Enriched Extract | Guava Leaf, Psidium guajava | 75, 150 mg/kg, p.o. daily for 7 days | Male C57BL/6 mice | 300 mg/kg, i.p. once | The Nrf2 and MAPK signaling pathways. | Li, et al., 2021 [186] |
| Compound 1 (1.1, 3.3, 10, and 30 μM), Compounds 6, 7, 16, and 17 (10 and 30 μM) | HepG2 cells | With 10 mM for another 48 h | ||||
| Hot-water Extract | Que Zui tea, Vaccinium dunalianum Wight | 200 and 600 mg/kg, i.g. once daily for 7 consecutive days | Male Kunming mice | 400 mg/kg, i.p. once | Enhance the expression of Nrf2, NQO1, and HO-1 proteins, suppressing the activation of MAPK signaling pathway. Enhancing the Bcl-2/Bax ratio and reducing caspase-3 and caspase-9 expressions in the liver tissues. | Wang, et al., 2021 [46] |
| Aqueous-ethanol Extract | 200 and 600 mg/kg, i.g. once daily for 7 consecutive days | |||||
| Lipid-soluble (chloroform) Extract | Dicranopteris linearis (Brum.f.) Underw., Dicranopteris dichotoma | 50, 250, or 500 mg/kg, p.o. for 7 successive days | Male SD rats | 3 g/kg, p.o. single dose | Hepatoprotective activity, a high antioxidant capacity, improve the endogenous antioxidant enzymatic defense system. | Zakaria, et al., 2021 [187] Zakaria, et al., 2020 [188] |
| Licorice Extract | Dried roots, Glycyrrhiza | 30 mg/kg, p.o. for 7 days | Male SD rats | 500 mg/kg, i.p. single dose | Activated PI3K/Akt/Nrf2/HO-I, Keapl/Nrf2-ARE pathway and inhibited NF-κB/MAPK pathway. | Ma, et al., 2021 [189] |
| 85% Ethanol Extract | Schisandra, Schisandra Michx. | 10 mL/kg, i.g. one time each day for 7 days. | Male ICR mice | 200 mg/kg, i.p. single dose | Attributed to the activation of ErbB signaling pathway and Arachidonic acid metabolism pathway, and inhibiting of oxidative stress, reducing ROS mediated DNA damage and inflammation, and reducing the metabolism of toxic substances of NAPQI. | Li, et al., 2021 [190] |
| 50% Ethanol Extract | 500 mg/kg, i.g. twice a day for three consecutive days. | Male C57BL/6J mice | 400 mg/kg, i.p. single dose | Reducing intrahepatic diglyceride and triglyceride levels. | Yan, et al., 2020 [191] | |
| Steam Shoot Extract | Ginseng, Panax ginseng C.A. Meyer | 50, 100, 150, 200, and 250 mg/kg, i.g. daily for seven consecutive days | Grade ICR mice | 250 mg/kg, i.p. single dose | Oxidative stress, reduced lipid peroxidation and inflammation, inhibited apoptosis. | Yao, et al., 2020 [192] |
| 50% Ethanol Extract | Isatis indigotica Fort, Isatidis Folium | 400 mg/kg, orally, for 7 consecutive days | Male ICR mice | 100 mg/kg, i.v. once | Reducing the hepatic content of NAPQI, accelerating the generation of GSH and NAPQI-GSH adduct. | Ding, et al., 2020 [193] |
| Juice Extraction | Opuntia | 800 mg/kg, p.o., single dose | Male Wistar rats | 500 mg/kg, i.p. single dose | Scavenges ROS, increases antioxidant gene expression, and regulates the expression of important ros genes, decreases Gadd45b expression. | Gonzalez-Ponce, et al., 2020 [194] |
| After 0.5 h, 10 mg/mL | Male Wistar rat primary hepatocytes | 10, 20 mmol/L | ||||
| 80% Ethanol Extract | Vachellia nilotica (L.) P. J. H. Hurter & Mabb, Acacia | 100 mg/kg, i.g. daily for a month | Wister albino rats | 750 mg/kg, i.g. daily for a month | Mechanism not explored. | Salman, et al., 2020 [195] |
| Stems, Leaves, and Roots Extract | Leptadenia hastata, Asclepiadaceae | 250 mg/kg, orally, once daily for one week | Male and female albino mice | 250 mg/kg, single dose | Antioxidant and anti-inflammatory activities. | Galani, et al., 2020 [196] |
| Ethanol Extract | aurantii fructus immaturus, Citrus aurantium L. | 6 g/kg, i.g. daily for 7 consecutive days | Male SD rats | 2 g/kg, p.o. single dose | Regulating lipid metabolism combined with conducting multi-targeted signaling pathway. | Shu, et al., 2020 [197] |
| Aqueous Extracts of Roots, Leaves, and Barks | Bixaceae, Bixa orellana L. | 250 mg/kg, i.g. 3 h after paracetamol treatments once a day for one week | Albino mice | 250 mg/kg, i.p. single dose, | Inhibited ERK signaling pathway, anti-inflammatory and antioxidant potential. | Gatsou Djibersou, et al., 2020 [198] |
| 50% Aqueous Ethanol | Yellow Chinese Chive | 25 and 100 mg/kg, p.o. daily for 7 days | Male ICR mice | 700 mg/kg, i.p. single dose. | Activated the Nrf2 signaling pathway. | Kawakami, et al., 2020 [199] |
| Fold Boiled Water Extract | Sonneratia apetala | 100, 200, and 400 mg/kg, i.g. per day for a week | Male Kunming mice | 220 mg/kg, i.p. single dose | Scavenging activity against DPPH radicals, and has counteracted oxidative stress. Hepatic GSH levels and GSH-Px activity increased. Attenuating oxidative stress and increasing antioxidant enzyme activity. Attenuated the inflammatory response. | Liu, et al., 2019 [200] |
| 70% Methanol Extract | Myristica fragrans Houtt., nutmeg | 300 mg/kg, p.o. daily for a week | Male Wistar albino rats | 2 g/kg, p.o. single dose | Enhance the antioxidant defence system, antioxidant, anti-inflammatory, and anti-apoptotic, promote the Nrf2/ARE pathway. | Dkhil, et al., 2019 [201] |
| 70% Ethanol Extract | Nasturtium officinale | 500 mg/kg, p.o. per day for a week | Male Wistar rats | 2 g/kg, p.o. single dose | Increasing T-SH content as well as enhancing GPx activity. Antioxidant activity as a free radical scavenger. | Azarmehr, et al., 2019 [202] |
| Polyphenolic-rich Fraction Extract | Lauridia tetragona (L.f) R.H. Archer | 500, 250, 125, and 62.5 μg/mL for 24 h | HepG2 cells | 100 mM, single dose | Activated antioxidant enzyme synthesis through the Nrf2 pathway. Antioxidant activity and/or polyphenolic contents. Induced the expression of HO-1 and Nrf2 protein levels. | Odeyemi, et al., 2019 [203] |
| 95% Ethanol Extract | Veronica ciliata Fisch. | 900, 600, or 300 mg/kg, p.o. per day for two weeks | Male Kunming mice | 150 mg/kg, i.p. single dose | Showed significant reversal by the decrease in hepatic functional enzyme activity, the inhibition of lipid peroxidation, and the increase in total serum antioxidant capacity (T-AOC) and antioxidant enzyme activity. | Lu, et al., 2019 [204] |
| 10, 25, and 100 μM, treated for 2 h | BRL-3A cells | 20 mM, treated for 12 h | Involved in the p62-Keap1-Nrf2 signal pathway. | |||
| Aqueous Extract | Allium sativum L., Garlic | 25, 50, 100 mg/kg, i.g. for 7 days | Male Kunming mice | 300 mg/kg, i.p. single dose | Inhibited CYP2E1, and activated Nrf2 pathway. | Zhao, et al., 2019 [205] |
4.4. Formula Extract
| Formula Extract | Dose, Route, and Duration of Administration | Animals | APAP Dose and Route of Administration | Findings and Key Mechanisms | Reference |
|---|---|---|---|---|---|
| GTS-LE | 55.00 mg/kg Sal B, 8.60 mg/kg TSN IIA, 33.13 mg/kg GA, i.g. | Male Kunming mice | 500 mg/kg, i.g. single dose | Inhibited oxidative stress and lipid peroxidation. | Zhang, et al., 2022 [208] |
| Pien Tze Huang | 75, 150, or 300 mg/kg, 2 times/day for 3 days, p.o | WT- C57BL/6 mice | 400 mg/kg, i.p. single dose | Inhibition the NF-κB pathway, NLRP3 inflammasome, and promoted autophagy. | Zhao, et al., 2023 [207] |
| HF | 100, 200, and 400 mg/kg, p.o. for 15 days | Either sex wistar rats | 640 mg/kg, p.o. single dose | Hepatoprotective and antioxidant. | Kataki, et al., 2022 [209] |
| LWWL | Schisandrin A or Schisandrin B 10, 20, and 40 μM | HepaG2 cells | 20 mM for 12 h | Inhibited the NF-κB signaling pathway and suppressed H2O2-induced cell apoptosis, and inhibited the release of ROS. | Gao, et al., 2021 [210] |
| 159.78 mg/kg Schisandrin A, 162.43 mg/kg Schisandrin B, 0.93 mg/kg esculetin, 8.56 mg/kg luteolin | Male SD rats | 300 mg/kg, i.p. single dose | |||
| Gamisoyo-san | 100–400 μg/mL for 2 h | BNL CL.2 cell | 20 mM, single dose | Lipid peroxidation. | Jin, et al., 2021 [211] |
| 700 mg/kg, 1400 mg/kg, p.o. for 7 days | Male BALB/c mice | 200 mg/kg, i.p. single dose | |||
| A multi-herbal combination (MHC) | 50, 100, and 200 mg/kg, i.g. once daily for seven consecutive days | Female Wistar rats | 3 g/kg, i.g. single dose | Triggering antioxidant defense systems, and stabilizing cell membranes by inhibiting LPO. | Kaur, et al., 2021 [206] |
| Yan-Gan-Wan (YGW) | Pre or post:100 mg/kg, i.g. once daily for 7 or 14 days | Male ICR mice | 400 mg/kg, i.p. single dose | Reducing collagen fiber formation, mitigates oxidative stress, inflammatory factors, and apoptosis, and inhibits the expression of TNF-α and caspase-3. | Yeh, et al., 2021 [212] |
5. Discussion and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. CB 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Bjornsson, E.S. Clinical management of patients with drug-induced liver injury (DILI). United Eur. Gastroenterol. J. 2021, 9, 781–786. [Google Scholar] [CrossRef]
- McGill, M.R.; Jaeschke, H. Animal models of drug-induced liver injury. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1031–1039. [Google Scholar] [CrossRef]
- Subramanya, S.B.; Venkataraman, B.; Meeran, M.F.N.; Goyal, S.N.; Patil, C.R.; Ojha, S. Therapeutic Potential of Plants and Plant Derived Phytochemicals against Acetaminophen-Induced Liver Injury. Int. J. Mol. Sci. 2018, 19, 3776. [Google Scholar] [CrossRef]
- Yoon, E.; Babar, A.; Choudhary, M.; Kutner, M.; Pyrsopoulos, N. Acetaminophen-Induced Hepatotoxicity: A Comprehensive Update. J. Clin. Transl. Hepatol. 2016, 4, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Chiew, A.L.; Reith, D.; Pomerleau, A.; Wong, A.; Isoardi, K.Z.; Soderstrom, J.; Buckley, N.A. Updated guidelines for the management of paracetamol poisoning in Australia and New Zealand. Med. J. Aust. 2020, 212, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Zempsky, W.T.; Bhagat, P.K.; Siddiqui, K. Practical Challenges-Use of Paracetamol in Children and Youth Who Are Overweight or Obese: A Narrative Review. Paediatr. Drugs 2020, 22, 525–534. [Google Scholar] [CrossRef]
- Heard, K.; Bui, A.; Mlynarchek, S.L.; Green, J.L.; Bond, G.R.; Clark, R.F.; Kozer, E.; Koff, R.S.; Dart, R.C. Toxicity from repeated doses of acetaminophen in children: Assessment of causality and dose in reported cases. Am. J. Ther. 2014, 21, 174–183. [Google Scholar] [CrossRef]
- Rosenblatt, R.; Brown, R.S., Jr. Nonviral or Drug-Induced Etiologies of Acute Liver Failure. Clin. Liver Dis. 2018, 22, 347–360. [Google Scholar] [CrossRef]
- Maclean, D.; Peters, T.J.; Brown, R.A.; McCathie, M.; Baines, G.F.; Robertson, P.G. Treatment of acute paracetamol poisoning. Lancet 1968, 19, 849–852. [Google Scholar] [CrossRef]
- Akintonwa, A.; Essien, A.R. Protective effects of Garcinia kola seed extract against paracetamol-induced hepatotoxicity in rats. J. Ethnopharmacol. 1990, 29, 207–211. [Google Scholar] [CrossRef]
- Chilvery, S.; Yelne, A.; Khurana, A.; Saifi, M.A.; Bansod, S.; Anchi, P.; Godugu, C. Acetaminophen induced hepatotoxicity: An overview of the promising protective effects of natural products and herbal formulations. Phytomed. Int. J. Phytother. Phytopharm. 2023, 108, 154510. [Google Scholar] [CrossRef]
- Cai, X.; Cai, H.; Wang, J.; Yang, Q.; Guan, J.; Deng, J.; Chen, Z. Molecular pathogenesis of acetaminophen-induced liver injury and its treatment options. J. Zhejiang Univ. Sci. B 2022, 23, 265–285. [Google Scholar] [CrossRef]
- Chen, W.; Koenigs, L.L.; Thompson, S.J.; Peter, R.M.; Rettie, A.E.; Trager, W.F.; Nelson, S.D. Oxidation of acetaminophen to its toxic quinone imine and nontoxic catechol metabolites by baculovirus-expressed and purified human cytochromes P450 2E1 and 2A6. Chem. Res. Toxicol. 1998, 11, 295–301. [Google Scholar] [CrossRef]
- Liao, J.; Lu, Q.; Li, Z.; Li, J.; Zhao, Q.; Li, J. Acetaminophen-induced liver injury: Molecular mechanism and treatments from natural products. Front. Pharmacol. 2023, 14, 1122632. [Google Scholar] [CrossRef]
- Nakagawa, H.; Maeda, S.; Hikiba, Y.; Ohmae, T.; Shibata, W.; Yanai, A.; Sakamoto, K.; Ogura, K.; Noguchi, T.; Karin, M.; et al. Deletion of apoptosis signal-regulating kinase 1 attenuates acetaminophen-induced liver injury by inhibiting c-Jun N-terminal kinase activation. Gastroenterology 2008, 135, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Win, S.; Than, T.A.; Min, R.W.; Aghajan, M.; Kaplowitz, N. c-Jun N-terminal kinase mediates mouse liver injury through a novel Sab (SH3BP5)-dependent pathway leading to inactivation of intramitochondrial Src. Hepatology 2016, 63, 1987–2003. [Google Scholar] [CrossRef] [PubMed]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxidative Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [PubMed]
- Masubuchi, Y.; Suda, C.; Horie, T. Involvement of mitochondrial permeability transition in acetaminophen-induced liver injury in mice. J. Hepatol. 2005, 42, 110–116. [Google Scholar] [CrossRef]
- Iorga, A.; Dara, L.; Kaplowitz, N. Drug-Induced Liver Injury: Cascade of Events Leading to Cell Death, Apoptosis or Necrosis. Int. J. Mol. Sci. 2017, 18, 1018. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Nelson, L.J.; Gomez Del Moral, M.; Martinez-Naves, E.; Cubero, F.J. Dissecting the molecular pathophysiology of drug-induced liver injury. World J. Gastroenterol. 2018, 24, 1373–1385. [Google Scholar] [CrossRef]
- McGill, M.R.; Williams, C.D.; Xie, Y.; Ramachandran, A.; Jaeschke, H. Acetaminophen-induced liver injury in rats and mice: Comparison of protein adducts, mitochondrial dysfunction, and oxidative stress in the mechanism of toxicity. Toxicol. Appl. Pharmacol. 2012, 264, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Chao, X.; Wang, H.; Jaeschke, H.; Ding, W.X. Role and mechanisms of autophagy in acetaminophen-induced liver injury. Liver Int. Off. J. Int. Assoc. Study Liver 2018, 38, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Torres, M.; Quintas, G.; Castell, J.V. The Potential Role of Metabolomics in Drug-Induced Liver Injury (DILI) Assessment. Metabolites 2022, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- Watkins, P.B.; Seligman, P.J.; Pears, J.S.; Avigan, M.I.; Senior, J.R. Using controlled clinical trials to learn more about acute drug-induced liver injury. Hepatology 2008, 48, 1680–1689. [Google Scholar] [CrossRef] [PubMed]
- Larrey, D. Drug-induced liver diseases. J. Hepatol. 2000, 32, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Church, R.J.; Kullak-Ublick, G.A.; Aubrecht, J.; Bonkovsky, H.L.; Chalasani, N.; Fontana, R.J.; Goepfert, J.C.; Hackman, F.; King, N.M.P.; Kirby, S.; et al. Candidate biomarkers for the diagnosis and prognosis of drug-induced liver injury: An international collaborative effort. Hepatology 2019, 69, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Zhai, T.; Zhang, J.; Zhang, J.; Liu, B.; Zhou, Z.; Liu, F.; Wu, Y. Cathelicidin promotes liver repair after acetaminophen-induced liver injury in mice. JHEP Rep. Innov. Hepatol. 2023, 5, 100687. [Google Scholar] [CrossRef]
- Yu, Y.; Chang, L.; Hu, Q.; Zhu, J.; Zhang, J.; Xia, Q.; Zhao, J. P2rx1 deficiency alleviates acetaminophen-induced acute liver failure by regulating the STING signaling pathway. Cell Biol. Toxicol. 2023. ahead of print. [Google Scholar] [CrossRef]
- Sun, M.; Chen, P.; Xiao, K.; Zhu, X.; Zhao, Z.; Guo, C.; He, X.; Shi, T.; Zhong, Q.; Jia, Y.; et al. Circulating Cell-Free DNAs as a Biomarker and Therapeutic Target for Acetaminophen-Induced Liver Injury. Adv. Sci. 2023, 10, e2206789. [Google Scholar] [CrossRef]
- Zhu, Y.; Lei, L.; Wang, X.; Chen, L.; Li, W.; Li, J.; Zhao, C.; Du, X.; Song, Y.; Gao, W.; et al. The E3 ubiquitin ligase NEDD4-1 protects against acetaminophen-induced liver injury by targeting VDAC1 for degradation. Acta Pharm. Sin. B 2023, 13, 1616–1630. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Sun, P.; Pan, B.; Qiu, J.; Zhang, X.; Shen, S.; Ke, X.; Tang, N. IL-33/ST2 antagonizes STING signal transduction via autophagy in response to acetaminophen-mediated toxicological immunity. Cell Commun. Signal. CCS 2023, 21, 80. [Google Scholar] [CrossRef] [PubMed]
- Cen, Y.; Lou, G.; Qi, J.; Li, M.; Zheng, M.; Liu, Y. Adipose-Derived Mesenchymal Stem Cells Inhibit JNK-Mediated Mitochondrial Retrograde Pathway to Alleviate Acetaminophen-Induced Liver Injury. Antioxidants 2023, 12, 158. [Google Scholar] [CrossRef]
- Zeng, Y.; Wu, R.; Wang, F.; Li, S.; Li, L.; Li, Y.; Qin, P.; Wei, M.; Yang, J.; Wu, J.; et al. Liberation of daidzein by gut microbial β-galactosidase suppresses acetaminophen-induced hepatotoxicity in mice. Cell Host Microbe 2023, 31, 766–780. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Phan, M.; Zhu, J.; Ma, X.; Manautou, J.E.; Zhong, X.B. Alterations of Cytochrome P450-Mediated Drug Metabolism during Liver Repair and Regeneration after Acetaminophen-Induced Liver Injury in Mice. Drug Metab. Dispos. Biol. Fate Chem. 2022, 50, 694–703. [Google Scholar] [CrossRef]
- He, X.; Kang, K.; Pan, D.; Sun, Y.; Chang, B. FTY720 attenuates APAP-induced liver injury via the JAK2/STAT3 signaling pathway. Int. J. Mol. Med. 2022, 49, 67. [Google Scholar] [CrossRef]
- Hou, X.; Liu, Q.; Gao, Y.; Yong, L.; Xie, H.; Li, W.; Zhou, Y.; Liu, J.; Feng, L.; Xu, L.; et al. Mesencephalic astrocyte-derived neurotrophic factor reprograms macrophages to ameliorate acetaminophen-induced acute liver injury via p38 MAPK pathway. Cell Death Dis. 2022, 13, 100. [Google Scholar] [CrossRef]
- Lei, X.; Xu, Q.; Li, C.; Niu, B.; Ming, Y.; Li, J.; Tang, Y.; Li, X.; Tang, J.; Wu, J.; et al. Egr1 confers protection against acetaminophen-induced hepatotoxicity via transcriptional upregulating of Acaa2. Int. J. Biol. Sci. 2022, 18, 3800–3817. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, L.; Pan, B.; Chen, Y.; Zhang, T.; Tang, N. Interleukin 33 mediates hepatocyte autophagy and innate immune response in the early phase of acetaminophen-induced acute liver injury. Toxicology 2021, 456, 152788. [Google Scholar] [CrossRef]
- Qian, H.; Bai, Q.; Yang, X.; Akakpo, J.Y.; Ji, L.; Yang, L.; Rulicke, T.; Zatloukal, K.; Jaeschke, H.; Ni, H.M.; et al. Dual roles of p62/SQSTM1 in the injury and recovery phases of acetaminophen-induced liver injury in mice. Acta Pharm. Sin. B 2021, 11, 3791–3805. [Google Scholar] [CrossRef]
- Shan, Z.; Li, L.K.; Atkins, C.L.; Wang, M.; Wen, Y.K.; Jeong, J.; Moreno, N.F.; Feng, D.C.; Gui, X.; Zhang, N.Y.; et al. Chitinase 3-like-1 contributes to acetaminophen-induced liver injury by promoting hepatic platelet recruitment. eLife 2021, 10, e68571. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, Y.; Wang, Z.; Sun, R.; Zou, B.; Li, R.; Liu, D.; Lin, M.; Zhou, J.; Ning, S.; et al. Peroxiredoxin 3 Inhibits Acetaminophen-Induced Liver Pyroptosis through the Regulation of Mitochondrial ROS. Front. Immunol. 2021, 12, 652782. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, E.; Gudd, C.L.; Mawhin, M.A.; Husbyn, H.C.; Trovato, F.M.; Siggins, M.K.; O’Connor, T.; Kudo, H.; Mukherjee, S.K.; Wendon, J.A.; et al. PD-1 blockade improves Kupffer cell bacterial clearance in acute liver injury. J. Clin. Investig. 2021, 131, e140196. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, X.; Jiao, T.; Li, W.; Chen, P.; Jiang, Y.; Sun, J.; Chen, Y.; Chen, P.; Guan, L.; et al. SIRT6 as a key event linking P53 and NRF2 counteracts APAP-induced hepatotoxicity through inhibiting oxidative stress and promoting hepatocyte proliferation. Acta Pharm. Sin. B 2021, 11, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, T.; Tong, Y.; Cui, R.; Qu, K.; Liu, C.; Zhang, J. Ulinastatin protects against acetaminophen-induced liver injury by alleviating ferroptosis via the SIRT1/NRF2/HO-1 pathway. Am. J. Transl. Res. 2021, 13, 6031–6042. [Google Scholar]
- Wang, Y.; Tian, L.; Wang, Y.; Zhao, T.; Khan, A.; Wang, Y.; Cao, J.; Cheng, G. Protective effect of Que Zui tea hot-water and aqueous ethanol extract against acetaminophen-induced liver injury in mice via inhibition of oxidative stress, inflammation, and apoptosis. Food Funct. 2021, 12, 2468–2480. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Jung, Y.S.; Yun, J.; Han, S.B.; Roh, Y.S.; Song, M.J.; Hong, J.T. Peroxiredoxin 6 mediates acetaminophen-induced hepatocyte death through JNK activation. Redox Biol. 2020, 32, 101496. [Google Scholar] [CrossRef]
- Li, L.; Wang, H.; Zhang, J.; Sha, Y.; Wu, F.; Wen, S.; He, L.; Sheng, L.; You, Q.; Shi, M.; et al. SPHK1 deficiency protects mice from acetaminophen-induced ER stress and mitochondrial permeability transition. Cell Death Differ. 2020, 27, 1924–1937. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, K.; Zhang, J.; Hai, Y.; Wang, P.; Wang, H.; Liu, Q.; Wong, C.C.L.; Yao, J.; Gao, Y.; et al. c-Jun NH(2)-Terminal Protein Kinase Phosphorylates the Nrf2-ECH Homology 6 Domain of Nuclear Factor Erythroid 2-Related Factor 2 and Downregulates Cytoprotective Genes in Acetaminophen-Induced Liver Injury in Mice. Hepatology 2020, 71, 1787–1801. [Google Scholar] [CrossRef]
- Gao, R.Y.; Wang, M.; Liu, Q.; Feng, D.; Wen, Y.; Xia, Y.; Colgan, S.P.; Eltzschig, H.K.; Ju, C. Hypoxia-Inducible Factor-2α Reprograms Liver Macrophages to Protect Against Acute Liver Injury through the Production of Interleukin-6. Hepatology 2020, 71, 2105–2117. [Google Scholar] [CrossRef]
- Li, M.; Sun, X.; Zhao, J.; Xia, L.; Li, J.; Xu, M.; Wang, B.; Guo, H.; Yu, C.; Gao, Y.; et al. CCL5 deficiency promotes liver repair by improving inflammation resolution and liver regeneration through M2 macrophage polarization. Cell. Mol. Immunol. 2020, 17, 753–764. [Google Scholar] [CrossRef]
- Wen, Y.; Wang, C.; Gu, J.; Yu, C.; Wang, K.; Sun, X.; Sun, Y.; Wu, H.; Tong, Y.; Xia, Q.; et al. Metabolic modulation of acetaminophen-induced hepatotoxicity by osteopontin. Cell. Mol. Immunol. 2019, 16, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ni, H.M.; Chao, X.; Ma, X.; Rodriguez, Y.A.; Chavan, H.; Wang, S.; Krishnamurthy, P.; Dobrowsky, R.; Xu, D.X.; et al. Double deletion of PINK1 and Parkin impairs hepatic mitophagy and exacerbates acetaminophen-induced liver injury in mice. Redox Biol. 2019, 22, 101148. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Ni, H.M.; Wang, L.; Ma, X.; Yu, J.; Ding, W.X.; Zhang, L. p53 Up-regulated Modulator of Apoptosis Induction Mediates Acetaminophen-Induced Necrosis and Liver Injury in Mice. Hepatology 2019, 69, 2164–2179. [Google Scholar] [CrossRef] [PubMed]
- Seglen, P.O. Preparation of isolated rat liver cells. Methods Cell Biol. 1976, 13, 29–83. [Google Scholar] [CrossRef] [PubMed]
- Godoy, P.; Hewitt, N.J.; Albrecht, U.; Andersen, M.E.; Ansari, N.; Bhattacharya, S.; Bode, J.G.; Bolleyn, J.; Borner, C.; Bottger, J.; et al. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch. Toxicol. 2013, 87, 1315–1530. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, A.; Liu, M.; Ren, X. Comparative Protective Effect of Lipoic Acid and Resveratrol on Acetaminophen-Induced Cellular Oxidative Damage in HepG2 Cells. Food Sci. 2020, 41, 153–158. [Google Scholar]
- Schmitt, B.; Vicenzi, M.; Garrel, C.; Denis, F.M. Effects of N-acetylcysteine, oral glutathione (GSH) and a novel sublingual form of GSH on oxidative stress markers: A comparative crossover study. Redox Biol. 2015, 6, 198–205. [Google Scholar] [CrossRef]
- Tang, F.; Wang, Z.; Zhou, J.; Yao, J. Salvianolic Acid A Protects against Acetaminophen-Induced Hepatotoxicity via Regulation of the miR-485-3p/SIRT1 Pathway. Antioxidants 2023, 12, 870. [Google Scholar] [CrossRef]
- Lin, M.; Zhai, X.; Wang, G.; Tian, X.; Gao, D.; Shi, L.; Wu, H.; Fan, Q.; Peng, J.; Liu, K.; et al. Salvianolic acid B protects against acetaminophen hepatotoxicity by inducing Nrf2 and phase II detoxification gene expression via activation of the PI3K and PKC signaling pathways. J. Pharmacol. Sci. 2015, 127, 203–210. [Google Scholar] [CrossRef]
- Wu, C.T.; Deng, J.S.; Huang, W.C.; Shieh, P.C.; Chung, M.I.; Huang, G.J. Salvianolic Acid C against Acetaminophen-Induced Acute Liver Injury by Attenuating Inflammation, Oxidative Stress, and Apoptosis through Inhibition of the Keap1/Nrf2/HO-1 Signaling. Oxidative Med. Cell. Longev. 2019, 2019, 9056845. [Google Scholar] [CrossRef]
- Wang, W.; Guan, C.; Sun, X.; Zhao, Z.; Li, J.; Fu, X.; Qiu, Y.; Huang, M.; Jin, J.; Huang, Z. Tanshinone IIA protects against acetaminophen-induced hepatotoxicity via activating the Nrf2 pathway. Phytomed. Int. J. Phytother. Phytopharm. 2016, 23, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Sharma, A.; Dogra, A.; Sharma, P.; Kumar, A.; Kotwal, P.; Bag, S.; Misra, P.; Singh, G.; Kumar, A.; et al. Glabridin attenuates paracetamol-induced liver injury in mice via CYP2E1-mediated inhibition of oxidative stress. Drug Chem. Toxicol. 2022, 45, 2352–2360. [Google Scholar] [CrossRef]
- Yang, W.; Liang, Z.; Wen, C.; Jiang, X.; Wang, L. Silymarin Protects against Acute Liver Injury Induced by Acetaminophen by Downregulating the Expression and Activity of the CYP2E1 Enzyme. Molecules 2022, 27, 8855. [Google Scholar] [CrossRef]
- Papackova, Z.; Heczkova, M.; Dankova, H.; Sticova, E.; Lodererova, A.; Bartonova, L.; Poruba, M.; Cahova, M. Silymarin prevents acetaminophen-induced hepatotoxicity in mice. PLoS ONE 2018, 13, e0191353. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zhang, S.; Sun, Z.; Tong, Z.; Ge, Y.; Zhou, L.; Xu, Q.; Zhou, H.; Wang, W. Preclinical validation of silibinin/albumin nanoparticles as an applicable system against acute liver injury. Acta Biomater. 2022, 146, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, T.; Gao, Z.; Wu, W.; Cao, Y.; Wang, L.; Zhang, Q. Study on the Protective Effect of Schizandrin B against Acetaminophen-Induced Cytotoxicity in Human Hepatocyte. Biol. Pharm. Bull. 2022, 45, 596–604. [Google Scholar] [CrossRef]
- Dai, W.Z.; Bai, Z.F.; He, T.T.; Zhan, X.Y.; Li, Q.; Zhao, J.; Xiao, X.H. Schisandrin C improves acetaminophen-induced liver injury in mice by regulating Nrf2 signaling pathway. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2022, 47, 5299–5305. [Google Scholar] [CrossRef]
- Li, Y.Z.; Ma, Z.N.; Sun, Y.S.; Ren, S.; Jiang, S.; Zhang, W.Z.; Wang, Z.; Li, W. Protective effects of extracts of Schisandra chinensis stems against acetaminophen-induced hepatotoxicity via regulation of MAPK and caspase-3 signaling pathways. Chin. J. Nat. Med. 2018, 16, 700–713. [Google Scholar] [CrossRef]
- Qiu, B.X.; Liu, K.; Zou, L.; Zhu, H.F.; Feng, S. Effects of Schisandrae Chinensis Fructus induced CYPs and Nrf2 activation on acute liver injury induced by acetaminophen. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Med. 2018, 43, 4908–4915. [Google Scholar] [CrossRef]
- Leung, K.W.; Wong, A.S. Pharmacology of ginsenosides: A literature review. Chin. Med. 2010, 5, 20. [Google Scholar] [CrossRef]
- Li, W.; Zhang, M.; Gu, J.; Meng, Z.J.; Zhao, L.C.; Zheng, Y.N.; Chen, L.; Yang, G.L. Hypoglycemic effect of protopanaxadiol-type ginsenosides and compound K on Type 2 diabetes mice induced by high-fat diet combining with streptozotocin via suppression of hepatic gluconeogenesis. Fitoterapia 2012, 83, 192–198. [Google Scholar] [CrossRef]
- Ning, C.; Gao, X.; Wang, C.; Kong, Y.; Liu, Z.; Sun, H.; Sun, P.; Huo, X.; Ma, X.; Meng, Q.; et al. Ginsenoside Rg1 protects against acetaminophen-induced liver injury via activating Nrf2 signaling pathway in vivo and in vitro. Regul. Toxicol. Pharmacol. RTP 2018, 98, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.N.; Xu, X.Y.; Li, W.; Wang, Y.M.; Liu, Y.; Wang, Z.; Wang, Y.P. Ginsenoside Rk1 ameliorates paracetamol-induced hepatotoxicity in mice through inhibition of inflammation, oxidative stress, nitrative stress and apoptosis. J. Ginseng Res. 2019, 43, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; Hu, J.N.; Liu, Z.; Zhang, R.; He, Y.F.; Hou, W.; Wang, Z.Q.; Yang, G.; Li, W. Saponins (Ginsenosides) from the Leaves of Panax quinquefolius Ameliorated Acetaminophen-Induced Hepatotoxicity in Mice. J. Agric. Food Chem. 2017, 65, 3684–3692. [Google Scholar] [CrossRef]
- Gao, Y.; Yan, J.; Li, J.; Li, X.; Yang, S.; Chen, N.; Li, L.; Zhang, L. Ginsenoside Rg3 ameliorates acetaminophen-induced hepatotoxicity by suppressing inflammation and oxidative stress. J. Pharm. Pharmacol. 2021, 73, 322–331. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, W.; Cheng, N.; Zhu, Y.; Li, H.; Zhang, S.; Guo, W.; Ge, G. Pectolinarigenin ameliorates acetaminophen-induced acute liver injury via attenuating oxidative stress and inflammatory response in Nrf2 and PPARa dependent manners. Phytomed. Int. J. Phytother. Phytopharm. 2023, 113, 154726. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhou, S.; Wang, Y.; Di, S.; Wang, Y.; Huang, X.; Chen, Y. Leonurine alleviates acetaminophen-induced acute liver injury by regulating the PI3K/AKT signaling pathway in mice. Int. Immunopharmacol. 2023, 120, 110375. [Google Scholar] [CrossRef]
- Liu, F.C.; Lee, H.C.; Liao, C.C.; Chou, A.H.; Yu, H.P. Role of NADPH Oxidase-Derived ROS-Mediated IL-6/STAT3 and MAPK/NF-κB Signaling Pathways in Protective Effect of Corilagin against Acetaminophen-Induced Liver Injury in Mice. Biology 2023, 12, 334. [Google Scholar] [CrossRef]
- Lv, H.; Hong, L.; Tian, Y.; Yin, C.; Zhu, C.; Feng, H. Corilagin alleviates acetaminophen-induced hepatotoxicity via enhancing the AMPK/GSK3β-Nrf2 signaling pathway. Cell Commun. Signal. CCS 2019, 17, 2. [Google Scholar] [CrossRef]
- Zhao, J.; Shi, S.; Zhang, X.; Liu, Y.; Yuan, M.; Cheng, G.; Wang, Y. Confusoside, a dihydrochalcone glucoside, prevents acetaminophen-induced liver injury by modulating the Nrf2/NF-κB/caspase signaling pathway. Food Funct. 2023, 14, 2432–2443. [Google Scholar] [CrossRef]
- Yao, Y.; Li, R.; Liu, D.; Long, L.; He, N. Rosmarinic acid alleviates acetaminophen-induced hepatotoxicity by targeting Nrf2 and NEK7-NLRP3 signaling pathway. Ecotoxicol. Environ. Saf. 2022, 241, 113773. [Google Scholar] [CrossRef]
- Yan, M.; Jin, S.; Liu, Y.; Wang, L.; Wang, Z.; Xia, T.; Chang, Q. Cajaninstilbene Acid Ameliorates Acetaminophen-Induced Liver Injury through Enhancing Sestrin2/AMPK-Mediated Mitochondrial Quality Control. Front. Pharmacol. 2022, 13, 824138. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Yang, X.; Han, Y.; Li, J.; Hu, C.; Liu, C.; Xiao, W. Sarmentosin promotes USP17 and regulates Nrf2-mediated mitophagy and cellular oxidative stress to alleviate APAP-induced acute liver failure. Phytomed. Int. J. Phytother. Phytopharm. 2022, 104, 154337. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ge, J.; Li, M.; Deng, S.; Li, J.; Ma, Y.; Zhang, J.; Zheng, Y.; Ma, L. Network pharmacology, molecular docking technology integrated with pharmacodynamic study to reveal the potential targets of Schisandrol A in drug-induced liver injury by acetaminophen. Bioorg. Chem. 2022, 118, 105476. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.C.; Yu, H.P.; Chou, A.H.; Lee, H.C.; Hu, L.M.; Liu, F.C. Gastrodin Alleviates Acetaminophen-Induced Liver Injury in a Mouse Model Through Inhibiting MAPK and Enhancing Nrf2 Pathways. Inflammation 2022, 45, 1450–1462. [Google Scholar] [CrossRef]
- Zhang, C.; Lin, J.; Zhen, C.; Wang, F.; Sun, X.; Kong, X.; Gao, Y. Amygdalin protects against acetaminophen-induced acute liver failure by reducing inflammatory response and inhibiting hepatocyte death. Biochem. Biophys. Res. Commun. 2022, 602, 105–112. [Google Scholar] [CrossRef]
- Deng, X.; Li, Y.; Li, X.; Zhang, Z.; Dai, S.; Wu, H.; Zhang, F.; Hu, Q.; Chen, Y.; Zeng, J.; et al. Paeoniflorin Protects against Acetaminophen-Induced Liver Injury in Mice via JNK Signaling Pathway. Molecules 2022, 27, 8534. [Google Scholar] [CrossRef]
- Mohamed Kamel, G.A.; Harahsheh, E.; Hussein, S. Diacerein ameliorates acetaminophen hepatotoxicity in rats via inhibiting HMGB1/TLR4/NF-κB and upregulating PPAR-γ signal. Mol. Biol. Rep. 2022, 49, 5863–5874. [Google Scholar] [CrossRef]
- Shen, P.; Han, L.; Chen, G.; Cheng, Z.; Liu, Q. Emodin Attenuates Acetaminophen-Induced Hepatotoxicity via the cGAS-STING Pathway. Inflammation 2022, 45, 74–87. [Google Scholar] [CrossRef]
- Hong, M.K.; Liu, H.H.; Chen, G.H.; Zhu, J.Q.; Zheng, S.Y.; Zhao, D.; Diao, J.; Jia, H.; Zhang, D.D.; Chen, S.X.; et al. Oridonin Alters Hepatic Urea Cycle via Gut Microbiota and Protects against Acetaminophen-Induced Liver Injury. Oxidative Med. Cell. Longev. 2021, 2021, 3259238. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jin, S.W.; Lee, G.H.; Han, E.H.; Hwang, Y.P.; Jeong, H.G. Rutaecarpine Protects against Acetaminophen-Induced Acute Liver Injury in Mice by Activating Antioxidant Enzymes. Antioxidants 2021, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Li, D.; Ma, H.; Mu, Y.; Huang, X.; Li, L. Guavinoside B from Psidium guajava alleviates acetaminophen-induced liver injury via regulating the Nrf2 and JNK signaling pathways. Food Funct. 2020, 11, 8297–8308. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.O.; Kim, J.H.; Che, D.N.; Kang, H.J.; Shin, J.Y.; Hao, S.; Park, J.H.; Wang, F.; Lee, Y.J.; Jang, S.I. Kushenol C Prevents Tert-Butyl Hydroperoxide and Acetaminophen-Induced Liver Injury. Molecules 2021, 26, 1635. [Google Scholar] [CrossRef]
- BinMowyna, M.N.; AlFaris, N.A. Kaempferol suppresses acetaminophen-induced liver damage by upregulation/activation of SIRT1. Pharm. Biol. 2021, 59, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chu, S.F.; Zhang, Z.; Ai, Q.D.; Xia, C.Y.; Huang, H.Y.; Chen, N.H. Ginsenoside Rg1 prevents acetaminophen-induced oxidative stress and apoptosis via Nrf2/ARE signaling pathway. J. Asian Nat. Prod. Res. 2019, 21, 782–797. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Song, C.; Chen, J.; Zhou, L.; Jiang, X.; Cao, X.; Sun, Y.; Zhang, Q. Limonin ameliorates acetaminophen-induced hepatotoxicity by activating Nrf2 antioxidative pathway and inhibiting NF-κB inflammatory response via upregulating Sirt1. Phytomedicine Int. J. Phytother. Phytopharm. 2020, 69, 153211. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, J.; Hu, C.; Wang, T.; Lu, J.; Wu, C.; Chen, L.; Jin, M.; Ji, G.; Cao, Q.; et al. Apigenin Prevents Acetaminophen-Induced Liver Injury by Activating the SIRT1 Pathway. Front. Pharmacol. 2020, 11, 514. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Y.; Jiao, F.Z.; Yang, F.; Li, X.; Wang, L.W. Sinomenine Attenuates Acetaminophen-Induced Acute Liver Injury by Decreasing Oxidative Stress and Inflammatory Response via Regulating TGF-β/Smad Pathway in vitro and in vivo. Drug Des. Dev. Ther. 2020, 14, 2393–2403. [Google Scholar] [CrossRef]
- Dong, S.; Ji, J.; Hu, L.; Wang, H. Dihydromyricetin alleviates acetaminophen-induced liver injury via the regulation of transformation, lipid homeostasis, cell death and regeneration. Life Sci. 2019, 227, 20–29. [Google Scholar] [CrossRef]
- Shu, G.; Qiu, Y.; Hao, J.; Fu, Q.; Deng, X. γ-Oryzanol alleviates acetaminophen-induced liver injury: Roles of modulating AMPK/GSK3β/Nrf2 and NF-κB signaling pathways. Food Funct. 2019, 10, 6858–6872. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, J.; Pan, L.; Chen, L.; Wang, Y.; Liu, X.; You, L.; Jia, Y.; Hu, C. Protective effect of 7,3′,4′-flavon-3-ol (fisetin) on acetaminophen-induced hepatotoxicity in vitro and in vivo. Phytomed. Int. J. Phytother. Phytopharm. 2019, 58, 152865. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.W.; Bai, T.; Yang, Y.; Zhang, Y.; Jiang, M.; Yang, H.X.; Wu, M.; Liu, J.; Qiao, C.Y.; Zhan, Z.Y.; et al. Thymoquinone Attenuates Acetaminophen Overdose-Induced Acute Liver Injury and Inflammation Via Regulation of JNK and AMPK Signaling Pathway. Am. J. Chin. Med. 2019, 47, 577–594. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Kuang, G.; Jiang, R.; Wu, S.; Zeng, T.; Wang, Y.; Xu, F.; Xiong, L.; Gong, X.; Wan, J. Geniposide protected hepatocytes from acetaminophen hepatotoxicity by down-regulating CYP 2E1 expression and inhibiting TLR 4/NF-κB signaling pathway. Int. Immunopharmacol. 2019, 74, 105625. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.D.; Chen, J.F.; Liu, J.J.; Xie, J.H.; Zhang, Z.B.; Gu, J.Y.; Zhuo, J.Y.; Huang, S.; Su, Z.R.; Sun, Z.H. Tetrahydrocurcumin and octahydrocurcumin, the primary and final hydrogenated metabolites of curcumin, possess superior hepatic-protective effect against acetaminophen-induced liver injury: Role of CYP2E1 and Keap1-Nrf2 pathway. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 123, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.Y.; Shin, J.K.; Lee, S.M. Pterostilbene protects against acetaminophen-induced liver injury by restoring impaired autophagic flux. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2019, 123, 536–545. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Zhang, J.; Chen, X.; Li, Z.; Liu, B.; Zhang, L. Shikonin Attenuates Acetaminophen-Induced Hepatotoxicity by Upregulation of Nrf2 through Akt/GSK3β Signaling. Molecules 2018, 24, 110. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Sun, J.; Li, D.; Hu, Y.; Yu, X.; Hua, H.; Jing, X.; Chen, F.; Jia, Z.; Xu, J. Shikonin attenuates acetaminophen-induced acute liver injury via inhibition of oxidative stress and inflammation. Biomed. Pharmacother. 2019, 112, 108704. [Google Scholar] [CrossRef]
- Yan, H.; Huang, Z.; Bai, Q.; Sheng, Y.; Hao, Z.; Wang, Z.; Ji, L. Natural product andrographolide alleviated APAP-induced liver fibrosis by activating Nrf2 antioxidant pathway. Toxicology 2018, 396-397, 1–12. [Google Scholar] [CrossRef]
- Zhou, H.C.; Wang, H.; Shi, K.; Li, J.M.; Zong, Y.; Du, R. Hepatoprotective Effect of Baicalein Against Acetaminophen-Induced Acute Liver Injury in Mice. Molecules 2018, 24, 131. [Google Scholar] [CrossRef]
- Wang, Z.; Hao, W.; Hu, J.; Mi, X.; Han, Y.; Ren, S.; Jiang, S.; Wang, Y.; Li, X.; Li, W. Maltol Improves APAP-Induced Hepatotoxicity by Inhibiting Oxidative Stress and Inflammation Response via NF-κB and PI3K/Akt Signal Pathways. Antioxidants 2019, 8, 395. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Park, J.S.; Lee, Y.S.; Han, J.; Lee, D.K.; Kwon, S.W.; Han, D.H.; Lee, Y.H.; Bae, S.H. SQSTM1/p62 activates NFE2L2/NRF2 via ULK1-mediated autophagic KEAP1 degradation and protects mouse liver from lipotoxicity. Autophagy 2020, 16, 1949–1973. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Huo, Y.; Yin, S.; Hu, H. Mechanisms of acetaminophen-induced liver injury and its implications for therapeutic interventions. Redox Biol. 2018, 17, 274–283. [Google Scholar] [CrossRef]
- Goldring, C.E.; Kitteringham, N.R.; Elsby, R.; Randle, L.E.; Clement, Y.N.; Williams, D.P.; McMahon, M.; Hayes, J.D.; Itoh, K.; Yamamoto, M.; et al. Activation of hepatic Nrf2 in vivo by acetaminophen in CD-1 mice. Hepatology 2004, 39, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Wang, Y.; Li, H.; Jia, W.; Man, K.; Lo, C.M.; Wang, Y.; Lam, K.S.; Xu, A. Fibroblast growth factor 21 protects against acetaminophen-induced hepatotoxicity by potentiating peroxisome proliferator-activated receptor coactivator protein-1α-mediated antioxidant capacity in mice. Hepatology 2014, 60, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Donatus, I.A.; Sardjoko; Vermeulen, N.P. Cytotoxic and cytoprotective activities of curcumin. Effects on paracetamol-induced cytotoxicity, lipid peroxidation and glutathione depletion in rat hepatocytes. Biochem. Pharmacol. 1990, 39, 1869–1875. [Google Scholar] [CrossRef]
- Sasaki, T.; Sato, Y.; Kumagai, T.; Yoshinari, K.; Nagata, K. Effect of health foods on cytochrome P450-mediated drug metabolism. J. Pharm. Health Care Sci. 2017, 3, 14. [Google Scholar] [CrossRef]
- Lee, J.S.; Surh, Y.J. Nrf2 as a novel molecular target for chemoprevention. Cancer Lett. 2005, 224, 171–184. [Google Scholar] [CrossRef]
- Li, G.; Chen, J.B.; Wang, C.; Xu, Z.; Nie, H.; Qin, X.Y.; Chen, X.M.; Gong, Q. Curcumin protects against acetaminophen-induced apoptosis in hepatic injury. World J. Gastroenterol. 2013, 19, 7440–7446. [Google Scholar] [CrossRef]
- Bulku, E.; Stohs, S.J.; Cicero, L.; Brooks, T.; Halley, H.; Ray, S.D. Curcumin exposure modulates multiple pro-apoptotic and anti-apoptotic signaling pathways to antagonize acetaminophen-induced toxicity. Curr. Neurovascular Res. 2012, 9, 58–71. [Google Scholar] [CrossRef]
- Mu, W.; Wang, Q.; Jia, M.; Dong, S.; Li, S.; Yang, J.; Liu, G. Hepatoprotective Effects of Albumin-Encapsulated Nanoparticles of a Curcumin Derivative COP-22 against Lipopolysaccharide/D-Galactosamine-Induced Acute Liver Injury in Mice. Int. J. Mol. Sci. 2022, 23, 4903. [Google Scholar] [CrossRef] [PubMed]
- Kheradpezhouh, E.; Panjehshahin, M.R.; Miri, R.; Javidnia, K.; Noorafshan, A.; Monabati, A.; Dehpour, A.R. Curcumin protects rats against acetaminophen-induced hepatorenal damages and shows synergistic activity with N-acetyl cysteine. Eur. J. Pharmacol. 2010, 628, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.L.; Guo, Y.N.; Lu, M.H.; Ding, K.N.; Liang, S.S.; Mou, R.W.; Yuan, S.; He, Y.M.; Tang, L.P. Acetaminophen-induced hepatotoxicity predominantly via inhibiting Nrf2 antioxidative pathway and activating TLR4-NF-κB-MAPK inflammatory response in mice. Ecotoxicol. Environ. Saf. 2023, 252, 114590. [Google Scholar] [CrossRef] [PubMed]
- Leng, J.; Wang, Z.; Fu, C.L.; Zhang, J.; Ren, S.; Hu, J.N.; Jiang, S.; Wang, Y.P.; Chen, C.; Li, W. NF-κB and AMPK/PI3K/Akt signaling pathways are involved in the protective effects of Platycodon grandiflorum saponins against acetaminophen-induced acute hepatotoxicity in mice. Phytother. Res. PTR 2018, 32, 2235–2246. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, R.H.; Cook, J.G. Stress Relief Techniques: p38 MAPK Determines the Balance of Cell Cycle and Apoptosis Pathways. Biomolecules 2021, 11, 1444. [Google Scholar] [CrossRef]
- Qin, S.; Lv, C.; Wang, Q.; Zheng, Z.; Sun, X.; Tang, M.; Deng, F. Extraction, identification, and antioxidant property evaluation of limonin from pummelo seeds. Anim. Nutr. 2018, 4, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yu, H.; Chen, J.; Zhu, J.; Song, C.; Zhou, L.; Sun, Y.; Zhang, Q. Limonin Attenuates LPS-Induced Hepatotoxicity by Inhibiting Pyroptosis via NLRP3/Gasdermin D Signaling Pathway. J. Agric. Food Chem. 2021, 69, 982–991. [Google Scholar] [CrossRef]
- Mahmoud, M.F.; Gamal, S.; El-Fayoumi, H.M. Limonin attenuates hepatocellular injury following liver ischemia and reperfusion in rats via toll-like receptor dependent pathway. Eur. J. Pharmacol. 2014, 740, 676–682. [Google Scholar] [CrossRef]
- Li, Y.C.; Qiao, J.Y.; Wang, B.Y.; Bai, M.; Shen, J.D.; Cheng, Y.X. Paeoniflorin Ameliorates Fructose-Induced Insulin Resistance and Hepatic Steatosis by Activating LKB1/AMPK and AKT Pathways. Nutrients 2018, 10, 1024. [Google Scholar] [CrossRef]
- Chen, L.; Wei, S.; Liu, H.; Li, J.; Jing, M.; Tong, Y.; Li, R.; Wen, J.; Zhan, H.; Zhao, Y. Paeoniflorin Protects against ANIT-Induced Cholestatic Liver Injury in Rats via the Activation of SIRT1-FXR Signaling Pathway. Evid.-Based Complement. Altern. Med. eCAM 2021, 2021, 8479868. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. MMBR 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal Transduct. Res. 2015, 35, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.L.; Lee, J.H.; Shu, L.; Kim, J.H.; Sacks, D.B.; Kong, A.N. The Ras GTPase-activating-like protein IQGAP1 mediates Nrf2 protein activation via the mitogen-activated protein kinase/extracellular signal-regulated kinase (ERK) kinase (MEK)-ERK pathway. J. Biol. Chem. 2013, 288, 22378–22386. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Lei, Z.; Huang, A.; Wu, Q.; Xie, S.; Awais, I.; Dai, M.; Wang, X.; Yuan, Z. Toxic metabolites, MAPK and Nrf2/Keap1 signaling pathways involved in oxidative toxicity in mice liver after chronic exposure to Mequindox. Sci. Rep. 2017, 7, 41854. [Google Scholar] [CrossRef]
- Zimta, A.A.; Cenariu, D.; Irimie, A.; Magdo, L.; Nabavi, S.M.; Atanasov, A.G.; Berindan-Neagoe, I. The Role of Nrf2 Activity in Cancer Development and Progression. Cancers 2019, 11, 1755. [Google Scholar] [CrossRef]
- Gadacha, W.; Ben-Attia, M.; Bonnefont-Rousselot, D.; Aouani, E.; Ghanem-Boughanmi, N.; Touitou, Y. Resveratrol opposite effects on rat tissue lipoperoxidation: Pro-oxidant during day-time and antioxidant at night. Redox Rep. Commun. Free Radic. Res. 2009, 14, 154–158. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, Y.; Fan, X.; Tan, H.; Zeng, H.; Wang, Y.; Chen, P.; Huang, M.; Bi, H. Hepato-protective effect of resveratrol against acetaminophen-induced liver injury is associated with inhibition of CYP-mediated bioactivation and regulation of SIRT1-p53 signaling pathways. Toxicol. Lett. 2015, 236, 82–89. [Google Scholar] [CrossRef]
- Kim, D.H.; Jung, Y.J.; Lee, J.E.; Lee, A.S.; Kang, K.P.; Lee, S.; Park, S.K.; Han, M.K.; Lee, S.Y.; Ramkumar, K.M.; et al. SIRT1 activation by resveratrol ameliorates cisplatin-induced renal injury through deacetylation of p53. Am. J. Physiol. Ren. Physiol. 2011, 301, F427–F435. [Google Scholar] [CrossRef]
- Du, K.; McGill, M.R.; Xie, Y.; Bajt, M.L.; Jaeschke, H. Resveratrol prevents protein nitration and release of endonucleases from mitochondria during acetaminophen hepatotoxicity. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2015, 81, 62–70. [Google Scholar] [CrossRef]
- Wang, X.; Sun, R.; Chen, Y.; Lian, Z.X.; Wei, H.; Tian, Z. Regulatory T cells ameliorate acetaminophen-induced immune-mediated liver injury. Int. Immunopharmacol. 2015, 25, 293–301. [Google Scholar] [CrossRef]
- Masubuchi, Y.; Sugiyama, S.; Horie, T. Th1/Th2 cytokine balance as a determinant of acetaminophen-induced liver injury. Chem. -Biol. Interact. 2009, 179, 273–279. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, J.; Zhao, C.N.; Gan, R.Y.; Li, H.B. Health Benefits and Molecular Mechanisms of Resveratrol: A Narrative Review. Foods 2020, 9, 340. [Google Scholar] [CrossRef]
- Qiao, H.; Chen, L.; Rui, T.; Wang, J.; Chen, T.; Fu, T.; Li, J.; Di, L. Fabrication and in vitro/in vivo evaluation of amorphous andrographolide nanosuspensions stabilized by D-α-tocopheryl polyethylene glycol 1000 succinate/sodium lauryl sulfate. Int. J. Nanomed. 2017, 12, 1033–1046. [Google Scholar] [CrossRef]
- Neamatallah, T.; Malebari, A.M.; Alamoudi, A.J.; Nazreen, S.; Alam, M.M.; Bin-Melaih, H.H.; Abuzinadah, O.A.; Badr-Eldin, S.M.; Alhassani, G.; Makki, L.; et al. Andrographolide nanophytosomes exhibit enhanced cellular delivery and pro-apoptotic activities in HepG2 liver cancer cells. Drug Deliv. 2023, 30, 2174209. [Google Scholar] [CrossRef]
- Pekthong, D.; Blanchard, N.; Abadie, C.; Bonet, A.; Heyd, B.; Mantion, G.; Berthelot, A.; Richert, L.; Martin, H. Effects of Andrographis paniculata extract and Andrographolide on hepatic cytochrome P450 mRNA expression and monooxygenase activities after in vivo administration to rats and in vitro in rat and human hepatocyte cultures. Chem.-Biol. Interact. 2009, 179, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Ooi, J.P.; Kuroyanagi, M.; Sulaiman, S.F.; Muhammad, T.S.; Tan, M.L. Andrographolide and 14-deoxy-11, 12-didehydroandrographolide inhibit cytochrome P450s in HepG2 hepatoma cells. Life Sci. 2011, 88, 447–454. [Google Scholar] [CrossRef]
- Mondal, M.; Sarkar, C.; Saha, S.; Hossain, M.N.; Norouzi, R.; Mubarak, M.S.; Siyadatpanah, A.; Wilairatana, P.; Hossain, R.; Islam, M.T.; et al. Hepatoprotective activity of andrographolide possibly through antioxidative defense mechanism in Sprague-Dawley rats. Toxicol. Rep. 2022, 9, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Shukla, B.; Visen, P.K.; Patnaik, G.K.; Dhawan, B.N. Choleretic effect of andrographolide in rats and guinea pigs. Planta Med. 1992, 58, 146–149. [Google Scholar] [CrossRef]
- Roy, P.; Das, S.; Auddy, R.G.; Saha, A.; Mukherjee, A. Engineered andrographolide nanoparticles mitigate paracetamol hepatotoxicity in mice. Pharm. Res. 2013, 30, 1252–1262. [Google Scholar] [CrossRef]
- He, S.; Cui, X.; Khan, A.; Liu, Y.; Wang, Y.; Cui, Q.; Zhao, T.; Cao, J.; Cheng, G. Activity Guided Isolation of Phenolic Compositions from Anneslea fragrans Wall. and Their Cytoprotective Effect against Hydrogen Peroxide Induced Oxidative Stress in HepG2 Cells. Molecules 2021, 26, 3690. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Wang, Y.; Su, Z.; Kou, R.; Xie, K.; Song, F. Activation of p62-keap1-Nrf2 antioxidant pathway in the early stage of acetaminophen-induced acute liver injury in mice. Chem.-Biol. Interact. 2018, 282, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.H.; Li, J.; Zhang, X.Y.; Shi, S.; Wang, L.; Yuan, M.L.; Liu, Y.P.; Wang, Y.D. Confusoside from Anneslea fragrans Alleviates Acetaminophen-Induced Liver Injury in HepG2 via PI3K-CASP3 Signaling Pathway. Molecules 2023, 28, 1932. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.A.; Hwang, Y.; Hwang, H.J.; Park, N. Hepatoprotective Effects of Radish (Raphanus sativus L.) on Acetaminophen-Induced Liver Damage via Inhibiting Oxidative Stress and Apoptosis. Nutrients 2022, 14, 5082. [Google Scholar] [CrossRef]
- Sarkar, C.; Mondal, M.; Al-Khafaji, K.; El-Kersh, D.M.; Jamaddar, S.; Ray, P.; Roy, U.K.; Afroze, M.; Moniruzzaman, M.; Khan, M.; et al. GC-MS analysis, and evaluation of protective effect of Piper chaba stem bark against paracetamol-induced liver damage in Sprague-Dawley rats: Possible defensive mechanism by targeting CYP2E1 enzyme through in silico study. Life Sci. 2022, 309, 121044. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.; Sun-Waterhouse, D. The potential of dandelion in the fight against gastrointestinal diseases: A review. J. Ethnopharmacol. 2022, 293, 115272. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Castejon, M.; Visioli, F.; Rodriguez-Casado, A. Diverse biological activities of dandelion. Nutr. Rev. 2012, 70, 534–547. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, L.; Wang, J.; Li, J.; Ding, J.; He, X. Protective Effect of Dandelion Leaf Water Extracts on APAP-Induced Liver Injury in Rats and Its Mechanism. Cell. Mol. Biol. 2022, 68, 24–33. [Google Scholar] [CrossRef]
- Ren, Y.S.; Zheng, Y.; Duan, H.; Lei, L.; Deng, X.; Liu, X.Q.; Mei, Z.N.; Deng, X.K. Dandelion polyphenols protect against acetaminophen-induced hepatotoxicity in mice via activation of the Nrf-2/HO-1 pathway and inhibition of the JNK signaling pathway. Chin. J. Nat. Med. 2020, 18, 103–113. [Google Scholar] [CrossRef]
- Zheng, Y.; Lei, L.; Liang, S.; Ai, J.; Deng, X.; Li, Y.Q.; Zhang, T.P.; Pu, S.B.; Ren, Y.S. Protective Effect of Fresh/Dry Dandelion Extracts on APAP-Overdose-Induced Acute Liver Injury. Chin. J. Integr. Med. 2022, 28, 683–692. [Google Scholar] [CrossRef]
- Fadil, H.A.E.; Behairy, A.; Ebraheim, L.L.M.; Abd-Elhakim, Y.M.; Fathy, H.H. The palliative effect of mulberry leaf and olive leaf ethanolic extracts on hepatic CYP2E1 and caspase-3 immunoexpression and oxidative damage induced by paracetamol in male rats. Environ. Sci. Pollut. Res. Int. 2023, 30, 41682–41699. [Google Scholar] [CrossRef]
- Ayenew, K.D.; Wasihun, Y. Hepatoprotective effect of methanol extract of Agave americana leaves on paracetamol induced hepatotoxicity in Wistar albino rats. BMC Complement. Med. Ther. 2023, 23, 99. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hong, J.; Lee, J.; Jeon, W.; Yeo, C.; Lee, Y.; Baek, S.; Ha, I. Curcuma aromatica Salisb. Protects from Acetaminophen-Induced Hepatotoxicity by Regulating the Sirt1/HO-1 Signaling Pathway. Nutrients 2023, 15, 808. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, J.; Jin, H.; Gu, D.; Wang, Q.; Liu, Y.; Zan, K.; Fan, J.; Wang, R.; Wei, F.; et al. Comparisons of physicochemical features and hepatoprotective potentials of unprocessed and processed polysaccharides from Polygonum multiflorum Thunb. Int. J. Biol. Macromol. 2023, 235, 123901. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yang, L.; Zhou, J.; Pan, X.; He, Z.; Liu, J.; Zhang, Y. Smilax china L. Polysaccharide Alleviates Oxidative Stress and Protects from Acetaminophen-Induced Hepatotoxicity via Activating the Nrf2-ARE Pathway. Front. Pharmacol. 2022, 13, 888560. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Hao, K.; Chen, X.; Wu, E.; Nie, D.; Zhang, G.; Si, H. Broussonetia papyrifera Polysaccharide Alleviated Acetaminophen-Induced Liver Injury by Regulating the Intestinal Flora. Nutrients 2022, 14, 2636. [Google Scholar] [CrossRef] [PubMed]
- Baponwa, O.; Amang, A.P.; Mezui, C.; Koubala, B.B.; Siwe, G.T.; Vandi, V.L.; Tan, P.V. Antioxidant Mechanism of Renal and Hepatic Failure Prevention Related to Paracetamol Overdose by the Aqueous Extract of Amblygonocarpus andongensis Stem Bark. BioMed Res. Int. 2022, 2022, 1846558. [Google Scholar] [CrossRef] [PubMed]
- Adetuyi, B.O.; Adebisi, O.A.; Adetuyi, O.A.; Ogunlana, O.O.; Toloyai, P.E.; Egbuna, C.; Uche, C.Z.; Khan, J.; Adumanya, O.C.U.; Patrick-Iwuanyanwu, K.C. Ficus exasperata Attenuates Acetaminophen-Induced Hepatic Damage via NF-κB Signaling Mechanism in Experimental Rat Model. BioMed Res. Int. 2022, 2022, 6032511. [Google Scholar] [CrossRef]
- Geresu, G.D.; Umer, S.; Arayaselassie, M.; Ashebir, G.; Makonnen, E. Hepatoprotective Effects of Crude Stem Bark Extracts and Solvent Fractions of Cordia africana against Acetaminophen-Induced Liver Injury in Rats. Can. J. Gastroenterol. Hepatol. 2022, 2022, 1449286. [Google Scholar] [CrossRef]
- Sahin, B.; Karabulut, S.; Filiz, A.K.; Ozkaraca, M.; Gezer, A.; Akpulat, H.A.; Ataseven, H. Galium aparine L. protects against acetaminophen-induced hepatotoxicity in rats. Chem.-Biol. Interact. 2022, 366, 110119. [Google Scholar] [CrossRef]
- Elsayed, H.E.; Ebrahim, H.Y.; Mady, M.S.; Khattab, M.A.; El-Sayed, E.K.; Moharram, F.A. Ethnopharmacological impact of Melaleuca rugulosa (Link) Craven leaves extract on liver inflammation. J. Ethnopharmacol. 2022, 292, 115215. [Google Scholar] [CrossRef]
- Ismail, S.; Naveed Mushtaq, M.; Shabbir, U.; Saima; Razzaq, A.; Ramzan, H.M.; Shoukat, A.; Ahsan, H.; Movafagh, A.; Kahrizi, D.; et al. Evaluation of Hepatoprotective and Gastroprotective Activities of Paspalidium flavidum Leaves Extract in Experimental Animal Models. Cell. Mol. Biol. 2022, 68, 8–14. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Aleem, A.; Hussain, S.A.; Manzoor, M.; Latif, S.; Sarwar, M.I.; Khan, M.; Talib, S. Hepatoprotective effect of methanolic extract of Iris florentina L. on paracetamol-induced liver toxicity in rats. Pak. J. Pharm. Sci. 2022, 35, 1399–1405. [Google Scholar] [PubMed]
- Feki, F.; Mahmoudi, A.; Denev, P.; Feki, I.; Ognyanov, M.; Georgiev, Y.; Choura, S.; Chamkha, M.; Trendafilova, A.; Sayadi, S. A jojoba (Simmondsia chinensis) seed cake extracts express hepatoprotective activity against paracetamol-induced toxicity in rats. Biomed. Pharmacother. 2022, 153, 113371. [Google Scholar] [CrossRef] [PubMed]
- Chariyakornkul, A.; Juengwiroj, W.; Ruangsuriya, J.; Wongpoomchai, R. Antioxidant Extract from Cleistocalyx nervosum var. paniala Pulp Ameliorates Acetaminophen-Induced Acute Hepatotoxicity in Rats. Molecules 2022, 27, 553. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Bhattacharya, S.; Choudhury, P.R.; Rahaman, A.; Sarkar, A.; Talukdar, A.D.; Mandal, D.P.; Bhattacharjee, S. Drynaria quercifolia suppresses paracetamol-induced hepatotoxicity in mice by inducing Nrf-2. Bratisl. Lek. Listy 2022, 123, 110–119. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, C.; Kou, J.; Wang, M.; Rong, X.; Pu, X.; Xie, X.; Han, G.; Pang, X. Chrysanthemi Flos extract alleviated acetaminophen-induced rat liver injury via inhibiting oxidative stress and apoptosis based on network pharmacology analysis. Pharm. Biol. 2021, 59, 1376–1385. [Google Scholar] [CrossRef]
- Sinaga, E.; Fitrayadi, A.; Asrori, A.; Rahayu, S.E.; Suprihatin, S.; Prasasty, V.D. Hepatoprotective effect of Pandanus odoratissimus seed extracts on paracetamol-induced rats. Pharm. Biol. 2021, 59, 31–39. [Google Scholar] [CrossRef]
- Ahmad, S.; Zeb, A.; Khan, S. Effects of aqueous extract of Medicago denticulata against paracetamol-induced hepatotoxicity in rabbits. J. Food Biochem. 2021, 45, e13985. [Google Scholar] [CrossRef]
- Zhou, Z.; Qi, J.; Zhao, J.; Seo, J.H.; Shin, D.G.; Cha, J.D.; Lim, C.W.; Kim, J.W.; Kim, B. Orostachys japonicus ameliorates acetaminophen-induced acute liver injury in mice. J. Ethnopharmacol. 2021, 265, 113392. [Google Scholar] [CrossRef] [PubMed]
- Park, D.W.; Jeon, H.; Kwon, J.E.; Lee, Y.G.; So, R.; Choe, T.H.; Jeong, Y.J.; Kang, S.C. Hepatoprotective effect of Centella asiatica 50% ethanol extract against acetaminophen-induced acute liver injury in BALB/c mice. Toxicol. Res. 2021, 37, 261–275. [Google Scholar] [CrossRef]
- Jiang, Z.; Han, Y.; Zhang, Y.; Li, J.; Liu, C. Sedum sarmentosum Bunge Attenuates Drug-Induced Liver Injury via Nrf2 Signaling Pathway: An Experimental Verification Based on Network Pharmacology Prediction. J. Healthc. Eng. 2021, 2021, 1142638. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.A.; Rahman, M.A.; Hossen, M.A.; Reza, A.; Islam, M.S.; Rashid, M.M.; Rafi, M.K.J.; Siddiqui, M.T.A.; Al-Noman, A.; Uddin, M.N. Epiphytic Acampe ochracea orchid relieves paracetamol-induced hepatotoxicity by inhibiting oxidative stress and upregulating antioxidant genes in in vivo and virtual screening. Biomed. Pharmacother. 2021, 143, 112215. [Google Scholar] [CrossRef]
- Bouhlali, E.D.T.; Derouich, M.; Hmidani, A.; Bourkhis, B.; Khouya, T.; Filali-Zegzouti, Y.; Alem, C. Protective Effect of Phoenix dactylifera L. Seeds against Paracetamol-Induced Hepatotoxicity in Rats: A Comparison with Vitamin C. Sci. World J. 2021, 2021, 6618273. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Yun, D.H.; Lee, J.H.; Kwon, Y.B.; Lee, Y.M.; Lee, D.H.; Kim, D.K. Extract of Triticum aestivum Sprouts Suppresses Acetaminophen-Induced Hepatotoxicity in Mice by Inhibiting Oxidative Stress. Molecules 2021, 26, 6336. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, D.M.; Emad, A.M.; Ali, S.F.; Ali, S.S.; Farag, M.A.; Meselhy, M.R.; Sattar, E.A. UPLC-PDA-ESI/MS metabolic profiling of dill shoots bioactive fraction; evidence of its antioxidant and hepatoprotective effects in vitro and in vivo. J. Food Biochem. 2021, 45, e13741. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Li, D.; Ma, H.; Mu, Y.; Zheng, D.; Huang, X.; Li, L. Chemical Characterization and Hepatoprotective Effects of a Standardized Triterpenoid-Enriched Guava Leaf Extract. J. Agric. Food Chem. 2021, 69, 3626–3637. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Z.A.; Sahmat, A.; Azmi, A.H.; Nur Zainol, A.S.; Omar, M.H.; Balan, T.; Sulistyorini, L.; Azizah, R.; Abdullah, M.N.H. Polyphenolics and triterpenes presence in chloroform extract of Dicranopteris linearis leaves attenuated paracetamol-induced liver intoxication in rat. BMC Complement. Med. Ther. 2021, 21, 35. [Google Scholar] [CrossRef]
- Zakaria, Z.A.; Kamisan, F.H.; Kek, T.L.; Salleh, M.Z. Hepatoprotective and antioxidant activities of Dicranopteris linearis leaf extract against paracetamol-induced liver intoxication in rats. Pharm. Biol. 2020, 58, 478–489. [Google Scholar] [CrossRef]
- Ma, H.C.; Ren, H.Z.; Wang, J.; Yuan, X.W.; Wu, X.Y.; Shi, X.L. Targeting PI3K/Akt/Nrf2 pathway by glabridin alleviates acetaminophen-induced hepatic injury in rats. Arab. J. Chem. 2021, 14, 102968. [Google Scholar] [CrossRef]
- Li, X.; Li, M.; Deng, S.; Yu, T.; Ma, Y.; Yang, H.; Zhang, J.; Zheng, Y.; Ma, L. A network pharmacology-integrated metabolomics strategy for clarifying the action mechanisms of Schisandrae Chinensis Fructus for treating drug-induced liver injury by acetaminophen. Bioorg. Med. Chem. 2021, 31, 115992. [Google Scholar] [CrossRef]
- Yan, C.; Guo, H.; Ding, Q.; Shao, Y.; Kang, D.; Yu, T.; Li, C.; Huang, H.; Du, Y.; Wang, H.; et al. Multiomics Profiling Reveals Protective Function of Schisandra Lignans against Acetaminophen-Induced Hepatotoxicity. Drug Metab. Dispos. Biol. Fate Chem. 2020, 48, 1092–1103. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Sun, J.; Cao, X.; Liu, M.; Liu, Y. Steamed ginseng shoot extract rich in less-polar ginsenosides ameliorated the acute hepatotoxicity caused by overdose of acetaminophen in mice. J. Funct. Foods 2020, 73, 104155. [Google Scholar] [CrossRef]
- Ding, C.H.; Zhu, H. Isatidis Folium alleviates acetaminophen-induced liver injury in mice by enhancing the endogenous antioxidant system. Environ. Toxicol. 2020, 35, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ponce, H.A.; Martinez-Saldana, M.C.; Tepper, P.G.; Quax, W.J.; Buist-Homan, M.; Faber, K.N.; Moshage, H. Betacyanins, major components in Opuntia red-purple fruits, protect against acetaminophen-induced acute liver failure. Food Res. Int. 2020, 137, 109461. [Google Scholar] [CrossRef]
- Salman, A.A.; El-Aleem, I.M.A.; El-Rahman, A.A.A.; El-Husseiny, T.S.; El-Hadary, A.E. Assessment of antioxidant traits and protective action of Egyptian acacia pods extracts against paracetamol-induced liver toxicity in rats. J. Food Biochem. 2020, 44, e13392. [Google Scholar] [CrossRef] [PubMed]
- Galani, B.R.T.; Owona, B.A.; Chuisseu, D.P.D.; Machewere, E.; Ngantchouko, C.B.N.; Moundipa, P.F. Hepatoprotective Activity of Leptadenia hastata (Asclepiadaceae) on Acetaminophen-Induced Toxicity in Mice: In Vivo Study and Characterization of Bioactive Compounds through Molecular Docking Approaches. BioMed Res. Int. 2020, 66, 357–363. [Google Scholar] [CrossRef]
- Shu, Y.; He, D.; Li, W.; Wang, M.; Zhao, S.; Liu, L.; Cao, Z.; Liu, R.; Huang, Y.; Li, H.; et al. Hepatoprotective Effect of Citrus aurantium L. Against APAP-induced Liver Injury by Regulating Liver Lipid Metabolism and Apoptosis. Int. J. Biol. Sci. 2020, 16, 752–765. [Google Scholar] [CrossRef]
- Gatsou Djibersou, D.; Rosnay Tietcheu Galani, B.; Dieudonne Djamen Chuisseu, P.; Yanou Njintang, N. Anti-oxidant and anti-inflammatory potential of aqueous extracts of leaves, barks and roots of Bixa orellana L. (Bixaceae) on acetaminophen-induced liver damage in mice. Avicenna J. Phytomed. 2020, 10, 428–439. [Google Scholar]
- Kawakami, K.; Moritani, C.; Hatanaka, T.; Suzaki, E.; Tsuboi, S. Hepatoprotective Activity of Yellow Chinese Chive against Acetaminophen-Induced Acute Liver Injury via Nrf2 Signaling Pathway. J. Nutr. Sci. Vitaminol. 2020, 66, 357–363. [Google Scholar] [CrossRef]
- Liu, J.; Luo, D.; Wu, Y.; Gao, C.; Lin, G.; Chen, J.; Wu, X.; Zhang, Q.; Cai, J.; Su, Z. The Protective Effect of Sonneratia apetala Fruit Extract on Acetaminophen-Induced Liver Injury in Mice. Evid.-Based Complement. Altern. Med. eCAM 2019, 2019, 6919834. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Abdel Moneim, A.E.; Hafez, T.A.; Mubaraki, M.A.; Mohamed, W.F.; Thagfan, F.A.; Al-Quraishy, S. Myristica fragrans Kernels Prevent Paracetamol-Induced Hepatotoxicity by Inducing Anti-Apoptotic Genes and Nrf2/HO-1 Pathway. Int. J. Mol. Sci. 2019, 20, 993. [Google Scholar] [CrossRef] [PubMed]
- Azarmehr, N.; Afshar, P.; Moradi, M.; Sadeghi, H.; Sadeghi, H.; Alipoor, B.; Khalvati, B.; Barmoudeh, Z.; Abbaszadeh-Goudarzi, K.; Doustimotlagh, A.H. Hepatoprotective and antioxidant activity of watercress extract on acetaminophen-induced hepatotoxicity in rats. Heliyon 2019, 5, e02072. [Google Scholar] [CrossRef] [PubMed]
- Odeyemi, S.; Dewar, J. Repression of Acetaminophen-Induced Hepatotoxicity in HepG2 Cells by Polyphenolic Compounds from Lauridia tetragona (L.f.) R.H. Archer. Molecules 2019, 24, 2118. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Tan, S.; Gu, W.; Li, F.; Hua, W.; Zhang, S.; Chen, F.; Tang, L. Phytochemical composition, isolation and hepatoprotective activity of active fraction from Veronica ciliata against acetaminophen-induced acute liver injury via p62-Keap1-Nrf2 signaling pathway. J. Ethnopharmacol. 2019, 243, 112089. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.J.; Li, M.J.; Zhang, M.P.; Wei, M.K.; Shen, L.P.; Jiang, M.; Zeng, T. Allyl methyl trisulfide protected against acetaminophen (paracetamol)-induced hepatotoxicity by suppressing CYP2E1 and activating Nrf2 in mouse liver. Food Funct. 2019, 10, 2244–2253. [Google Scholar] [CrossRef]
- Kaur, P.; Shergill, R.; Mehta, R.G.; Singh, B.; Arora, S. Biofunctional significance of multi-herbal combination against paracetamol-induced hepatotoxicity in Wistar rats. Environ. Sci. Pollut. Res. Int. 2021, 28, 61021–61046. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, Q.; Liu, W.; Lin, Y.; He, Y.; Chang, D.; Li, S.; Xu, W.; Lin, Y.; Zheng, Y.; et al. Pien Tze Huang attenuated acetaminophen-induced liver injury by autophagy mediated-NLRP3 inflammasome inhibition. J. Ethnopharmacol. 2023, 311, 116285. [Google Scholar] [CrossRef]
- Zhang, X.R.; Lin, T.; Wang, X.L.; Wang, X.J.; Gu, H. Preparation of salvianolic acid B, tanshinone II_A, and glycyrrhetinic acid lipid emulsion and its protective effect against acute liver injury induced by acetaminophen. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Medica 2022, 47, 4634–4642. [Google Scholar] [CrossRef]
- Kataki, M.S.; Kakoti, B.B. An Optimized Herbal Formula Reverses the Hepatotoxicity Induced by Acetaminophen. J. Young Pharm. 2022, 14, 56–61. [Google Scholar] [CrossRef]
- Gao, Y.; Shi, W.; Yao, H.; Ai, Y.; Li, R.; Wang, Z.; Liu, T.; Dai, W.; Xiao, X.; Zhao, J.; et al. An Integrative Pharmacology Based Analysis of Refined Liuweiwuling Against Liver Injury: A Novel Component Combination and Hepaprotective Mechanism. Front. Pharmacol. 2021, 12, 747010. [Google Scholar] [CrossRef]
- Jin, S.E.; Shin, H.K.; Ha, H. Hepatoprotective effects of Gamisoyo-san against acetaminophen-induced liver injuries. Integr. Med. Res. 2021, 10, 100466. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.W.; Wu, W.J.; Lu, C.W.; Wang, S.E.; Chuang, W.C.; Lee, M.C.; Wu, C.H. Traditional Chinese Medicine Yang-Gan-Wan Alleviated Experimental Hepatic Damage by Inhibiting Oxidation, Inflammation, and Apoptosis in Cell and Mouse Models. Evid.-Based Complement. Altern. Med. eCAM 2021, 2021, 2556352. [Google Scholar] [CrossRef] [PubMed]
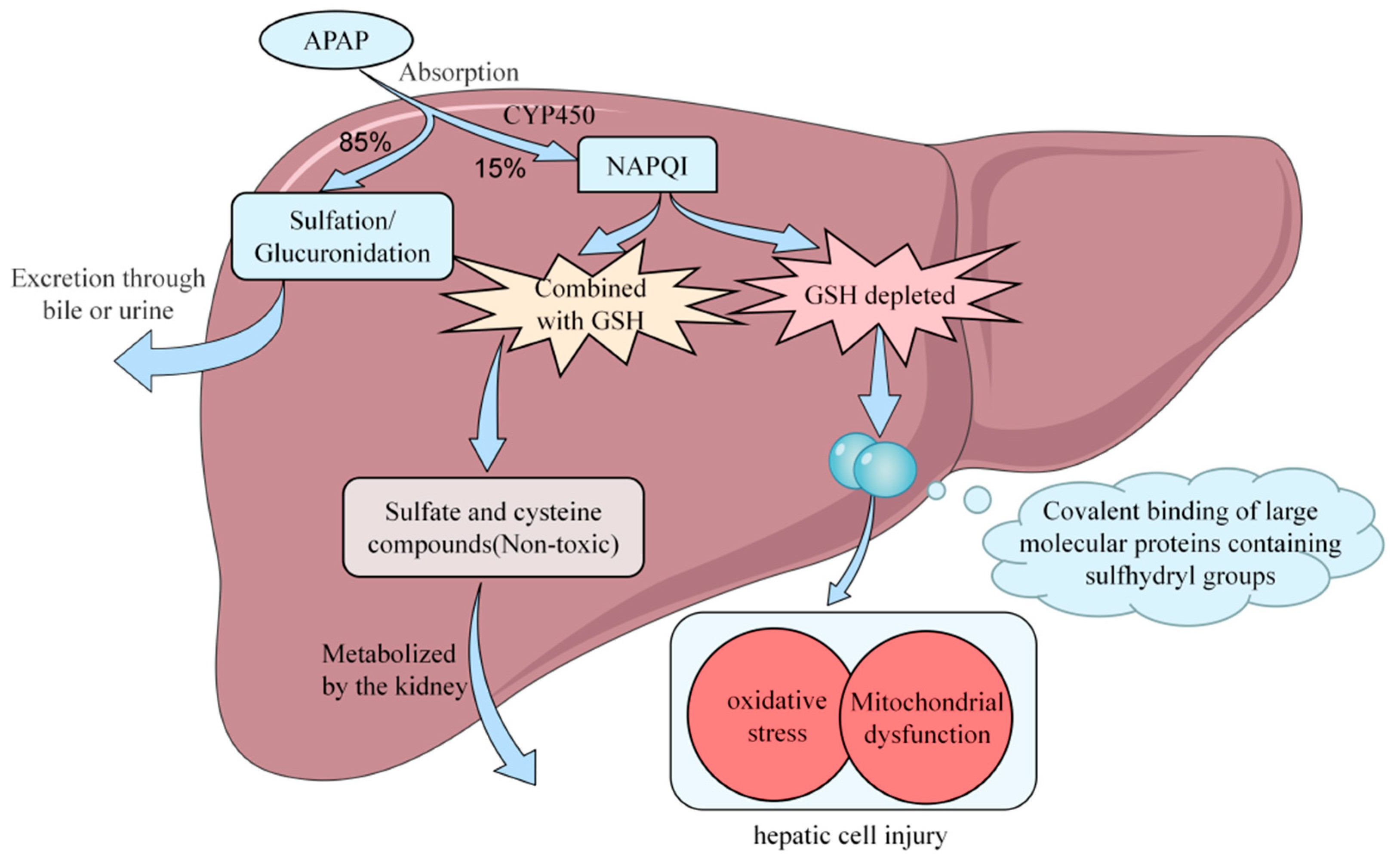

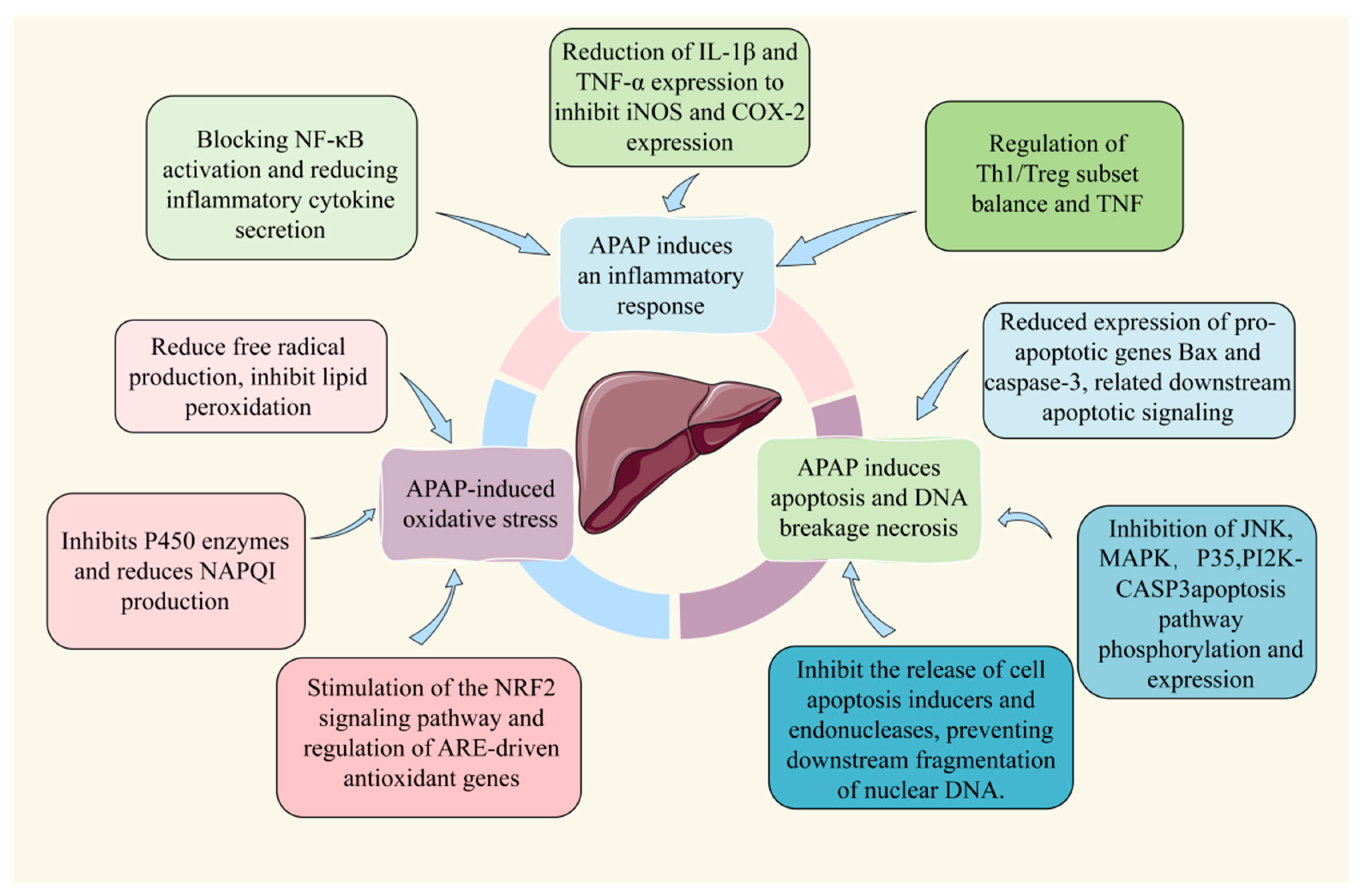

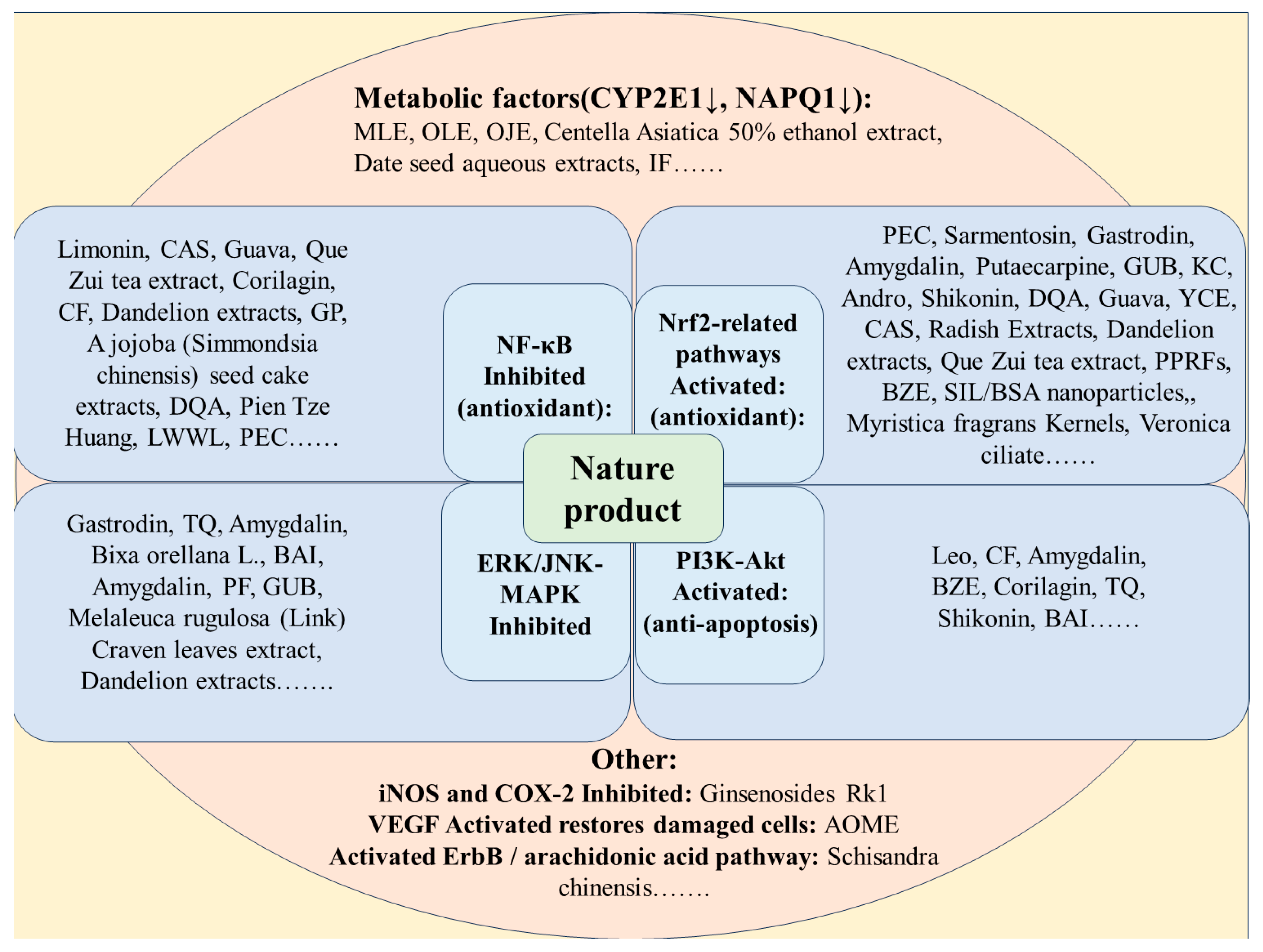
| Phytochemicals | Dose, Route, and Duration of Administration | Model | APAP Dose and Route of Administration | Findings and Key Mechanisms | Reference |
|---|---|---|---|---|---|
| Pectolinarigenin (PEC) | 5 and 10 mg/kg daily, i.p. for 7 days | Male C6BL/8 mice | 400 mg/kg, i.p. single dose, on 7th day after the final dose 2 h | Activate Nrf2 and PPARα signaling, decreasing hepatic oxidative stress and inflammation, increasing phase IIdetoxification enzymes related to APAP metabolism. | Li, et al., 2023 [77] |
| 0, 0.25, 0.5, 1, 2, and 5 μM for 48 h before APAP administration | HepG2 cells | 0, 2.5, 5, 10, 20, and 40 mM for 6, 12, and 24 h. | |||
| Leonurine (Leo) | 20 and 40 mg/kg i.g. for 7 days | Male ICR mice | 300 mg/kg, i.p. single dose | Alleviated ALI by modulating the PI3K/AKT signaling pathway. | Yu, et al., 2023 [78] |
| 10 μΜ Leo for 12 h | Mouse primary hepatocytes | DMEM with 10 mM APAP for 24 h | |||
| Corilagin (CL) | 0, 1, 5, or 10 mg/kg i.p. after APAP administration | Male C57BL/6C mice | 300 mg/kg, i.p. single dose | Involve the regulation of IL-6/STAT3 and MAPK/NF-κB pathways through NOX-derived ROS. | Liu, et al., 2023 [79] |
| 15, 30, 60 mg/kg i.p. twice for one day | Male C57BL/6 mice | 400, 900 mg/kg, i.p. single dose | Inhibited AMPK, upregulated GSK3β, and activated the Nrf2 pathway. | Lv, et al., 2019 [80] | |
| 7.5, 15, or 30 μM, treated for 25 h | HepG2 cells | 15 mM, treated for 24 h | |||
| Confusoside (CF) | 50, 100 μg/mL | Hepatoma HepG2 cells | 10 mM | Inhibiting the NF-κB-regulated inflammatory response and PI3K/Akt-regulated apoptosis. | Zhao, et al., 2023 [81] |
| Rosmarinic acid (RA) | 20, 40, or 80 mg/kg, Unknown route of administration, for seven days | Male Kunming mice | 500 mg/kg, i.p. single dose | Upregulated Nrf2 pathway and inhibited NEK7-NLRP3 pathway. | Yao, et al., 2022 [82] |
| 20, 40, or 80 μM | HepG2 cells | 25 mM | |||
| Cajaninstilbene acid (CSA) | 75 mg/kg, p.o. after APAP administration | Male C57BL/6 mice | 300 mg/kg, i.p. single dose | Activated Sestrin2-LKB1-AMPK pathway, enhanced mitochondrial quality control, and inhibited oxidative stress. | Yan, et al., 2022 [83] |
| 50 μM | Mouse primary hepatocytes | 5 and 10 mM | |||
| Sarmentosin (SMT) | 0.5, 1, 2 μM to L-02 and 5, 10, 20 μM to AML-12 cells for 2 h | L-02 and AML-12 cells | 20 mM for 2 h single dose | Regulating Nrf2 to alleviate APAP-induced hepatocytes oxidative damage. | Jiang, et al., 2022 [84] |
| 20, 40, and 80 mg/kg for a week | Male ICR mice | 300 mg/kg i.p. single dose | |||
| Silybin A/B (SB A/B) | 20 mg/kg, single dose, 30 min after APAP administration. | BALB/c mice | 300 mg/kg, i.p. single dose | Regulated the Nrf2/ARE pathway, inhibited mitochondrial damage and apoptosis pathway associated with caspase-9/3. | Ding, et al., 2022 [66] |
| Schisandrin C (Sc C) | 200 mg/kg, i.g. for 7 days | Male C57BL/6 mice | 400 mg/kg, i.p. single dose | Regulated the Nrf2 signaling pathway and reduced the expression of inflammatory factors and CYP2E1. | Dai, et al., 2022 [68] |
| Schisandrol A (Sc A) | 10 g/kg, i.g. one time each day for 7 days. | Male ICR mice | 200 mg/kg, i.p. single dose, after the last intragastric administration for 1 h | Activated TNF signaling pathway, and inhibited the activities of cytochrome P450 enzymes. | Li, et al., 2022 [85] |
| Gastrodin (GAS) | 0, 15, 30, or 45 mg/kg, i.p. single dose, at 30 min after APAP administration. | Adult C57BL/6 (B6) mice | 300 mg/kg, i.p. single dose | Inhibited ERK/JNK MAPK signaling pathway while activating Nrf2 signaling pathway | Liao, et al., 2022 [86] |
| Amygdalin (AG) | 2.5, 5 mg/kg i.p. at the same time | Male C57BL/6 mice | 400 mg/kg, i.p. after fasting for 15–17 h; the mice were intraperitoneally injected. | Inhibit apoptosis and necrosis, alleviate inflammation and oxidative stress, and even reverse liver injury by activating the AKT/JNK/Nrf2 pathway. | Zhang, et al., 2022 [87] |
| Paeoniflorin (PF) | 60 mg/kg, i.g., for 5 days, once a day. | Male C57BL/6 mice | 250 mg/kg, i.p. single dose | Regulated CYP2E1/JNK signaling. | Deng, et al., 2022 [88] |
| Diacerein (DIA) | 25 and 50 mg/kg, p.o. for 7 days | Male albino rats | 1 g/kg, p.o. single dose | Reduced oxidative stress, anti-inflammatory, anti-apoptotic, elevated PPAR-γ expression, and inhibited HMGB-1/TLR4/NF-κB signaling pathway. | Mohamed Kamel, et al., 2022 [89] |
| Emodin (EMO) | 15 and 30 mg/kg, p.o. for 5 consecutive days. | Male C57BL/6 mice | 300 mg/kg, i.p. single dose | Inhibited cCGAS-STING pathways. | Shen, et al., 2022 [90] |
| Oridonin (OD) | 20 mg/kg, i.p. after APAP administration 1 h | Male C57BL/6 mice | 300 mg/kg, i.g. single dose | Targeted the B. vulgatus–urea cycle–Nrf2 axis and enriched the gut microbiota regulation of B. vulgatus. | Hong, et al., 2021 [91] |
| Rutaecarpine (RCP) | 5 or 20 mg/kg once daily for 7 days | Male ICR mice | 300 mg/kg, i.p. single dose | Inhibited CYP2E1 expression, inhibited lipid peroxidation, and activation of Nrf2 pathway. | Choi, et al., 2021 [92] |
| Guavinoside B (GUB) | 3.3, 10, 30, and 100 μM | HepG2 cells | 5 mM. | Activated the Nrf2 pathway, and inhibited the JNK pathway. | Li, et al., 2020 [93] |
| 100 mg/kg/day, i.g. for 7 consecutive days | Male C57BL/6 mice | 300 mg/kg, i.p. single dose, after the final administration | |||
| kushenol C (KC) | 1, 10, 20 mg/kg, p.o. every day for seven days. | Male BALB/c mice | 500 mg/kg, i.p. single dose, after the final administration | Regulated Caspase 3, Bax/Bcl-2, GSH, and ROS production and activated the AKT-mediated Nrf2 pathway. | Cho, et al., 2021 [94] |
| Various concentrations for 1 h | HepG2 cells | 1 mM tBHP, for 24 h | |||
| Kaempferol (KP) | 250 mg/kg, p.o. for 7 days | Male Wistar rats | 800 mg/kg, i.p. single dose | Activated SIRT1, inhibited PARP1 and CYP2E1. | BinMowyna, et al., 2021 [95] |
| Ginsenoside Rg3 (GR3) | 5, 10, and 20 mg/kg, p.o. after administration APAP | Male C57BL/6J mice | 350 mg/kg, p.o. single dose | Regulated NLRP3 pathway. | Gao, et al., 2021 [76] |
| Ginsenoside Rk1 (GK1) | 10 mg/kg, 20 mg/kg, i.g. per day for a week | Male ICR mice | 250 mg/kg, i.p. single dose | Inhibited the increase of iNOS and COX-2 expression. Regulated Bax and Bcl-2. | Hu, et al., 2019 [74] |
| Ginsenoside Rg1 (GR1) | Treated for 24 h | HepG2 and HEK293 cells | 16 mM for 24 h | Activated Nrf2/ARE signaling pathway. Decreased oxidative stress. Inhibited ROS. Decreased the activation of the downstream caspase-8/9 to inhibit apoptosis of cells. | Gao, et al., 2019 [96] |
| No treatment | Male C57BL/6 mice | 500 mg/kg, i.g. single dose | |||
| Limonin (LM) | 10, 25, 50 μM, for 2 h | L-02 cells | 7.5 mM, treated for 24 h | Upregulated Sirt1, inhibited NF-κB inflammatory response, and activated Nrf2 antioxidant signaling. | Yang, et al., 2020 [97] |
| 40, 80 mg/kg, p.o. for 1 h | Male C57BL/6 mice | 300 mg/kg, i.p. single dose | |||
| Apigenin (API) | 20 and 80 mg/kg for 7 consecutive days. | Male C57BL/6 mice | 400 mg/kg, p.o. single dose | Interacted with SIRT1 and activated autophagy. | Zhao, et al., 2020 [98] |
| 5, 50 μM | L-02 cells | 10 mM, for 48 h | |||
| Sinomenine (SIN) | 25, 50, and 100 mg/kg, unknown route of administration, for 7 days | Male C57BL/6 mice | 250 mg/kg, i.p. single dose | Inhibit the activation of NLRP3. Activated TGF-B/Smad pathway. | Chen, et al., 2020 [99] |
| 10, 50, and 100 μg/mL for 24 h. | BRL-3A cell | 75 mM, single dose | |||
| Dihydromyricetin (DHM) | 25, 100, 200 mg/kg, i.g. daily for 5 days | Male C57BL/6 mice | 300 mg/kg, i.p. single dose | Inhibited hepatocyte death, promoted regeneration, and regulated the lipid homeostasis imbalance mediated by PPARs and SREBP-1c. | Dong, et al., 2019 [100] |
| γ-Oryzanol (ORY) | 5 and 10 μg/mL for 24 h | L-02 cells | 15 mM | Inhibited NF-κB p65 pathway, inhibited AMPK, upregulated GSK3β, and activated the Nrf2 pathway. | Shu, et al., 2019 [101] |
| 7 and 14 mg/kg, i.g. for 7 days | Male Kunming mice | 300 mg/kg, i.p. single dose | |||
| Fisetin (FST) | 10, 20, and 40 mg/kg, i.g. for 7 days. | Male C57BL/6 mice | 400 mg/kg, i.p. single dose | Induced the increasing of glutathione metabolism and enhanced expression of downstream antioxidative enzymes. | Zhao, et al., 2019 [102] |
| Treated with FST for 15 min | L-02 cells | Incubated with APAP for 24 h | |||
| Thymoquinone (TQ) | 5 or 20 mg/kg, i.p. single dose at 2 h before the administration of APAP | Male Kunming mice | 300 mg/kg, i.p. single dose | Inhibited MAPK and JNK phosphorylation, and activated the AMPK signaling pathway to inhibit the PI3K/mTOR signaling pathways. | Cui, et al. 2019 [103] |
| Geniposide (GP) | 10, 30, and 100 mg/kg, p.o. for three times at 24 h before the injection of APAP | Male C57BL/6 mice | 350 mg/kg, i.p. single dose | Downregulated CYP2E1 expression, ameliorate oxidative stress. Suppressed the TLR4/NF-κB signaling pathway. | Yang, et al., 2019 [104] |
| Tetrahydrocurcu-min (THC) | 25, 50, and 100 mg/kg, i.p. single dose at 30 min before the administration of APAP | Male Kunming mice | 220 mg/kg, i.p. single dose | Inhibited CYP2E1 activity and activated the Nrf2 pathway. | Luo, et al., 2019 [105] |
| Octahydrocurcumin (OHC) | |||||
| Pterostilbene (PTE) | 15, 30, and 60 mg/kg, i.p., single dose at 1 h after the administration | Male ICR mice | 400 mg/kg, i.p. single dose | Restoring impaired autophagic flux. | Kang, et al., 2019 [106] |
| Shikonin (SK) | 12.5, 25.0 mg/kg, i.p., single dose at 1 h before the injection of APAP | Male C57BL/6 mice | 500 mg/Kg, i.p. single dose | Inhibited oxidative stress through AKT/GSK3β pathway-dependent Nrf2 upregulation. | Li, et al., 2018 [107] |
| 0.25, 0.375, and 0.50 μM for 24 h | L-02 cells | 10 mM, for 24 h | |||
| 12.5 mg/kg, i.p. for 2 h | Male BALB/c mice | 300 mg/kg, i.p. single dose | Inhibited the expression of TLR9 and NLRP3, and suppressed oxidative stress. | Guo, et al., 2019 [108] | |
| 5 μM | AML-12 cells | 0, 5, 10, 20, 50, 100 mM for 24 h | |||
| Andrographolide (Andro) | 20, 40 mg/kg, i.g. for 4 weeks | C57BL/6 mice | 300 mg/kg, i.g. single dose | Induced Nrf2 activation and attenuating hepatic oxidative stress injury. | Yan, et al., 2018 [109] |
| Baicalein (BAI) | 100, 50 mg/kg, p.o. per day for a week | Kunming male mice | 350 mg/kg, i.p. single dose | Related to the regulation of AKT-mTOR, MAPK, and JAK2/STAT3 signaling pathways. | Zhou, et al., 2018 [110] |
| Salvianolic acid C (SAC) | 5 mg/kg, 10 mg/kg, and 20 mg/kg, i.p. daily for 6 days | Male ICR mice | 400 mg/kg, i.p. single dose | Activated Nrf2 pathway, inhibited MAPK and NF-κB pathway, inhibited apoptosis and inflammation. | Wu, et al., 2019 [61] |
| Maltol (MT) | 50 and 100 mg/kg, i.p. for 7 days | Male ICR mice | 250 mg/kg, i.p. single dose | Inhibited oxidative-stress-mediated NF-κB pathway activation and apoptosis, and activated PI3K/Akt pathway. | Wang, et al., 2019 [111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Lao, R.; Lei, J.; Chen, Y.; Zhou, Q.; Wang, T.; Tong, Y. Natural Products for Acetaminophen-Induced Acute Liver Injury: A Review. Molecules 2023, 28, 7901. https://doi.org/10.3390/molecules28237901
Li X, Lao R, Lei J, Chen Y, Zhou Q, Wang T, Tong Y. Natural Products for Acetaminophen-Induced Acute Liver Injury: A Review. Molecules. 2023; 28(23):7901. https://doi.org/10.3390/molecules28237901
Chicago/Turabian StyleLi, Xiaoyangzi, Ruyang Lao, Jiawei Lei, Yuting Chen, Qi Zhou, Ting Wang, and Yingpeng Tong. 2023. "Natural Products for Acetaminophen-Induced Acute Liver Injury: A Review" Molecules 28, no. 23: 7901. https://doi.org/10.3390/molecules28237901
APA StyleLi, X., Lao, R., Lei, J., Chen, Y., Zhou, Q., Wang, T., & Tong, Y. (2023). Natural Products for Acetaminophen-Induced Acute Liver Injury: A Review. Molecules, 28(23), 7901. https://doi.org/10.3390/molecules28237901