


Statistical Improvement of rGILCC 1 and rPOXA 1B Laccases Activity Assay Conditions Supported by Molecular Dynamics

 , ,
, ,  ,
,
Abstract
:
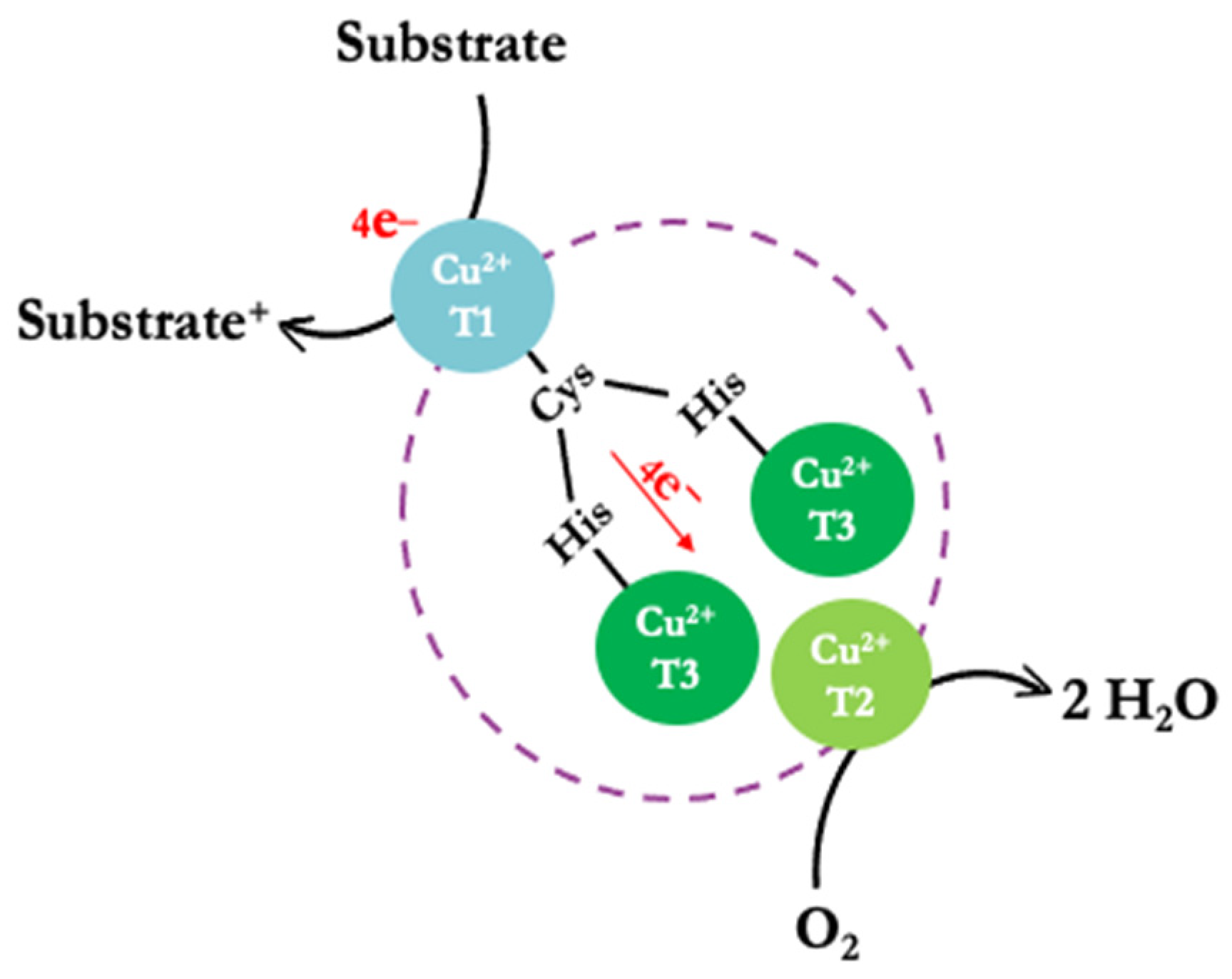
1. Introduction
2. Results
2.1. Optimal Experimental Designs (OED)
2.2. Low-pH Assay for rGILCC 1 and rPOXA 1B in Acetate and Citrate-Phosphate Buffers
2.3. Mean Comparison
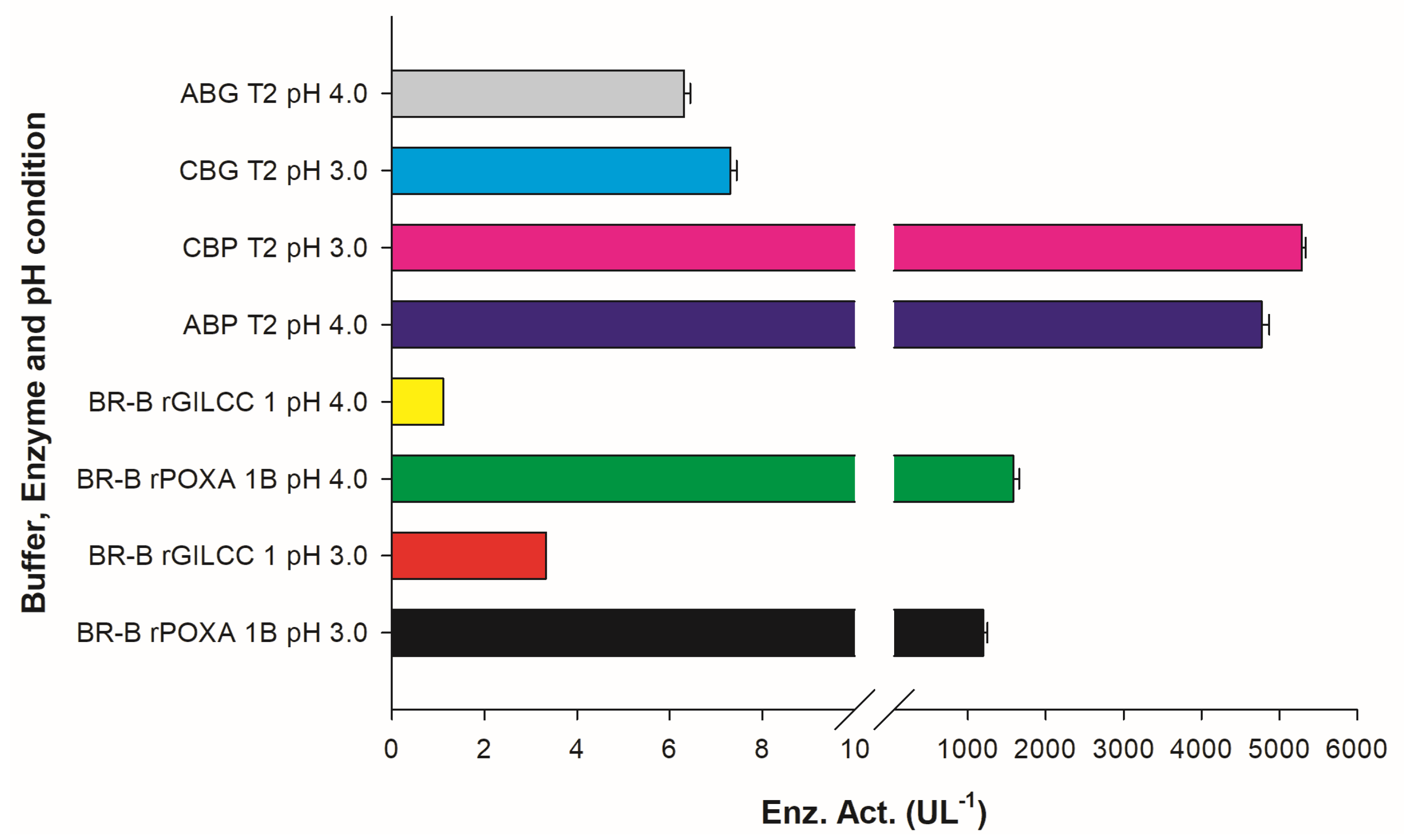
2.4. Assay for rGILCC 1 and rPOXA 1B in Britton–Robinson Buffer at pH 3.0 and 4.0 ± 0.2
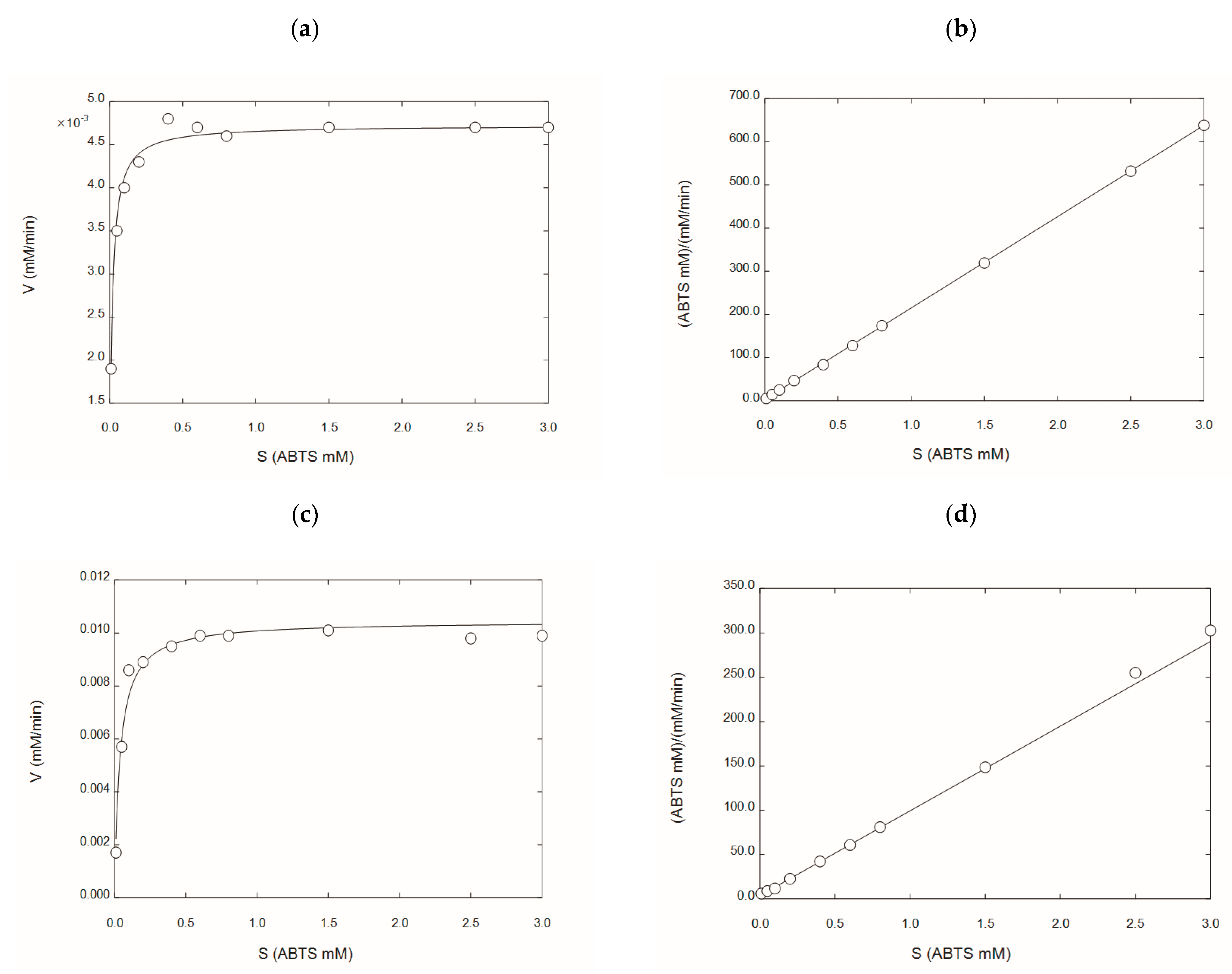
2.5. Kinetic Characterization of rGILCC 1 and rPOXA 1B under Best Conditions Found at the Mean Comparison
2.6. Molecular Docking of the 3D Models of GILCC 1 and POXA 1B with ABTS at pH 3.0
2.7. Molecular Dynamics of 3D Models of GILCC 1 and POXA 1B the Complexes GILCC 1–ABTS and POXA 1B–ABTS at pH 3.0
3. Discussion
3.1. Optimal Experimental Designs of rGILCC 1 and rPOXA 1B in Acetate Buffer
3.2. Optimal Experimental Designs of rGILCC 1 and rPOXA 1B in Citrate Buffer
3.3. Low-pH Assay for rGILCC 1 and rPOXA 1B in Acetate and Citrate-Phosphate Buffers
3.4. Kinetic Characterization of rGILCC 1 and rPOXA 1B under Best Conditions Found at the Mean Comparison
3.5. GILCC 1 and POXA 1B Laccases—ABTS Molecular Docking and Molecular Dynamic
4. Materials and Methods
4.1. Concentrates of Recombinant Laccases
4.2. Optimal Experimental Designs
4.3. Low-pH Assay (One-Factor Experimental Designs, OFED) for rGILCC 1 and rPOXA 1B Assayed in Acetate and Citrate-Phosphate
4.4. Assay for rGILCC1 and rPOXA 1B in Britton–Robinson Buffer at pH 3.0 and 4.0 ± 0.2
4.5. Laccase Activity Assay Using Acetate Buffer
4.6. Laccase Activity Assay Using Citrate Buffer
4.7. Mean Comparison
4.8. Kinetic Characterization of rGILCC 1 and rPOXA 1B under Best Conditions Found at the Mean Comparison
4.9. Three-dimensional Computational Homology and Metal Ion Modelling of GILCC 1 and POXA 1B Laccases
4.10. GILCC 1–ABTS and POXA 1B–ABTS Molecular Docking and Dynamics
4.10.1. Enzyme and Ligand Preparation
4.10.2. Molecular Docking
4.10.3. Molecular Dynamics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Asano, T.; Taoka, M.; Yamauchi, Y.; Everroad, R.C.; Seto, Y.; Isobe, T.; Kamo, M.; Chosa, N. Re-examination of a a-chymotrypsin-solubilized laccase in the pupal cuticle of the silkworm, Bombyx mori: Insights into the regulation system for laccase activation during the ecdysis process. Insect Biochem. Mol. Biol. 2014, 55, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Otto, B.; Schlosser, D. First laccase in green algae: Purification and characterization of an extracellular phenol oxidase from Tetracystis aeria. Planta 2014, 240, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Gupte, S.; Gahlout, M.; Gupte, A. Purification and characterization of an extracellular laccase from solid-state culture of Pleurotus ostreatus HP-1. 3Biotech 2014, 4, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hoyos, C.M.; Morales-Álvarez, E.D.; Poutou-Piñales, R.A.; Pedroza-Rodríguez, A.M.; Rodríguez-Vázquez, R.; Delgado-Boada, J.M. Fungal laccases. Fung. Biol. Rev. 2013, 27, 67–82. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Gorbacheva, M.A.; Shleev, S.V.; Yaropolov, A.I. “Blue” laccases. Biochem 2007, 72, 1136–1150. [Google Scholar] [CrossRef]
- Mateljak, I.; Monza, E.; Lucas, M.F.; Guallar, V.; Aleksejeva, O.; Ludwig, R.; Leech, D.; Shleev, S.; Alcalde, M. Increasing redox potential, redox mediator activity, and stability in a fungal laccase by computer-guided mutagenesis and directed evolution. ACS Catal. 2019, 9, 4561–4572. [Google Scholar] [CrossRef]
- Rivera-Hoyos, C.M.; Morales-Álvarez, E.D.; Abelló-Esparza, J.; Buitrago-Pérez, D.F.; Martínez-Aldana, N.; Salcedo-Reyes, J.C.; Poutou-Piñales, R.A.; Pedroza-Rodríguez, A.M. Detoxification of pulping black liquor with Pleurotus ostreatus or recombinant Pichia pastoris followed by CuO/TiO2/visible photocatalysis. Sci. Rep. 2018, 8, 3503. [Google Scholar] [CrossRef]
- Aghaie-Khouzani, M.; Forootanfar, H.; Moshfegh, M.; Khoshayand, M.R.; Faramarzi, M.A. Decolorization of some synthetic dyes using optimized culture broth of laccase producing ascomycete Paraconiothyrium variabile. Biochem. Eng. J. 2012, 60, 9–15. [Google Scholar] [CrossRef]
- Forootanfar, H.; Rezaei, S.; Zeinvand-Lorestani, H.; Tahmasbi, H.; Mogharabi, M.; Ameri, A.; Faramarzi, M.A. Studies on the laccase-mediated decolorization, kinetic, and microtoxicity of some synthetic azo dyes. J. Environ. Health Sci. 2016, 14, 7. [Google Scholar] [CrossRef]
- Zeng, S.; Qin, X.; Xia, L. Degradation of the herbicide isoproturon by laccase-mediator systems. Biochem. Eng. J. 2017, 119, 92–100. [Google Scholar] [CrossRef]
- Mora-Gamboa, M.P.C.; Rincón-Gamboa, S.M.; Ardila-Leal, L.D.; Poutou-Piñales, R.A.; Pedroza-Rodríguez, A.M.; Quevedo-Hidalgo, B.E. Impact of antibiotics as waste, physical, chemical, and enzymatical degradation: Use of laccases. Molecules 2022, 27, 4436. [Google Scholar] [CrossRef]
- Sumathi, T.; Viswanath, B.; Sri Lakshmi, A.; SaiGopal, D.V. Production of laccase by Cochliobolus sp. isolated from plastic dumped soils and their ability to degrade low molecular weight PVC. Biochem. Res. Int. 2016, 2016, 9519527. [Google Scholar] [CrossRef] [PubMed]
- Ferraroni, M.; Westphal, A.H.; Borsari, M.; Tamayo-Ramos, J.A.; Briganti, F.; de Graaff, L.H.; van Berkel, W.J.H. Structure and function of Aspergillus niger laccase McoG. Biocatal. Agric. Biotechnol. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Britos, C.N.; Trelles, J.A. Development of strong enzymatic biocatalysts for dye decolorization. Biocat. Agricult. Biotechnol. 2016, 7, 228–233. [Google Scholar] [CrossRef]
- Pietra, F. On dioxygen permeation of MaL laccase from the thermophilic fungus Melanocarpus albomyces: An all-atom molecular dynamics investigation. Chem. Biodiv. 2016, 13, 493–1501. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Solomon, E.I. Electron transfer and reaction mechanism of laccases. CMLS 2015, 72, 869–883. [Google Scholar] [CrossRef]
- Hakulinen, N.; Rouvinen, J. Three-dimensional structures of laccases. CMLS 2015, 72, 857–868. [Google Scholar] [CrossRef]
- Komori, H.; Higuchi, Y. Structure and molecular evolution of multicopper blue proteins. Biomol. Concepts 2010, 1, 31–40. [Google Scholar] [CrossRef]
- Tonin, F.; Melis, R.; Cordes, A.; Sanchez-Amat, A.; Pollegioni, L.; Rosini, E. Comparison of different microbial laccases as tools for industrial uses. New Biotechnol. 2016, 33, 387–398. [Google Scholar] [CrossRef]
- Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases: A never-ending story. Cell Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef]
- Theerachat, M.; Guieysse, D.; Morel, S.; Remaud-Simeon, M.; Chulalaksananukul, W. Laccases from Marine Organisms and Their Applications in the Biodegradation of Toxic and Environmental Pollutants: A Review. Appl. Biochem. Biotechnol. 2019, 187, 583–611. [Google Scholar] [CrossRef]
- Dwivedi, U.N.; Singh, P.; Pandey, V.P.; Kumar, A. Structure–function relationship among bacterial, fungal and plant laccases. J. Mol. Catal B Enz. 2011, 68, 117–128. [Google Scholar] [CrossRef]
- Bisswanger, H. Enzyme assays. Perspect Sci. 2014, 1, 41–55. [Google Scholar] [CrossRef]
- Ezike, T.C.; Ezugwu, A.L.; Udeh, J.O.; Ugwuoke, K.C.; Eze, S.O.O.; Chilaka, F.C. Substrate specificity of a new laccase from Trametes polyzona WRF03. Heliyon 2021, 7, e06080. [Google Scholar] [CrossRef]
- Galli, C.; Madzak, C.; Vadala, R.; Jolivalt, C.; Gentili, P. Concerted electron/proton transfer mechanism in the oxidation of phenols by laccase. Chembiochem 2013, 14, 2500–2505. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Li, Y.; Xia, Y.L.; Ai, S.M.; Liang, J.; Sang, P.; Ji, X.L.; Liu, S.Q. Insights into protein-ligand interactions: Mechanisms, models, and methods. Int. J. Mol. Sci. 2016, 17, 144. [Google Scholar] [CrossRef]
- Alzate-Morales, J.H.; Contreras, R.; Soriano, A.; Tuñon, I.; Silla, E. A Computational Study of the Protein-Ligand Interactions in CDK2 Inhibitors: Using Quantum Mechanics/Molecular Mechanics Interaction Energy as a Predictor of the Biological Activity. Biophys. J. 2007, 92, 430–439. [Google Scholar] [CrossRef]
- Meng, X.-Y.; Zhang, H.-X.; Mezei, M.; Cui, M. Molecular Docking: A powerful approach for structure-based drug discovery. Curr. Comp.-Aided. Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Bhatt, P.; Bhatt, K.; Chen, W.J.; Huang, Y.; Xiao, Y.; Wu, S.; Lei, Q.; Zhong, J.; Zhu, X.; Chen, S. Bioremediation potential of laccase for catalysis of glyphosate, isoproturon, lignin, and parathion: Molecular docking, dynamics, and simulation. J. Hazard Mater. 2023, 443, 130319. [Google Scholar] [CrossRef]
- Morales-Álvarez, E.D.; Rivera-Hoyos, C.M.; Cardozo-Bernal, Á.M.; Poutou-Piñales, R.A.; Pedroza-Rodríguez, A.M.; Díaz-Rincón, D.J.; Rodríguez-López, A.; Alméciga-Díaz, C.J.; Cuervo-Patiño, C.L. Plackett-Burman design for rGILCC1 laccase activity enhancement in Pichia pastoris: Concentrated enzyme kinetic characterization. Enz. Res. 2017, 2017, 5947581. [Google Scholar] [CrossRef]
- Ardila-Leal, L.D.; Albarracín-Pardo, D.A.; Rivera-Hoyos, C.M.; Morales, E.D.; Poutou-Piñales, R.A.; Cardozo-Bernal, A.M.; Quevedo-Hidalgo, B.E.; Pedroza-Rodríguez, A.M.; Díaz-Rincón, D.J.; Rodríguez-Lopez, A.; et al. Media improvement for 10 L bioreactor production of rPOXA 1B laccase by P. pastoris. 3Biotech 2019, 9, 447. [Google Scholar] [CrossRef] [PubMed]
- Ardila-Leal, L.D. Producción a Escala Piloto (10L) y Caracterización de un Concentrado Enzimático de rPOXA 1B Para la Remoción de Colorantes. In Microbiology Department; Pontificia Universidad Javeriana: Bogotá, Colombia, 2021; p. 361. [Google Scholar]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminformat. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Pinar, O.; Tamerler, C.; Karataş, A.Y. Heterologous expression and characterization of a high redox potential laccase from Coriolopsis polyzona MUCL 38443. Turkish J. Biol. 2017, 41, 278–291. [Google Scholar] [CrossRef]
- Bertrand, B.; Trejo-Hernández, M.; Morales-Guzmán, D.; Caspeta, L.; Suárez Rodríguez, R.; Martínez-Morales, F. Functional expression, production, and biochemical characterization of a laccase using yeast surface display technology. Fungal. Biol. 2016, 120, 1609–1622. [Google Scholar] [CrossRef]
- Piscitelli, A.; Giardina, P.; Mazzoni, C.; Sannia, G. Recombinant expression of Pleurotus ostreatus laccases in Kluyveromyces lactis and Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2005, 69, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Acker, M.G.; Auld, D.S. Considerations for the design and reporting of enzyme assays in high-throughput screening applications. Perspect Sci. 2014, 1, 56–73. [Google Scholar] [CrossRef]
- Zbacnik, T.J.; Holcomb, R.E.; Katayama, D.S.; Murphy, B.M.; Payne, R.W.; Coccaro, R.C.; Evans, G.J.; Matsuura, J.E.; Henry, C.S.; Manning, M.C. Role of buffers in protein formulations. J. Pharm. Sci. 2017, 106, 713–733. [Google Scholar] [CrossRef]
- Itoh, N.; Takagi, S.; Miki, A.; Kurokawa, J. Characterization and cloning of laccase gene from Hericium coralloides NBRC 7716 suitable for production of epitheaflagallin 3-O-gallate. Enz. Microb. Technol. 2016, 82, 125–132. [Google Scholar] [CrossRef]
- Childs, R.E.; Bardsley, W.G. The steady-state kinetics of peroxidase with 2,2′-Azino-di-(3-ethylbenzthiazoline- 6-sulphonic acid) as chromogen. Biochem. J. 1975, 145, 93–103. [Google Scholar] [CrossRef]
- Wolfenden, B.S.; Willson, R.L. Radical-cations as reference chromogens in kinetic studies of oneelectron transfer reactions: Pulse radiolysis studies of 2,2’-azinobisethyl benzthiazoline-6-sulphonate). J. Chem. Soc.-Perkin Transact 2 1982, 7, 805–812. [Google Scholar] [CrossRef]
- Tinoco, R.; Pickard, M.A.; Vazquez-Duhalt, R. Kinetic differences of purified laccases from six Pleurotus ostreatus strains. Lett. Appl. Microbiol. 2001, 32, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, B.; Martínez-Morales, F.; Trejo-Hernández, M.ı.R. Upgrading laccase production and biochemical properties: Strategies and challenges. Biotechnol. Prog. 2017, 33, 1015–1034. [Google Scholar] [CrossRef]
- Morales-Álvarez, E.D.; Rivera-Hoyos, C.M.; Chaparro-Núñez, L.E.; Daza, C.E.; Poutou-Piñales, R.A.; Pedroza-Rodríguez, A.M. Decolorization and detoxification of malachite green by Ganoderma lucidum: Key operating parameters and adsorption studies. J. Env. Eng. 2017, 143, 04016093. [Google Scholar] [CrossRef]
- Salem, A.A.; Soliman, A.A.; El-Haty, I.A. New spectrophotometric method for determining nitrogen dioxide in air using 2,2-azino-bis(3-ethyl benzothiazoline)-6-sulfonic acid-diammonium salt and passive sampling. Anal Chem. Insights 2011, 6, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Cox, M.M. Lehninger, Principles of Biochemistry, 7th ed.; Freeman, W.H., Ed.; Macmillan Higher Education: New York, NY, USA, 2017. [Google Scholar]
- Bourbonnais, R.; Paice, M.G. Demethylation and delignification of kraft pulp by Trametes versicolor laccase in the presence of 2,2’-azinobis-(3-ethylbenzthiazoline-6-sulphonate). Appl. Microbiol. Biotechnol. 1992, 36, 823–827. [Google Scholar] [CrossRef]
- Pezzella, C.; Giacobelli, V.G.; Lettera, V.; Olivieri, G.; Cicatiello, P.; Sannia, G.; Piscitelli, A. A step forward in laccase exploitation: Recombinant production and evaluation of techno-economic feasibility of the process. J. Biotechnol. 2017, 259, 175–181. [Google Scholar] [CrossRef]
- Gu, C.; Zheng, F.; Long, L.; Wang, J.; Ding, S. Engineering the expression and characterization of two novel laccase isoenzymes from Coprinus comatus in Pichia pastoris by fusing an additional ten amino acids tag at N-terminus. PLoS ONE 2014, 9, e93912. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, L.; Zhang, H.; Wang, S.; Zhang, X.; Geng, A. A novel homodimer laccase from Cerrena unicolor BBP6: Purification, characterization, and potential in dye decolorization and denim bleaching. PLoS ONE 2018, 13, e0202440. [Google Scholar] [CrossRef] [PubMed]
- Farnet, A.M.; Gil, G.; Ferre, E. Effects of pollutants on laccase activities of Marasmius quercophilus, a white-rot fungus isolated from a Mediterranean schlerophyllous litter. Chemosphere 2008, 70, 895–900. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.M.H.; Pinto, I.S.S.; Soares, E.V.; Soares, H.M.V.M. (Un)suitability of the use of pH buffers in biological, biochemical and environmental studies and their interaction with metal ions—a review. RSC Adv. 2015, 5, 30989–31003. [Google Scholar] [CrossRef]
- Eichlerová, I.; Šnajdr, J.; Baldrian, P. Laccase activity in soils: Considerations for the measurement of enzyme activity. Chemosphere 2012, 88, 1154–1160. [Google Scholar] [CrossRef]
- Salony; Mishra, S.; Bisaria, V.S. Production and characterization of laccase from Cyathus bulleri and its use in decolourization of recalcitrant textile dyes. Appl. Microbiol. Biotechnol. 2006, 71, 646–653. [Google Scholar] [CrossRef]
- Salis, A.; Monduzzi, M. Not only pH. Specific buffer effects in biological systems. Curr. Opin. Colloid. Interf. Sci. 2016, 23, 1–9. [Google Scholar] [CrossRef]
- Ahmad, I.; Sheraz, M.A.; Ahmed, S.; Kazi, S.H.; Mirza, T.; Aminuddin, M. Stabilizing effect of citrate buffer on the photolysis of riboflavin in aqueoussolution. Res. Pharma. Sci. 2011, 1, 11–15. [Google Scholar] [CrossRef]
- Warner, R.C.; Weber, I. The cupric and ferric citrate complexes. J. Amer. Chem. Soc. 1953, 75, 5086–5094. [Google Scholar] [CrossRef]
- Sahbaz, F.; Somer, G. The effect of citrate anions on the kinetics of cupric ion-catalysed oxidation of ascorbic acid. Food Chem. 1993, 47, 345–349. [Google Scholar] [CrossRef]
- Garg, N.; Bieler, N.; Kenzom, T.; Chhabra, M.; Ansorge-Schumacher, M.; Mishra, S. Cloning, sequence analysis, expression of Cyathus bulleri laccase in Pichia pastoris and characterization of recombinant laccase. BMC Biotechnol. 2012, 12, 75. [Google Scholar] [CrossRef]
- Li, Q.; Pei, J.; Zhao, L.; Xie, J.; Cao, F.; Wang, G. Overexpression and characterization of laccase from Trametes versicolor in Pichia pastoris. Appl. Biochem. Microbiol. 2014, 50, 140–147. [Google Scholar] [CrossRef]
- Li, Q.; Ge, L.; Cai, J.; Pei, J.; Xie, J.; Zhao, L. Comparison of Two Laccases from Trametes versicolor for Application in the Decolorization of Dyes. J. Microbiol. Biotechnol. 2014, 24, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Velásquez-Quintero, C.; Merino-Restrepo, A.; Hormaza-Anaguano, A. Production, extraction, and quantification of laccase obtained from an optimized solid-state fermentation of corncob with white-rot fungi. J. Clean Prod. 2022, 370, 133598. [Google Scholar] [CrossRef]
- Cross, R.F.; Cao, J. Salt effects in capillary zone electrophoresis. IV. Resolution versus time and the effect of potassium phosphate and its concentration in the high ionic strength separation of sulphonamides. J. Chromatogr. A 1999, 849, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.I.; Heppner, D.E.; Johnston, E.M.; Ginsbach, J.W.; Cirera, J.; Qayyum, M.; Kieber-Emmons, M.T.; Kjaergaard, C.H.; Hadt, R.G.; Tian, L. Copper active sites in biology. Chem. Rev. 2014, 114, 3659–3853. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Peng, R.-H.; Xiong, A.-S.; Tian, Y.; Zhao, W.; Xu, H.; Liu, D.-T.; Chen, J.-M.; Yao, Q.-H. Secretory expression and characterization of a soluble laccase from the Ganoderma lucidum strain 7071-9 in Pichia pastoris. Mol. Biol. Rep. 2012, 39, 3807–3814. [Google Scholar] [CrossRef] [PubMed]
- Giardina, P.; Palmieri, G.; Scaloni, A.; Fontanella, B.; Faraco, V.; Cennamo, G.; Sannia, G. Protein and gene structure of a blue laccase from Pleurotus ostreatus. Biochem. J. 1999, 341, 655–663. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Nguyen, T.H.; Pham, T.N.H.; Huy, N.T.; Bay, M.V.; Pham, M.Q.; Nam, P.C.; Vu, V.V.; Ngo, S.T. Autodock vina adopts more accurate binding pose but Autodock4 forms better binding affinity. J. Chem. Informat. Model 2020, 60, 204–211. [Google Scholar] [CrossRef]
- Patil, R.; Das, S.; Stanley, A.; Yadav, L.; Sudhakar, A.; Varma, A.K. Optimized hydrophobic interactions and hydrogen bonding at the target-ligand interface leads the pathways of drug-designing. PLoS ONE 2010, 5, e12029. [Google Scholar] [CrossRef] [PubMed]
- Hongyan, L.; Zexiong, Z.; Shiwei, X.; He, X.; Yinian, Z.; Haiyun, L.; Zhongsheng, Y. Study on transformation and degradation of bisphenol A by Trametes versicolor laccase and simulation of molecular docking. Chemosphere 2019, 224, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hoyos, C.M.; Morales-Álvarez, E.D.; Poveda-Cuevas, S.A.; Reyes-Guzmán, E.A.; Poutou-Piñales, R.A.; Reyes-Montaño, E.A.; Pedroza-Rodríguez, A.M.; Rodríguez-Vázquez, R.; Cardozo-Bernal, Á.M. Computational analysis and low-scale constitutive expression of laccases synthetic genes GlLCC1 from Ganoderma lucidum and POXA 1B from Pleurotus ostreatus in Pichia pastoris. PLoS ONE 2015, 10, e0116524. [Google Scholar] [CrossRef]
- Christensen, N.J.; Kepp, K.P. Stability mechanisms of a thermophilic laccase probed by molecular dynamics. PLoS ONE 2013, 8, e61985. [Google Scholar] [CrossRef]
- Enguita, F.J.; Marcal, D.; Martins, L.; Grenha, R.; Henriques, A.; Lindley, P.; Carrondo, M. Substrate and dioxygen binding to the endospore coat laccase from Bacillus subtilis. J. Biol. Chem. 2004, 279, 23472–23476. [Google Scholar] [CrossRef] [PubMed]
- Cambria, M.; Marino, D.; Falconi, M.; Garavaglia, S.; Cambria, A. Docking simulation and competitive experiments validate the interaction between the 2,5-Xylidine inhibitor and Rigidoporus lignosus laccase. J. Biomol. Strcut. Din. 2010, 27, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.K.; Tambolia, A.S.; Sambhare, S.B.; Jeon, B.-H.; Govindwar, S.P. Enzymatic analysis, structural study and molecular docking of laccase and catalase from B. subtilis SK1 after textile dye exposure. Ecol. Informat. 2018, 48, 269–280. [Google Scholar] [CrossRef]
- Mehra, R.; Meyer, A.S.; Kepp, K.P. Molecular dynamics derived life times of active substrate binding poses explain K (M) of laccase mutants. RSC Adv. 2018, 8, 36915–36926. [Google Scholar] [CrossRef] [PubMed]
- Christensen, N.J.; Kepp, K.P. Setting the stage for electron transfer: Molecular basis of ABTS-binding to four laccases from Trametes versicolor at variable pH and protein oxidation state. J. Mol. Catal B Enz. 2014, 100, 68–77. [Google Scholar] [CrossRef]
- Fernandez-Remacha, D.; Gonzalez-Riancho, C.; Lastra Osua, M.; Gonzalez Arce, A.; Montanchez, I.; Garcia-Lobo, J.M.; Estrada-Tejedor, R.; Kaberdin, V.R. Analysis of laccase-like enzymes secreted by fungi isolated from a cave in northern Spain. Microbiol. Open 2022, 11, e1279. [Google Scholar] [CrossRef] [PubMed]
- Jandova, Z.; Vargiu, A.V.; Bonvin, A. Native or non-native protein-protein docking models? molecular dynamics to the rescue. J. Chem. Theory Comput. 2021, 17, 5944–5954. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Re, S.; Sugita, Y. Prediction of protein-ligand Binding pose and affinity using the gREST+FEP method. J. Chem. Informat. Model 2020, 60, 5382–5394. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; He, Y.; Lu, H.P. Interrogating the activities of conformational deformed enzyme by single-molecule fluorescence-magnetic tweezers microscopy. Proc. Natl. Acad. Sci. USA 2015, 112, 13904–13909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ma, G.; Zhu, Y.; Zeng, L.; Ahmad, A.; Wang, C.; Pang, B.; Fang, H.; Zhao, L.; Hao, Q. Active-site conformational fluctuations promote the enzymatic activity of NDM-1. Antimicrob. Agents Chemother. 2018, 62, e01579-18. [Google Scholar] [CrossRef]
- Karshikoff, A.; Nilsson, L.; Ladenstein, R. Rigidity versus flexibility: The dilemma of understanding protein thermal stability. FEBS J. 2015, 282, 3899–3917. [Google Scholar] [CrossRef] [PubMed]
- Ardila-Leal, L.D.; Monterey-Gutiérrez, P.A.; Poutou-Piñales, R.A.; Quevedo-Hidalgo, B.E.; Galindo, J.F.; Pedroza-Rodríguez, A.M. Recombinant laccase rPOXA 1B real-time, accelerated and molecular dynamics stability study. BMC Biotechnol. 2021, 21, 37. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, I.; Endo, K.; Yamamoto, E.; Hirano, Y.; Yasuoka, K. Differences in ligand-induced protein dynamics extracted from an unsupervised deep learning approach correlate with protein-ligand binding affinities. Commun. Biol. 2022, 5, 481. [Google Scholar] [CrossRef] [PubMed]
- Bulbul, E.F.; Melesina, J.; Ibrahim, H.S.; Abdelsalam, M.; Vecchio, A.; Robaa, D.; Zessin, M.; Schutkowski, M.; Sippl, W. Docking, Binding Free Energy Calculations and In Vitro Characterization of Pyrazine Linked 2-Aminobenzamides as Novel Class I Histone Deacetylase (HDAC) Inhibitors. Molecules 2022, 27, 2526. [Google Scholar] [CrossRef]
- De Rienzo, F.; Gabdoulline, R.R.; Wade, R.C.; Sola, M.; Menziani, M.C. Computational approaches to structural and functional analysis of plastocyanin and other blue copper proteins. Cell Mol. Life Sci. 2004, 61, 1123–1142. [Google Scholar] [CrossRef] [PubMed]
- Burquillo, F.J.; Holgado, M.; Bardsley, W.G. Uso del paquete estadístico SIMFIT en la enseñanza del análisis de datos en ciencias experimentales. J. Sci. Edu. 2003, 4, 8–14. [Google Scholar]
- Li, P.; Merz, K.M. MCPB.py: A python based metal center parameter builder. J. Chem. Inf. Model. 2016, 56, 599–604. [Google Scholar] [CrossRef]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, I.T.E.; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Giambasu, G.; et al. AMBER 2020; University of California: San Francisco, CA, USA, 2020. [Google Scholar]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16, Revision A.03; Gaussian Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Ngan, C.H.; Hall, D.R.; Zerbe, B.; Grove, L.E.; Kozakov, D.; Vajda, S. FTSite: High accuracy detection of ligand binding sites on unbound protein structures. Bioinformatics 2012, 28, 286–287. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Montes-Robledo, A.; Baldiris-Avila, R.; Galindo, J.F. D-Mannoside FimH inhibitors as non-antibiotic alternatives for uropathogenic Escherichia coli. Antibiotics 2021, 10, 1072. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | rGILCC 1 (Acetate Buffer) | rPOXA 1B (Acetate Buffer) | ||||||||
| F | p-value | Effect | % | F | p-value | Effect | % | |||
| Value | Prob > F | Contribution | Value | Prob > F | Contribution | |||||
| Model | 51.40 | <0.0001 | 202.14 | <0.0001 | ||||||
| A-pH | 166.98 | <0.0001 | −1.36 | 46.11 | 783.01 | <0.0001 | −1524.96 | 53.62 | ||
| B-wavelength | 82.66 | <0.0001 | −0.96 | 22.83 | 290.63 | <0.0001 | −929.06 | 19.90 | ||
| C-ABTS | 74.37 | <0.0001 | 0.91 | 20.54 | 302.73 | <0.0001 | 948.21 | 20.73 | ||
| AB | 5.89 | 0.0320 | 0.26 | 1.63 | 35.71 | <0.0001 | 325.65 | 2.45 | ||
| AC | 15.88 | 0.0018 | −0.42 | 4.39 | 26.96 | 0.0002 | 282.98 | 1.85 | ||
| BC | 1.49 | 0.2460 | 0.13 | 0.41 | 7.27 | 0.0194 | 146.97 | 0.50 | ||
| ABC | 2.85 | 0.1172 | −0.18 | 0.79 | 1.95 | 0.1874 | −76.19 | 0.13 | ||
| Lack of fit | 7.44 | 0.0629 | 0.09 | 0.9978 | ||||||
| R-squared | 0.9677 | 0.9916 | ||||||||
| Adj R-squared | 0.9489 | 0.9867 | ||||||||
| Pred R-squared | 0.8224 | 0.9774 | ||||||||
| Adeq precision | 18.983 | 39.593 | ||||||||
| Source | rGILCC1 (Citrate Buffer) | rPOXA 1B (Citrate Buffer) | ||||||||
| F | p-value | F | p-value | |||||||
| Value | Prob > F | Value | Prob > F | |||||||
| Model | 100.44 | <0.0001 | 432.14 | <0.0001 | ||||||
| A-pH | 306.42 | <0.0001 | 1740.36 | <0.0001 | ||||||
| B-wavelength | 351.98 | <0.0001 | 621.23 | <0.0001 | ||||||
| C-ABTS | 74.58 | <0.0001 | 487.05 | <0.0001 | ||||||
| AB | 20.03 | 0.0002 | 55.33 | <0.0001 | ||||||
| AC | 13.64 | 0.0010 | 44.23 | <0.0001 | ||||||
| BC | 0.03 | 0.8583 | 2.19 | 0.167 | ||||||
| ABC | 0.50 | 0.6205 | 8.34 | 0.006 | ||||||
| Lack of fit | 1.18 | 0.4944 | 4.28 | 0.130 | ||||||
| R-squared | 0.9892 | 0.9977 | ||||||||
| Adj R-squared | 0.9831 | 0.9954 | ||||||||
| Pred R-squared | 0.9686 | 0.9879 | ||||||||
| Adeq precision | 38.2892 | 60.7103 | ||||||||
| Treatments | rGILCC 1 at Optimal Experimental Designs (OED) | |||||||
| Factors | Enz. Act. (UL−1) | |||||||
| A | B | C | Acetate Buffer (AB) | Citrate Buffer (CB) | ||||
| Acetate Buffer | Citrate Buffer | pH | Wavelength (nm) | ABTS (mM) | Observed | Predicted | Observed | Predicted |
| CBG T1 | 3.00 | 420 | 0.50 | 7.04 ± 0.00 a | 7.12 ± 0.00 | |||
| CBG T2 | 3.00 | 420 | 2.00 | 7.31 ± 0.13 a | 7.30 ± 0.00 | |||
| CBG T3 | 3.00 | 436 | 0.50 | 5.35 ± 0.16 d,e,f | 5.33 ± 0.00 | |||
| CBG T4 | 3.00 | 436 | 2.00 | 5.46 ± 0.00 d,e | 5.40 ± 0.00 | |||
| ABG T1 | CBG T5 | 4.00 | 420 | 0.50 | 5.31 ± 0.06 d,e,f | 5.31 ± 0.00 | 4.72 ± 0.13 g | 4.57 ± 0.00 |
| ABG T2 | CBG T6 | 4.00 | 420 | 2.00 | 6.30 ± 0.17 c | 6.31 ± 0.00 | 5.56 ± 0.00 d | 5.63 ± 0.00 |
| ABG T3 | CBG T7 | 4.00 | 436 | 0.50 | 3.74 ± 0.04 j | 3.79 ± 0.00 | 3.64 ± 0.32 j | 3.72 ± 0.00 |
| ABG T4 | CBG T8 | 4.00 | 436 | 2.00 | 5.08 ± 0.00 f | 5.44 ± 0.00 | 4.70 ± 0.35 g | 4.61 ± 0.00 |
| ABG T5 | CBG T9 | 4.50 | 420 | 0.50 | 3.93 ± 0.07 i,j | 3.93 ± 0.00 | 4.63 ± 0.00 g | 4.71 ± 0.00 |
| ABG T6 | CBG T10 | 4.50 | 420 | 2.00 | 4.47 ± 0.04 g,h | 4.46 ± 0.00 | 5.19 ± 0.00 e,f | 5.13 ± 0.00 |
| ABG T7 | CBG T11 | 4.50 | 436 | 0.50 | 2.81 ± 0.11 k | 3.30 ± 0.00 | 3.75 ± 0.16 j | 3.70 ± 0.00 |
| ABG T8 | CBG T12 | 4.50 | 436 | 2.00 | 3.69 ± 0.05 j | 3.70 ± 0.00 | 4.21 ± 0.16 h,i | 3.70 ± 0.00 |
| Treatments | rPOXA 1B at Optimal Experimental Designs (OED) | |||||||
| Factors | Enz. Act. (UL−1) | |||||||
| A | B | C | Acetate Buffer (AB) | Citrate Buffer (CB) | ||||
| Acetate Buffer | Citrate Buffer | pH | Wavelength (nm) | ABTS (mM) | Observed | Predicted | Observed | Predicted |
| CBP T1 | 3.00 | 420 | 0.50 | 4962.96 ± 13.09 b | 4958.89 ± 0.00 | |||
| CBP T2 | 3.00 | 420 | 2.00 | 5291.67 ± 45.83 a | 5291.68 ± 0.00 | |||
| CBP T3 | 3.00 | 436 | 0.50 | 2870.37 ± 130.95 e | 2870.39 ± 0.00 | |||
| CBP T4 | 3.00 | 436 | 2.00 | 3180.56 ± 124.40 d | 3184.61 ± 0.00 | |||
| ABP T1 | CBP T5 | 4.00 | 420 | 0.50 | 4378.60 ± 28.13 c | 4378.60 ± 0.00 | 2236.11 ± 32.74 i,j | 2244.25 ± 0.00 |
| ABP T2 | CBP T6 | 4.00 | 420 | 2.00 | 4773.66 ± 95.15 b | 4794.42 ± 0.00 | 3870.37 ± 13.09 d | 3866.30 ± 0.00 |
| ABP T3 | CBP T7 | 4.00 | 436 | 0.50 | 2889.64 ± 238.37 g | 2910.40 ± 0.00 | 1300.93 ± 72.02 k | 1296.86 ± 0.00 |
| ABP T4 | CBP T8 | 4.00 | 436 | 2.00 | 3804.51 ± 1.34 d | 3786.64 ± 0.00 | 2067.90 ± 95.03 h | 2073.32 ± 0.00 |
| ABP T5 | CBP T9 | 4.50 | 420 | 0.50 | 2160.49 ± 25.26 i,j | 2160.49 ± 0.00 | 1643.52 ± 32.74 k | 1639.45 ± 0.00 |
| ABP T6 | CBP T10 | 4.50 | 420 | 2.00 | 3320.47 ± 59.46 f | 3320.47 ± 0.00 | 2254.63 ± 32.74 i | 2258.68 ± 0.00 |
| ABP T7 | CBP T11 | 4.50 | 436 | 0.50 | 1518.77 ± 12.07 k | 1500.89 ± 0.00 | 912.04 ± 19.64 l | 916.09 ± 0.00 |
| ABP T8 | CBP T12 | 4.50 | 436 | 2.00 | 2773.35 ± 162.24 g | 2788.35 ± 0.00 | 1666.67 ± 52.38 j | 1658.56 ± 0.00 |
| Treatments | rGILCC 1 and rPOXA 1B at low-pH assay | |||||||
| Factor | Fixed terms | Enz. Act. (UL−1) | ||||||
| A | B | C | Acetate Buffer (AB OFED) | Citrate-Phosphate Buffer (C-PB OFED) | ||||
| pH | Wavelength (nm) | ABTS (mM) | Observed | Observed | ||||
| ABG OFED | 3.6 | 420 | 2 | 6.67 ± 0.19 b | ||||
| ABP OFED | 3.6 | 420 | 2 | 4819.44 ± 55.56 b | ||||
| C-PBG OFED | 2.6 | 420 | 2 | 7.037 ± 0.32 a | ||||
| C-PBP OFED | 2.6 | 420 | 2 | 4884.26 ± 48.78 b | ||||
| Enzyme | Buffer | Apparent Kinetic Parameters | SEM | Confidence Limit 95% | p | References | ||
|---|---|---|---|---|---|---|---|---|
| rGILCC 1 | Citrate | Vmax (mM min−1) | 4.72 × 10−3 | 1.67 × 10−7 | 4.72 × 10−3 | 4.72 × 10−3 | <0.001 | This study |
| Km (mM) | 1.49 × 10−2 | 2.34 × 10−6 | 1.48 × 10−2 | 1.49 × 10−2 | <0.001 | |||
| Acetate | Vmax (mM min−1) | 6.87 × 10−5 | 1.16 × 10−5 | 3.92 × 10−3 | 3.99 × 10−3 | <0.001 | [30] | |
| Km (mM) | 5.36 × 10−2 | 5.17 × 10−4 | 4.75 × 10−2 | 5.03 × 10−2 | <0.001 | |||
| rPOXA 1B | Citrate | Vmax (mM min−1) | 1.05 × 10−2 | 3.20 × 10−4 | 9.70 × 10−3 | 1.12 × 10−2 | <0.001 | This study |
| Km (mM) | 3.72 × 10−2 | 5.07 × 10−3 | 2.56 × 10−2 | 4.89 × 10−2 | <0.001 | |||
| Acetate | Vmax (mM min−1) | 3.16 × 10−2 | 1.0 × 10−3 | 2.90 × 10−2 | 3.40 × 10−2 | <0.001 | [31] | |
| Km (mM) | 1.72 | 1.21 × 10−1 | 1.44 | 1.99 | <0.001 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mora-Gamboa, M.P.C.; Ferrucho-Calle, M.C.; Ardila-Leal, L.D.; Rojas-Ojeda, L.M.; Galindo, J.F.; Poutou-Piñales, R.A.; Pedroza-Rodríguez, A.M.; Quevedo-Hidalgo, B.E. Statistical Improvement of rGILCC 1 and rPOXA 1B Laccases Activity Assay Conditions Supported by Molecular Dynamics. Molecules 2023, 28, 7263. https://doi.org/10.3390/molecules28217263
Mora-Gamboa MPC, Ferrucho-Calle MC, Ardila-Leal LD, Rojas-Ojeda LM, Galindo JF, Poutou-Piñales RA, Pedroza-Rodríguez AM, Quevedo-Hidalgo BE. Statistical Improvement of rGILCC 1 and rPOXA 1B Laccases Activity Assay Conditions Supported by Molecular Dynamics. Molecules. 2023; 28(21):7263. https://doi.org/10.3390/molecules28217263
Chicago/Turabian StyleMora-Gamboa, María P. C., María C. Ferrucho-Calle, Leidy D. Ardila-Leal, Lina M. Rojas-Ojeda, Johan F. Galindo, Raúl A. Poutou-Piñales, Aura M. Pedroza-Rodríguez, and Balkys E. Quevedo-Hidalgo. 2023. "Statistical Improvement of rGILCC 1 and rPOXA 1B Laccases Activity Assay Conditions Supported by Molecular Dynamics" Molecules 28, no. 21: 7263. https://doi.org/10.3390/molecules28217263
APA StyleMora-Gamboa, M. P. C., Ferrucho-Calle, M. C., Ardila-Leal, L. D., Rojas-Ojeda, L. M., Galindo, J. F., Poutou-Piñales, R. A., Pedroza-Rodríguez, A. M., & Quevedo-Hidalgo, B. E. (2023). Statistical Improvement of rGILCC 1 and rPOXA 1B Laccases Activity Assay Conditions Supported by Molecular Dynamics. Molecules, 28(21), 7263. https://doi.org/10.3390/molecules28217263