
Research Advances in Clinical Applications, Anticancer Mechanism, Total Chemical Synthesis, Semi-Synthesis and Biosynthesis of Paclitaxel

 ,
,
Abstract
1. Introduction
2. Anticancer Mechanism and Clinical Applications
3. Sources and Production Methods of Paclitaxel
3.1. Extraction from Taxus Plants
3.2. Total Synthesis
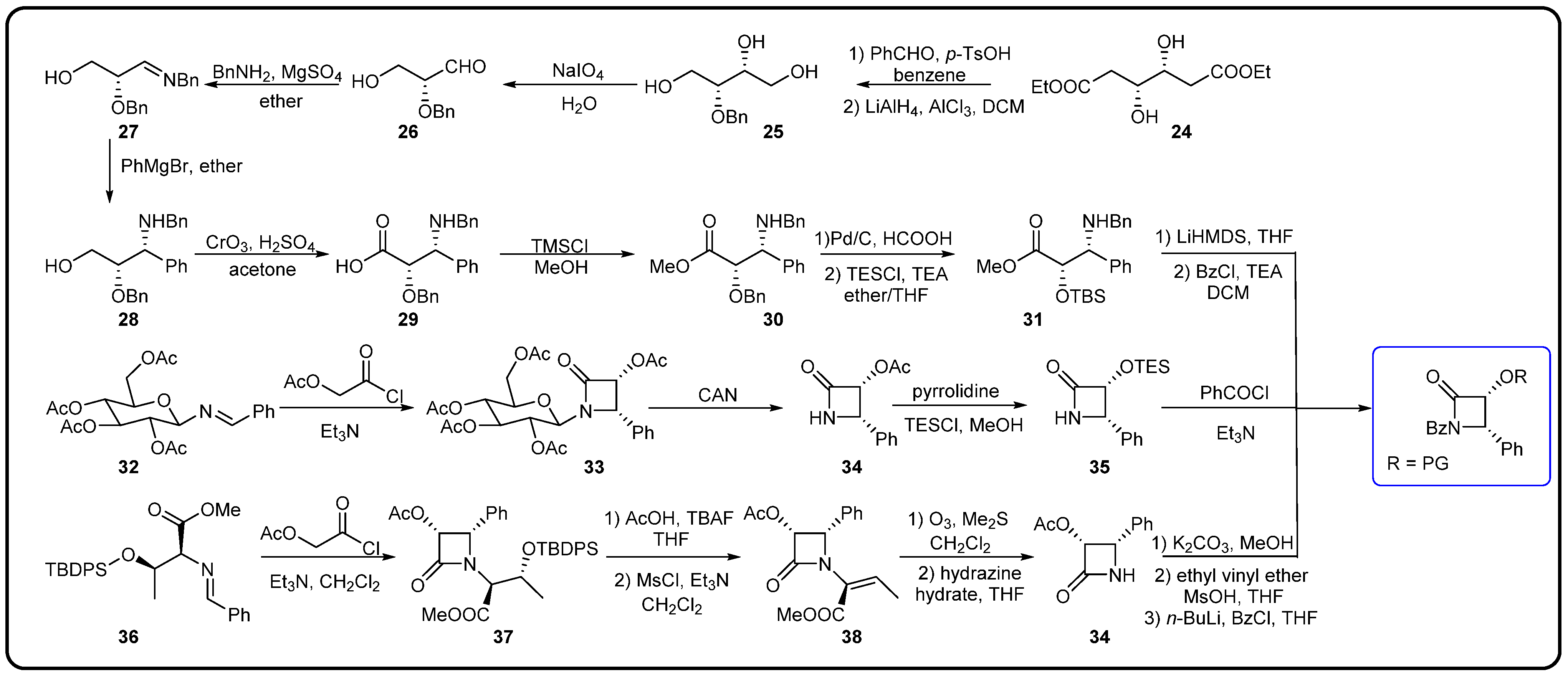
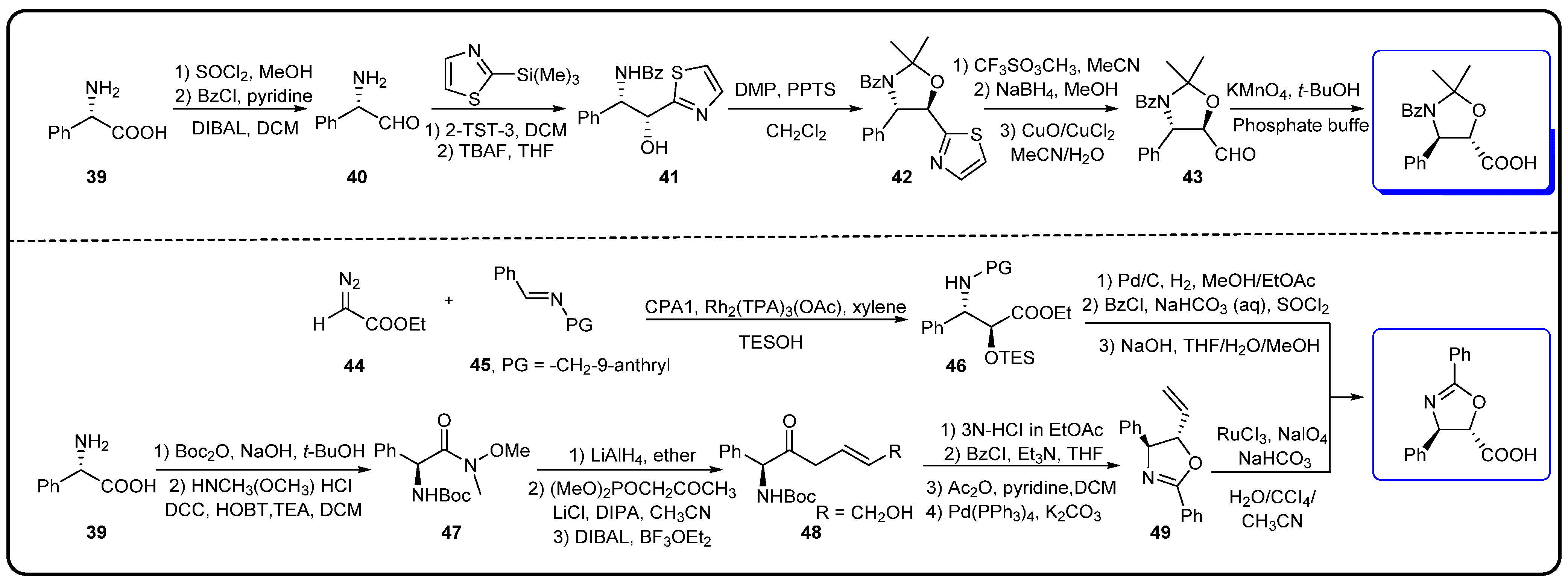
3.3. Semi-Synthesis
3.4. Tissue and Cell Culture
3.5. Paclitaxel-Producing Endophytic Fungi
4. Synthetic Biology Studies of Paclitaxel
4.1. Biosynthetic Pathways
4.2. Ab Initio Biosynthesis of Paclitaxel by Heterologous Systems
4.3. Semi-Synthesis by Microbial Systems
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perez-Matas, E.; Hanano, A.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazon, J. Insights into the control of taxane metabolism: Molecular, cellular, and metabolic changes induced by elicitation in Taxus baccata cell suspensions. Front. Plant Sci. 2022, 13, 942433. [Google Scholar] [CrossRef] [PubMed]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; McPhail, A.T. Plant antitumor agents. VI. Isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef] [PubMed]
- Zefirova, O.N.; Nurieva, E.V.; Ryzhov, A.N.; Zyk, N.V.; Zefirov, N.S. Taxol: Synthesis, Bioactive Conformations, and Structure-Activity Relationships in Its Analogs. Russ. J. Org. Chem. 2005, 41, 315–351. [Google Scholar] [CrossRef]
- Expósito, O.; Bonfill, M.; Moyano, E.; Onrubia, M.; Mirjalili, M.H.; Cusidó, R.M.; Palazón, J. Biotechnological production of taxol and related taxoids: Current state and prospects. Anticancer Agents Med. Chem. 2009, 9, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Schiff, P.B.; Fant, J.; Horwitz, S.B. Promotion of microtubule assembly in vitro by taxol. Nature 1979, 277, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Khanna, C.; Rosenberg, M.; Vail, D.M. A Review of Paclitaxel and Novel Formulations Including Those Suitable for Use in Dogs. J. Vet. Intern. Med. 2015, 29, 1006–1012. [Google Scholar] [CrossRef]
- Jeong, I.S.D.; Mo, H.; Nguyen, A.; Chong, E.G.; Tsai, H.H.C.; Moyers, J.; Kim, M.; Lacy, C.; Shah, V.; Lau, E.; et al. Primary chemoradiation with cisplatin versus cetuximab for locally advanced head and neck cancer: A retrospective cohort study. Exp. Hematol. Oncol. 2020, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Oi, H.; Matsuda, T.; Kimura, T.; Morise, M.; Yamano, Y.; Yokoyama, T.; Kataoka, K.; Kondoh, Y. Weekly nanoparticle albumin-bound paclitaxel and paclitaxel for relapsed small cell lung cancer: A retrospective observational study. Medicine 2022, 101, e28863. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Mao, J.-W.; Tan, X.-L. Research progress on the source, production, and anti-cancer mechanisms of paclitaxel. Chin. J. Nat. Med. 2020, 18, 890–897. [Google Scholar] [CrossRef]
- Wheeler, N.C.; Jech, K.; Masters, S.; Brobst, S.W.; Alvarado, A.B.; Hoover, A.J.; Snader, K.M. Effects of Genetic, Epigenetic, and Environmental Factors on Taxol Content in Taxus brevifolia and Related Species. J. Nat. Prod. 1992, 55, 432–440. [Google Scholar] [CrossRef]
- Croteau, R.; Ketchum, R.E.B.; Long, R.M.; Kaspera, R.; Wildung, M.R. Taxol Biosynthesis and Molecular Genetics. Phytochem. Rev. 2006, 5, 75–97. [Google Scholar] [CrossRef] [PubMed]
- Mutanda, I.; Li, J.; Xu, F.; Wang, Y. Recent Advances in Metabolic Engineering, Protein Engineering, and Transcriptome-Guided Insights toward Synthetic Production of Taxol. Front. Bioeng. Biotechnol. 2021, 9, 632269. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Joo, H.-G. Protective effects of fucoidan on the cell death of spleen cells induced by taxol. J. Prev. Vet. Med. 2015, 39, 185–188. (In Chinese) [Google Scholar] [CrossRef]
- Veerabhadrappa, B.; Subramanian, S.; Sudharshan, S.J.; Dyavaiah, M. Evaluating the genetic basiss of anti-cancer property of Taxol in Saccharomyces cerevisiae model. FEMS Microbiol. Lett. 2021, 368, fnab077. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-P.H.; Horwitz, S.B. Taxol®: The First Microtubule Stabilizing Agent. Int. J. Mol. Sci. 2017, 18, 1733. [Google Scholar] [CrossRef]
- Feng, R.; Morine, Y.; Ikemoto, T.; Imura, S.; Iwahashi, S.; Saito, Y.; Shimada, M. Nab-paclitaxel interrupts cancer-stromal interaction through C-X-C motif chemokine 10-mediated interleukin-6 downregulation in vitro. Cancer Sci. 2018, 109, 2509–2519. [Google Scholar] [CrossRef]
- Muraro, E.; Comaro, E.; Talamini, R.; Turchet, E.; Miolo, G.; Scalone, S.; Militello, L.; Lombardi, D.; Spazzapan, S.; Perin, T.; et al. Improved Natural Killer cell activity and retained anti-tumor CD8+ T cell responses contribute to the induction of a pathological complete response in HER2-positive breast cancer patients undergoing neoadjuvant chemotherapy. J. Transl. Med. 2015, 13, 204. [Google Scholar] [CrossRef]
- Wang, S.Q.; Wang, C.; Chang, L.M.; Zhou, K.R.; Wang, J.W.; Ke, Y.; Yang, D.X.; Shi, H.G.; Wang, R.; Shi, X.L.; et al. Geridonin and paclitaxel act synergistically to inhibit the proliferation of gastric cancer cells through ROS-mediated regulation of the PTEN/PI3K/Akt pathway. Oncotarget 2016, 7, 72990–73002. [Google Scholar] [CrossRef]
- Papadopoulos, E.I.; Scorilas, A. Cisplatin and Paclitaxel Alter the Expression Pattern of miR-143/145 and miR-183/96/182 Clusters in T24 Bladder Cancer Cells. Clin. Transl. Sci. 2015, 8, 668–675. [Google Scholar] [CrossRef]
- Xin, Z.; Tong, Z.; Tan, J.; Liu, C. MicroRNA-145-5p aggravates cell apoptosis and oxidative stress in tongue squamous cell carcinoma. Exp. Ther. Med. 2021, 21, 373. [Google Scholar] [CrossRef]
- Lee, Y.; Na, J.; Lee, M.S.; Cha, E.Y.; Sul, J.Y.; Park, J.B.; Lee, J.S. Combination of pristimerin and paclitaxel additively induces autophagy in human breast cancer cells via ERK1/2 regulation. Mol. Med. Rep. 2018, 18, 4281–4288. [Google Scholar] [CrossRef]
- Prencipe, M.; Fitzpatrick, P.; Gorman, S.; Mosetto, M.; Klinger, R.; Furlong, F.; Harrison, M.; O’Connor, D.; Roninson, I.B.; O’Sullivan, J.; et al. Cellular senescence induced by aberrant MAD2 levels impacts on paclitaxel responsiveness in vitro. Br. J. Cancer 2009, 101, 1900–1908. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Ren, J.J.; Liang, C.; Wang, F.; Li, W.; Li, X.F. GSDME influences sensitivity of breast cancer MCF-7 cells to paclitaxel by regulating cell pyroptosis. Chin. J. Cancer Biother 2019, 26, 146–151. (In Chinese) [Google Scholar]
- Weiner-Gorzel, K.; Dempsey, E.; Milewska, M.; McGoldrick, A.; Toh, V.; Walsh, A.; Lindsay, S.; Gubbins, L.; Cannon, A.; Sharpe, D.; et al. Overexpression of the microRNA miR-433 promotes resistance to paclitaxel through the induction of cellular senescence in ovarian cancer cells. Cancer Med. 2015, 4, 745–758. [Google Scholar] [CrossRef]
- You, J.H.; Lee, J.; Roh, J.-L. PGRMC1-dependent lipophagy promotes ferroptosis in paclitaxel-tolerant persister cancer cells. J. Exp. Clin. Cancer Res. 2021, 40, 350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.C.; Li, C.G.; Wang, Y.F.; Xu, L.H.; He, X.H.; Zeng, Q.Z.; Zeng, C.Y.; Mai, F.Y.; Hu, B.; Ouyang, D.Y. Chemotherapeutic paclitaxel and cisplatin differentially induce pyroptosis in A549 lung cancer cells via caspase-3/GSDME activation. Apoptosis 2019, 24, 312–325. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, T.; Liu, R.; Ning, T.; Yang, H.; Liu, D.; Zhang, Q.; Lin, D.; Ge, S.; Bai, M.; et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol. Cancer 2020, 19, 43. [Google Scholar] [CrossRef]
- Liu, W.C.; Gong, T.; Zhu, P. Advances in exploring alternative Taxol sources. RSC Adv. 2016, 6, 48800–48809. [Google Scholar] [CrossRef]
- Ojima, I.; Lichtenthal, B.; Lee, S.; Wang, C.; Wang, X. Taxane anticancer agents: A patent perspective. Expert Opin. Ther. Pat. 2016, 26, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.X.; Yu, L.J.; Bo, S.J. Paclitaxel: A Landmark Natural Anticancer Drug. World Sci. Technol. Mod. Tradit. Chin. Med. Mater. Medica 2017, 19, 941–949. (In Chinese) [Google Scholar]
- Ni, Z.Y.; Li, Y.; Wang, Y.F.; Wang, S.M.; Dong, M.; Shi, Q.W. Cancer-Fighting Molecules-Taxol and its Analogs. Curr. Org. Chem. 2012, 16, 2038–2052. [Google Scholar] [CrossRef]
- Ren, S.; Wang, Y.; Wang, J.; Gao, D.; Zhang, M.; Ding, N.; Li, Y. Synthesis and biological evaluation of novel larotaxel analogues. Eur. J. Med. Chem. 2018, 156, 692–710. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, S.; Wang, C.; Li, Z.; Ma, Y.; Liu, G. Antagonizing NOD2 Signaling with Conjugates of Paclitaxel and Muramyl Dipeptide Derivatives Sensitizes Paclitaxel Therapy and Significantly Prevents Tumor Metastasis. J. Med. Chem. 2017, 60, 1219–1224. [Google Scholar] [CrossRef]
- Ferrandina, G.; Corrado, G.; Vitrano, G.; Gallotta, V.; Palluzzi, E.; Distefano, M.; Scambia, G. Dose-dense paclitaxel/carboplatin as neo-adjuvant chemotherapy followed by radical surgery in locally advanced cervical cancer: A prospective phase II study. Cancer Chemother. Pharmacol. 2019, 83, 431–438. [Google Scholar] [CrossRef]
- Park, S.-A.; Kim, L.K.; Park, H.M.; Kim, H.J.; Heo, T.-H. Inhibition of GP130/STAT3 and EMT by combined bazedoxifene and paclitaxel treatment in ovarian cancer. Oncol. Rep. 2022, 47, 52. [Google Scholar] [CrossRef]
- Yan, X.; Sheng, X.; Chi, Z.; Si, L.; Cui, C.; Kong, Y.; Tang, B.; Mao, L.; Wang, X.; Lian, B.; et al. Randomized Phase II Study of Bevacizumab in Combination with Carboplatin Plus Paclitaxel in Patients with Previously Untreated Advanced Mucosal Melanoma. J. Clin. Oncol. 2021, 39, 881–889. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, B.; Zhou, Z.; Meng, L.; Sun, Z.; Xu, Y.; Xu, Q.; Yuan, A.; Yu, L.; Qian, H.; et al. Enhancement of radiotherapy efficacy by pleiotropic liposomes encapsulated paclitaxel and perfluorotributylamine. Drug Deliv. 2017, 24, 1419–1428. [Google Scholar] [CrossRef] [PubMed]
- Kassem, M.A.R.; Hassan, A.A.; Ibrahim, N.Y.; Toeama, A.A.M. 217P—Concurrent paclitaxel and radiotherapy for node positive breast cancer. Ann. Oncol. 2016, 27, vi65. [Google Scholar] [CrossRef]
- Hussain, T.; Paranthaman, S.; Rizvi, S.M.; Moin, A.; Gowda, D.V.; Subaiea, G.M.; Ansari, M.; Alanazi, A.S. Fabrication and Characterization of Paclitaxel and Resveratrol Loaded Soluplus Polymeric Nanoparticles for Improved BBB Penetration for Glioma Management. Polymers 2021, 13, 3210. [Google Scholar] [CrossRef]
- Jiang, Q.; Yang, M.; Qu, Z.; Zhou, J.; Zhang, Q. Resveratrol enhances anticancer effects of paclitaxel in HepG2 human liver cancer cells. BMC Complement. Altern. Med. 2017, 17, 477–488. [Google Scholar] [CrossRef]
- Calaf, G.M.; Ponce-Cusi, R.; Carrión, F. Curcumin and paclitaxel induce cell death in breast cancer cell lines. Oncol. Rep. 2018, 40, 2381–2388. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Pu, X.; Zhao, L. Preclinical studies for the combination of paclitaxel and curcumin in cancer therapy (Review). Oncol. Rep. 2017, 37, 3159–3166. [Google Scholar] [CrossRef] [PubMed]
- Fratantonio, D.; Molonia, M.S.; Bashllari, R.; Muscarà, C.; Ferlazzo, G.; Costa, G.; Saija, A.; Cimino, F.; Speciale, A. Curcumin potentiates the antitumor activity of Paclitaxel in rat glioma C6 cells. Phytomedicine 2019, 55, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shen, Z.; Zhu, T.; Lu, W.; Fu, Y. Curcumin enhances the anti-cancer efficacy of paclitaxel in ovarian cancer by regulating the miR-9-5p/BRCA1 axis. Front. Pharmacol. 2022, 13, 1014933. [Google Scholar] [CrossRef]
- Boztas, A.O.; Karakuzu, O.; Galante, G.; Ugur, Z.; Kocabas, F.; Altuntas, C.Z.; Yazaydin, A.O. Synergistic interaction of paclitaxel and curcumin with cyclodextrin polymer complexation in human cancer cells. Mol. Pharm. 2013, 10, 2676–2683. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, F.; Kikuchi, Y.; Ohira, T.; Yatagai, M. Accelerated Solvent Extraction of Paclitaxel and Related Compounds from the Bark of Taxus cuspidata. J. Nat. Prod. 1999, 62, 244–247. [Google Scholar] [CrossRef]
- McPartland, T.J.; Patil, R.A.; Malone, M.F.; Roberts, S.C. Liquid–liquid extraction for recovery of paclitaxel from plant cell culture: Solvent evaluation and use of extractants for partitioning and selectivity. Biotechnol. Prog. 2012, 28, 990–997. [Google Scholar] [CrossRef]
- Tan, Z.; Li, Q.; Wang, C.; Zhou, W.; Yang, Y.; Wang, H.; Yi, Y.; Li, F. Ultrasonic Assisted Extraction of Paclitaxel from Taxus × media Using Ionic Liquids as Adjuvants: Optimization of the Process by Response Surface Methodology. Molecules 2017, 22, 1483. [Google Scholar] [CrossRef]
- Min, H.-S.; Kim, J.-H. Process synthesis and optimization for the isolation and purification of paclitaxel from Taxus chinensis. Korean J. Chem. Eng. 2022, 39, 3389–3398. [Google Scholar] [CrossRef]
- Shin, E.-J.; Kim, J.-H. Ultrasound- and Negative Pressure-assisted Fractional Precipitation of Paclitaxel from Taxus chinensis. Biotechnol. Bioproc. Eng. 2023, 28, 336–344. [Google Scholar] [CrossRef]
- Jeon, H.-J.; Kim, J.-H. Effect of Gas Bubbles on the Recovery Efficiency of Paclitaxel from Biomass of Taxus chinensis in Ultrasonic Extraction. Biotechnol. Bioproc. Eng. 2023, 28, 545–553. [Google Scholar] [CrossRef]
- Min, H.-S.; Kim, H.-G.; Kim, J.-H. Ultrasound-negative pressure cavitation extraction of paclitaxel from Taxus chinensis. Korean J. Chem. Eng. 2022, 39, 398–407. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Nantermet, P.G.; Ueno, H.; Guy, R.K.; Couladouros, E.A.; Sorensen, E.J. Total Synthesis of Taxol. 1. Retrosynthesis, Degradation, and Reconstitution. J. Am. Chem. Soc. 1995, 117, 624–633. [Google Scholar] [CrossRef]
- Holton, R.A.; Somoza, C.; Kim, H.B.; Liang, F.; Biediger, R.J.; Boatman, P.D.; Shindo, M.; Smith, C.C.; Kim, S. First total synthesis of taxol. 1. Functionalization of the B ring. J. Am. Chem. Soc. 1994, 116, 1597–1598. [Google Scholar] [CrossRef]
- Holton, R.A.; Kim, H.B.; Somoza, C.; Liang, F.; Biediger, R.J.; Boatman, P.D.; Shindo, M.; Smith, C.C.; Kim, S. First total synthesis of taxol. 2. Completion of the C and D rings. J. Am. Chem. Soc. 1994, 116, 1599–1600. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, J.; Li, W.-D.Z. Diverse strategic approaches en route to Taxol total synthesis. Chin. Chem. Lett. 2022, 33, 4957–4968. [Google Scholar] [CrossRef]
- Min, L.; Han, J.-C.; Zhang, W.; Gu, C.-C.; Zou, Y.-P.; Li, C.-C. Strategies and Lessons Learned from Total Synthesis of Taxol. Chem. Rev. 2023, 123, 4934–4971. [Google Scholar] [CrossRef] [PubMed]
- Danishefsky, S.J.; Masters, J.J.; Young, W.B.; Link, J.T.; Snyder, L.B.; Magee, T.V.; Jung, D.K.; Isaacs, R.C.A.; Bornmann, W.G.; Alaimo, C.A.; et al. Total Synthesis of Baccatin III and Taxol. J. Am. Chem. Soc. 1996, 118, 2843–2859. [Google Scholar] [CrossRef]
- Wender, P.A.; Badham, N.F.; Conway, S.P.; Floreancig, P.E.; Glass, T.E.; Gränicher, C.; Houze, J.B.; Jänichen, J.; Lee, D.; Marquess, D.G.; et al. The Pinene Path to Taxanes. 5. Stereocontrolled Synthesis of a Versatile Taxane Precursor. J. Am. Chem. Soc. 1997, 119, 2755–2756. [Google Scholar] [CrossRef]
- Wender, P.A.; Badham, N.F.; Conway, S.P.; Floreancig, P.E.; Glass, T.E.; Houze, J.B.; Krauss, N.E.; Lee, D.; Marquess, D.G.; McGrane, P.L.; et al. The Pinene Path to Taxanes. 6. A Concise Stereocontrolled Synthesis of Taxol. J. Am. Chem. Soc. 1997, 119, 2757–2758. [Google Scholar] [CrossRef]
- Kusama, H.; Hara, R.; Kawahara, S.; Nishimori, T.; Kashima, H.; Nakamura, N.; Morihira, K.; Kuwajima, I. Enantioselective Total Synthesis of (−)-Taxol. J. Am. Chem. Soc. 2000, 122, 3811–3820. [Google Scholar] [CrossRef]
- Mukaiyama, T.; Shiina, I.; Iwadare, H.; Saitoh, M.; Nishimura, T.; Ohkawa, N.; Sakoh, H.; Nishimura, K.; Tani, Y.-i.; Hasegawa, M.; et al. Asymmetric Total Synthesis of Taxol\R. Chem. Eur. J. 1999, 5, 121–161. [Google Scholar] [CrossRef]
- Doi, T.; Fuse, S.; Miyamoto, S.; Nakai, K.; Sasuga, D.; Takahashi, T. A Formal Total Synthesis of Taxol Aided by an Automated Synthesizer. Chem. Asian J. 2006, 1, 370–383. [Google Scholar] [CrossRef]
- Hirai, S.; Utsugi, M.; Iwamoto, M.; Nakada, M. Formal Total Synthesis of (−)-Taxol through Pd-Catalyzed Eight-Membered Carbocyclic Ring Formation. Chem. Eur. J. 2015, 21, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Fukaya, K.; Tanaka, Y.; Sato, A.C.; Kodama, K.; Yamazaki, H.; Ishimoto, T.; Nozaki, Y.; Iwaki, Y.M.; Yuki, Y.; Umei, K.; et al. Synthesis of Paclitaxel. 1. Synthesis of the ABC Ring of Paclitaxel by SmI2-Mediated Cyclization. Org. Lett. 2015, 17, 2570–2573. [Google Scholar] [CrossRef]
- Fukaya, K.; Kodama, K.; Tanaka, Y.; Yamazaki, H.; Sugai, T.; Yamaguchi, Y.; Watanabe, A.; Oishi, T.; Sato, T.; Chida, N. Synthesis of Paclitaxel. 2. Construction of the ABCD Ring and Formal Synthesis. Org. Lett. 2015, 17, 2574–2577. [Google Scholar] [CrossRef]
- Kanda, Y.; Nakamura, H.; Umemiya, S.; Puthukanoori, R.K.; Murthy Appala, V.R.; Gaddamanugu, G.K.; Paraselli, B.R.; Baran, P.S. Two-Phase Synthesis of Taxol. J. Am. Chem. Soc. 2020, 142, 10526–10533. [Google Scholar] [CrossRef]
- Hu, Y.J.; Gu, C.C.; Wang, X.F.; Min, L.; Li, C.C. Asymmetric Total Synthesis of Taxol. J. Am. Chem. Soc. 2021, 143, 17862–17870. [Google Scholar] [CrossRef]
- Iiyama, S.; Fukaya, K.; Yamaguchi, Y.; Watanabe, A.; Yamamoto, H.; Mochizuki, S.; Saio, R.; Noguchi, T.; Oishi, T.; Sato, T.; et al. Total Synthesis of Paclitaxel. Org. Lett. 2022, 24, 202–206. [Google Scholar] [CrossRef]
- Imamura, Y.; Takaoka, K.; Komori, Y.; Nagatomo, M.; Inoue, M. Total Synthesis of Taxol Enabled by Inter- and Intramolecular Radical Coupling Reactions. Angew. Chem. Int. Ed. 2023, 62, e202219114. [Google Scholar] [CrossRef]
- Denis, J.N.; Greene, A.E.; Guenard, D.; Gueritte-Voegelein, F.; Mangatal, L.; Potier, P. Highly efficient, practical approach to natural taxol. J. Am. Chem. Soc. 1988, 110, 5917–5919. [Google Scholar] [CrossRef]
- Deng, X.M.; Cao, H.; Shen, H.W.; Zhong, G.X.; Xia, C.N. Overview of the semisynthesis of paclitaxel and its derivatives. Zhejiang Chem. 2018, 49, 1–8. (In Chinese) [Google Scholar]
- Yan, J.-Q. Chiral synthesis of the Taxol side chain and semi synthesis of Taxol. Fine Spec. Chem. 2005, 13, 1–6+11. (In Chinese) [Google Scholar]
- Borah, C.J.; Boruwa, J.; Barua, C.N. Synthesis of the C-13 Side-Chain of Taxol. Curr. Org. Synth. 2007, 4, 175–199. [Google Scholar] [CrossRef]
- Shi, T.; Li, X.; Li, Y.; Fu, X.; Wu, L.; Wu, D.; Hu, W. An asymmetric catalytic multi-component reaction enabled the green synthesis of isoserine derivatives and semi-synthesis of paclitaxel. Green Synth. Catal. 2023, 4, 58–63. [Google Scholar] [CrossRef]
- Zhang, H.; Gan, H.; Wu, Z.; Meng, Z.; Dou, G. Study on biotransformation of 7-xylosyl-10deacetyltaxol. Chin. J. New Drugs 2013, 22, 1029–1033. (In Chinese) [Google Scholar]
- Xue, B.; Zhao, J.; Fan, Y.; Chen, S.; Li, W.; Chen, J.; Li, Z.; Wang, H.; Kong, H. Synthesis of Taxol and Docetaxel by Using 10-Deacetyl-7-xylosyltaxanes. Chem. Biodivers. 2020, 17, e1900631. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.V. Semi-synthesis of paclitaxel from naturally occurring glycosidic precursors. J. Heterocycl. Chem. 1997, 34, 675–680. [Google Scholar] [CrossRef]
- Christen, A.A.; Gibson, D.M.; Bland, J. Production of Taxol or Taxol-Like Compounds in Cell Culture. U.S. Patent 5,019,504, 28 May 1991. [Google Scholar]
- Bamneshin, M.; Mirjalili, M.H.; Naghavi, M.R.; Cusido, R.M.; Palazón, J. Gene expression pattern and taxane biosynthesis in a cell suspension culture of Taxus baccata L. subjected to light and a phenylalanine ammonia lyase (PAL) inhibitor. J. Photochem. Photobiol. B Biol. 2022, 234, 112532. [Google Scholar] [CrossRef] [PubMed]
- Golinejad, S.; Mirjalili, M.H.; Rezadoost, H.; Ghorbanpour, M. Molecular, biochemical, and metabolic changes induced by gold nanoparticles in Taxus baccata L. cell culture. Ind. Crops Prod. 2023, 192, 115988. [Google Scholar] [CrossRef]
- Hazrati, R.; Zare, N.; Asghari, R.; Sheikhzadeh, P.; Johari-Ahar, M. Biologically synthesized CuO nanoparticles induce physiological, metabolic, and molecular changes in the hazel cell cultures. Appl. Microbiol. Biotechnol. 2022, 106, 6017–6031. [Google Scholar] [CrossRef] [PubMed]
- Yukimune, Y.; Tabata, H.; Higashi, Y.; Hara, Y. Methyl jasmonate-induced overproduction of paclitaxel and baccatin III in Taxus cell suspension cultures. Nat. Biotechnol. 1996, 14, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, H.; Li, T.; Li, C.; Zhou, Y.; Zhong, X. The selection and stability analysis of stable and high Taxol-producing cell lines from Taxus cuspidata. J. For. Res. 2018, 29, 65–71. [Google Scholar] [CrossRef]
- Hoffman, A.; Khan, W.; Worapong, J.; Strobel, G.; Griffin, D.; Arbogast, B.; Barofsky, B.; Boone, D.; Ning, R.B.; Zheng, P.; et al. Bioprospecting for taxol in angiosperm plant extracts: Using high performance liquid chromatography–thermospray mass spectrometry to detect the anticancer agent and its related metabolites in filbert trees. Spectroscopy 1998, 13, 22–32. [Google Scholar]
- Gallego, A.; Imseng, N.; Bonfill, M.; Cusido, R.M.; Palazon, J.; Eibl, R.; Moyano, E. Development of a hazel cell culture-based paclitaxel and baccatin III production process on a benchtop scale. J. Biotechnol. 2015, 195, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, S.; Moieni, A.; Safaie, N.; Sabet, M.S.; Salehi, M. Fungal Cell Wall and Methyl-β–Cyclodextrin Synergistically Enhance Paclitaxel Biosynthesis and Secretion in Corylus avellana Cell Suspension Culture. Sci. Rep. 2020, 10, 5427. [Google Scholar] [CrossRef]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; El-Sayed, M.T.; Rady, A.; Zein, N.; Enan, G.; Shindia, A.; El-Hefnawy, S.; Sitohy, M.; Sitohy, B. Exploiting the Biosynthetic Potency of Taxol from Fungal Endophytes of Conifers Plants; Genome Mining and Metabolic Manipulation. Molecules 2020, 25, 3000. [Google Scholar] [CrossRef]
- Subban, K.; Kempken, F. Insights into Taxol® biosynthesis by endophytic fungi. Appl. Microbiol. Biotechnol. 2023, 107, 6151–6162. [Google Scholar] [CrossRef]
- Shankar Naik, B. Developments in taxol production through endophytic fungal biotechnology: A review. Orient. Pharm. Exp. Med. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Strobel, G.; Yang, X.; Sears, J.; Kramer, R.; Sidhu, R.S.; Hess, W.M. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 1996, 142, 435–440. [Google Scholar] [CrossRef]
- Tian, R.P.; Yang, Q.; Zhou, G.L. Taxonomic study on a taxol producing fungus isolated from the bark of Taxus chinensis var. mairei. J. Wuhan Bot. Res. 2006, 24, 541–545. [Google Scholar]
- Strobel, G.A.; Hess, W.M.; Ford, E.; Sidhu, R.S.; Yang, X. Taxol from fungal endophytes and the issue of biodiversity. J. Ind. Microbiol. 1996, 17, 417–423. [Google Scholar] [CrossRef]
- Fu, Y.; Li, X.; Yuan, X.; Zhang, Z.; Wei, W.; Xu, C.; Song, J.; Gu, C. Alternaria alternata F3, a Novel Taxol-Producing Endophytic Fungus Isolated from the Fruits of Taxus cuspidata: Isolation, Characterization, Taxol Yield Improvement, and Antitumor Activity. Appl. Biochem. Biotechnol. 2023, 195. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Zhang, Z.; Wang, Y.; Su, Y.; Zhang, R. Screening endophytic fungus to produce taxol from Taxus Yunnanensis. Biotechnol. 2003, 13, 10–11. (In Chinese) [Google Scholar]
- Zhang, P.; Zhou, P.-P.; Yu, L.-J. An Endophytic Taxol-Producing Fungus from Taxus media, Cladosporium cladosporioides MD2. Curr. Microbiol. 2009, 59, 227–232. [Google Scholar] [CrossRef]
- Zhang, P.; Zhou, P.-P.; Yu, L.-J. An endophytic taxol-producing fungus from Taxus × media, Aspergillus candidus MD3. FEBS Microbiol. Lett. 2009, 293, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Singh, B.; Thakur, V.; Thakur, A.; Thakur, N.; Pandey, D.; Chand, D. Hyper-production of taxol from Aspergillus fumigatus, an endophytic fungus isolated from Taxus sp. of the Northern Himalayan region. Biotechnol. Rep. 2019, 24, e00395. [Google Scholar] [CrossRef]
- Zhao, K.; Ping, W.X.; Li, Q. Aspergillus niger var. taxi, a new species variant of taxol-producing fungus isolated from Taxus cuspidata in China. J. Appl. Microbiol. 2009, 107, 1202–1207. [Google Scholar] [CrossRef]
- Chen, J.H.; Liu, J.J.; Zang, G.G. Screening of taxol-producing endophytic fungi and regulation of fermentation conditions. J. Cent. South Univ. Nat. Sci. 2004, 35, 65–69. [Google Scholar]
- Zhou, X.; Zhu, H.; Liu, L.; Lin, J.; Tang, K. A review: Recent advances and future prospects of taxol-producing endophytic fungi. Appl. Microbiol. Biotechnol. 2010, 86, 1707–1717. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Hu, C.H. Separation and identification of a new Taxus chinensis var. mairei endophytic fungus (Bionectria sp.) and the activity of its metabolites. J. Southwest Univ. Nat. Sci. Ed. 2007, 29, 131–135. [Google Scholar]
- Venkatachalam, R.; Subban, K.; Paul, M.J. Taxol from Botryodiplodia theobromae (BT 115)-an endophytic fungus of Taxus baccata. J. Biotechnol. 2008, 136, 189–190. [Google Scholar] [CrossRef]
- Zhao, K.; Zhao, L.; Jin, Y.; Wei, H.; Ping, W.; Zhou, D. Isolation of a taxol-producing endophytic fungus and inhibiting effect of the fungus metabolites on HeLa cell. Mycosystema 2008, 27, 735–744. (In Chinese) [Google Scholar]
- Hu, K.; Tan, F.; Tang, K.; Zhu, S.; Wang, W. Isolation and screening of endophytic fungi synthesizing taxol from Taxus chinensis var. mairei. J. Southwest China Normal Univ. Nat. Sci. Ed. 2006, 31, 134–137. [Google Scholar]
- Wang, Y.; Ma, X.Y.; Ping, W.; Zhou, D. A new species of Botrytis. J. Fungal. Res. 2006, 4, 62–64. (In Chinese) [Google Scholar]
- Wang, Y.; Tang, K. A new endophytic taxol- and baccatin III-producing fungus isolated from Taxus chinensis var. mairei. Afr. J. Biotechnol. 2011, 10, 16379–16386. [Google Scholar]
- Li, C.T.; Li, Y.; Wang, Q.; Sung, C. Taxol production by Fusarium Arthrosporioides isolated from yew, Taxus cuspidata. J. Med. Biochem. 2008, 27, 454–458. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Ali, D.M.I.; Yassin, M.A.; Zayed, R.A.; Ali, G.S. Sterol inhibitor “Fluconazole” enhance the Taxol yield and molecular expression of its encoding genes cluster from Aspergillus flavipes. Process Biochem. 2019, 76, 55–67. [Google Scholar] [CrossRef]
- Dai, W.L.; Tao, W.Y. Preliminary study on fermentation conditions of taxol-producing endophytic fungus. Chem. Ind. Eng. Prog. 2008, 27, 883–886. (In Chinese) [Google Scholar]
- Garyali, S.; Kumar, A.; Reddy, M.S. Taxol production by an endophytic fungus Fusarium redolens, isolated from Himalayan Yew. J. Microbiol. Biotechnol. 2013, 23, 1372–1380. [Google Scholar] [CrossRef]
- Deng, B.W.; Liu, K.H.; Chen, W.Q.; Ding, X.W.; Xie, X.C. Fusarium solani, Tax-3, a new endophytic taxol-producing fungus from Taxus chinensis. World J. Microbiol. Biotechnol. 2009, 25, 139–143. [Google Scholar] [CrossRef]
- Lu, L.Y.; Qin, Z.; Xu, J.K.; Li, X.S. Production of taxol by an endophytic fungus isolated from Taxus chinensis var. Mairei. Chin. Med. Biotechnol. 2010, 5, 202–207. (In Chinese) [Google Scholar]
- Cheng, L.; Ma, Q.M.; Tao, G.J.; Tao, W.Y.; Wang, R.M.; Yang, J.; Guo, X.L. Systematic identification of a paclitaxel-producing endofungus. Ind. Microbiol. 2007, 37, 23–30. (In Chinese) [Google Scholar]
- Xiong, Z.Q.; Yang, Y.Y.; Zhao, N. Diversity of endophytic fungi and screening of fungal paclitaxel producer from Anglo jap yew, Taxus × media. BMC Microbiol. 2013, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.Y.; Zhang, P.; Liu, B.; Xu, M.; Zhou, P.P.; Yu, L.J. Isolation and identification of a taxol-producing endophytic fungus Z58. Chin. J. Biochem. Mol. Biol. 2012, 28, 1141–1146. (In Chinese) [Google Scholar]
- Liu, W.H.; Yao, B.; Zhu, S.Q. Advances in studies on biosynthetic pathway of taxol precursor and its correlative biotechnology. Chin. Tradit. Herb Drugs 2009, 40, 1327–1331. [Google Scholar]
- Miao, Z.; Wang, Y.; Yu, X.; Guo, B.; Tang, K. New endophytic taxane production fungus from T. chinensis. Appl. Biochem. Microbiol. 2009, 45, 81–86. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, B.; Xu, M.; Ao, M.Z.; Fu, C.H.; Yu, L.J.; Gu, Y.C. Isolation and identification of a taxol-producing endophytic fungus O60B1. Hubei Agric. Sci. 2012, 51, 5315–5317. (In Chinese) [Google Scholar]
- Zhou, D.; Zhao, K.; Ping, W. Study on the mutagenesis of protoplasts from taxol-producing fungus Nodulisporium sylviforme. J. Am. Sci. 2005, 1, 55–62. [Google Scholar]
- Zhao, K.; Sun, L.X.; Wang, X. Screening of high taxol producing fungi by mutagenesis and construction of subtracted cDNA library by Suppression substracted hybridization for differentially expressed genes. Acta. Microbiol. Sin. 2011, 51, 923–933. [Google Scholar]
- Guo, B.H.; Wang, Y.C.; Zhou, X.W. An endophytic taxol-producing fungus BT2 isolated from Taxus chinensis var. mairei. Afr. J. Biotechnol. 2006, 5, 875–877. [Google Scholar]
- Zhang, P.; Liu, B.; Zhou, B.B.; Wang, C.L.; Jiang, C.; Yu, L.J. Isolation and identification of a taxol-producing endophytic fungus YN6. Chin. J. Biochem. Mol. Biol. 2011, 27, 961–967. (In Chinese) [Google Scholar]
- Gangadevi, V.; Muthumary, J. Taxol production by Pestalotiopsis terminaliae, an endophytic fungus of Terminalia arjuna (arjun tree). Biotechnol. Appl. Biochem. 2009, 52, 9–15. [Google Scholar] [CrossRef]
- Kumaran, R.S.; Hur, B.K. Screening of species of the endophytic fungus Phomopsis for the production of the anticancer drug taxol. Biotechnol. Appl. Biochem. 2009, 54, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Li, T.Q.; Zhang, Z.J.; Zhang, P.; Wang, C.L.; Liu, B.; Liu, T.T.; Fu, C.H.; Yu, L.J. Isolation and identification of a taxol-producing endophytic fungus identified from Taxus media. Agric. Sci. Technol. 2010, 11, 38–40+68. (In Chinese) [Google Scholar]
- Mirjalili, M.H.; Farzaneh, M.; Bonfill, M.; Rezadoost, H.; Ghassempour, A. Isolation and characterization of Stemphylium sedicola SBU-16 as a new endophytic taxol-producing fungus from Taxus baccata grown in Iran. FEMS Microbiol. Lett. 2012, 328, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Staniek, A.; Woerdenbag, H.J.; Kayser, O. Taxomyces andreanae: A presumed paclitaxel producer demystified. Planta. Med. 2009, 75, 1561–1566. [Google Scholar] [CrossRef]
- Wang, J.F.; Li, G.L.; Lu, H.Y. Taxol from Tubercularia sp. Strain TF5, an endophytic fungus of Taxus mairei. FEMS Microbiol. Lett. 2000, 193, 249–253. [Google Scholar] [CrossRef]
- Bi, J.; Ji, Y.; Pan, J.; Yu, Y.; Chen, H.; Zhu, X. A new taxol-producing fungus (Pestalotiopsis malicola) and evidence for taxol as a transient product in the culture. Afr. J. Biotechnol. 2011, 10, 6647–6654. [Google Scholar]
- Strobel, G.A.; Hess, W.M.; Li, J.Y.; Ford, E.; Sears, J.; Sidhu, R.S.; Summerell, B. Pestalotiopsis guepinii, a taxol-producing endophyte of the Wollemi pine, Wollemia nobilis. Aust. J. Bot. 1997, 45, 1073–1082. [Google Scholar] [CrossRef]
- Li, J.Y.; Strobel, G.; Sidhu, R. Endophytic taxol-producing fungi from bald cypress, Taxodium distichum. Microbiology 1996, 142, 2223–2236. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, H.; Barrero, R.A.; Zhang, B.; Sun, G.; Wilson, I.W.; Xie, F.; Walker, K.D.; Parks, J.W.; Bruce, R. Genome sequencing and analysis of the paclitaxel-producing endophytic fungus Penicillium aurantiogriseum NRRL 62431. BMC Genom. 2014, 15, 69. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, R.S.; Choi, Y.K.; Lee, S.; Jeon, H.J.; Jung, H.; Kim, H.J. Isolation of taxol, an anticancer drug produced by the endophytic fungus, Phoma betae. Afr. J. Biotechnol. 2012, 11, 950–960. [Google Scholar]
- Kumaran, R.S.; Muthumary, J.; Hur, B.K. Production of taxol from Phyllosticta spinarum, an endophytic fungus of Cupressus sp. Eng. Life Sci. 2008, 8, 438–446. [Google Scholar] [CrossRef]
- Sun, D.F.; Ran, X.Q.; Wang, J.F. Isolation and identifcation of a taxol-producing endophytic fungus from Podocarpus. Acta. Microbiol. Sin. 2008, 48, 589–595. [Google Scholar]
- Gangadevi, V.; Murugan, M.; Muthumary, J. Taxol determination from Pestalotiopsis pauciseta, a fungal endophyte of a medicinal plant. Chin. J. Biotechnol. 2008, 24, 1433–1438. [Google Scholar] [CrossRef]
- Gangadevi, V.; Muthumary, J. A novel endophytic taxolproducing fungus Chaetomella raphigera isolated from a medicinal plant, Terminalia arjuna. Appl. Biochem. Biotechnol. 2009, 158, 675–684. [Google Scholar] [CrossRef]
- Kumaran, R.S.; Muthumary, J.; Hur, B.K. Isolation and identifcation of an anticancer drug, taxol from Phyllosticta tabernaemontanae, a leaf spot fungus of an angiosperm Wrightia tinctoria. J. Microbiol. 2009, 47, 40–49. [Google Scholar] [CrossRef]
- Gokul Raj, K.; Manikandan, R.; Arulvasu, C.; Pandi, M. Anti-proliferative effect of fungal taxol extracted from Cladosporium oxysporum against human pathogenic bacteria and human colon cancer cell line HCT 15. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 138, 667–674. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, Y.; Ma, X.; Wang, C.; Yue, G.; Zhang, Y.; Zhang, Y.; Li, S.; Ling, S.; Liu, X.; et al. Greater Taxol Yield of Fungus Pestalotiopsis hainanensis from Dermatitic Scurf of the Giant Panda (Ailuropoda melanoleuca). Appl. Biochem. Biotechnol. 2015, 175, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Garyali, S.; Kumar, A.; Reddy, M.S. Enhancement of taxol production from endophytic fungus Fusarium redolens. Biotechnol. Bioproc. E. 2014, 19, 908–915. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Mosa, K.A.; El-Keblawy, A.A.; Husseiny, M.I. Exogenous and endogenous increase in fungal GGPP increased fungal Taxol production. Appl. Microbiol. Biotechnol. 2017, 101, 7523–7533. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Ling, F.; Yu, L.; Huang, Y.; Wang, T. Enhancing taxol production in a novel endophytic fungus, Aspergillus aculeatinus Tax-6, isolated from Taxus chinensis var. mairei. Fungal Biol. 2017, 121, 1037–1044. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Shindia, A.A.; AbouZeid, A.; Koura, A.; Hassanein, S.E.; Ahmed, R.M. Triggering the biosynthetic machinery of Taxol by Aspergillus flavipes via cocultivation with Bacillus subtilis: Proteomic analyses emphasize the chromatin remodeling upon fungal-bacterial interaction. Environ. Sci. Pollut. Res. 2021, 28, 39866–39881. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Fathalla, M.; Yassin, M.A.; Zein, N.; Morsy, S.; Sitohy, M.; Sitohy, B. Conjugation of Aspergillus flavipes Taxol with Porphyrin Increases the Anticancer Activity of Taxol and Ameliorates Its Cytotoxic Effects. Molecules 2020, 25, 263. [Google Scholar] [CrossRef]
- Subban, K.; Subramani, R.; Srinivasan, V.P.M.; Johnpaul, M.; Chelliah, J. Salicylic acid as an effective elicitor for improved taxol production in endophytic fungus Pestalotiopsis microspora. PLoS ONE 2019, 14, e0212736. [Google Scholar] [CrossRef]
- El-Sayed, A.; Enan, G.; Al-Mohammadi, A.-R.; Moustafa, A.H.; El-Gazzar, N. Detection, Purification and Elucidation of Chemical Structure and Antiproliferative Activity of Taxol Produced by Penicillium chrysogenum. Molecules 2020, 25, 4822. [Google Scholar] [CrossRef]
- Yang, N.; Pan, X.W.; Chen, G.J.; Sarsaiya, S.; Yu, J.; Fan, X.K.; Jin, L.L.; Zhang, B.H.; Shi, J.; Chen, J.S. Fermentation Engineering for Enhanced Paclitaxel Production by Taxus Media Endophytic Fungus MF-5 (Alternaria sp.). J. Biobased Mater. Biol. 2018, 12, 545–550. [Google Scholar] [CrossRef]
- Abdel-Fatah, S.S.; El-Sherbiny, G.M.; Khalaf, M.; Baz, A.F.E.; El-Sayed, A.S.A.; El-Batal, A.I. Boosting the Anticancer Activity of Aspergillus flavus “endophyte of Jojoba” Taxol via Conjugation with Gold Nanoparticles Mediated by γ-Irradiation. Appl. Biochem. Biotechnol. 2022, 194, 3558–3581. [Google Scholar] [CrossRef]
- Chakravarthi, B.V.S.K.; Singh, S.; Kamalraj, S.; Gupta, V.K.; Jayabaskaran, C. Evaluation of spore inoculum and confirmation of pathway genetic blueprint of T13αH and DBAT from a Taxol-producing endophytic fungus. Sci. Rep. 2020, 10, 21139. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.S.; Greenwood, J.S.; Bombarely, A.; Mueller, L.A.; Tsao, R.; Mosser, D.D.; Raizada, M.N. An Endophyte Constructs Fungicide-Containing Extracellular Barriers for Its Host Plant. Curr. Biol. 2015, 25, 2570–2576. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.S.M.; Raizada, M.N. Darkness: A Crucial Factor in Fungal Taxol Production. Front. Microbiol. 2018, 9, 353. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, L.; Zhuang, W.; Zhang, F.; Shu, X.; Wang, N.; Wang, Z. Recent Research Progress in Taxol Biosynthetic Pathway and Acylation Reactions Mediated by Taxus Acyltransferases. Molecules 2021, 26, 2855. [Google Scholar] [CrossRef]
- Muchiri, R.; Walker, K.D. Taxol Biosynthesis: Tyrocidine Synthetase A Catalyzes the Production of Phenylisoserinyl CoA and Other Amino Phenylpropanoyl Thioesters. Cell Chem. Biol. 2012, 19, 679–685. [Google Scholar] [CrossRef][Green Version]
- Walker, K.; Fujisaki, S.; Long, R.; Croteau, R. Molecular cloning and heterologous expression of the C-13 phenylpropanoid side chain-CoA acyltransferase that functions in Taxol biosynthesis. Proc. Natl. Acad. Sci. USA 2002, 99, 12715–12720. [Google Scholar] [CrossRef]
- Ajikumar, P.K.; Xiao, W.-H.; Tyo, K.E.J.; Wang, Y.; Simeon, F.; Leonard, E.; Mucha, O.; Phon, T.H.; Pfeifer, B.; Stephanopoulos, G. Isoprenoid Pathway Optimization for Taxol Precursor Overproduction in Escherichia coli. Science 2010, 330, 70–74. [Google Scholar] [CrossRef]
- Biggs, B.W.; Lim, C.G.; Sagliani, K.; Shankar, S.; Stephanopoulos, G.; De Mey, M.; Ajikumar, P.K. Overcoming heterologous protein interdependency to optimize P450-mediated Taxol precursor synthesis in Escherichia coli. Proc. Natl. Acad. Sci. USA 2016, 113, 3209–3214. [Google Scholar] [CrossRef]
- Huang, Q.; Roessner, C.A.; Croteau, R.; Scott, A.I. Engineering Escherichia coli for the synthesis of taxadiene, a key intermediate in the biosynthesis of taxol. Bioorg. Med. Chem. 2001, 9, 2237–2242. [Google Scholar] [CrossRef]
- Zhou, K.; Qiao, K.; Edgar, S.; Stephanopoulos, G. Distributing a metabolic pathway among a microbial consortium enhances production of natural products. Nat. Biotechnol. 2015, 33, 377–383. [Google Scholar] [CrossRef]
- Abdallah, I.I.; Pramastya, H.; van Merkerk, R.; Sukrasno; Quax, W.J. Metabolic Engineering of Bacillus subtilis toward Taxadiene Biosynthesis as the First Committed Step for Taxol Production. Front. Microbiol. 2019, 10, 218. [Google Scholar] [CrossRef]
- Dejong, J.M.; Liu, Y.; Bollon, A.P.; Long, R.M.; Jennewein, S.; Williams, D.; Croteau, R.B. Genetic engineering of taxol biosynthetic genes in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2006, 93, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Engels, B.; Dahm, P.; Jennewein, S. Metabolic engineering of taxadiene biosynthesis in yeast as a first step towards Taxol (Paclitaxel) production. Metab. Eng. 2008, 10, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.-z.; Yan, H.-f.; Li, L.-f.; Zhai, F.; Shang, L.-q.; Yin, Z.; Yuan, Y.-j. Biosynthesis of Taxadiene in Saccharomyces cerevisiae: Selection of Geranylgeranyl Diphosphate Synthase Directed by a Computer-Aided Docking Strategy. PLoS ONE 2014, 9, e109348. [Google Scholar] [CrossRef] [PubMed]
- Reider Apel, A.; d’Espaux, L.; Wehrs, M.; Sachs, D.; Li, R.A.; Tong, G.J.; Garber, M.; Nnadi, O.; Zhuang, W.; Hillson, N.J.; et al. A Cas9-based toolkit to program gene expression in Saccharomyces cerevisiae. Nucleic Acids Res. 2017, 45, 496–508. [Google Scholar] [CrossRef]
- Nowrouzi, B.; Li, R.A.; Walls, L.E.; d’Espaux, L.; Malcı, K.; Liang, L.; Jonguitud-Borrego, N.; Lerma-Escalera, A.I.; Morones-Ramirez, J.R.; Keasling, J.D.; et al. Enhanced production of taxadiene in Saccharomyces cerevisiae. Microb. Cell Fact. 2020, 19, 200. [Google Scholar] [CrossRef]
- Walls, L.E.; Malcı, K.; Nowrouzi, B.; Li, R.A.; d’Espaux, L.; Wong, J.; Dennis, J.A.; Semião, A.J.C.; Wallace, S.; Martinez, J.L.; et al. Optimizing the biosynthesis of oxygenated and acetylated Taxol precursors in Saccharomyces cerevisiae using advanced bioprocessing strategies. Biotechnol. Bioeng. 2021, 118, 279–293. [Google Scholar] [CrossRef]
- Walls, L.E.; Martinez, J.L.; Rios-Solis, L. Enhancing Saccharomyces cerevisiae Taxane Biosynthesis and Overcoming Nutritional Stress-Induced Pseudohyphal Growth. Microorganisms 2022, 10, 163. [Google Scholar] [CrossRef]
- Besumbes, O.; Sauret-Güeto, S.; Phillips, M.A.; Imperial, S.; Rodríguez-Concepción, M.; Boronat, A. Metabolic engineering of isoprenoid biosynthesis in Arabidopsis for the production of taxadiene, the first committed precursor of Taxol. Biotechnol. Bioeng. 2004, 88, 168–175. [Google Scholar] [CrossRef]
- Kovacs, K.; Zhang, L.; Linforth, R.S.; Whittaker, B.; Hayes, C.J.; Fray, R.G. Redirection of carotenoid metabolism for the efficient production of taxadiene [taxa-4(5),11(12)-diene] in transgenic tomato fruit. Transgenic Res. 2007, 16, 121–126. [Google Scholar] [CrossRef]
- Rontein, D.; Onillon, S.; Herbette, G.; Lesot, A.; Werck-Reichhart, D.; Sallaud, C.; Tissier, A. CYP725A4 from yew catalyzes complex structural rearrangement of taxa-4(5),11(12)-diene into the cyclic ether 5(12)-oxa-3(11)-cyclotaxane. J. Biol. Chem. 2008, 283, 6067–6075. [Google Scholar] [CrossRef]
- Hasan, M.M.; Kim, H.-S.; Jeon, J.-H.; Kim, S.H.; Moon, B.; Song, J.-Y.; Shim, S.H.; Baek, K.-H. Metabolic engineering of Nicotiana benthamiana for the increased production of taxadiene. Plant Cell Rep. 2014, 33, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, X.; Li, X.; Shi, H.; Wang, F.; Zhang, Y.; Li, X. Combinatorial Engineering of Mevalonate Pathway and Diterpenoid Synthases in Escherichia coli for cis-Abienol Production. J. Agric. Food Chem. 2019, 67, 6523–6531. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Xu, W.; Huang, W.; Wang, B.; Li, S.; Zhang, J.; Chang, L. Importation of taxadiene synthase into chloroplast improves taxadiene production in tobacco. Planta 2021, 253, 107. [Google Scholar] [CrossRef] [PubMed]
- Cha, M.; Shim, S.H.; Kim, S.H.; Kim, O.T.; Lee, S.W.; Kwon, S.Y.; Baek, K.H. Production of taxadiene from cultured ginseng roots transformed with taxadiene synthase gene. BMB Rep. 2012, 45, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Anterola, A.; Shanle, E.; Perroud, P.F.; Quatrano, R. Production of taxa-4(5),11(12)-diene by transgenic Physcomitrella patens. Transgenic Res. 2009, 18, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Yuan, Y.; Tao, H.; Shi, X.; Zhong, X.; Han, Y.; Fu, S.; Fang, C.; Deng, Z.; Liu, T. Production of taxadiene by engineering of mevalonate pathway in Escherichia coli and endophytic fungus Alternaria alternata TPF6. Biotechnol. J. 2017, 12, 1–12. [Google Scholar] [CrossRef]
- Kirby, J.; Keasling, J.D. Biosynthesis of Plant Isoprenoids: Perspectives for Microbial Engineering. Annu. Rev. Plant Biol. 2009, 60, 335–355. [Google Scholar] [CrossRef]
- Li, B.J.; Wang, H.; Gong, T.; Chen, J.-J.; Chen, T.-J.; Yang, J.-L.; Zhu, P. Improving 10-deacetyl baccatin III-10-beta-O-acetyltransferase catalytic fitness for Taxol production. Nat. Commun. 2017, 8, 15544. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition or Disease | Partner Drugs | Condition or Disease | Partner Drugs |
|---|---|---|---|
| Lung cancer | Cisplatinum | Lymphoma | Cisplatinum |
| Gastric cancer | Tegafur | Adriamycin | |
| Capecitabine | Capecitabine | ||
| Head and neck tumors | Cisplatinum | Gemcitabine | |
| Esophageal cancer | Cisplatinum | Adriamycin + cyclophosphamide | |
| Capecitabine | Pancreatic | Gemcitabine |
| Partner Drugs | Condition or Disease | Phase | Clinical Trial Identifier |
|---|---|---|---|
| Gemcitabine | Refractory solid tumors | I | NCT03507491 |
| Raltitrexed | Advanced pancreatic cancer | II | NCT04581876 |
| Apatinib and camrelizumab | Advanced gastric cancer | I/II | NCT04286711 |
| AZD2014 | Advanced cancer | I | NCT02193633 |
| Fostamatinib | Ovarian cancer | I | NCT03246074 |
| Tilelizumab | High-risk non-muscle-invasive urothelial bladder carcinoma that is not completely resectable | II | NCT04730232 |
| LDE225 | Recurrent ovarian cancer | I | NCT02195973 |
| Camrelizumab | Non-small cell lung cancer | II | NCT04167774 |
| Gemcitabine and ficlatuzumab | Pancreatic cancer | I | NCT03316599 |
| Durvalumab | Squamous cell carcinoma of the head and neck | II | NCT03723967 |
| fruquintinib | Gastric cancer | III | NCT03223376 |
| Chiauranib | Ovarian cancer | III | NCT04921527 |
| Lovastatin | Ovarian cancer | II | NCT00585052 |
| Capibasertib | Locally advanced (inoperable) or metastatic triple-negative breast cancer | III | NCT03997123 |
| Pembrolizumab and carboplatin | Recurrent/metastatic head and neck squamous cell carcinoma | IV | NCT04489888 |
| Research Groups | Year | Synthetic Strategy | Starting Materials | Total Steps | Refs. |
|---|---|---|---|---|---|
| Nicolaou et al. | 1994 | (1) Coupling of the A and C rings by Shapiro reaction at the C1–C2 positions; (2) formation of the B ring by McMurry coupling at the C9–C10 positions; (3) and finally, formation of the side chain, selective oxidation of the C13 position and formation of the D ring. | Ethyl 4-hydroxy-2-methylbut-2-enoate and 3-hydroxy-2-pyrone | 51 | [53] |
| Holton et al. | 1994 | (1) Formation of AB ring by epoxy alcohol cleavage; (2) formation of the C ring by Dieckmann condensation; (3) formation of the D ring based on intramolecular SN2 cyclization; (4) introduction of the C9 oxygen functional group and side chain. | Camphor | 41 | [54,55] |
| Danishefsky et al. | 1996 | (1) Coupling of A and CD rings at the C1–C2 sites via 1,2-addition reactions; (2) generation of B rings via Heck coupling at the C9–C10 sites; (3) selective oxidation of C9 and C13 and formation of side chains. | 2-Methyl-1,3-cyclohexandione | 47 | [58] |
| Wender et al. | 1997 | (1) Formation of the AB ring by Grob-type fragmentation; (2) formation of the C ring by aldol cyclization reaction; (3) formation of the D ring based on intramolecular SN2 cyclization and introduction of the side chain. | Verbenone | 37 | [59,60] |
| Kuwajima et al. | 1998 | (1) Coupling of the A ring and C ring at the C1–C2 site by 1,2-addition reaction; (2) generation of the B ring at the C9–C10 site by Vinylogous Mukaiyama aldol reaction; (3) formation of the D ring and C13 side chain by introducing C19-methyl and the C3 standing center. | 2-(Prop-2-yn-1-yloxy)tetrahydro-2H-pyran | 47 | [61] |
| Mukaiyama et al. | 1999 | (1) Formation of octacycles at the C3–C8 sites by intramolecular aldol cyclization of SmI2; (2) formation of C rings at the C7–C8 sites based on Michael addition and intramolecular hydroxyl aldol cyclization; (3) formation of A rings at the C11–C12 sites based on pinacol coupling cyclization; (4) final selective oxidation of C13 as well as the formation of D rings and side chains. | Methyl 3-hydroxy-2,2-dimethylpropanoate | 38 | [62] |
| Kishi et al. | 2000 | (1) Introduction of a C8 all-carbon quaternary center by [2,3] rearrangement; (2) Coupling of A and C rings at the C1–C2 sites by 1,2-addition reaction; (3) generation of the B ring by NHK coupling at C9–C10 sites; (4) formation of the D ring and side chain, oxidation of C13. | 3-Methylcyclohex-2-en-1-ol | 45 | [56] |
| Takahashi et al. | 2006 | (1) Coupling of the A ring and CD ring at the C1–C2 sites by 1,2-addition reaction; (2) generation of the B ring by microwave-assisted alkylation at the C9–C10 sites; (3) selective oxidation of C9 and formation of the D ring. | Geraniol | 47 | [63] |
| Nakada et al. | 2015 | (1) Coupling of the A and C rings at the C1–C2 sites based on 1,2-addition reactions; (2) generation of the B ring by palladium-catalyzed alkenylation at the C9–C10 sites; (3) formation of the D ring by SN2 cyclization. | Acetal aldehyde and vinyl iodide | 37 | [64] |
| Chida et al. | 2015 | (1) Linking the A and C rings by 1,2-addition at the C1–C2 site; (2) forming the B ring by palladium-catalyzed alkenylation at the C10–C11 bond; (3) constructing the D ring of oxetane by SN2 cyclization. | Tri-O-acethl-D-glucal and 1,3-cyclohexanedione | 42 | [65,66] |
| Baran et al. | 2020 | (1) Type II intramolecular Diels–Alder reaction to form the ABC framework; (2) stereoselective oxidation to C13, C5, C10 and C9 sites; (3) dioxane-mediated C–H oxidation to produce bridging tertiary alcohols at the C1 site; (4) formation of the D ring and side chain. | 2,3-Dimethylbut-2-ene; 3-ethoxy-2-cyclohexen-1-one; CHBr3; acrylaldehyde | 24 | [67] |
| Li et al. | 2021 | (1) Asymmetric synthesis to form the AC ring; (2) SmI2-mediated pinacol coupling reaction to form the B ring; (3) generation of C3 stereocenters by the Hutchins–Kabalka method; (4) formation of the D ring as well as the introduction of the C13 side chain. | (2R, 3S)-2-Allyl-3-hydroxy-2-methylcyclohexan-1-one | 21 | [68] |
| Chida et al. | 2022 | (1) Linking the A and C rings by 1,2-addition at the C1–C2 site; (2) forming the B ring by palladium-catalyzed allylation at the C10–C11 bond; (3) forming the C13 and C5 hydroxyl groups by Rubottom oxidation; and (4) forming the D ring by a novel sliver-promoted cyclization method. | Tri-O-acethl-D-glucal | 22 | [69] |
| Inoue et al. | 2023 | (1) Intermolecular and intramolecular radical coupling processes to link and cyclize the A- and C-ring fragments, respectively; (2) efficient decoration of the A- and C-ring functional groups using newly discovered chemo-, regio- and stereoselective processes; (3) finally, D ring formation and conjugation with amino-acid-delivered taxol. | 2,2-Dimethylcyclohexane-1,3-dione | 34 | [70] |
| Family | Fungus | Host | Strain | Yield (μg/L) | Reference |
|---|---|---|---|---|---|
| Taxaceae | Alternaria alternata | T. hicksii | Tbp-9 | 0.13 | [92] |
| Alternaria alternata | T. hicksii | - | 332–512 | [89] | |
| Alternaria alternata | T. chinensis var mairei | TPF6 | 84.5 | [93] | |
| Alternaria sp. | T. cuspidata | Ja-69 | 0.16 | [94] | |
| Alternaria alternata | T. cuspidata | F3 | 195.4 | [95] | |
| Anthina Fr. | T. yunnanensis | Tax-15 | 6.23 | [96] | |
| Aspergillus candidus | Taxus media | MD2 | 112 | [97] | |
| Aspergillus candidus | T. media | MD3 | 73 | [98] | |
| Aspergillus fumigatus | Taxus sp. | TPF-06 | 1590.00 | [99] | |
| Aspergillus niger | T. cuspidata | HD86-9 | 273.46 | [100] | |
| Aspergillus niger | Taxus yunnanensis | IBFC-Z3S | 1000 | [101] | |
| Aspergillus niger var taxi | T. cuspidata | - | 91 | [102] | |
| Bionectria sp. | T. chinensis var mairei | XH004 | 33.90–430.46 | [103] | |
| Botryodiplodia theobromae | T. baccata | BT115 | 280.5 | [104] | |
| Botrytis sp. | T. cuspidata | HD181-23 | 206.34 | [105] | |
| Botrytis sp. | T. chinensis var mairei | XT-2 | 161.24 | [106] | |
| Botrytis taxi | T. cuspidata | HD104 | - | [107] | |
| Cephalosporium sp. | T. yunnanensis | Tax-36 | 3.781 | [96] | |
| Chaetomium sp. | T. yunnanensis | Tax-60 | 21.1 | [96] | |
| Cladosporium cladosporioides | T. media | MD2 | 80 | [97] | |
| Didymostilbe sp. | T. chinensis var mairei | DF110 | - | [108] | |
| Dimemasporium sp. | T. yunnanensis | Tax-35 | 3.34 | [96] | |
| Ectostroma sp. | T. chinensis var mairei | XT 5 | 276.75 | [106] | |
| Ectostroma sp. | T. yunnanensis | Tax-16 | 4.092 | [96] | |
| Ectostroma sp. | T. yunnanensis | Tax-25 | 2.16 | [96] | |
| Fusarium anthrosporioides | T. cuspidata | F-40 | 131 | [109] | |
| Fusarium lateritium | T. baccata | Tbp-9 | 0.13 | [94] | |
| Fusarium mairei | T. chinensis | - | 78 | [110] | |
| Fusarium mairei | Taxus × media | UH23 | 20 | [111] | |
| Fusarium redolens | T. baccata | TBPJ-B | 66 | [112] | |
| Fusarium solani | T. chinensis | Tax-3 | 164 | [113] | |
| Fusarium sp. | T. chinensis var mairei | D62 | 148.95 | [114] | |
| Fusarium sp. | T. chinensis var mairei | Y1117 | 2.70 | [115] | |
| Gliocladium sp. | T. baccata | - | 90 | [110] | |
| Gonatobotrys sp. | T. yunnanensis | Tax-13 | 4.092 | [96] | |
| Guignardia mangiferae | Taxus × media | HAA 11, HBA 29 | - | [116] | |
| Hypocrea sp. | T. media | Z58 | 2.50–3.00 | [117] | |
| Metarhizium anisopliae | T. chinensis | H-27 | 846.10 | [118] | |
| Monochaetia sp. | T. baccata | Tbp-2 | 0.1 | [92] | |
| Mucor rouxianus | T. chinensis | DA10 | 30 | [119] | |
| Mucor sp. | T. yunnanensis | Tax-56 | 1.08 | [96] | |
| Mucor sp. | T. media | 060B1 | 2.50–3.00 | [120] | |
| Nodulisporium sylviforme | T. cuspidata | HQD33, HQD48 | 51.06–125.70 | [121] | |
| Nodulisporium sylviforme | T. cuspidata | NCEU-1 | 314 | [121] | |
| Nodulisporium sylviforme | T. cuspidata | UV40-19, UL50-6 | 392 | [121] | |
| Nodulisporium sylviforme | T. cuspidata | HDF68 | 468.62 | [122] | |
| Nodulisporium sylviforme | T. cuspidata | - | 450 | [89] | |
| Nodulisporium sylviforme | T. cuspidata | HDFS4-26 | 516.37 | [105] | |
| Ozonium sp. | T. chinensis var mairei | BT 2 | 4–18 | [123] | |
| Papulaspora sp. | T. chinensis var mairei | XT17 | 10.25 | [106] | |
| Penicillium sp. | T. yunnanensis | Tax-20 | 8.24 | [96] | |
| Pestalotia bicilia | T. baccata | Tbx-2 | 1.08 | [92] | |
| Pestalotiopsis microspora | T. walichiana | Ne-32 | 50.00 | [92] | |
| Pestalotiopsis microspora | T. cuspidata | Ja-73 | 0.27 | [92] | |
| Pestalotiopsis sp. | T. yunnanensis | YN6 | 120–140 | [124] | |
| Pestalotiopsis terminaliae | T. arjuna | TAP 15 | 211.10 | [125] | |
| Phoma sp. | T. yunnanensis | Tax-26 | 18.56 | [96] | |
| Phoma sp. | T. yunnanensis | Tax-47 | 47.302 | [96] | |
| Phomopsis sp. | T. cuspidata | BKH27 | 418 | [126] | |
| Pithomyces sp. | T. sumatrana | P-96 | 0.095 | [92] | |
| Placodium sp. | T. yunnanensis | Tax-24 | 13.63 | [96] | |
| Placodium sp. | T. yunnanensis | Tax-49 | 31.06 | [96] | |
| Placodium sp. | T. yunnanensis | Tax-55 | 0.46 | [96] | |
| Placodium sp. | T. yunnanensis | Tax-63 | 3.11 | [96] | |
| Placodium sp. | T. yunnanensis | Tax-65 | 6.27 | [96] | |
| Rhizoctonia sp. | T. yunnanensis | Tax-1 | 1.43 | [96] | |
| Rhizopus | T. media | M57 | 45.00–50.00 | [127] | |
| Stemphylium sedicola | T. baccata | SBU-16 | 6.90 | [128] | |
| Taxomyces andreanae | T. brevifolia | Tbp-2 | 0.02–0.05 | [129] | |
| Trichoderma sp. | T. yunnanensis | Tax-23 | 19.59 | [96] | |
| Tubercularia sp. | T. chinensis var mairei | TF-5 | 185.40 | [130] | |
| Rhizosphere | Alternaria sp. | Rhizosphere | - | 4.2 | [110] |
| Aspergillus flavipes | Rhizosphere | - | 185–850 | [110] | |
| Aspergillus flavus | Rhizosphere | - | 2.8 | [110] | |
| Aspergillus oryzae | Rhizosphere | - | 3.2 | [110] | |
| Penicillium chrysogenum | Rhizosphere | - | 85 | [110] | |
| Pestalotiopsis malicola | Rhizosphere | - | 186 | [131] | |
| Bromeliaceae | Fusarium proliferatum | Tillandsia usneoides | - | 165 | [110] |
| Pestalotiopsis humus 133 | Tillandsia usneoides | - | 6.1 | [110] | |
| Pestalotiopsis humus 154 | Tillandsia usneoides | - | 5.7 | [110] | |
| Pestalotiopsis sp. 118 | Tillandsia usneoides | - | 8.9 | [110] | |
| Pestalotiopsis sp. 107 | Tillandsia usneoides | - | 89 | [110] | |
| Pestalotiopsis sp. 155 | Tillandsia usneoides | - | 4.3 | [110] | |
| Pestalotiopsis sp. 163 | Tillandsia usneoides | - | 4.0 | [110] | |
| Phomopsis sp. 116 | Tillandsia usneoides | - | 22 | [110] | |
| Araucariaceae | Pestalotiopsis guepinii | Wollemia nobilis | w-1, f-2 | 0.49 | [132] |
| Pestalotiopsis sp. | Wollemia nobilis | w-x-3 | 0.13 | [132] | |
| Pestalotiopsis sp. | Wollemia nobilis | w-1, f-1 | 0.17 | [132] | |
| Phomopsis sp. | Wollemia nobilis | - | 170 | [129] | |
| Cupressaceae | Fusarium mairei | Taxodium distichum | UH23 | 20.00 | [111] |
| Pestalotiopsis microspora | Taxodium distichum | Cp-4 | 0.01–1.49 | [133] | |
| Rutaceae | Bartalinia robillardoides | Aegle mamelos | - | 187.6 | [134] |
| Phyllosticta citricarpa | Citrus media | - | 265.00 | [110] | |
| Ginkgoaceae | Phoma betae | Ginkgo biloba | SBU-16 | 795.00 | [135] |
| Phomopsis sp. | Ginkgo biloba | - | 372 | [136] | |
| Rubiaceae Juss. | Lasiodiplodia theobromae | Morinda citrifolia | - | 120 | [110] |
| Pestalotiopsis microspora | Maguireothamnus speciosus | - | 0.11 | [92] | |
| Podocarpaceae | Aspergillus fumigatus | Podocarpus sp. | EPTP-1 | 560.00 | [137] |
| Sapindaceae Juss. | Pestalotiopsis pauciseta | Cardiospermum helicacabum | CHP-11 | 113.30 | [138] |
| Combretaceae R. Br. | Pestalotiopsis terminaliae | Terminalia arjuna | TAP-15 | 211.10 | [139] |
| Apocynaceae Juss. | Phyllosticta tabernaemontanae | Wrightia tinctoria | - | 461.00 | [140] |
| Sterculiaceae | Phyllostica melochiae | Melochia corchorifolia | - | 478 | [110] |
| Malvaceae Juss. | Phyllosticta dioscorea | Hibiscus rosa-sinensis | - | 298 | [136] |
| Moringa Adans. | Cladosporium oxysporum | Moringa oleifera | - | 550 | [141] |
| Betulaceae Gray | Penicillium aurantiogriseum | Corylus avellana | NRRL 62431 | 70 | [134] |
| Products | Concentration | Host | Reference |
|---|---|---|---|
| Taxadiene | 1.0 g/L | E. coli | [158] |
| Oxygenated taxanes | 570 mg/L | E. coli | [159] |
| Taxadiene | 1.3 mg/L | E. coli | [160] |
| Oxygenated taxanes | 33 mg/L | E. coli and S. cerevisiae | [161] |
| Taxadiene | 1.98 mg/L | Bacillus subtilis | [162] |
| Taxadiene and taxadiene-5α-ol | 1.0 mg/L and~25 μg/L | S. cerevisiae | [163] |
| Taxadiene | 8.7 mg/L | S. cerevisiae | [164] |
| Taxadiene | 72.8 mg/L | S. cerevisiae | [165] |
| Taxadiene | 20 mg/L | S. cerevisiae | [166] |
| Taxadiene | 129 mg/L | S. cerevisiae | [167] |
| Taxadien-5α-yl-acetate and total oxygenated taxane | 3.7 mg/L and 78 mg/L | S. cerevisiae | [168] |
| Taxadiene and taxadien-5α-yl-acetate | 71 mg/L and 21 mg/L | S. cerevisiae | [169] |
| Taxadiene | 600 ng/g DW | A. thaliana | [170] |
| Taxadiene | 160 mg/kg | Tomato fruits | [171] |
| Taxadiene and 5(12)-oxa-(11)-cyclotaxane | no yield | Tobacco (Nicotiana sylvestris) | [172] |
| Taxadiene | 27 μg/g DW | Tobacco (Nicotiana benthamiana) | [173] |
| Taxadiene and taxadiene-5α-ol | 56.6 μg/g and 1.3 μg/g FW | Tobacco (Nicotiana benthamiana) | [174] |
| Taxadiene | 87.7 μg/g DW | Nicotiana tabacum cv. Petit Havana | [175] |
| TS-transgenic ginseng | 14.6–15.9 μg/g DW | Ginseng (Panax ginseng) roots | [176] |
| Taxadiene | 0.05% FW of plant tissue | Physcomitrella patens (moss) | [177] |
| Taxadiene | 61.9 μg/L | Alternaria alternata (endophytic fungus) | [178] |
| Methods | Advantages | Disadvantages |
|---|---|---|
| Extraction from plants |
|
|
| Total synthesis |
|
|
| Semi-synthesis |
|
|
| Tissue and cell culture |
|
|
| endophytic fungi method |
|
|
| Synthetic biology method |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Ye, T.; Liu, Y.; Hou, G.; Wang, Q.; Zhao, F.; Li, F.; Meng, Q. Research Advances in Clinical Applications, Anticancer Mechanism, Total Chemical Synthesis, Semi-Synthesis and Biosynthesis of Paclitaxel. Molecules 2023, 28, 7517. https://doi.org/10.3390/molecules28227517
Zhang S, Ye T, Liu Y, Hou G, Wang Q, Zhao F, Li F, Meng Q. Research Advances in Clinical Applications, Anticancer Mechanism, Total Chemical Synthesis, Semi-Synthesis and Biosynthesis of Paclitaxel. Molecules. 2023; 28(22):7517. https://doi.org/10.3390/molecules28227517
Chicago/Turabian StyleZhang, Shengnan, Taiqiang Ye, Yibin Liu, Guige Hou, Qibao Wang, Fenglan Zhao, Feng Li, and Qingguo Meng. 2023. "Research Advances in Clinical Applications, Anticancer Mechanism, Total Chemical Synthesis, Semi-Synthesis and Biosynthesis of Paclitaxel" Molecules 28, no. 22: 7517. https://doi.org/10.3390/molecules28227517
APA StyleZhang, S., Ye, T., Liu, Y., Hou, G., Wang, Q., Zhao, F., Li, F., & Meng, Q. (2023). Research Advances in Clinical Applications, Anticancer Mechanism, Total Chemical Synthesis, Semi-Synthesis and Biosynthesis of Paclitaxel. Molecules, 28(22), 7517. https://doi.org/10.3390/molecules28227517