
Antioxidant and Antiproliferative Activities of Eclipta prostrata (L.) L. Extract and Isolated Compounds


Abstract
:1. Introduction
2. Results and Discussion
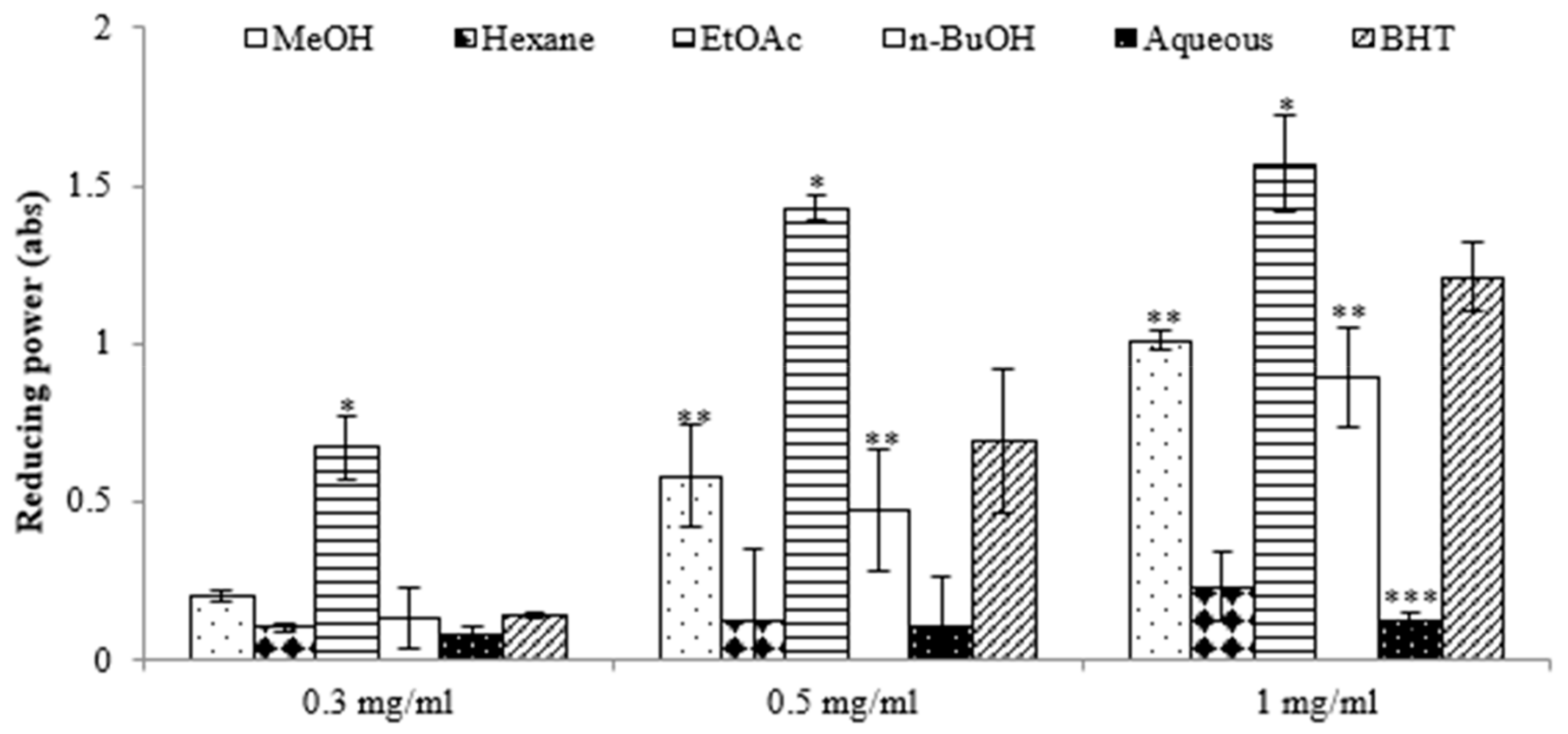
2.1. Antioxidant Activities of E. prostrata Extracts
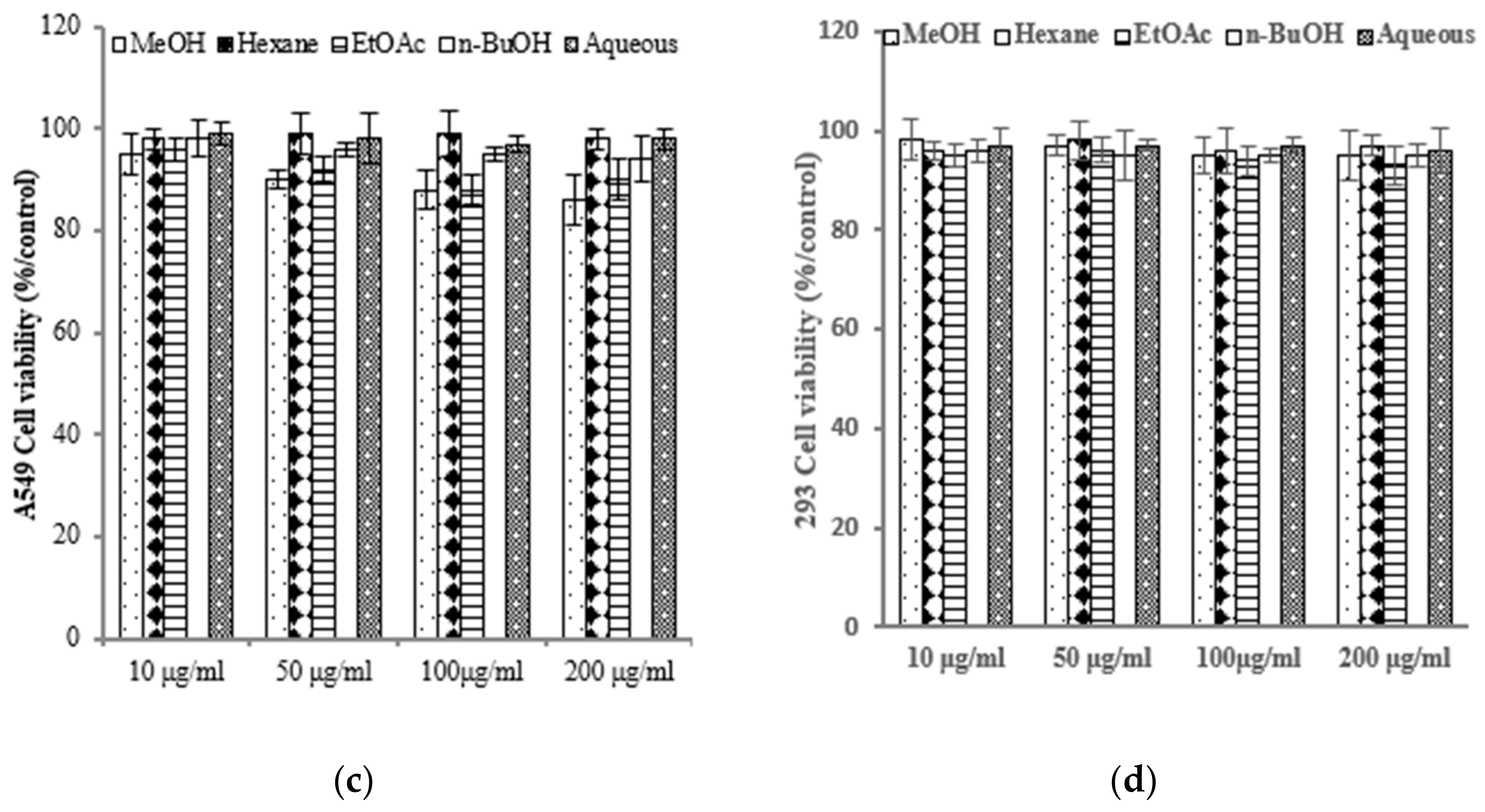
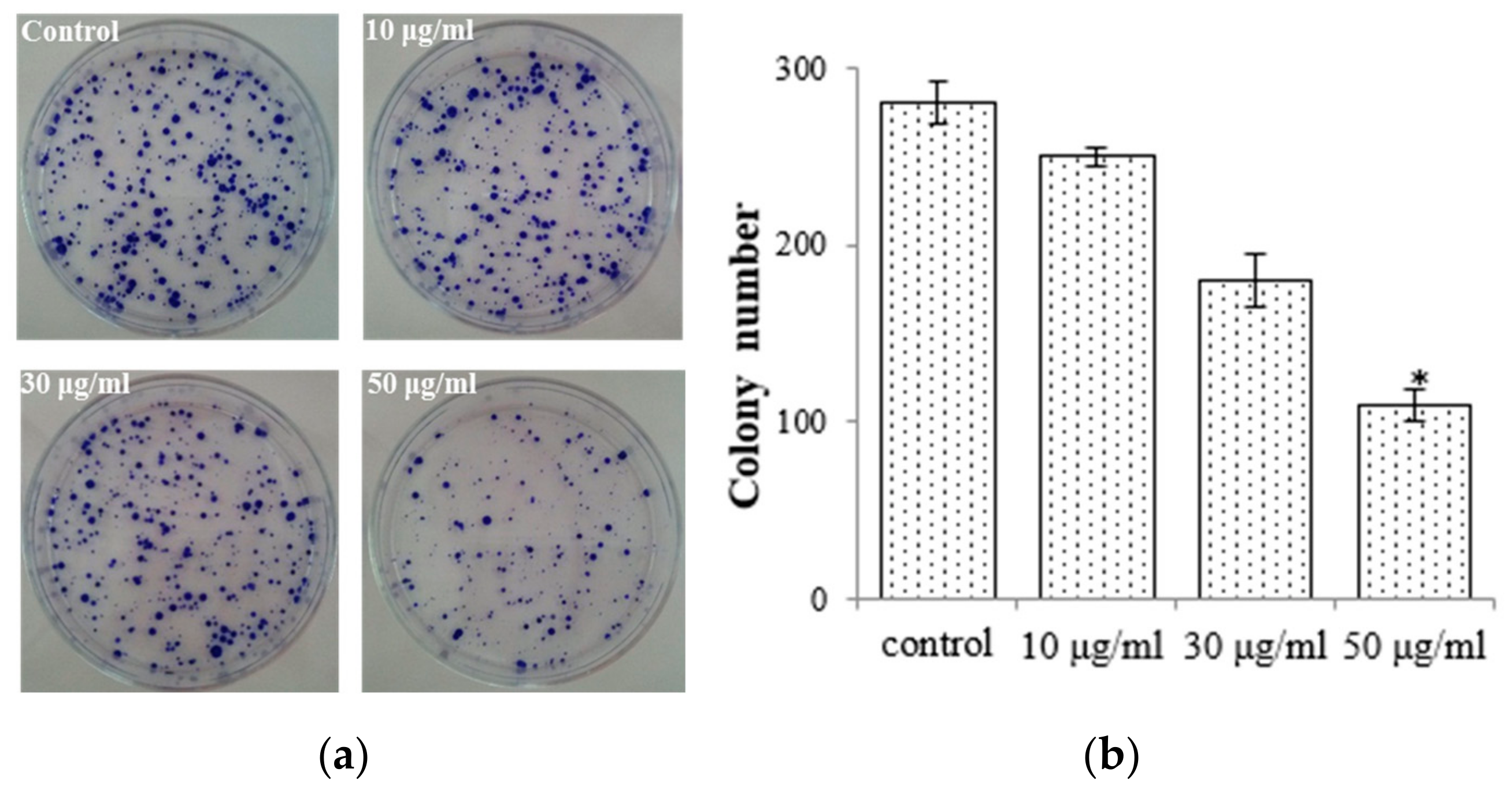
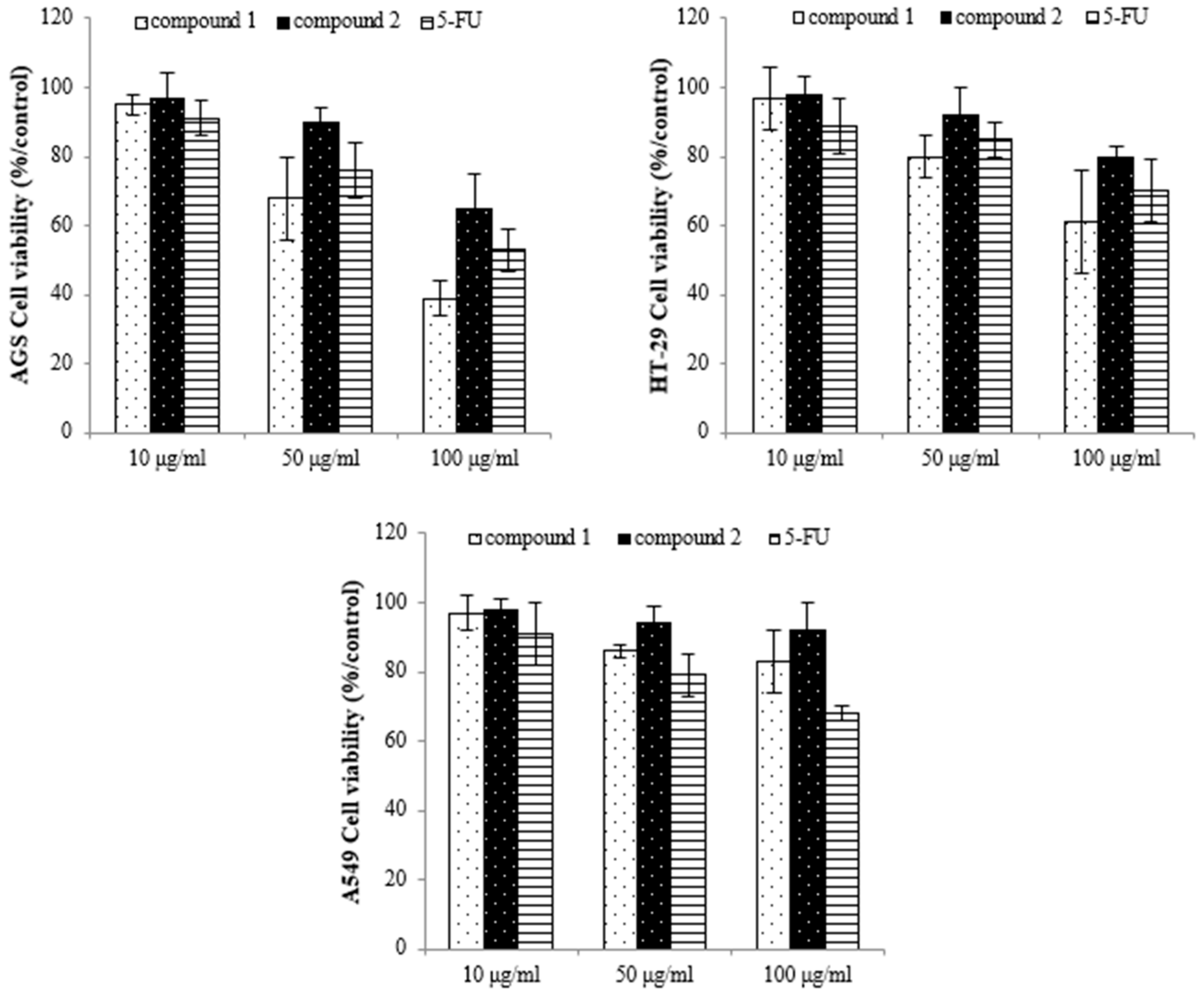
2.2. E. prostrata Affects Cancer Cell Growth
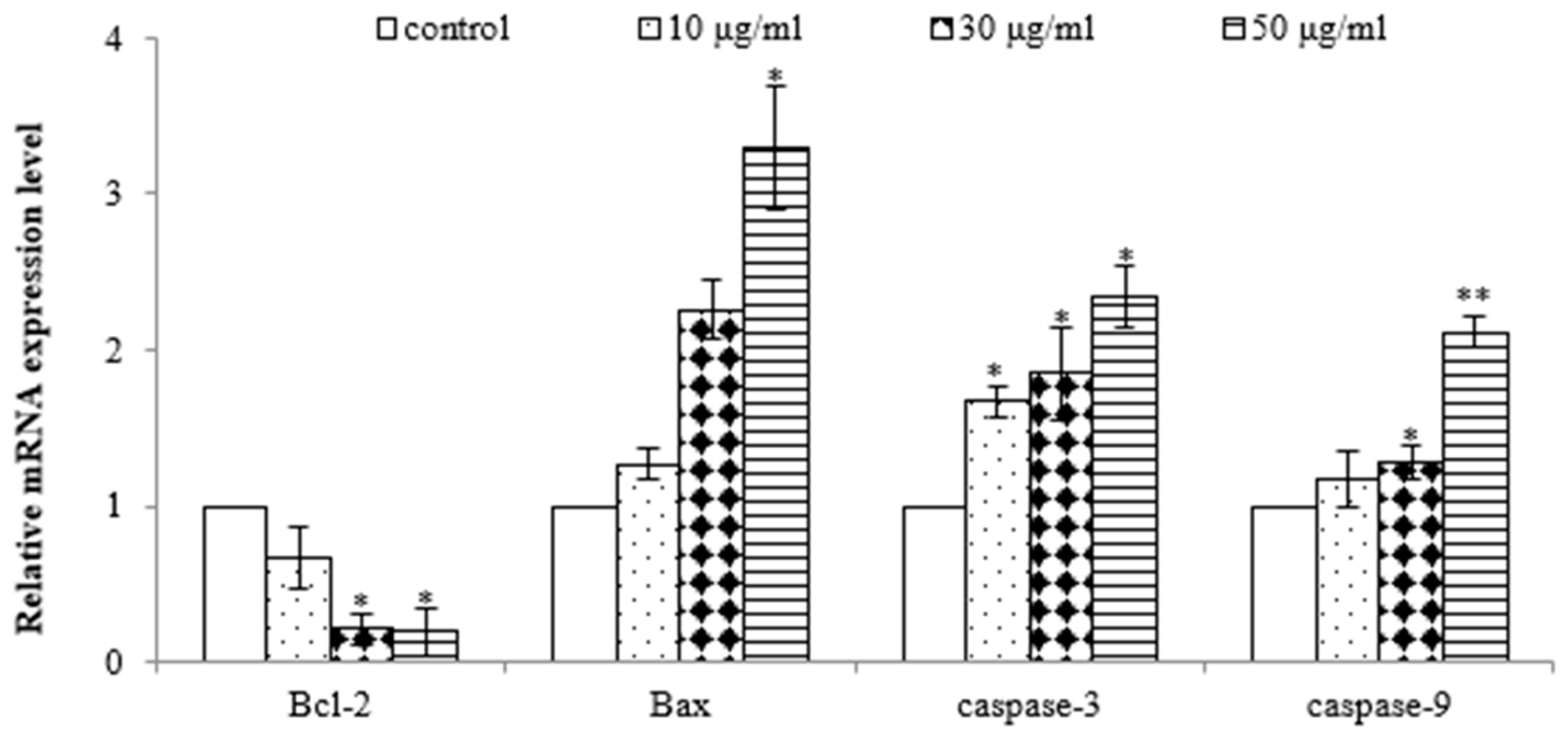
2.3. E. prostrata Induces Apoptosis of Gastric Carcinoma Cells
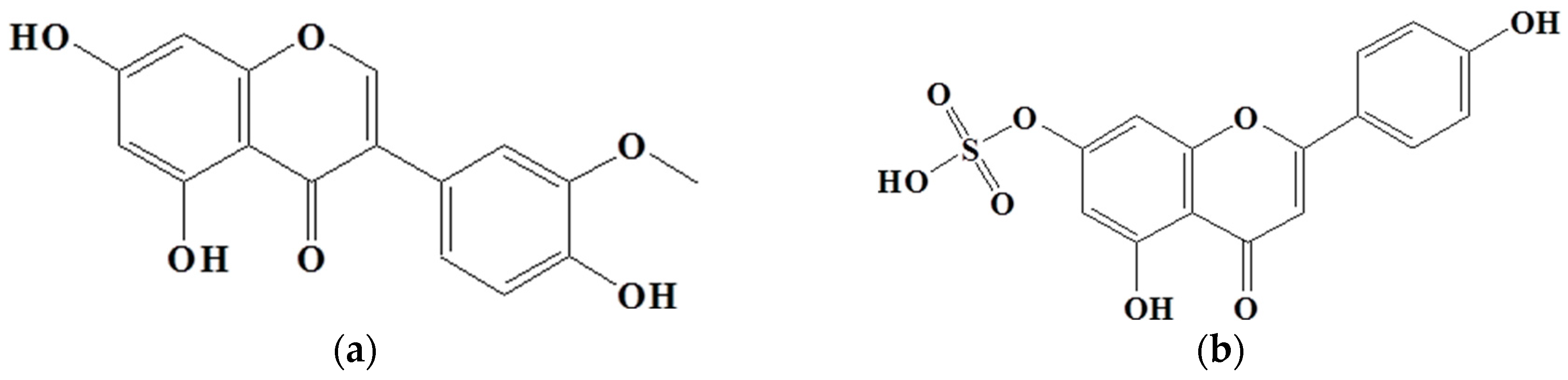
2.4. Isolation of Active Compounds
3. Materials and Methods
3.1. Plant Materials
3.2. DPPH Free Radical Scavenging Activity
3.3. Analysis of Reducing-Power Activity
3.4. Total Phenolic and Flavonoid Contents
3.5. Cell Culture
3.6. Cell Viability
3.7. Colony Formation Assay
3.8. Cell Cycle Assay
3.9. qRT-PCR Analysis
3.10. Statistics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deepak, T.; Hari, P.D. Eclipta prostrata (L.) L. (Asteraceae): Ethnomedicinal Uses, Chemical Constituents, and Biological Activities. Biomolecules 2021, 22, 1738. [Google Scholar]
- Li, F.; Zhai, Y.Y.; Xu, J.; Li, W.F. A review on traditional uses, phytochemistry and pharmacology of Eclipta prostrata (L.) L. J. Ethnopharmacol. 2019, 245, 112109. [Google Scholar]
- Adhikari, M.; Thapa, R.; Kunwar, R.M.; Devkota, H.P.; Poudel, P. Ethnomedicinal uses of plant resources in the Machhapuchchhre rural municipality of Kaski District. Nepal. Med. 2019, 6, 69. [Google Scholar]
- Diogo, L.C.; Fernandes, R.S.; Marcussi, S.; Menaldo, D.L.; Roberto, P.G.; Matrangulo, P.V.F.; Pereira, P.S.; França, S.C.; Giuliatti, S.; Soares, A.M. Inhibition of Snake Venoms and Phospholipases A2 by Extracts from Native and Genetically Modified Eclipta alba: Isolation of Active Coumestans. Basic Clin. Pharmacol. Toxicol. 2009, 104, 293–299. [Google Scholar] [CrossRef]
- Chung, I.M.; Rajakumar, G.; Lee, J.H.; Kim, S.H.; Thiruvengadam, M. Ethnopharmacological uses, phytochemistry, biological activities, and biotechnological applications of Eclipta prostrata. Appl. Microbiol. Biotechnol. 2017, 101, 5247–5257. [Google Scholar] [PubMed]
- Wang, L.; Yuan, S.; Rui, W.; Deding, S.; Tang, M.F.; Liu, Y.D.; Li, Z.G. Antioxidant activity and healthy benefits of natural pigments in fruits: A review. Int. J. Mol. Sci. 2021, 22, 4945. [Google Scholar]
- Gao, Z.; Gao, W.; Zeng, S.L.; Li, P.; Liu, E.H. Chemical structures, bioactivities and molecular mechanisms of citrus poly methoxyflavones. J. Funct. Foods 2018, 40, 498–509. [Google Scholar] [CrossRef]
- Duan, L.; Dou, L.L.; Yu, K.Y.; Guo, L.; Bai, C.; Li, P.; Liu, E.H. Polymethoxyflavones in peel of Citrus reticulata ‘Chachi’ and their biological activities. Food Chem. 2017, 234, 254–261. [Google Scholar]
- Albuquerque, M.A.C.; Levit, R.; Beres, C.; Bedani, R.; LeBlanc, A.D.M.D.; Saad, S.M.I.; LeBlanc, J.G. Tropical fruit by-products water extracts as sources of soluble fibres and phenolic compounds with potential antioxidant, anti-inflammatory, and functional properties. J. Funct. Foods 2019, 52, 724–733. [Google Scholar] [CrossRef]
- Kim, H.Y.; Kim, H.M.; Ryu, B.; Lee, J.S.; Choi, J.H.; Jang, D.S. Constituents of the aerial parts of Eclipta prostrata and their cytotoxicity on human ovarian cancer cells in vitro. Arch Pharm. Res. 2015, 38, 1963–1969. [Google Scholar] [PubMed]
- Ardestani, A.; Yazdanparast, R. Antioxidant and free radical scavenging potential of Achillea santolina extracts. Food Chem. 2007, 104, 21–29. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Res. Int. 2020, 132, 109114. [Google Scholar] [CrossRef]
- Bakoyiannis, I.; Daskalopoulou, A.; Pergialiotis, V.; Perrea, D. Phytochemicals and cognitive health: Are flavonoids doing the trick? Biomed. Pharmacother. 2019, 109, 1488–1497. [Google Scholar] [CrossRef]
- Lei, M.; Hua, X.; Xiao, M.; Ding, J.; Han, Q.Y. Impairments of astrocytes are involved in the D-galactose-induced brain aging. Bioch. Bioph. Res. Commun. 2008, 369, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.L.; Yin, Y.G. In vivo antioxidant activity of total flavonoids from indocalamus leaves in aging mice caused by D-galactose. Food Chem. Toxicol. 2012, 50, 3814–3818. [Google Scholar] [CrossRef] [PubMed]
- Tae, K.H.; Kim, M.K.; Lee, H.Y.; Kim, Y.J.; Kim, E.Y.; Kim, J.S. Evaluation of anti-oxidant and anti-cancer properties of Dendropanax morbifera Léveille. Food Chem. 2013, 141, 1947–1955. [Google Scholar]
- Fleischauer, A.T.; Simonsen, N.; Arab, L. Antioxidant supplements and risk of breast cancer recurrence and breast cancer-related mortality among postmenopausal women. Nutr. Cancer 2003, 46, 15–22. [Google Scholar] [CrossRef]
- Nishihara, H.; Kizaka-Kondoh, S.; Insel, P.A.; Eckmann, L. Inhibition of apoptosis in normal and transformed intestinal epithelial cells by cAMP through induction of inhibitor of apoptosis protein (IAP)-2. Proc. Natl. Acad. Sci. USA 2003, 100, 8921–8926. [Google Scholar] [CrossRef]
- Primchanien, M.; Nuttavut, K.; Sineenart, K.; Omboon, L.; Narongchai, P.X. Antiproliferation, antioxidation and induction of apoptosis by Garcinia mangostana (mangosteen) on SKBR3 human breast cancer cell line. J. Ethnopharmacol. 2006, 92, 161–166. [Google Scholar]
- Wang, X.J.; Yuan, S.L.; Wang, J.; Lin, P.; Liu, G.J.; Wei, Y.Q. Anticancer activity of litchi fruit pericarp extract against human breast cancer in vitro and in vivo. Toxicol. Appl. Pharm. 2006, 215, 168–178. [Google Scholar] [CrossRef]
- Saeed, N.; Muhammad, R.K.; Maria, S. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts. BMC Comp. Alt. Med. 2012, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.Y.; Yu, X.M.; Meenu, M.; Xu, B.J. Total phenolics and antioxidants profiles of commonly consumed edible flowers in China. Int. J. Food Prop. 2018, 21, 1524–1540. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Sahreen, S.; Khan, M.R.; Khan, R.A. Phenolic compounds and antioxidant activities of Rumex hastatus D. Don. Leaves. J. Med. Plants Res. 2011, 5, 2755–2765. [Google Scholar]
- Abu Bakar, M.F.; Ahmad, N.E.; Suleiman, M.; Rahmat, A.; Isha, A. Garcinia dulcis fruit extract induced cytotoxicity and apoptosis in HepG2 liver cancer cell line. BioMed Res. Int. 2015, 2015, 916902. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, C.; Shang, L.; Zhang, Y.; Zou, R.; Zhan, Y.; Bi, B. Sulforaphane induces apoptosis and inhibits invasion in U251MG glioblastoma cells. Springerplus 2016, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.Z.; Adebayo, I.A.; Mohamed, W.A.S.; Mohamad Zain, N.N.; Arsad, H. Christia vespertilionis extract induced antiproliferation and apoptosis in breast cancer (MCF7) cells. Mol. Biol. Rep. 2021, 48, 7361–7370. [Google Scholar] [CrossRef]
- Cui, D.; Liang, T.; Sun, L.; Meng, L.; Yang, C.; Wang, L.; Li, Q. Green synthesis of selenium nanoparticles with extract of hawthorn fruit induced HepG2 cells apoptosis. Pharm. Biol. 2018, 56, 528–534. [Google Scholar] [CrossRef]
- Ahmad, S.; Riaz, N.; Saleem, M.; Jabbar, A.; Ashraf, M. Antioxidant flavonoids from Alhagi maurorum. J. Asian. Nat. Prod. Res. 2010, 12, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Deng, X.; Zhang, L.; Shu, P.; Qin, M. A new coumestan with immunosuppressive activities from Flemingia philippinensis. Fitoterapia 2011, 82, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wu, Y.; Xue, S.; Wang, S.; Zhang, C.; Cao, Z. 3′-O-Methylorobol inhibits the voltage-gated sodium channel Nav1.7 with anti-itch efficacy in a histamine-dependent itch mouse model. Int. J. Mol. Sci. 2019, 20, 6058. [Google Scholar] [CrossRef]
- Seo, J.W.; Ham, D.Y.; Lee, J.G.; Kim, N.Y.; Kim, M.J.; Yu, C.Y.; Seong, E.S. Antioxidant Activity, Phenolic Content, and Antioxidant Gene Expression in Genetic Resources of Sorghum Collected from Australia, Former Soviet Union, USA, Sudan and Guadeloupe. Agronomy 2023, 13, 1698. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Park, Y.K.; Koo, M.H.; Ikegaki, M.; Contado, J.L. Comparison of the flavonoid aglycone contents of Apis mellifera propolis from various regions of Brazil. Arq. Biol. Tecnol. 1997, 40, 97–106. [Google Scholar]
- Arung, E.T.; Shimizu, K.; Kondo, R. Inhibitory effect of artocarpanone from Artocarpus heterophyllus on melanin biosynthesis. Biol. Pharm. Bull. 2006, 29, 1966–1969. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | DPPH Radical Scavenging Activity IC50 (μg/mL) | Total Phenolic Contents (mg GAE/g) | Total Flavonoid Contents (mg QE/g) |
|---|---|---|---|
| MeOH | 88.53 ± 0.83 b | 63.22 ± 2.90 b | 15.62 ± 0.29 b |
| Hexane | 189.77 ± 5.77 c | 14.82 ± 2.09 d | 2.11 ± 0.01 d |
| EtOAc | 26.12 ± 1.83 a | 124.56 ± 1.35 a | 24.69 ± 0.48 a |
| n-BuOH | 396.67 ± 8.72 d | 37.39 ± 4.23 c | 5.22 ± 0.64 c |
| Aqueous | ND | 5.27 ± 0.22 e | 1.58 ± 0.25 d |
| BHT | 109.61 ± 2.98 |
| Primer | Sequences | |
|---|---|---|
| Bax | Forward | 5′-GTTGTCGCCCTTTTCTACTTTG-3′ |
| Reverse | 5′-GCACTTTCTTCGCAGTTTCC-3′ | |
| Bcl-2 | Forward | 5′-GTGGATGACTGAGTACCTGAAC-3′ |
| Reverse | 5′-GGACATCAGTCGCTTCAGTG-3′ | |
| Caspase-3 | Forward | 5′-GGATGGGTGCTATTGTGAGG-3′ |
| Reverse | 5′-TGGGATTTCAAGGCGACG-3′ | |
| Caspase-9 | Forward | 5′-AGAGATTCGCAAACCAGAGG-3′ |
| Reverse | 5′-CACGGCAGAAGTTCACATTG-3′ | |
| β-actin | Forward | 5′-ACCACACCTTCTACAATGAGC-3′ |
| Reverse | 5′-GCGTACAGGGATAGCACAG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Kim, J.S.; Kwon, Y.S.; Seong, E.S.; Kim, M.J. Antioxidant and Antiproliferative Activities of Eclipta prostrata (L.) L. Extract and Isolated Compounds. Molecules 2023, 28, 7354. https://doi.org/10.3390/molecules28217354
Yang J, Kim JS, Kwon YS, Seong ES, Kim MJ. Antioxidant and Antiproliferative Activities of Eclipta prostrata (L.) L. Extract and Isolated Compounds. Molecules. 2023; 28(21):7354. https://doi.org/10.3390/molecules28217354
Chicago/Turabian StyleYang, Jinfeng, Joo Seok Kim, Yong Soo Kwon, Eun Soo Seong, and Myong Jo Kim. 2023. "Antioxidant and Antiproliferative Activities of Eclipta prostrata (L.) L. Extract and Isolated Compounds" Molecules 28, no. 21: 7354. https://doi.org/10.3390/molecules28217354
APA StyleYang, J., Kim, J. S., Kwon, Y. S., Seong, E. S., & Kim, M. J. (2023). Antioxidant and Antiproliferative Activities of Eclipta prostrata (L.) L. Extract and Isolated Compounds. Molecules, 28(21), 7354. https://doi.org/10.3390/molecules28217354