
Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review


 ,
,  ,
,
Abstract

1. Introduction
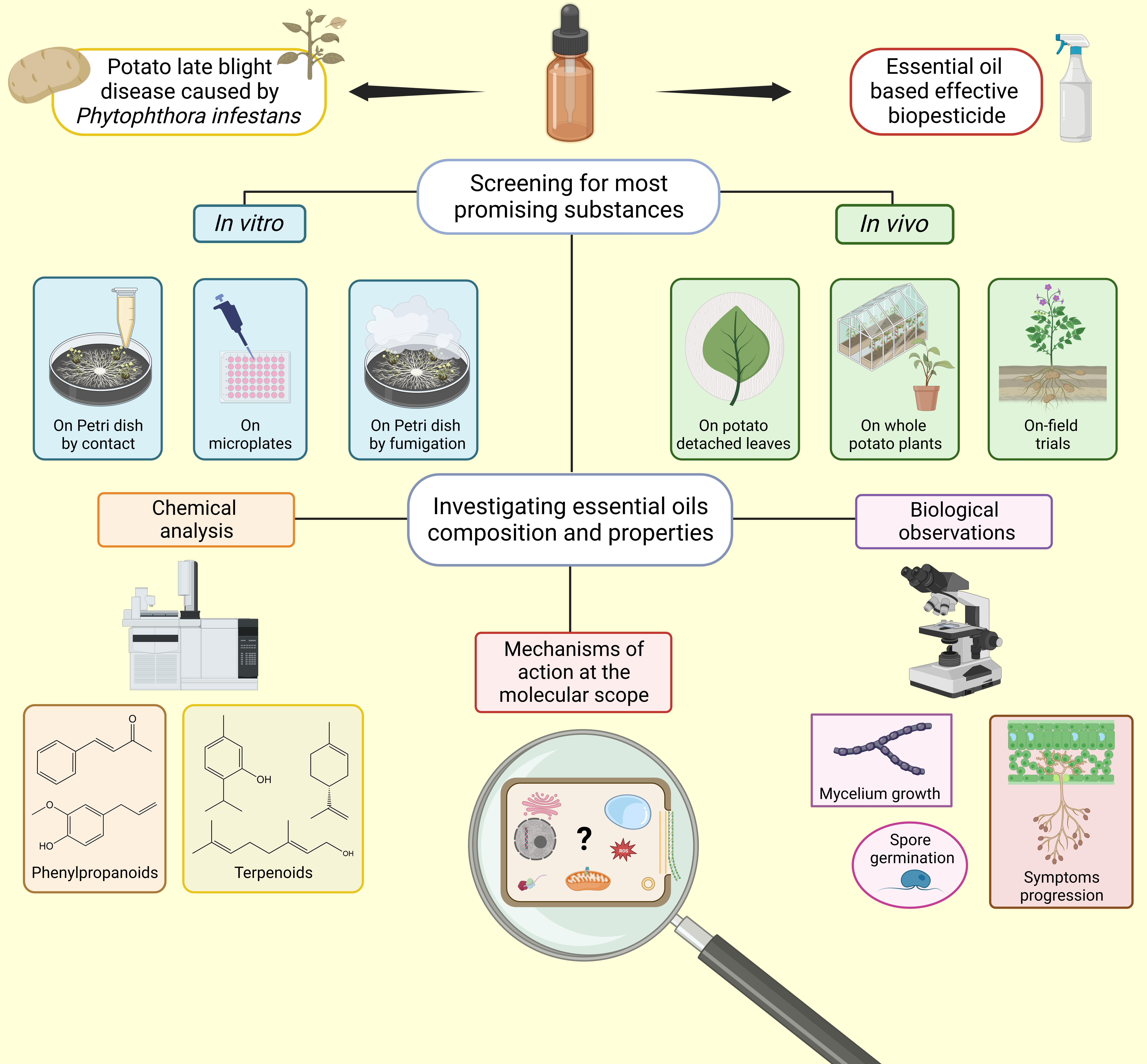
2. Essential Oils as Alternative Management against P. infestans
2.1. Essential Oils Description and Fields of Application
2.2. Assessing Anti-Oomycete Activities of Natural Substances at Different Laboratory Scales
2.3. Insights of Essential Oils Activity on P. infestans
3. Investigating Mechanisms of Action of Essential Oil Components
3.1. Chemical Composition and Variability of Essential Oils
3.2. Cellular Impacts of Essential Oil Components on Oomycetes
3.3. VOCs Interactions Modulate Biological Activities
4. Overcoming Essential Oils Challenges for Biopesticides Development
5. Discussion
6. Conclusions
- Context on potato and associated diseases: Solanum tuberosum is one of the most important crops in terms of human consumption and Phytophthora infestans—an oomycete causing potato late blight—represents its main threat.
- Challenges for late blight control: Synthetics pesticides are harmful to human health, the environment, and biodiversity; thus, biocontrol tools, in particular, natural molecules extracted from plants, such as essential oils, are gaining interest.
- Current research status: Numerous in vitro studies demonstrated the efficacy of essential oils, but in vivo trials are still lacking.
- Inconsistencies in the results: Essential oils tested against P. infestans are not unanimous in their effectiveness and do not systematically present same potential at different experimental and practical scales.
- Incomplete understanding of mechanisms of action: Essential oil components primarily target cell walls and membranes but also other cellular structures, which must be further explored.
- High diversity of VOCs composition: Investigation on essential oil major compounds may allow better comprehension of the global mechanisms of action.
- Main challenges for EO-based biopesticides: Finding substances that specifically disturb Phytophthora cellular machinery without impacting the host plant (phytotoxicity) nor other living organisms (ecotoxicity).
- Need for optimal formulation: EO requires appropriate physico-chemical methods to ensure stability, target-specific delivery, and long-term activity.
- Take-home message: Essential oils definitely present high anti-oomycete potential to cure diseases such as late blight caused by Phytophthora infestans; however, cellular sites of action must be better understood, and appropriate formulations developed to obtain effective biopesticides.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devaux, A.; Goffart, J.-P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global Food Security, Contributions from Sustainable Potato Agri-Food Systems. In The Potato Crop: Its Agricultural, Nutritional and Social Contribution to Humankind; Campos, H., Ortiz, O., Eds.; Springer International Publishing: Cham, Swizerland, 2020; pp. 3–35. ISBN 978-3-03028-683-5. [Google Scholar]
- Haverkort, A.J.; Struik, P.C. Yield Levels of Potato Crops: Recent Achievements and Future Prospects. Field Crops Res. 2015, 182, 76–85. [Google Scholar] [CrossRef]
- Raymundo, R.; Asseng, S.; Robertson, R.; Petsakos, A.; Hoogenboom, G.; Quiroz, R.; Hareau, G.; Wolf, J. Climate Change Impact on Global Potato Production. Eur. J. Agron. 2018, 100, 87–98. [Google Scholar] [CrossRef]
- Djaman, K.; Koudahe, K.; Koubodana, H.D.; Saibou, A.; Essah, S. Tillage Practices in Potato (Solanum tuberosum L.) Production: A Review. Am. J. Potato Res. 2022, 99, 1–12. [Google Scholar] [CrossRef]
- Haverkort, A.J. Ecology of Potato Cropping Systems in Relation to Latitude and Altitude. Agric. Syst. 1990, 32, 251–272. [Google Scholar] [CrossRef]
- Dupuis, J.H.; Liu, Q. Potato Starch: A Review of Physicochemical, Functional and Nutritional Properties. Am. J. Potato Res. 2019, 96, 127–138. [Google Scholar] [CrossRef]
- Navarre, D.A.; Goyer, A.; Shakya, R. Chapter 14—Nutritional Value of Potatoes: Vitamin, Phytonutrient, and Mineral Content. In Advances in Potato Chemistry and Technology; Singh, J., Kaur, L., Eds.; Washington State University: Prosser, WA, USA, 2009; pp. 395–424. ISBN 978-0-12-374349-7. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A Molecular Phylogeny of Phytophthora and Related Oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Kroon, L.P.N.M.; Brouwer, H.; de Cock, A.W.A.M.; Govers, F. The Genus Phytophthora Anno 2012. Phytopathology 2012, 102, 348–364. [Google Scholar] [CrossRef]
- Sogin, M.L.; Silberman, J.D. Evolution of the Protists and Protistan Parasites from the Perspective of Molecular Systematics. Int. J. Parasitol. 1998, 28, 11–20. [Google Scholar] [CrossRef]
- Crous, P.W.; Rossman, A.Y.; Aime, M.C.; Allen, W.C.; Burgess, T.; Groenewald, J.Z.; Castlebury, L.A. Names of Phytopathogenic Fungi: A Practical Guide. Phytopathology 2021, 111, 1500–1508. [Google Scholar] [CrossRef]
- Carlile, M.J. The Success of the Hypha and Mycelium. In The Growing Fungus; Gow, N.A.R., Gadd, G.M., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 3–19. ISBN 978-0-585-27576-5. [Google Scholar]
- Werner, S.; Steiner, U.; Becher, R.; Kortekamp, A.; Zyprian, E.; Deising, H.B. Chitin Synthesis during in Planta Growth and Asexual Propagation of the Cellulosic Oomycete and Obligate Biotrophic Grapevine Pathogen Plasmopara Viticola. FEMS Microbiol. Lett. 2002, 208, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Chérif, M.; Benhamou, N.; Belanger, R. Occurrence of Cellulose and Chitin in the Hyphal Walls of Pythium Ultimum: A Comparative Study with Other Plant Pathogenic Fungi. Can. J. Microbiol. 2011, 39, 213–222. [Google Scholar] [CrossRef]
- Ivanov, A.A.; Ukladov, E.O.; Golubeva, T.S. Phytophthora infestans: An Overview of Methods and Attempts to Combat Late Blight. J. Fungi 2021, 7, 1071. [Google Scholar] [CrossRef]
- Goss, E.M.; Tabima, J.F.; Cooke, D.E.L.; Restrepo, S.; Fry, W.E.; Forbes, G.A.; Fieland, V.J.; Cardenas, M.; Grünwald, N.J. The Irish Potato Famine Pathogen Phytophthora infestans Originated in Central Mexico Rather than the Andes. Proc. Natl. Acad. Sci. USA 2014, 111, 8791–8796. [Google Scholar] [CrossRef] [PubMed]
- Grünwald, N.J.; Flier, W.G. The Biology of Phytophthora infestans at Its Center of Orgin. Annu. Rev. Phytopathol. 2005, 43, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Ristaino, J.B. Tracking Historic Migrations of the Irish Potato Famine Pathogen, Phytophthora infestans. Microbes Infect. 2002, 4, 1369–1377. [Google Scholar] [CrossRef]
- Schumann, G.L.; D’Arcy, C.J. CHAPTER 1: The Irish Potato Famine: The Birth of Plant Pathology. In Hungry Planet: Stories of Plant Diseases; General Plant Pathology; The American Phytopathological Society: Saint Paul, MN, USA, 2017; pp. 1–19. ISBN 978-0-89054-490-7. [Google Scholar]
- Nowicki, M.; Foolad, M.R.; Nowakowska, M.; Kozik, E.U. Potato and Tomato Late Blight Caused by Phytophthora infestans: An Overview of Pathology and Resistance Breeding. Plant Dis. 2012, 96, 4–17. [Google Scholar] [CrossRef]
- Fawke, S.; Doumane, M.; Schornack, S. Oomycete Interactions with Plants: Infection Strategies and Resistance Principles. Microbiol. Mol. Biol. Rev. 2015, 79, 263–280. [Google Scholar] [CrossRef]
- Fry, W.E.; Birch, P.R.J.; Judelson, H.S.; Grünwald, N.J.; Danies, G.; Everts, K.L.; Gevens, A.J.; Gugino, B.K.; Johnson, D.A.; Johnson, S.B.; et al. Five Reasons to Consider Phytophthora infestans a Reemerging Pathogen. Phytopathology 2015, 105, 966–981. [Google Scholar] [CrossRef]
- Olanya, M.; Anwar, M.; He, Z.; Larkin, R.; Honeycutt, C. Survival Potential of Phytophthora infestans Sporangia in Relation to Environmental Factors and Late Blight Occurrence. J. Plant Prot. Res. 2016, 56, 73–81. [Google Scholar] [CrossRef]
- Hagman, J.E.; Mårtensson, A.; Grandin, U. Cultivation Practices and Potato Cultivars Suitable for Organic Potato Production. Potato Res. 2009, 52, 319–330. [Google Scholar] [CrossRef]
- Fry, W. Phytophthora infestans: The Plant (and R Gene) Destroyer. Mol. Plant Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Ren, X.; Yun, L.; Hou, Q.; Feng, F.; Liu, H. Simple and Inexpensive Long-Term Preservation Methods for Phytophthora infestans. J. Microbiol. Methods 2018, 152, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Gavino, P.D.; Smart, C.D.; Sandrock, R.W.; Miller, J.S.; Hamm, P.B.; Lee, T.Y.; Davis, R.M.; Fry, W.E. Implications of Sexual Reproduction for Phytophthora infestans in the United States: Generation of an Aggressive Lineage. Plant Dis. 2000, 84, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Leesutthiphonchai, W.; Vu, A.L.; Ah-Fong, A.M.V.; Judelson, H.S. How Does Phytophthora infestans Evade Control Efforts? Modern Insight Into the Late Blight Disease. Phytopathology 2018, 108, 916–924. [Google Scholar] [CrossRef]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 Oomycete Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef]
- Fry, W.E.; McGrath, M.T.; Seaman, A.; Zitter, T.A.; McLeod, A.; Danies, G.; Small, I.M.; Myers, K.; Everts, K.; Gevens, A.J.; et al. The 2009 Late Blight Pandemic in the Eastern United States—Causes and Results. Plant Dis. 2013, 97, 296–306. [Google Scholar] [CrossRef]
- Cooke, D.E.L.; Cano, L.M.; Raffaele, S.; Bain, R.A.; Cooke, L.R.; Etherington, G.J.; Deahl, K.L.; Farrer, R.A.; Gilroy, E.M.; Goss, E.M.; et al. Genome Analyses of an Aggressive and Invasive Lineage of the Irish Potato Famine Pathogen. PLoS Pathog. 2012, 8, e1002940. [Google Scholar] [CrossRef]
- Fry, W.E. Phytophthora infestans: New Tools (and Old Ones) Lead to New Understanding and Precision Management. Annu. Rev. Phytopathol. 2016, 54, 529–547. [Google Scholar] [CrossRef]
- Beninal, L.; Bouznad, Z.; Corbière, R.; Belkhiter, S.; Mabon, R.; Taoutaou, A.; Keddad, A.; Runno-Paurson, E.; Andrivon, D. Distribution of Major Clonal Lineages EU_13_A2, EU_2_A1, and EU_23_A1 of Phytophthora infestans Associated with Potato Late Blight across Crop Seasons and Regions in Algeria. Plant Pathol. 2022, 71, 458–469. [Google Scholar] [CrossRef]
- Shattock, R.C. Phytophthora infestans: Populations, Pathogenicity and Phenylamides. Pest Manag. Sci. 2002, 58, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Gisi, U.; Cohen, Y. Resistance to Phenylamide Fungicides: A Case Study with Phytophthora infestans Involving Mating Type and Race Structure. Annu. Rev. Phytopathol. 1996, 34, 549–572. [Google Scholar] [CrossRef] [PubMed]
- Elansky, S.; Pobedinskaya, M.A.; Kokaeva, L.; Statsyuk, N.; Dyakov, Y.T. Phytophthora infestans Populations from the European Part of Russia: Genotypic Structure and Metalaxyl Resistance. J. Plant Pathol. 2015, 97, 449–456. [Google Scholar] [CrossRef]
- Troussieux, S.; Gilgen, A.; Souche, J.-L. A New Biocontrol Tool to Fight Potato Late Blight Based on Willaertia Magna C2c Maky Lysate. Plants 2022, 11, 2756. [Google Scholar] [CrossRef] [PubMed]
- Puidet, B.; Mabon, R.; Guibert, M.; Kiiker, R.; Soonvald, L.; Le, V.H.; Eikemo, H.; Dewaegeneire, P.; Saubeau, G.; Chatot, C.; et al. Examining Phenotypic Traits Contributing to the Spread in Northern European Potato Crops of EU_41_A2, a New Clonal Lineage of Phytophthora infestans. Phytopathology 2022, 112, 414–421. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide Pesticide Usage and Its Impacts on Ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Miller, S.A.; Ferreira, J.P.; LeJeune, J.T. Antimicrobial Use and Resistance in Plant Agriculture: A One Health Perspective. Agriculture 2022, 12, 289. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial Plant Compounds, Extracts and Essential Oils: An Updated Review on Their Effects and Putative Mechanisms of Action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef]
- Devrnja, N.; Milutinović, M.; Savić, J. When Scent Becomes a Weapon—Plant Essential Oils as Potent Bioinsecticides. Sustainability 2022, 14, 6847. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural Sustainability and Intensive Production Practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Choudhary, S.; Yamini, N.R.; Yadav, S.K.; Kamboj, M.; Sharma, A. A Review: Pesticide Residue: Cause of Many Animal Health Problems. J. Entomol. Zool. Stud. 2018, 6, 330–333. [Google Scholar]
- Tsedaley, B. Late Blight of Potato (Phytophthora infestans) Biology, Economic Importance and Its Management Approaches. J. Biol. Agric. Healthc. 2014, 4, 215–225. [Google Scholar]
- Forbes, G.A. Using Host Resistance to Manage Potato Late Blight with Particular Reference to Developing Countries. Potato Res. 2012, 55, 205–216. [Google Scholar] [CrossRef]
- Runno-Paurson, E.; Williams, I.H.; Metspalu, L.; Kaart, T.; Mänd, M. Current Potato Varieties Are Too Susceptible to Late Blight to Be Grown without Chemical Control under North-East European Conditions. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2013, 63, 80–88. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Boonekamp, P.M.; Hutten, R.; Jacobsen, E.; Lotz, L.A.P.; Kessel, G.J.T.; Visser, R.G.F.; van der Vossen, E.A.G. Societal Costs of Late Blight in Potato and Prospects of Durable Resistance Through Cisgenic Modification. Potato Res. 2008, 51, 47–57. [Google Scholar] [CrossRef]
- Kefelegn, H.; Chala, A.; Kassa, B.; Pananjay, G.; Tiwari, K. Evaluation of Different Potato Variety and Fungicide Combinations for the Management of Potato Late Blight (Phytophthora infestans) in Southern Ethiopia. Int. J. Life Sci. 2012, 1, 8–15. [Google Scholar]
- Mekonen, S.; Tadesse, T. Effect of Varieties and Fungicides on Potato Late Blight (Phytophthora infestans, (Mont.) de Bary) Management. Agrotechnology 2018, 7, 1–4. [Google Scholar] [CrossRef]
- Tähtjärv, T.; Tsahkna, A.; Tamm, S. Comparison of Late Blight Resistance and Yield of Potato Varieties. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2013, 67, 254–258. [Google Scholar] [CrossRef]
- Gedlu, D.; Hailu, N.; Kefelegn, H. Integrated Management of Potato Late Blight (Phytophthora infestans (Mont) de Bary) through Resistant Varieties and Fungicides in North Shewa, Ethiopia. J. Plant Pathol. 2023, 105, 95–106. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef]
- What Is Biocontrol; IBMA-GLOBAL: Brussels, Belgium, 2023; Available online: https://ibma-global.org/what-is-biocontrol (accessed on 17 July 2023).
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for Unifying the Terminology in Biological Control. BioControl 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Seiber, J.N.; Coats, J.; Duke, S.O.; Gross, A.D. Biopesticides: State of the Art and Future Opportunities. J. Agric. Food Chem. 2014, 62, 11613–11619. [Google Scholar] [CrossRef] [PubMed]
- Amichot, M.; Joly, P.; Martin-Laurent, F.; Siaussat, D.; Lavoir, A.-V. Biocontrol, New Questions for Ecotoxicology? Environ. Sci. Pollut. Res. 2018, 25, 33895–33900. [Google Scholar] [CrossRef]
- Thomas, M.B.; Willis, A.J. Biocontrol—Risky but Necessary? Trends Ecol. Evol. 1998, 13, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P.A. Screening for Novel Biocontrol Agents Applicable in Plant Disease Management—A Review. Biol. Control 2020, 144, 104240. [Google Scholar] [CrossRef]
- Reddy, P.P. Biological Control of Plant Pathogens. In Sustainable Crop Protection under Protected Cultivation; Reddy, P.P., Ed.; Springer: Singapore, 2016; pp. 61–69. ISBN 978-9-81287-952-3. [Google Scholar]
- Hashemi, M.; Tabet, D.; Sandroni, M.; Benavent-Celma, C.; Seematti, J.; Andersen, C.B.; Grenville-Briggs, L.J. The Hunt for Sustainable Biocontrol of Oomycete Plant Pathogens, a Case Study of Phytophthora infestans. Fungal Biol. Rev. 2022, 40, 53–69. [Google Scholar] [CrossRef]
- Stenberg, J.A.; Sundh, I.; Becher, P.G.; Björkman, C.; Dubey, M.; Egan, P.A.; Friberg, H.; Gil, J.F.; Jensen, D.F.; Jonsson, M.; et al. When Is It Biological Control? A Framework of Definitions, Mechanisms, and Classifications. J. Pest Sci. 2021, 94, 665–676. [Google Scholar] [CrossRef]
- Caulier, S.; Gillis, A.; Colau, G.; Licciardi, F.; Liépin, M.; Desoignies, N.; Modrie, P.; Legrève, A.; Mahillon, J.; Bragard, C. Versatile Antagonistic Activities of Soil-Borne Bacillus Spp. and Pseudomonas Spp. against Phytophthora infestans and Other Potato Pathogens. Front. Microbiol. 2018, 9, 143. [Google Scholar] [CrossRef]
- Najdabbasi, N.; Mirmajlessi, S.M.; Dewitte, K.; Ameye, M.; Mänd, M.; Audenaert, K.; Landschoot, S.; Haesaert, G. Green Leaf Volatile Confers Management of Late Blight Disease: A Green Vaccination in Potato. J. Fungi 2021, 7, 312. [Google Scholar] [CrossRef]
- Gfeller, A.; Fuchsmann, P.; De Vrieze, M.; Gindro, K.; Weisskopf, L. Bacterial Volatiles Known to Inhibit Phytophthora infestans Are Emitted on Potato Leaves by Pseudomonas Strains. Microorganisms 2022, 10, 1510. [Google Scholar] [CrossRef] [PubMed]
- Alfiky, A.; L’Haridon, F.; Abou-Mansour, E.; Weisskopf, L. Disease Inhibiting Effect of Strain Bacillus Subtilis EG21 and Its Metabolites against Potato Pathogens Phytophthora infestans and Rhizoctonia solani. Phytopathology 2022, 112, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.; Ficke, A.; Asiimwe, T.; Höfte, M.; Raaijmakers, J.M. Role of the Cyclic Lipopeptide Massetolide A in Biological Control of Phytophthora infestans and in Colonization of Tomato Plants by Pseudomonas Fluorescens. New Phytol. 2007, 175, 731–742. [Google Scholar] [CrossRef] [PubMed]
- De Vrieze, M.; Germanier, F.; Vuille, N.; Weisskopf, L. Combining Different Potato-Associated Pseudomonas Strains for Improved Biocontrol of Phytophthora infestans. Front. Microbiol. 2018, 9, 2573. [Google Scholar] [CrossRef] [PubMed]
- Aqil, F.; Zahin, M.; Ahmad, I.; Owais, M.; Khan, M.S.A.; Bansal, S.S.; Farooq, S. Antifungal Activity of Medicinal Plant Extracts and Phytocompounds: A Review. In Combating Fungal Infections: Problems and Remedy; Ahmad, I., Owais, M., Shahid, M., Aqil, F., Eds.; Springer: Berlin, Germany, 2010; pp. 449–484. ISBN 978-3-64212-173-9. [Google Scholar]
- Kumar, J.; Ramlal, A.; Mallick, D.; Mishra, V. An Overview of Some Biopesticides and Their Importance in Plant Protection for Commercial Acceptance. Plants 2021, 10, 1185. [Google Scholar] [CrossRef]
- Abdelgaleil, S.; Zoghroban, A.; El-Bakry, A.; Shehata, M. Insecticidal and Antifungal Activities of Crude Extracts and Pure Compounds from Rhizomes of Curcuma longa L. (Zingiberaceae). J. Agric. Sci. Technol. A 2019, 21, 1049–1061. [Google Scholar]
- Dakole, C.; Nguefack, J.; Dongmo Lekagne, J.B.; Joseph Hubert, G.; Rene, A.U.; Somda, I.; Henry, A. Antifungal Potential of Essential Oils, Aqueous and Ethanol Extracts of Thirteen Plants against Fusarium oxysporum f. Sp Lycopersici and Phytophtora infestans (Mont.) de Bary as Major Tomato Pathogens in Cameroon. Int. J. Curr. Sci. 2016, 19, 128–145. [Google Scholar]
- Hubert, G.Y.J.; Julienne, N.; Charles, D.D.; Daniel, F.; Sandrine, P.T.; Romain, F.F.; Henry, A.Z. Antifungal Potential and Phytochemical Analysis of Extracts from Seven Cameroonian Plants against Late Blight Pathogen. Phytophthora Infestans 2013, 2, 140–154. [Google Scholar]
- Jacquin, J.; Moureu, S.; Deweer, C.; Hakem, A.; Paguet, A.-S.; Bonneau, N.; Bordage, S.; Dermont, C.; Sahpaz, S.; Muchembled, J.; et al. Hop (Humulus lupulus L.) Specialized Metabolites: Extraction, Purification, Characterization in Different Plant Parts and In Vitro Evaluation of Anti-Oomycete Activities against Phytophthora infestans. Agronomy 2022, 12, 2826. [Google Scholar] [CrossRef]
- Ndala, R.; Mbega, E.; Ndakidemi, P. Different Plant Extracts against Phytophthora infestans (Mont.) de Bary in Tomato in Vitro. Am. J. Plant Sci. 2019, 10, 698–708. [Google Scholar] [CrossRef][Green Version]
- Bálint, J.; Turóczi, B.; Máthé, I.; Benedek, K.; Szabó, K.-A.; Adalbert, B. In Vitro and In Vivo Effect of Poplar Bud (Populi Gemma) Extracts on Late Blight (Phytophthora infestans). Acta Univ. Sapientiae Agric. Environ. 2014, 6, 1–8. [Google Scholar] [CrossRef]
- Turóczi, B.; Bakonyi, J.; Szabó, K.-A.; Bálint, J.; Máthé, I.; Lányi, S.; Balog, A. In Vitro and In Vivo Effect of Poplar Bud Extracts on Phytophthora infestans: A New Effective Biological Method in Potato Late Blight Control. Plants 2020, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Fometu, S.; Shittu, S.; Herman, R.; Ayepa, E. Essential Oils and Their Applications—A Mini Review. Adv. Nutr. Food Sci. 2019, 4, 1–13. [Google Scholar]
- Masango, P. Cleaner Production of Essential Oils by Steam Distillation. J. Clean. Prod. 2005, 13, 833–839. [Google Scholar] [CrossRef]
- Gavahian, M.; Chu, Y.-H.; Mousavi Khaneghah, A. Recent Advances in Orange Oil Extraction: An Opportunity for the Valorisation of Orange Peel Waste a Review. Int. J. Food Sci. Technol. 2019, 54, 925–932. [Google Scholar] [CrossRef]
- Maes, C.; Bouquillon, S.; Fauconnier, M.-L. Encapsulation of Essential Oils for the Development of Biosourced Pesticides with Controlled Release: A Review. Molecules 2019, 24, 2539. [Google Scholar] [CrossRef]
- Slavković, F.; Bendahmane, A. Floral Phytochemistry: Impact of Volatile Organic Compounds and Nectar Secondary Metabolites on Pollinator Behavior and Health. Chem. Biodivers. 2023, 20, e202201139. [Google Scholar] [CrossRef]
- De Clerck, C.; Genva, M.; Jijakli, M.H.; Fauconnier, M.-L. Use of Essential Oils and Volatile Compounds as Biological Control Agents. Foods 2021, 10, 1062. [Google Scholar] [CrossRef]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of Plant Secondary Metabolites: A Historical Perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of Essential Oils in Food Safety: Antimicrobial and Antioxidant Applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Preedy, V.R. Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2015; ISBN 978-0-12416-644-8. [Google Scholar]
- Assadpour, E.; Can Karaça, A.; Fasamanesh, M.; Mahdavi, S.A.; Shariat-Alavi, M.; Feng, J.; Kharazmi, M.S.; Rehman, A.; Jafari, S.M. Application of Essential Oils as Natural Biopesticides; Recent Advances. Crit. Rev. Food Sci. Nutr. 2023, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Bassolé, I.H.N.; Juliani, H.R. Essential Oils in Combination and Their Antimicrobial Properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, M.L. The Mode of Antibacterial Action of Essential Oils. Sci. Against Microb. Pathog. Commun. Curr. Res. Technol. Adv. 2011, 2, 1143–1156. [Google Scholar]
- Fenibo, E.O.; Ijoma, G.N.; Matambo, T. Biopesticides in Sustainable Agriculture: A Critical Sustainable Development Driver Governed by Green Chemistry Principles. Front. Sustain. Food Syst. 2021, 5, 619058. [Google Scholar] [CrossRef]
- Najdabbasi, N.; Mirmajlessi, S.M.; Dewitte, K.; Landschoot, S.; Mänd, M.; Audenaert, K.; Ameye, M.; Haesaert, G. Biocidal Activity of Plant-Derived Compounds against Phytophthora infestans: An Alternative Approach to Late Blight Management. Crop Prot. 2020, 138, 105315. [Google Scholar] [CrossRef]
- Soylu, E.M.; Soylu, S.; Kurt, S. Antimicrobial Activities of the Essential Oils of Various Plants against Tomato Late Blight Disease Agent Phytophthora infestans. Mycopathologia 2006, 161, 119–128. [Google Scholar] [CrossRef]
- Quintanilla, P.; Rohloff, J.; Iversen, T.-H. Influence of Essential Oils on Phytophthora infestans. Potato Res. 2002, 45, 225–235. [Google Scholar] [CrossRef]
- Aksit, H.; Bayar, Y.; Simsek, S.; Ulutas, Y. Chemical Composition and Antifungal Activities of the Essential Oils of Thymus Species (Thymus Pectinatus, Thymus Convolutus, Thymus Vulgaris) Against Plant Pathogens. J. Essent. Oil Bear. Plants 2022, 25, 200–207. [Google Scholar] [CrossRef]
- Carrillo, Y.A.; Gòmez, M.I.; Cotes, J.M.; Ñustez, C.E. Effect of Some Essential Oils on the Growth of Phytophthora infestans (Mont.) de Bary under Laboratory Conditions. Agron. Colomb. 2010, 28, 245–253. [Google Scholar]
- Deweer, C.; Sahmer, K.; Muchembled, J. Anti-Oomycete Activities from Essential Oils and Their Major Compounds on Phytophthora infestans. Environ. Sci. Pollut. Res. 2023. [Google Scholar] [CrossRef]
- Lu, M.; Han, Z.; Yao, L. In Vitro and in Vivo Antimicrobial Efficacy of Essential Oils and Individual Compounds against Phytophthora parasitica Var. Nicotianae. J. Appl. Microbiol. 2013, 115, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Jiang, H.; Hausbeck, M.K.; Hao, J.J. Inhibitory Effects of Essential Oils for Controlling Phytophthora Capsici. Plant Dis. 2012, 96, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Maissa, B.J.; Walid, H. Antifungal Activity of Chemically Different Essential Oils from Wild Tunisian Thymus Spp. Nat. Prod. Res. 2015, 29, 869–873. [Google Scholar] [CrossRef] [PubMed]
- Belay, D.W.; Asfaw, Z.; Molla, E.L.; Kassa, B.; Kifele, H. Evaluation of Essential Oils Against Potato Late Blight (Phytophthora infestans (Mont.) de Bary) at Holleta, Ethiopia. Turk. J. Agric.—Food Sci. Technol. 2022, 10, 1006–1018. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Zikeli, F.; Scarascia Mugnozza, G.; Vinciguerra, V.; Tabet, D.; Romagnoli, M. Lignin Nanoparticles Containing Essential Oils for Controlling Phytophthora Cactorum Diseases. For. Pathol. 2022, 52, e12739. [Google Scholar] [CrossRef]
- Thanh, V.M.; Bui, L.M.; Bach, L.G.; Nguyen, N.T.; Thi, H.L.; Hoang Thi, T.T. Origanum Majorana L. Essential Oil-Associated Polymeric Nano Dendrimer for Antifungal Activity against Phytophthora infestans. Materials 2019, 12, 1446. [Google Scholar] [CrossRef]
- Pitarokili, D.; Tzakou, O.; Loukis, A. Composition of the Essential Oil of Spontaneous Rosmarinus Officinalis from Greece and Antifungal Activity Against Phytopathogenic Fungi. J. Essent. Oil Res. 2008, 20, 457–459. [Google Scholar] [CrossRef]
- Amini, J.; Farhang, V.; Javadi, T.; Nazemi, J. Antifungal Effect of Plant Essential Oils on Controlling Phytophthora Species. Plant Pathol. J. 2016, 32, 16–24. [Google Scholar] [CrossRef]
- Kamsu, F.P.N.; Ndondoni Dikongue, F.J.; Ngouana, V.; Tchinda, E.S.; Jiogue, M.B.; Ambata, H.T.A.; Tchameni, S.N.; Sameza, M.L.; Dongmo Jazet, P.M. Effectiveness of Massep (Ocimum gratissimum L.) Essential Oil and Its Nanoemulsion toward Sclerotium Rolfsii, Phytophthora infestans and Alternaria solani, Pathogens Associated with Tomato Rot Diseases. Biocatal. Agric. Biotechnol. 2023, 47, 102591. [Google Scholar] [CrossRef]
- Al-Mughrabi, K.I.; Coleman, W.K.; Vikram, A.; Poirier, R.; Jayasuriya, K.E. Effectiveness of Essential Oils and Their Combinations with Aluminum Starch Octenylsuccinate on Potato Storage Pathogens. J. Essent. Oil Bear. Plants 2013, 16, 23–31. [Google Scholar] [CrossRef]
- Banihashemi, Z.; Abivardi, C. Evaluation of Fungicidal and Fungistatic Activity of Plant Essential Oils towards Plant Pathogenic and Saprophytic Fungi. Phytopathol. Mediterr. 2011, 50, 245–256. [Google Scholar]
- De Clerck, C.; Maso, S.D.; Parisi, O.; Dresen, F.; Zhiri, A.; Jijakli, M.H. Screening of Antifungal and Antibacterial Activity of 90 Commercial Essential Oils against 10 Pathogens of Agronomical Importance. Foods 2020, 9, 1418. [Google Scholar] [CrossRef]
- Mohammadi, A.; Hashemi, M.; Hosseini, S.M. Integration between Chitosan and Zataria Multiflora or Cinnamomum Zeylanicum Essential Oil for Controlling Phytophthora Drechsleri, the Causal Agent of Cucumber Fruit Rot. LWT—Food Sci. Technol. 2016, 65, 349–356. [Google Scholar] [CrossRef]
- Nana, W.L.; Eke, P.; Fokom, R.; Bakanrga-Via, I.; Begoude, D.; Tchana, T.; Tchameni, N.S.; Kuate, J.; Menut, C.; Fekam Boyom, F. Antimicrobial Activity of Syzygium Aromaticum and Zanthoxylum Xanthoxyloides Essential Oils Against Phytophthora Megakarya. J. Phytopathol. 2015, 163, 632–641. [Google Scholar] [CrossRef]
- Hong, Z.; Talib, K.M.; Mujtaba, K.G.; Dabin, H.; Yahya, F.; Congying, Z.; Fukai, W. Antifungal Potential of Cinnamon Essential Oils against Phytophthora Colocasiae Causing Taro Leaf Blight. Chem. Biol. Technol. Agric. 2021, 8, 39. [Google Scholar] [CrossRef]
- Mugao, L.G.; Muturi, P.W.; Gichimu, B.M.; Njoroge, E.K. In Vitro Control of Phytophthora infestans and Alternaria Solani Using Crude Extracts and Essential Oils from Selected Plants. Int. J. Agron. 2020, 2020, e8845692. [Google Scholar] [CrossRef]
- Wang, B.; Liu, F.; Li, Q.; Xu, S.; Zhao, X.; Xue, P.; Feng, X. Antifungal Activity of Zedoary Turmeric Oil against Phytophthora Capsici through Damaging Cell Membrane. Pestic. Biochem. Physiol. 2019, 159, 59–67. [Google Scholar] [CrossRef]
- Mugao, L.G.; Gichimu, B.M.; Muturi, P.W.; Njoroge, E.K. Essential Oils as Biocontrol Agents of Early and Late Blight Diseases of Tomato under Greenhouse Conditions. Int. J. Agron. 2021, 2021, e5719091. [Google Scholar] [CrossRef]
- Kalhoro, M.T.; Zhang, H.; Kalhoro, G.M.; Wang, F.; Chen, T.; Faqir, Y.; Nabi, F. Fungicidal Properties of Ginger (Zingiber Officinale) Essential Oils against Phytophthora Colocasiae. Sci. Rep. 2022, 12, 2191. [Google Scholar] [CrossRef]
- Han, X.-B.; Zhao, J.; Cao, J.-M.; Zhang, C.-S. Essential Oil of Chrysanthemum indicum L.: Potential Biocontrol Agent against Plant Pathogen Phytophthora Nicotianae. Environ. Sci. Pollut. Res. 2019, 26, 7013–7023. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, K.; Han, X.; Zhao, D.; Zheng, Y.; Chao, J.; Gou, J.; Kong, F.; Zhang, C.-S. The Antifungal Effect of Garlic Essential Oil on Phytophthora Nicotianae and the Inhibitory Component Involved. Biomolecules 2019, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- Messgo-Moumene, S.; Li, Y.; Bachir, K.; Houmani, Z.; Bouznad, Z.; Chemat, F. Antifungal Power of Citrus Essential Oils against Potato Late Blight Causative Agent. J. Essent. Oil Res. 2015, 27, 169–176. [Google Scholar] [CrossRef]
- El-Gamal, N.G. Effect of Orange Essential Oil and Its Constitutes on Late Blight Disease of Potato Plants under Field Conditions. Arch. Phytopathol. Plant Prot. 2011, 44, 260–268. [Google Scholar] [CrossRef]
- Tchameni, S.N.; Mbiakeu, S.N.; Sameza, M.L.; Jazet, P.M.D.; Tchoumbougnang, F. Using Citrus Aurantifolia Essential Oil for the Potential Biocontrol of Colocasia Esculenta (Taro) Leaf Blight Caused by Phytophthora Colocasiae. Environ. Sci. Pollut. Res. Int. 2018, 25, 29929–29935. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Q.; Li, L.; Li, P.; Yin, M.; Xu, S.; Chen, Y.; Feng, X.; Wang, B. Chemical Composition and Antifungal Activity of Zanthoxylum Armatum Fruit Essential Oil against Phytophthora Capsici. Molecules 2022, 27, 8636. [Google Scholar] [CrossRef]
- Wang, B.; Li, P.; Yang, J.; Yong, X.; Yin, M.; Chen, Y.; Feng, X.; Wang, Q. Inhibition Efficacy of Tetradium Glabrifolium Fruit Essential Oil against Phytophthora Capsici and Potential Mechanism. Ind. Crops Prod. 2022, 176, 114310. [Google Scholar] [CrossRef]
- Snoussi, A.; Koubaier, H.; Bouacida, S.; Essaidi, I.; Kachouri, F.; Bouzouita, N. In Vitro Antimicrobial Activity of Carum Carvi L. Seed Essential Oil against Pink Potato Spoilage Flora. Chem. Naissensis 2020, 3, 92–102. [Google Scholar] [CrossRef]
- Benlembarek, K.; Lograda, T.; Ramdani, M.; Figueredo, G.; Chalard, P. Chemical Composition and Biological Activities of Anethum graveolens L. Essential Oil from Algeria. J. Essent. Oil Bear. Plants 2022, 25, 728–740. [Google Scholar] [CrossRef]
- Rubio-Covarrubias, O.A.; Douches, D.S.; Hammerschmidt, R.; daRocha, A.; Kirk, W.W. Effect of Temperature and Photoperiod on Symptoms Associated with Resistance to Phytophthora infestans after Leaf Penetration in Susceptible and Resistant Potato Cultivars. Am. J. Potato Res. 2005, 82, 139–146. [Google Scholar] [CrossRef]
- Odilbekov, F.; Carlson-Nilsson, U.; Liljeroth, E. Phenotyping Early Blight Resistance in Potato Cultivars and Breeding Clones. Euphytica 2014, 197, 87–97. [Google Scholar] [CrossRef]
- Mihovilovich, E.; Munive, S.; Bonierbale, M. Influence of Day-Length and Isolates of Phytophthora infestans on Field Resistance to Late Blight of Potato. Theor. Appl. Genet. 2010, 120, 1265–1278. [Google Scholar] [CrossRef] [PubMed]
- Colon, L.T.; Turkensteen, L.J.; Prummel, W.; Budding, D.J.; Hoogendoorn, J. Durable Resistance to Late Blight (Phytophthora infestans) in Old Potato Cultivars. Eur. J. Plant Pathol. 1995, 101, 387–397. [Google Scholar] [CrossRef]
- Orłowska, E.; Fiil, A.; Kirk, H.-G.; Llorente, B.; Cvitanich, C. Differential Gene Induction in Resistant and Susceptible Potato Cultivars at Early Stages of Infection by Phytophthora infestans. Plant Cell Rep. 2012, 31, 187–203. [Google Scholar] [CrossRef]
- Olanya, O.M.; Larkin, R.P. Efficacy of Essential Oils and Biopesticides on Phytophthora infestans Suppression in Laboratory and Growth Chamber Studies. Biocontrol Sci. Technol. 2006, 16, 901–917. [Google Scholar] [CrossRef]
- Gisi, U.; Walder, F.; Resheat-Eini, Z.; Edel, D.; Sierotzki, H. Changes of Genotype, Sensitivity and Aggressiveness in Phytophthora infestans Isolates Collected in European Countries in 1997, 2006 and 2007. J. Phytopathol. 2011, 159, 223–232. [Google Scholar] [CrossRef]
- Slusarenko, A.J.; Patel, A.; Portz, D. Control of Plant Diseases by Natural Products: Allicin from Garlic as a Case Study. Eur. J. Plant Pathol. 2008, 121, 313–322. [Google Scholar] [CrossRef]
- Lubbe, A.; Verpoorte, R. Cultivation of Medicinal and Aromatic Plants for Specialty Industrial Materials. Ind. Crops Prod. 2011, 34, 785–801. [Google Scholar] [CrossRef]
- Cavanagh, H. Antifungal Activity of the Volatile Phase of Essential Oils: A Brief Review. Nat. Prod. Commun. 2007, 2, 1297–1302. [Google Scholar] [CrossRef]
- Werrie, P.-Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.-L. Phytotoxicity of Essential Oils: Opportunities and Constraints for the Development of Biopesticides. A Review. Foods 2020, 9, 1291. [Google Scholar] [CrossRef]
- Sangwan, N.S.; Farooqi, A.H.A.; Shabih, F.; Sangwan, R.S. Regulation of Essential Oil Production in Plants. Plant Growth Regul. 2001, 34, 3–21. [Google Scholar] [CrossRef]
- Moghaddam, M.; Mehdizadeh, L. Chapter 13—Chemistry of Essential Oils and Factors Influencing Their Constituents. In Soft Chemistry and Food Fermentation; Grumezescu, A.M., Holban, A.M., Eds.; Handbook of Food Bioengineering; Academic Press: Cambridge, MA, USA, 2017; pp. 379–419. ISBN 978-0-12811-412-4. [Google Scholar]
- Rehman, R.; Hanif, M.A.; Mushtaq, Z.; Al-Sadi, A.M. Biosynthesis of Essential Oils in Aromatic Plants: A Review. Food Rev. Int. 2016, 32, 117–160. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.; Wink, M.; Gershenzon, J. Biochemistry of Terpenoids: Monoterpenes, Sesquiterpenes and Diterpenes. In Annual Plant Reviews Volume 40: Biochemistry of Plant Secondary Metabolism; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 258–303. ISBN 978-1-44432-050-3. [Google Scholar]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.-J.; Reddy, M.S.S.; Wang, L. The Phenylpropanoid Pathway and Plant Defence—A Genomics Perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef] [PubMed]
- De Cássia da Silveira e Sá, R.; Andrade, L.N.; Dos Reis Barreto de Oliveira, R.; De Sousa, D.P. A Review on Anti-Inflammatory Activity of Phenylpropanoids Found in Essential Oils. Molecules 2014, 19, 1459–1480. [Google Scholar] [CrossRef] [PubMed]
- Barra, A. Factors Affecting Chemical Variability of Essential Oils: A Review of Recent Developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.; Draxler, L.; Göhler, I.; Franz, C.M. Essential Oil Composition of Vitex Agnus-Castus—Comparison of Accessions and Different Plant Organs. Flavour Fragr. J. 2005, 20, 186–192. [Google Scholar] [CrossRef]
- Boira, H.; Blanquer, A. Environmental Factors Affecting Chemical Variability of Essential Oils in Thymus piperella L. Biochem. Syst. Ecol. 1998, 26, 811–822. [Google Scholar] [CrossRef]
- Melito, S.; Petretto, G.L.; Podani, J.; Foddai, M.; Maldini, M.; Chessa, M.; Pintore, G. Altitude and Climate Influence Helichrysum Italicum Subsp. Microphyllum Essential Oils Composition. Ind. Crops Prod. 2016, 80, 242–250. [Google Scholar] [CrossRef]
- Llorens-Molina, J.A.; Vacas, S. Effect of Drought Stress on Essential Oil Composition of Thymus vulgaris L. (Chemotype 1, 8-Cineole) from Wild Populations of Eastern Iberian Peninsula. J. Essent. Oil Res. 2017, 29, 145–155. [Google Scholar] [CrossRef]
- Najar, B.; Pistelli, L.; Ferri, B.; Angelini, L.G.; Tavarini, S. Crop Yield and Essential Oil Composition of Two Thymus Vulgaris Chemotypes along Three Years of Organic Cultivation in a Hilly Area of Central Italy. Molecules 2021, 26, 5109. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Letchamo, W.; Ward, W.; Heard, B.; Heard, D. Essential Oil of Valeriana officinalis L. Cultivars and Their Antimicrobial Activity As Influenced by Harvesting Time under Commercial Organic Cultivation. J. Agric. Food Chem. 2004, 52, 3915–3919. [Google Scholar] [CrossRef] [PubMed]
- Zouari, N. Essential Oils Chemotypes: A Less Known Side. Med. Aromat. Plants 2013, 2, 1000.e145. [Google Scholar] [CrossRef]
- Kaloustian, J.; Abou, L.; Mikail, C.; Amiot, M.J.; Portugal, H. Southern French Thyme Oils: Chromatographic Study of Chemotypes. J. Sci. Food Agric. 2005, 85, 2437–2444. [Google Scholar] [CrossRef]
- Lis-Balchin, M.; Deans, S.G.; Eaglesham, E. Relationship between Bioactivity and Chemical Composition of Commercial Essential Oils. Flavour Fragr. J. 1998, 13, 98–104. [Google Scholar] [CrossRef]
- Marriott, P.J.; Shellie, R.; Cornwell, C. Gas Chromatographic Technologies for the Analysis of Essential Oils. J. Chromatogr. A 2001, 936, 1–22. [Google Scholar] [CrossRef]
- Pérez-Izquierdo, C.; Serrano-Pérez, P.; del Carmen Rodríguez-Molina, M. Chemical Composition, Antifungal and Phytotoxic Activities of Cistus Ladanifer L. Essential Oil and Hydrolate. Biocatal. Agric. Biotechnol. 2022, 45, 102527. [Google Scholar] [CrossRef]
- Dan, Y.; Liu, H.-Y.; Gao, W.-W.; Chen, S.-L. Activities of Essential Oils from Asarum heterotropoides Var. Mandshuricum against Five Phytopathogens. Crop Prot. 2010, 29, 295–299. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic Effects and Mechanism of Action of Essential Oils and Terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef]
- Taheri, P.; Soweizy, M.; Tarighi, S. Application of Essential Oils to Control Some Important Fungi and Bacteria Pathogenic on Cereals. J. Nat. Pestic. Res. 2023, 6, 100052. [Google Scholar] [CrossRef]
- Samuels, L.; Kunst, L.; Jetter, R. Sealing Plant Surfaces: Cuticular Wax Formation by Epidermal Cells. Annu. Rev. Plant Biol. 2008, 59, 683–707. [Google Scholar] [CrossRef] [PubMed]
- Cristani, M.; D’Arrigo, M.; Mandalari, G.; Castelli, F.; Sarpietro, M.G.; Micieli, D.; Venuti, V.; Bisignano, G.; Saija, A.; Trombetta, D. Interaction of Four Monoterpenes Contained in Essential Oils with Model Membranes: Implications for Their Antibacterial Activity. J Agric. Food Chem. 2007, 55, 6300–6308. [Google Scholar] [CrossRef] [PubMed]
- Brennan, T.C.R.; Kr�mer, J.O.; Nielsen, L.K. Physiological and Transcriptional Responses of Saccharomyces Cerevisiae to D-Limonene Show Changes to the Cell Wall but Not to the Plasma Membrane. Appl. Environ. Microbiol. 2013, 79, 3590–3600. [Google Scholar] [CrossRef]
- Ziogas, B.N.; Markoglou, A.N.; Theodosiou, D.I.; Anagnostou, A.; Boutopoulou, S. A High Multi-Drug Resistance to Chemically Unrelated Oomycete Fungicides in Phytophthora infestans. Eur. J. Plant Pathol. 2006, 115, 283–292. [Google Scholar] [CrossRef]
- Mei, X.; Liu, Y.; Huang, H.; Du, F.; Huang, L.; Wu, J.; Li, Y.; Zhu, S.; Yang, M. Benzothiazole Inhibits the Growth of Phytophthora Capsici through Inducing Apoptosis and Suppressing Stress Responses and Metabolic Detoxification. Pestic. Biochem. Physiol. 2019, 154, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Ben Jabeur, M.; Somai-Jemmali, L.; Hamada, W. Thyme Essential Oil as an Alternative Mechanism: Biofungicide-causing Sensitivity of Mycosphaerella Graminicola. J. Appl. Microbiol. 2017, 122, 932–939. [Google Scholar] [CrossRef]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A Comprehensive Review of the Antibacterial, Antifungal and Antiviral Potential of Essential Oils and Their Chemical Constituents against Drug-Resistant Microbial Pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Liang, J.; Wu, L.; Gao, W.; Jiang, J. Iturin A Extracted From Bacillus Subtilis WL-2 Affects Phytophthora infestans via Cell Structure Disruption, Oxidative Stress, and Energy Supply Dysfunction. Front. Microbiol. 2020, 11, 536083. [Google Scholar] [CrossRef]
- Yu, L.M. Elicitins from Phytophthora and Basic Resistance in Tobacco. Proc. Natl. Acad. Sci. USA 1995, 92, 4088–4094. [Google Scholar] [CrossRef]
- Vandana, V.V.; Suseela Bhai, R.; Ramakrishnan Nair, R.; Azeez, S. Role of Cell Wall and Cell Membrane Integrity in Imparting Defense Response against Phytophthora Capsici in Black Pepper (Piper nigrum L.). Eur. J. Plant Pathol. 2019, 154, 359–375. [Google Scholar] [CrossRef]
- Jing, C.; Gou, J.; Han, X.; Wu, Q.; Zhang, C. In Vitro and in Vivo Activities of Eugenol against Tobacco Black Shank Caused by Phytophthora Nicotianae. Pestic. Biochem. Physiol. 2017, 142, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of Terpenes and Terpenoids Present in Essential Oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Zhong, W.; Yang, L.; He, M.; Jiang, S.; Yin, D.; Guo, J.; Gao, Z. In Vitro and in Vivo Anti-Oomycetes Activities and Mechanisms of Linalool against Saprolegnia Ferax. Aquaculture 2024, 578, 740031. [Google Scholar] [CrossRef]
- Gao, T.; Zhou, H.; Zhou, W.; Hu, L.; Chen, J.; Shi, Z. The Fungicidal Activity of Thymol against Fusarium Graminearum via Inducing Lipid Peroxidation and Disrupting Ergosterol Biosynthesis. Molecules 2016, 21, 770. [Google Scholar] [CrossRef]
- Zhang, J.; Hao, Y.; Lu, H.; Li, P.; Chen, J.; Shi, Z.; Xie, Y.; Mo, H.; Hu, L. Nano-Thymol Emulsion Inhibits Botrytis Cinerea to Control Postharvest Gray Mold on Tomato Fruit. Agronomy 2022, 12, 2973. [Google Scholar] [CrossRef]
- Pei, S.; Liu, R.; Gao, H.; Chen, H.; Wu, W.; Fang, X.; Han, Y. Inhibitory Effect and Possible Mechanism of Carvacrol against Colletotrichum Fructicola. Postharvest Biol. Technol. 2020, 163, 111126. [Google Scholar] [CrossRef]
- Crowley, L.C.; Scott, A.P.; Marfell, B.J.; Boughaba, J.A.; Chojnowski, G.; Waterhouse, N.J. Measuring Cell Death by Propidium Iodide Uptake and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, 647–651. [Google Scholar] [CrossRef]
- Davis, D.J.; Burlak, C.; Money, N.P. Osmotic Pressure of Fungal Compatible Osmolytes. Mycol. Res. 2000, 104, 800–804. [Google Scholar] [CrossRef]
- Lippincott-Schwartz, J.; Phair, R.D. Lipids and Cholesterol as Regulators of Traffic in the Endomembrane System. Annu. Rev. Biophys. 2010, 39, 559–578. [Google Scholar] [CrossRef]
- Han, Y.; Sun, Z.; Chen, W. Antimicrobial Susceptibility and Antibacterial Mechanism of Limonene against Listeria Monocytogenes. Molecules 2020, 25, 33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Feng, R.; Li, L.; Zhou, X.; Li, Z.; Jia, R.; Song, X.; Zou, Y.; Yin, L.; He, C.; et al. The Antibacterial Mechanism of Terpinen-4-Ol Against Streptococcus Agalactiae. Curr. Microbiol. 2018, 75, 1214–1220. [Google Scholar] [CrossRef] [PubMed]
- Vila, R.; Freixa, B.; Cañigueral, S. Antifungal Compounds from Plants. In Phytochemical Resources for Medicine and Agriculture; Springer: Boston, MA, USA, 2013; Volume 3, pp. 23–43. ISBN 978-8-17895-605-3. [Google Scholar]
- Song, W.; Yin, Z.; Lu, X.; Shen, D.; Dou, D. Plant Secondary Metabolite Citral Interferes with Phytophthora Capsici Virulence by Manipulating the Expression of Effector Genes. Mol. Plant Pathol. 2023, 24, 932–946. [Google Scholar] [CrossRef] [PubMed]
- Ghalem, B.R. Essential Oils as Antimicrobial Agents against Some Important Plant Pathogenic Bacteria and Fungi. In Plant-Microbe Interaction: An Approach to Sustainable Agriculture; Choudhary, D.K., Varma, A., Tuteja, N., Eds.; Springer: Singapore, 2016; pp. 271–296. ISBN 978-9-81102-854-0. [Google Scholar]
- Ngegba, P.M.; Cui, G.; Khalid, M.Z.; Zhong, G. Use of Botanical Pesticides in Agriculture as an Alternative to Synthetic Pesticides. Agriculture 2022, 12, 600. [Google Scholar] [CrossRef]
- Turek, C.; Stintzing, F.C. Stability of Essential Oils: A Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]
- Lucia, A.; Guzmán, E. Emulsions Containing Essential Oils, Their Components or Volatile Semiochemicals as Promising Tools for Insect Pest and Pathogen Management. Adv. Colloid Interface Sci. 2021, 287, 102330. [Google Scholar] [CrossRef]
- Dunan, L.; Malanga, T.; Benhamou, S.; Papaiconomou, N.; Desneux, N.; Lavoir, A.-V.; Michel, T. Effects of Essential Oil-Based Formulation on Biopesticide Activity. Ind. Crops Prod. 2023, 202, 117006. [Google Scholar] [CrossRef]
- Pavoni, L.; Perinelli, D.R.; Bonacucina, G.; Cespi, M.; Palmieri, G.F. An Overview of Micro- and Nanoemulsions as Vehicles for Essential Oils: Formulation, Preparation and Stability. Nanomaterials 2020, 10, 135. [Google Scholar] [CrossRef]
- Tadros, T.; Izquierdo, P.; Esquena, J.; Solans, C. Formation and Stability of Nano-Emulsions. Adv. Colloid Interface Sci. 2004, 108–109, 303–318. [Google Scholar] [CrossRef]
- Anjali, C.H.; Sharma, Y.; Mukherjee, A.; Chandrasekaran, N. Neem Oil (Azadirachta indica) Nanoemulsion—A Potent Larvicidal Agent against Culex Quinquefasciatus. Pest Manag. Sci. 2012, 68, 158–163. [Google Scholar] [CrossRef]
- Wu, J.-E.; Lin, J.; Zhong, Q. Physical and Antimicrobial Characteristics of Thyme Oil Emulsified with Soluble Soybean Polysaccharide. Food Hydrocoll. 2014, 39, 144–150. [Google Scholar] [CrossRef]
- Sugumar, S.; Singh, S.; Mukherjee, A.; Chandrasekaran, N. Nanoemulsion of Orange Oil with Non Ionic Surfactant Produced Emulsion Using Ultrasonication Technique: Evaluating against Food Spoilage Yeast. Appl. Nanosci. 2016, 6, 113–120. [Google Scholar] [CrossRef]
- Tang, C.; Li, Y.; Pun, J.; Mohamed Osman, A.S.; Tam, K.C. Polydopamine Microcapsules from Cellulose Nanocrystal Stabilized Pickering Emulsions for Essential Oil and Pesticide Encapsulation. Colloids Surf. A Physicochem. Eng. Asp. 2019, 570, 403–413. [Google Scholar] [CrossRef]
- Rajkumar, V.; Gunasekaran, C.; Paul, C.A.; Dharmaraj, J. Development of Encapsulated Peppermint Essential Oil in Chitosan Nanoparticles: Characterization and Biological Efficacy against Stored-Grain Pest Control. Pestic. Biochem. Physiol. 2020, 170, 104679. [Google Scholar] [CrossRef] [PubMed]
- Campos, E.V.R.; Proença, P.L.F.; Oliveira, J.L.; Pereira, A.E.S.; de Morais Ribeiro, L.N.; Fernandes, F.O.; Gonçalves, K.C.; Polanczyk, R.A.; Pasquoto-Stigliani, T.; Lima, R.; et al. Carvacrol and Linalool Co-Loaded in β-Cyclodextrin-Grafted Chitosan Nanoparticles as Sustainable Biopesticide Aiming Pest Control. Sci. Rep. 2018, 8, 7623. [Google Scholar] [CrossRef] [PubMed]
- Ez-Zoubi, A.; Ez Zoubi, Y.; Bentata, F.; El-Mrabet, A.; Ben Tahir, C.; Labhilili, M.; Farah, A. Preparation and Characterization of a Biopesticide Based on Artemisia Herba-Alba Essential Oil Encapsulated with Succinic Acid-Modified Beta-Cyclodextrin. J. Chem. 2023, 2023, e3830819. [Google Scholar] [CrossRef]
- López, M.D.; Cantó-Tejero, M.; Pascual-Villalobos, M.J. New Insights Into Biopesticides: Solid and Liquid Formulations of Essential Oils and Derivatives. Front. Agron. 2021, 3, 763530. [Google Scholar] [CrossRef]
- Barradas, T.N.; de Holanda e Silva, K.G. Nanoemulsions of Essential Oils to Improve Solubility, Stability and Permeability: A Review. Environ. Chem. Lett. 2021, 19, 1153–1171. [Google Scholar] [CrossRef]
- Hardham, A.R. The Cell Biology behind Phytophthora Pathogenicity. Australas. Plant Pathol. 2001, 30, 91–98. [Google Scholar] [CrossRef]
- Isman, M.B. Plant Essential Oils for Pest and Disease Management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M. Targeting Pathogen Sterols: Defence and Counterdefence? PLoS Pathog. 2017, 13, e1006297. [Google Scholar] [CrossRef] [PubMed]
- Gaulin, E.; Bottin, A.; Dumas, B. Sterol Biosynthesis in Oomycete Pathogens. Plant Signal. Behav. 2010, 5, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, X.; Govers, F. The Mysterious Route of Sterols in Oomycetes. PLoS Pathog. 2021, 17, e1009591. [Google Scholar] [CrossRef]
- Wang, K.; Senthil-Kumar, M.; Ryu, C.-M.; Kang, L.; Mysore, K.S. Phytosterols Play a Key Role in Plant Innate Immunity against Bacterial Pathogens by Regulating Nutrient Efflux into the Apoplast. Plant Physiol. 2012, 158, 1789–1802. [Google Scholar] [CrossRef]
- Amborabé, B.-E.; Rossard, S.; Pérault, J.-M.; Roblin, G. Specific Perception of Ergosterol by Plant Cells. Comptes Rendus Biol. 2003, 326, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, P.; Srivastava, V.; Ekengren, S.; McKee, L.S.; Bulone, V. Comparative Analysis of Sterol Acquisition in the Oomycetes Saprolegnia Parasitica and Phytophthora infestans. PLoS ONE 2017, 12, e0170873. [Google Scholar] [CrossRef] [PubMed]
- Giunti, G.; Benelli, G.; Palmeri, V.; Laudani, F.; Ricupero, M.; Ricciardi, R.; Maggi, F.; Lucchi, A.; Guedes, R.N.C.; Desneux, N.; et al. Non-Target Effects of Essential Oil-Based Biopesticides for Crop Protection: Impact on Natural Enemies, Pollinators, and Soil Invertebrates. Biol. Control 2022, 176, 105071. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Cai, J.; Liu, D.; Yan, Y.; Zhang, H.; Li, L.; Wang, X.; Xiang, W.; Zhang, J. Discovery of Febrifugine with Specific Anti-Phytophthora Oomycete Activity Isolated from Dichroa Febrifuga Lour. Ind. Crops Prod. 2022, 178, 114651. [Google Scholar] [CrossRef]
- Stanley, J.; Preetha, G. Pesticide Toxicity to Microorganisms: Exposure, Toxicity and Risk Assessment Methodologies. In Pesticide Toxicity to Non-Target Organisms: Exposure, Toxicity and Risk Assessment Methodologies; Stanley, J., Preetha, G., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 351–410. ISBN 978-9-40177-752-0. [Google Scholar]
- Carlile, B. Pesticide Selectivity, Health and the Environment; Cambridge University Press: Cambridge, UK, 2006; ISBN 978-1-13945-756-9. [Google Scholar]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef]
- Michael, O.T.; Adebayo, O. Plant Essential Oil: An Alternative to Emerging Multidrug Resistant Pathogens. J. Microbiol. Exp. 2017, 5, 163–174. [Google Scholar] [CrossRef]
- Saltos-Rezabala, L.A.; Silveira, P.R.D.; Tavares, D.G.; Moreira, S.I.; Magalhães, T.A.; Botelho, D.M.D.S.; Alves, E. Thyme Essential Oil Reduces Disease Severity and Induces Resistance against Alternaria Linariae in Tomato Plants. Horticulturae 2022, 8, 919. [Google Scholar] [CrossRef]
- Kesraoui, S.; Andrés, M.F.; Berrocal-Lobo, M.; Soudani, S.; Gonzalez-Coloma, A. Direct and Indirect Effects of Essential Oils for Sustainable Crop Protection. Plants 2022, 11, 2144. [Google Scholar] [CrossRef] [PubMed]
- Rienth, M.; Crovadore, J.; Ghaffari, S.; Lefort, F. Oregano Essential Oil Vapour Prevents Plasmopara Viticola Infection in Grapevine (Vitis Vinifera) and Primes Plant Immunity Mechanisms. PLoS ONE 2019, 14, e0222854. [Google Scholar] [CrossRef] [PubMed]
- Lengnick, L. The Vulnerability of the US Food System to Climate Change. J. Environ. Stud. Sci. 2015, 5, 348–361. [Google Scholar] [CrossRef]
- Malik, A.; Suryapani, S.; Ahmad, J. Chemical Vs Organic Cultivation of Medicinal and Aromatic Plants: The Choice Is Clear. Int. J. Med. Aromat. Plants 2011, 1, 5–13. [Google Scholar]
- Balog, A.; Hartel, T.; Loxdale, H.D.; Wilson, K. Differences in the Progress of the Biopesticide Revolution between the EU and Other Major Crop-Growing Regions. Pest Manag. Sci. 2017, 73, 2203–2208. [Google Scholar] [CrossRef]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef]
- Gounaris, Y. Biotechnology for the Production of Essential Oils, Flavours and Volatile Isolates. A Review. Flavour Fragr. J. 2010, 25, 367–386. [Google Scholar] [CrossRef]
- Teper-Bamnolker, P.; Dudai, N.; Fischer, R.; Belausov, E.; Zemach, H.; Shoseyov, O.; Eshel, D. Mint Essential Oil Can Induce or Inhibit Potato Sprouting by Differential Alteration of Apical Meristem. Planta 2010, 232, 179–186. [Google Scholar] [CrossRef]
- Thoma, J.; Zheljazkov, V.D. Sprout Suppressants in Potato Storage: Conventional Options and Promising Essential Oils—A Review. Sustainability 2022, 14, 6382. [Google Scholar] [CrossRef]
- Kundu, R.; Dutta, D.; Nanda, M.K.; Chakrabarty, A. Near Real Time Monitoring of Potato Late Blight Disease Severity Using Field Based Hyperspectral Observation. Smart Agric. Technol. 2021, 1, 100019. [Google Scholar] [CrossRef]
- Laothawornkitkul, J.; Jansen, R.M.C.; Smid, H.M.; Bouwmeester, H.J.; Muller, J.; van Bruggen, A.H.C. Volatile Organic Compounds as a Diagnostic Marker of Late Blight Infected Potato Plants: A Pilot Study. Crop Prot. 2010, 29, 872–878. [Google Scholar] [CrossRef]
- Popp, J.; Pető, K.; Nagy, J. Pesticide Productivity and Food Security. A Review. Agron. Sustain. Dev. 2013, 33, 243–255. [Google Scholar] [CrossRef]
- Dassanayake, M.K.; Chong, C.H.; Khoo, T.-J.; Figiel, A.; Szumny, A.; Choo, C.M. Synergistic Field Crop Pest Management Properties of Plant-Derived Essential Oils in Combination with Synthetic Pesticides and Bioactive Molecules: A Review. Foods 2021, 10, 2016. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oil Origin | Tested | References | ||||
|---|---|---|---|---|---|---|
| Botanical Family | Vernacular Name | Plant Species | In Vitro Experiments (EO Concentration—Results Obtained) | In Vivo Experiments (EO Concentration—Results Obtained) | On Phytophthora | |
| Lamiaceae | Thyme | Thymus vulgaris | Sporangial germination on microplate (ED50 ≈ 0.3 µL/mL) Mycelium growth inhibition on Petri dish (80% MGI from 0.41 µL/mL) | Detached leave assays (DSI at 0% from 3.33 µL/mL) Greenhouse assays on potato plants (DSR = 80% at 3.33 µL/mL) | infestans | [93] |
| Fumigation test against mycelium growth (total inhibition from 0.3 µg/mL air) Contact test against mycelium growth (total inhibition from 6.4 µg/mL) Contact effect on sporangia production (Absence of sporangia from 1.6 µg/mL) | - | [94] | ||||
| Mycelium growth inhibition on Petri dish (90% inhibition CTC with 4 µL/plate after 22 days) | Greenhouse experiments on 2 potato cultivars (1:500 v/v—reduction of 30 and 40% of DSI CTC) | [95] | ||||
| Fumigation test (100% inhibition at 1 µL/Petri dish and LC50 = 0.467 µL/mL air) | - | [96] | ||||
| Fumigation test on mycelium growth (Mycelium area ≈ −85% CTC after 19 days) | - | [97] | ||||
| Sporangia development on microplate (IC50 = 99.41 mg/L) | - | [98] | ||||
| Mycelium growth inhibition on agar (Inhibition of 55% CTC at 100 ppm) | - | [24] | ||||
| Mycelium growth inhibition on Petri dish (95% inhibition at 144 and ED50 ≈ 70 mg/L) Sporangia development (Completely blocked from 72 mg/L) Zoospores production and germination (100% inhibited from 72 mg/mL) | - | parasitica | [99] | |||
| Mycelium growth inhibition (EC50 ≈ 0.14 µg/mL by contact and EC50 ≈ 0.11 µg/mL by fumigation) Sporangia and zoospores production ((EC50 ≈ 0.0475 µg/mL) Sporangia and zoospores germination (EC50 ≈ 0.095 µg/mL) | - | capsici | [100] | |||
| Thymus satureioides | Mycelium growth inhibition on agar (Inhibition of 80% CTC at 100 ppm) | - | infestans | [24] | ||
| Thymus convoltus | Fumigation test on Petri dish (60% inhibition at 4 µL/Petri after 7 days LC50 ND) | - | [96] | |||
| Thymus pectipatus | Fumigation test on Petri dish (100% inhibition CTC at 2 µL/Petri after 7 days, LC50 = 0.452 µL/mL air) | - | [96] | |||
| Thymus capitus | Antifungal activity on mycelium growth (IC50 = 107 µL/L) | - | [101] | |||
| Thymus algeriensis | Antifungal activity on mycelium growth (ND) | - | [101] | |||
| Thymus schimperi | - | On-field assays on 2 potato cultivars with ≠ levels of resistance (DSI equal to controls from 46 DAP) | [102] | |||
| Thymus serpyllum | Test by contact in Petri dish with EO encapsulated in lignin nanoparticles (LNP) (EC50 = 120 µg/mL for EO alone and EC50 = 88 µg/mL for EO-LNP) | Greenhouse tests on black pine plantlets (−20% mortality CTC with EO and no mortality with EO-LNP after 10 days) | cactorum | [103] | ||
| Oregano | Origanum vulgare | Fumigation test against mycelium growth (total inhibition from 0.3 µg/mL air) Contact test against mycelium growth (total inhibition from 6.4 µg/mL) Contact effect on sporangia production (Absence of sporangia from 0.8 µg/mL) | - | infestans | [94] | |
| Fumigation test on mycelium growth (Mycelium area ≈ −50% CTC after 19 days) | - | infestans | [97] | |||
| Mycelium growth inhibition on Petri dish (60% inhibition CTC with 4 µL/plate after 22 days) | - | [95] | ||||
| Mycelium growth inhibition on agar (Inhibition of 90% CTC at 100 ppm) | Potato plants in growth chamber on (25% disease suppression CTC at 0.2%) | [24] | ||||
| Origanum compactum | Sporangia development on microplate (IC50 = 96.5 mg/L) | - | [98] | |||
| Origanum syriacum | Mycelium growth inhibition (EC50 ≈ 0.07 µg/mL by contact and EC50 ≈ 0.09 µg/mL by fumigation) Sporangia and zoospores production (EC50 ≈ 0.0475 µg/mL) and germination (EC50 ≈ 0.095 µg/mL) | - | capsici | [100] | ||
| Origanum majorana | Fumigation test on mycelium growth (Mycelium area ≈ −35% CTC after 19 days) | - | infestans | [97] | ||
| Mycelium growth inhibition on agar (Inhibition EO < formulation EO + PANAM) | - | [104] | ||||
| Savory | Satureja montana | Sporangia development on microplate (IC50 = 74.65 mg/L) | - | [98] | ||
| Rosemary | Rosmarinus officinalis | Sporangial germination on microplate (ED50 ≈ 0.6 µL/mL) Mycelium growth inhibition on Petri dish (80% inhibition from 1.66 µL/mL) | Detached potato leaves assays (−30% DSI CTC from 1.66 µL/mL) Greenhouse assays on potato plants (DSR = 90% from 3.33 µL/mL) | [93] | ||
| Fumigation test on mycelium growth (Mycelium area ≈ −15% CTC after 19 days) | - | [97] | ||||
| Fumigation test against mycelium growth (total inhibition from 1.2 µg/mL air) Contact test against mycelium growth (total inhibition from 12.8 µg/mL) Contact effect on sporangia production (Absence of sporangia from 6.4 µg/mL) | - | [94] | ||||
| - | On-field assays with 2 potato cultivars presenting ≠ levels of resistance (DSI equal to the control from 46 DAP) | infestans | [102] | |||
| Mycelium growth inhibition on Petri dish (EC50 ≈ 172 µL/L) | - | nicotianae | [105] | |||
| Sage | Salvia officinalis | Fumigation test on mycelium growth Mycelium area ≈ −30% CTC after 19 days) | - | infestans | [97] | |
| Mycelium growth inhibition (EC50 ≈ 4.86 µg/mL by contact and EC50 ≈ 1.28 µg/mL by fumigation) | - | capsici | [100] | |||
| Basil | Ocinum basilicum | Fumigation test on mycelium growth (Mycelium area ≈ −50% CTC after 19 days) | - | infestans | [97] | |
| Mycelium growth inhibition (ED50 ≈ 120 mg/L) | - | parasitica | [99] | |||
| Mycelium growth inhibition on Petri dish (Total inhibition around 400 ppm for all three strains and EC50 ≈ 135, 200 and 191 ppm, respectively) | Assays in greenhouse on whole plants of pepper, cucumber and melon (DSI reduced by50, 36 and 44% CTC after 50 mL at 100 ppm applied on the roots of inoculated plants) | capsici dreshleris melonis | [106] | |||
| Massep | Ocinum gratissimum | Mycelium growth inhibition on Petri dish (Total inhibition up to 10 days at 300 µL/L with pure EO and at 250 µL/L with nano-emulsion) | Tests on artificially infected tomato fruits (Disease reduction of 47% and 100% with 900 µL/L of pure EO and nano-emulsion for preventive tests and of 100% with 900 µL/L of both treatments for curative) | infestans | [107] | |
| Mycelium growth inhibition on Petri dish (Total inhibition from 6250 µg/mL after 14 days) | - | [74] | ||||
| Pepper Mint | Mentha piperita | Mycelium growth inhibition on Petri dish (65% inhibition CTC with 4 µL/plate after 22 days) | Greenhouse experiments on 2 potato cultivars (1:500 v/v—reduction of 10 and 25% of DSI CTC over 22 days) | [95] | ||
| Fumigation test on mycelium growth (Mycelium area ≈ −85% CTC after 19 days) | - | infestans | [97] | |||
| Mycelium growth inhibition by fumigation (Total inhibition with 100 µL/Petri dish) | - | [108] | ||||
| Mycelium growth inhibition by contact (Total inhibition at 1 µL/mL for all spp.) Mycelium growth inhibition by fumigation (Total inhibition at 25 µL/L air for all spp.) | - | capsici melonis nicotianae cinnamoni citrophthora | [109] | |||
| Green mint | Mentha spicata | Sporangia development on microplate (IC50 = 130.56 mg/L) | - | infestans | [98] | |
| Mycelium growth inhibition by fumigation (Total inhibition with 100 µL/Petri dish) | - | [108] | ||||
| Mentha pulegium | Sporangial germination on microplate (Total inhibition after 120 h at 1000 ppm) | - | [110] | |||
| Lemon balm | Melissa officinalis | Mycelium growth inhibition on Petri dish (70% inhibition CTC 4 µL/plate at day 22) | - | [95] | ||
| Lavender | Lavendula officinalis | Fumigation test against mycelium growth (Total inhibition from 1.6 µg/mL air) Contact test against mycelium growth (Total inhibition from 25.6 µg/mL) Contact effect on sporangia production (Absence of sporangia from 6.4 µg/mL) | - | [94] | ||
| Mycelium growth inhibition on agar (100 ppm—inhibition of 20% CTC) | - | [24] | ||||
| Mycelium growth inhibition by contact (Total inhibition at 5 µL/mL for all spp.) Mycelium growth inhibition by fumigation (Total inhibition at 250 µL/L air for all spp.) | - | capsici melonis nicotianae cinnamoni citrophthora | [109] | |||
| Patchouli | Pogostemon cablin | Fumigation test on mycelium growth (Mycelium area ≈ −25% CTC after 19 days) | - | infestans | [97] | |
| Hyssop | Hyssopus officinalis | Mycelium growth inhibition on Petri dish (45% inhibition CTC with 4 µL/plate after 22 days) | Greenhouse experiments on 2 potato cultivars (1:500 v/v—reduction of 70 and 85% of DSI CTC) | infestans | [95] | |
| Zataria | Zataria multiflora | Mycelium growth inhibition on Petri dish in association with chitosan (CS) (EO IC50 = 0.039% and EO + CS IC50 = 0.011% | Assays on wounded cucumber fruits artificially infected (DSI −20% CTC with EO alone and −75% CTC with EO-CS after 7 days at 4 °C and then 2 days at 24°) | drechsleri | [111] | |
| Myrtaceae | Clove | Syzygium aromaticum | Sporangial germination on microplate (ED50 ≈ 4.5 µL/mL) Mycelium growth inhibition on Petri dish (80% MGI from 0.41 µL/mL) | Detached potato leaves assays (DSI −30% CTC from 3.33 µL/mL) Greenhouse assays on potato plants (DSR = 40% at 6.66 µL/mL) | infestans | [93] |
| Sporangia development on microplate (IC50 = 28.42 mg/L) | - | [98] | ||||
| Mycelium growth inhibition (Total inhibition from 250 µL/L) | Tests on cocoa pod husk pieces (DSI −70% CTC at 1000 µL/L after 2 weeks) | megakarya | [112] | |||
| Eucalyptus | Eucalyptus globulus | - | On-field assays on 2 potato cultivars with ≠ levels of resistance (DSI −25% on sensitive 53 DAP and DSI −33% on resistant cultivar 60 DAP) | infestans | [102] | |
| Eucalyptus citriodora | Sporangia development on microplate (IC50 = 122.11 mg/L) | - | [98] | |||
| Eucalyptus tereticornis | Mycelium growth inhibition on Petri dish (Total inhibition from 12,500 µg/mL after 14 days) | - | [74] | |||
| Tea tree | Melaleuca alternifolia | Mycelium growth inhibition (EC50 ≈ 3.59 µg/mL by contact and EC50 ≈ 10.07 µg/mL by fumigation) | - | capsici | [100] | |
| Sporangia development on microplate (IC50 = 476.37 mg/L) | - | infestans | [98] | |||
| Lauraceae | Laurel | Laurus nobilis | Fumigation test against mycelium growth (Total inhibition from 2.0 µg/mL air) Contact test against mycelium growth (Total inhibition from 51.2 µg/mL) Contact effect on sporangia production (Absence of sporangia from 12.8 µg/mL) | - | infestans | [94] |
| Cinnamon | Cinnamomum cassia | Sporangial germination on microplate (ED50 ≈ 0.5 µL/mL) Mycelium growth inhibition on Petri dish (80% MGI from 1.66 µL/mL) | Detached potato leaves assays (DSI −40% CTC at 6.66 µL/mL) Greenhouse assays on potato plants (DSR = 20% at 6.66 µL/mL) | [93] | ||
| Sporangial germination on microplates (Total inhibition after 120 h at 1000 ppm) | - | [110] | ||||
| Mycelium growth inhibition on Petri dish (Total inhibition at 72 and ED50 ≈ 40 mg/L) Sporangia and zoospores production (Completely blocked from 144 mg/L) Zoospores germination (Totally inhibited from 72 mg/mL) | - | parasitica. | [99] | |||
| Cinnamomum zeylanicum | Sporangial germination on microplate (Total inhibition after 120 h at 1000 ppm) | - | infestans | [110] | ||
| Mycelium growth inhibition on Petri dish in association with chitosan (CS) (EO IC50 = 0.039% and EO + CS IC50 = 0.011%) | Assays on wounded cucumber fruits (DSI −35% CTC with EO alone and −85% CTC with EO-CS after 7 days at 4 °C and then 2 days at 24°) | drechsleri | [111] | |||
| Mycelium growth inhibition (EC50 ≈ 0.19 µg/mL by contact and EC50 ≈ 0.28 µg/mL by fumigation) | - | capsici | [100] | |||
| Mycelium growth inhibition on Petri dish (Total inhibition from 0.625 mg/mL) Zoospores’ germination (Totally inhibited from 0.625 mg/mL) Sporangia production (Totally impeded from 1.25 mg/mL) | Effect on leaf necrosis and sporulation on taro aerial part leaves (Disease symptoms completely inhibited—leaf necrosis diameter = 0—and sporulation entirely blocked from 1.25 mg/mL) | colocasiae | [113] | |||
| Cupressaceae | Juniper | Juniperus communis | Sporangial germination on microplate (ED50 ≈ 2.3 µL/mL) Mycelium growth inhibition on Petri dish (30% MGI from 3.33 µL/mL) | Detached potato leaves assays (DSI −25% CTC from 3.33 µL/mL) Greenhouse assays on potato plants (DSR around 40% at 3.33 µL/mL) | infestans | [93] |
| Verbenaceae | Common lantana | Lantana camara | Mycelium growth on Petri dish (40% inhibition CTC at 2 mL/L after 7 days) | - | [114] | |
| Piperaceae | Pepper | Piper nigrum | Sporangial germination on microplates (ED50 ≈ 1.2 µL/mL) Mycelium growth inhibition on Petri dish (30% MGI at 6.66 µL/mL) | Detached leave assays (DSI −30% CTC from 6.66 µL/mL) Greenhouse assays on potato plants (DSI around 50% at 3.33 µL/mL) | [93] | |
| Zingiberaceae | Turmeric | Curcuma longa | Sporangial germination on microplates (ED50 ≈ 2.5 µL/mL) Mycelium growth inhibition on Petri dish (60% MGI from 3.33 µL/mL) | Detached leave assays (DSI −25% CTC from 3.33 µL/mL) Greenhouse assays on potato plants (DSR = 75% from 3.33 µL/mL) | [93] | |
| Curcuma phaeocaulis | Mycelium growth inhibition on Petri dish (EC50 = 4.9 µg/mL and EC90 = 34.3 µg/mL) | - | [115] | |||
| Mycelium growth inhibition on Petri dish (EC50 = 0.5 µg/mL and EC90 = 7.1 µg/mL) Investigation of activity against sporangial and zoospore production and germination (No sporangial nor zoospore production at 20 µg/mL and spores’ takes 4× more time to achieve germination at 20 µg/mL CTC) | Protective and curative assays on detached cucumber leaves (Control efficacy > 90% CTC for both preventive and curative activities from 100 µg/mL after 72 h of incubation) | capsici | [115] | |||
| Ginger | Zingiber officinalis | Mycelium growth on Petri dish (100% inhibition at 2 mL/L for 7 days) | Greenhouse assays on tomato plants (DSI −80% CTC after 10 weeks) | infestans | [114,116] | |
| Mycelium growth inhibition on Petri dish (Total inhibition from 1250 ppm) Inhibition of sporangia and zoospores (Total inhibition for at 625 ppm) | Assessment of necrosis on taro leaves (Diameter of necrosis ≈0 and no from 1250 ppm sporangia after 72 h) Reduction in symptoms on taro corms (−80% DSI CTC at 625 ppm after 7 days) | colocasiae | [117] | |||
| Asteraceae | Mexican marigold | Tagetes erecta | Mycelium growth inhibition on Petri dish (40% inhibition CTC at 2 mL/L after 7 days) | Greenhouse assays on tomato plants (DSI −80% CTC after 10 weeks) | infestans | [114,116] |
| Indian chrysanthemum | Chrysanthemum indicum | Mycelium growth inhibition on Petri dish (100% inhibition CTC from 200 µL/L) Spore germination (100% inhibition CTC from 200 µL/L) Fumigation test on mycelium growth (100% inhibition from 100 µL/L) | - | nicotianae | [118] | |
| Amaryllidaceae | Garlic | Allium sativum | - | On-field assays on 2 potato cultivars presenting ≠ levels of resistance (DSI −33% CTC 53 DAP on susceptible −33% CTC up to 60 DAP on resistant) | infestans | [102] |
| Mycelium growth on Petri dish (100% inhibition at 2 mL/L after 7 days) | Greenhouse assays on tomato plants (DSI −80% CTC after 10 weeks) | [114,116] | ||||
| Mycelium growth inhibition on Petri dish in DMSO 2% (EC50 ≈ 1 108 µL/L after 4 days) | On-pot experiments on tobacco roots (Disease control effect of 46% by root- irrigation at 1:500 v/v and of 49% by fumigation at 1:500 v/v) | nicotianae | [119] | |||
| Rutaceae | Lemon | Citrus limon | Mycelium growth inhibition on Petri dish (35% inhibition CTC after 7 days) Inhibition rate of sporulation (10% inhibition CTC at 1:100 v/v at day 21) | Inhibition of the infection on potato foliar discs after soaking in EO solutions at 3 dilution rates (Average inhibition of 5% CTC) | infestans | [120] |
| Orange | Citrus sinensis | Mycelium growth inhibition on Petri dish (No inhibition at all tested concentrations 1.15; 2.5; 5; 7.5 mL/L) | Greenhouse experiments (−80% DSI CTC for protective at 5 mL/L but no curative effect at 7.5 mL/L) | [121] | ||
| Mycelium growth inhibition on Petri dish (50% inhibition CTC after 7 days) Inhibition rate of sporulation (90% inhibition CTC at 1:100 v/v at day 21) | Inhibition of the infection on potato foliar discs after soaking in EO solutions at 3 dilution rates (Average inhibition of 65% CTC) | [120] | ||||
| Mycelium growth inhibition by contact (No inhibition for none of the spp. even at the highest tested concentration of 1 µL/mL) | - | capsici melonis nicotianae cinnamoni citrophthora | [109] | |||
| Bergamot | Citrus bergamia | Mycelium growth inhibition on Petri dish (55% inhibition CTC after 7 days) Inhibition rate of sporulation (50% inhibition CTC at 1:100 v/v at day 21) | Inhibition of the infection on potato foliar discs after soaking in EO solutions at 3 dilution rates (Average inhibition of 40% CTC) | infestans | [120] | |
| Lime | Citrus aurantifolia | Mycelium growth inhibition on Petri dish (MGI > 95% at 400 ppm after 7 days) Inhibition of sporangium production (−50% sporangia at 250 ppm CTC) | Necrosis inhibition tests on taro foliar discs (At 5000 ppm total necrosis inhibition for preventive and 50% for curative test) | colocasiae | [122] | |
| Bottle brush | Callistermon citrinus | Mycelium growth inhibition on Petri dish (Total inhibition from 312.5 µg/mL after 14 days) | - | infestans | [74] | |
| Prickly ash | Zanthoxylum armatum | Mycelium growth inhibition on Petri dish (Total inhibition at 2.5 µL/mL from 48 h) Investigation of activity against sporangial and zoospore production and germination (No sporangial nor zoospore production and germination at 2.0 µg/mL) | Protective and curative tests on pepper fruits (Control efficacy > 90% CTC for protective and 80% for curative at 200 µL/mL after 3 days of incubation) | capsici | [123] | |
| Zanthoxylum xanthoxyloides | Mycelium growth inhibition on Petri dish (Total inhibition from 350 µL/L) | Tests on cocoa pod husk pieces (DSI −64% CTC at 2000 µL/L after 2 weeks) | megakarya | [112] | ||
| / | Tetradium glabrifolium | Mycelium growth inhibition on Petri dish (Total inhibition at 20 mg/L up to 96 h) Activity against spores’ production (No spores produced at all at 20 mg/L) Inhibition of spores’ germination (3× more time to germinate at 10 mg/L CTC) | Activity test on detached pepper leaves (Efficacy ≈ 100% CTC at 500 mg/L for both protective and curative after 96 h) Activity test on pepper fruits (Efficacy ≈ 100% CTC at 500 mg/L for both protective and curative after 96 h) | capsici | [124] | |
| Poaceae | Lemon Grass | Cymbopogon nardus | Mycelium growth inhibition (EC50 ≈ 0.44 µg/mL by contact and EC50 ≈ 0.25 µg/mL by fumigation) | - | capsici | [100] |
| Cymbopogon flexuosus | Sporangial germination on microplates (Total inhibition for 120 h at 1000 ppm) | - | infestans | [110] | ||
| Cymbopogon citratus | Mycelium growth inhibition on Petri dish (Total inhibition around 72.5 ppm for all three strains and EC50 ≈135, 200 and 191 ppm, respectively) | On-plants assays in greenhouse (DSI reduced by 30, 70 and 35% CTC after 50 mL at 100 ppm applied on the roots of inoculated plants) | capsici dreschsleri melonis | [106] | ||
| Mycelium growth inhibition on Petri dish (No growth at 6250 µg/mL after 14 days) | - | infestans | [74] | |||
| Mycelium growth inhibition on Petri dish (Total inhibition at 244 and ED50 ≈ 60 mg/L) Sporangia and zoospores production (Totally hampered at 144 mg/L) Zoospores germination (Totally inhibited from 72 mg/mL) | - | parasitica | [99] | |||
| Palmarosa | Cymbopogon Martini | Mycelium growth inhibition (EC50 ≈ 0.10 µg/mL by contact and EC50 ≈ 0.15 µg/mL by fumigation) Sporangia and zoospores production ((EC50 ≈ 0.04 µg/mL) and germination (EC50 ≈ 0.08 µg/mL) | - | capsici | [100] | |
| Euphorbiaceae | Croton | Croton macrostachyrus | - | On-field assays on 2 potato cultivars presenting ≠ levels of resistance (DSI −25% CTC 53 DAP on susceptible and −15% CTC 60 DAP on resistant) | infestans | [102] |
| Apiaceae | Caraway | Carum carvi | Mycelium growth inhibition on Petri dish (4 µL/plate—no inhibition at all CTC after 22 days) | Greenhouse experiments on 2 potatoes cultivars (1:500 v/v—reduction of 15 and 35% of DSI CTC over 22 days) | [95] | |
| Mycelium growth inhibition (ND) | - | [125] | ||||
| Fumigation test against mycelium growth (total inhibition from 0.4 µg/mL air) Contact test against mycelium growth (total inhibition from 6.4 µg/mL) Contact effect on sporangia production (Absence of sporangia from 3.2 µg/mL) | - | [94] | ||||
| Cumin | Cuminum cymimum | Mycelium growth inhibition on Petri dish (Total inhibition at 216 and ED50 ≈60 mg/L) Sporangia and zoospores production (80% inhibited from 144 mg/L) Zoospores germination (Totally inhibited from 144 mg/mL) | - | parasitica | [99] | |
| Mycelium growth inhibition on Petri dish (10% inhibition CTC with 4 µL/plate after 22 days) | Greenhouse experiments on 2 potatoes cultivars (DSR of 20 and 30% CTC after 22 days at 1:500 v/v) | infestans | [95] | |||
| Fennel | Foeniculum vulgare | Mycelium growth inhibition (EC50 ≈ 8.10 µg/mL by contact but no inhibition at all by fumigation) | - | capsici | [100] | |
| Dill | Anethum graveolens | Mycelium growth inhibition on Petri dish (ND) | [126] | |||
| Cannabaceae | Hop | Humulus lupulus | Mycelium growth on twelve-well plates (IC50 > 1000 mg/L) Spores’ germination on microplates (IC50 > 5000 mg/L) | - | infestans | [76] |
| Geraniaceae | / | Pelagornium graveolens | Mycelium growth inhibition (ED50 ≈ 140 mg/L) | - | parasitica | [99] |
| Geranium | Geranium spp. (ND) | Mycelium growth inhibition by contact (Total inhibition at 1 µL/mL for all spp.) Mycelium growth inhibition by fumigation (Total inhibition at 100 µL/L air for all spp.) | - | capsici melonis nicotianae cinnamoni citrophthora | [109] | |
| Chemical Class | Metabolite | CAS Number | Found as Major Component in the Essential Oils of | Reference |
|---|---|---|---|---|
| Terpenoids | ||||
| Hydrocarbons | D-limonene | 5989−27-5 | Citrus limon | [120] |
| Citrus bergamia | ||||
| Citrus sinensis | ||||
| Zanthoxylum armatum | [123] | |||
| Tetradium glabrifolium | [124] | |||
| Citrus aurantifolia | [122] | |||
| α-pinene | 80-56-8 | Rosmarinus officinalis | [102,105] | |
| Cistus ladanifer | [157] | |||
| α-terpinene | 99-86-5 | Origanum majorana | [104] | |
| α-selinene | 473-13-2 | Chrysanthemum indicum | [118] | |
| α-humulene | 6753-98-6 | Humulus lupulus | [76] | |
| γ-terpinene | 99-85-4 | Thymus vulgaris | [96] | |
| Ocimum gratissimum | [107] | |||
| €-β-caryophyllene | 87-44-5 | Humulus lupulus | [76] | |
| α-phellandrene | 99-83-2 | Anethum graveolens | [126] | |
| p-cymene | 99-87-6 | Thymus pectipatus | [96] | |
| Origanum marjorana | [104] | |||
| β-ocimene | 3779-61-1 | Ocimum basilicum | [106] | |
| β-elemene | 33880-83-0 | Tetradium glabrifolium | [124] | |
| δ-cadimène | 483-76-1 | Chrysanthemum indicum | [118] | |
| Phenolic compounds | Carvacrol | 499-75-2 | Thymus vulgaris | [94,96] |
| Thymus schimperi | [102] | |||
| Origanum vulgaris | [94] | |||
| Origanum compactum | [98] | |||
| Satureja montana | [98] | |||
| Thymus capitatus | [101] | |||
| Thymus serpyllum | [103] | |||
| Thymol | 89-83-8 | Thymus vulgaris | [98,99,100,101] | |
| Thymus pectinatus | [96] | |||
| Ocimum gratissimum | [107] | |||
| Zataria multiflora | [111] | |||
| Alcohol | Borneol | 507-70-0 | Thymus satureioides | [132] |
| Ocinum basilicum | [106] | |||
| Rosmarinus officinalis | [94] | |||
| Linalol | 78-70-6 | Ocinum basilicum | [99] | |
| Zanthoxylum armatum | [123] | |||
| Terpinen-4-ol | 562-74-3 | Melaleuca alternifolia | [98] | |
| Citronellol | 106-22-9 | Pelargonium graveolens | [99] | |
| Zanthoxylum xanthoxyloides | [112] | |||
| Geraniol | 106-24-1 | Citrus aurantifolia | [122] | |
| Curcumol | 4871-97-0 | Curcuma zedoaria | [115] | |
| Ether | Eucalyptol | 470-82-6 | Eucalyptus globulus | [102] |
| Laurus nobilis | [94] | |||
| Thymus convoltus | [96] | |||
| Origanum majorana | [104] | |||
| Curcuma zedoaria | [115] | |||
| Dill ether | 74410-10-9 | Anethum graveolens | [126] | |
| Ketone | L-carvone | 6485-40-1 | Mentha spicata | [98,108] |
| D-carvone | 2244-16-8 | Anethum graveolens | [126] | |
| Carum carvi | [125] | |||
| L-menthone | 89-78-1 | Mentha piperita | [108] | |
| Camphor | 76-22-2 | Lavendula officinalis | [94] | |
| Thymus convolutus | [96] | |||
| Aldehyde | Citronellal | 106-23-0 | Eucalyptus citriodara | [98,105] |
| Neral—Citral B | 106-26-3 | Cymbopogon citratus | [106] | |
| Geranial—Citral A | 5392-40-5 | Cymbopogon citratus | [99,106,121] | |
| Citrus sinensis | [121,122] | |||
| Zingiber officinale | [117] | |||
| Ester | Bornyl acetate | 76-49-3 | Citrus aurantifolia | [122] |
| Phenylpropanoids | ||||
| Ether | Eugenol | 97-53-0 | Syzygium aromaticum | [93,98,112] |
| Syringa oblata | [158] | |||
| Methyleugenol | 93-15-2 | Asarum heterotropoides | [159] | |
| Anethole | 104-46-1 | Foeniculum vulgare | [94] | |
| Aldehyde | Cuminaldehyde | 122-03-2 | Cuminum cyminum | [99] |
| Cinnamaldehyde | 14371-10-9 | Cinnamomum cassia | [99] | |
| Cinnamomum zeylanicum | [111,111] | |||
| Other metabolic pathway | ||||
| Sulfur compounds | Diallyl disulfide | 2179-57-9 | Allium sativum | [119] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martini, F.; Jijakli, M.H.; Gontier, E.; Muchembled, J.; Fauconnier, M.-L. Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review. Molecules 2023, 28, 7302. https://doi.org/10.3390/molecules28217302
Martini F, Jijakli MH, Gontier E, Muchembled J, Fauconnier M-L. Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review. Molecules. 2023; 28(21):7302. https://doi.org/10.3390/molecules28217302
Chicago/Turabian StyleMartini, Florian, M. Haïssam Jijakli, Eric Gontier, Jérôme Muchembled, and Marie-Laure Fauconnier. 2023. "Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review" Molecules 28, no. 21: 7302. https://doi.org/10.3390/molecules28217302
APA StyleMartini, F., Jijakli, M. H., Gontier, E., Muchembled, J., & Fauconnier, M.-L. (2023). Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review. Molecules, 28(21), 7302. https://doi.org/10.3390/molecules28217302