

Advances in 4-Hydroxyphenylacetate-3-hydroxylase Monooxygenase

Abstract
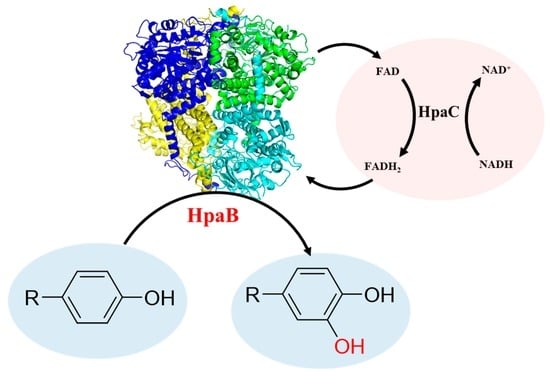
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
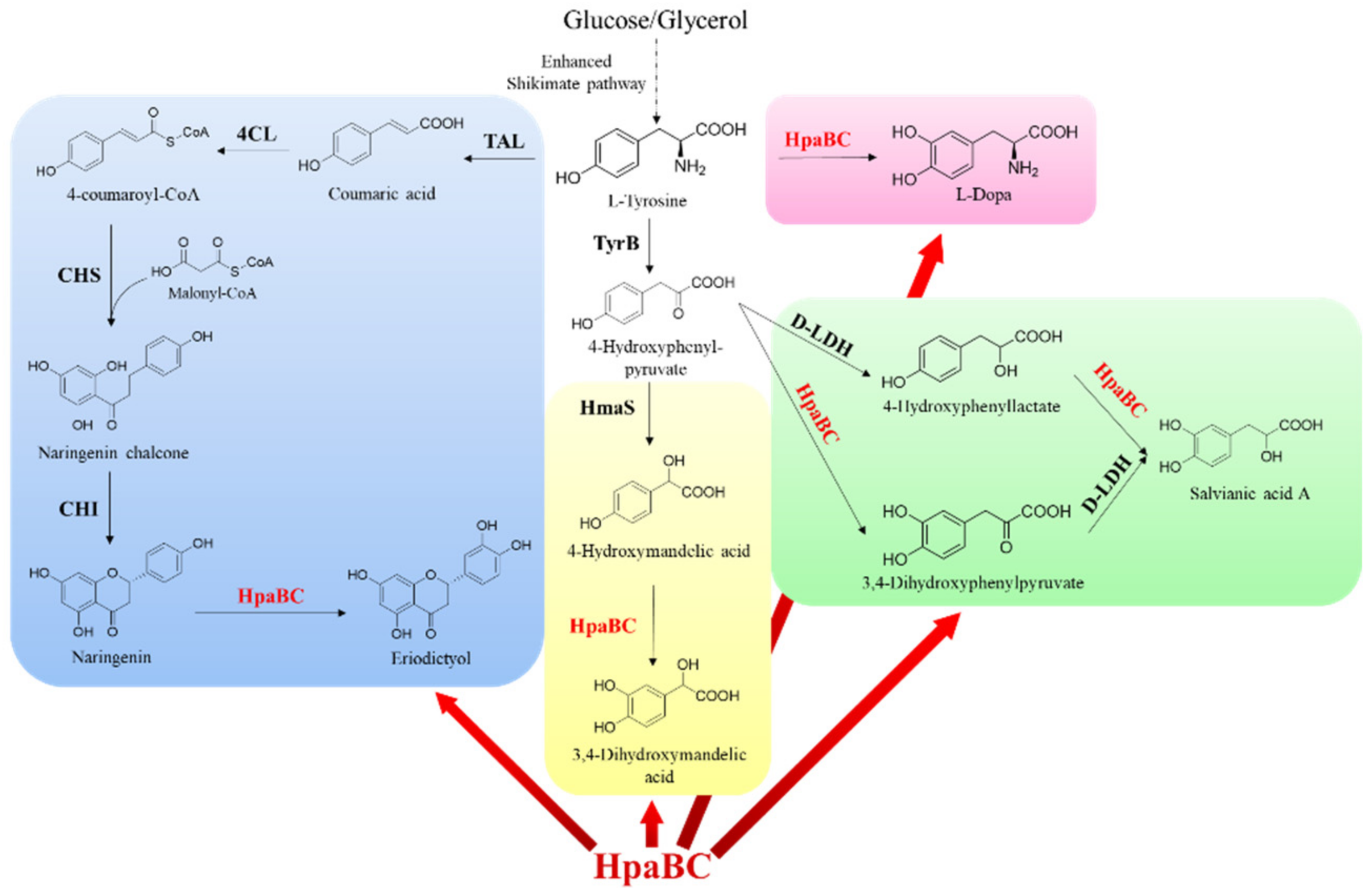{kind=link}
1. Introduction
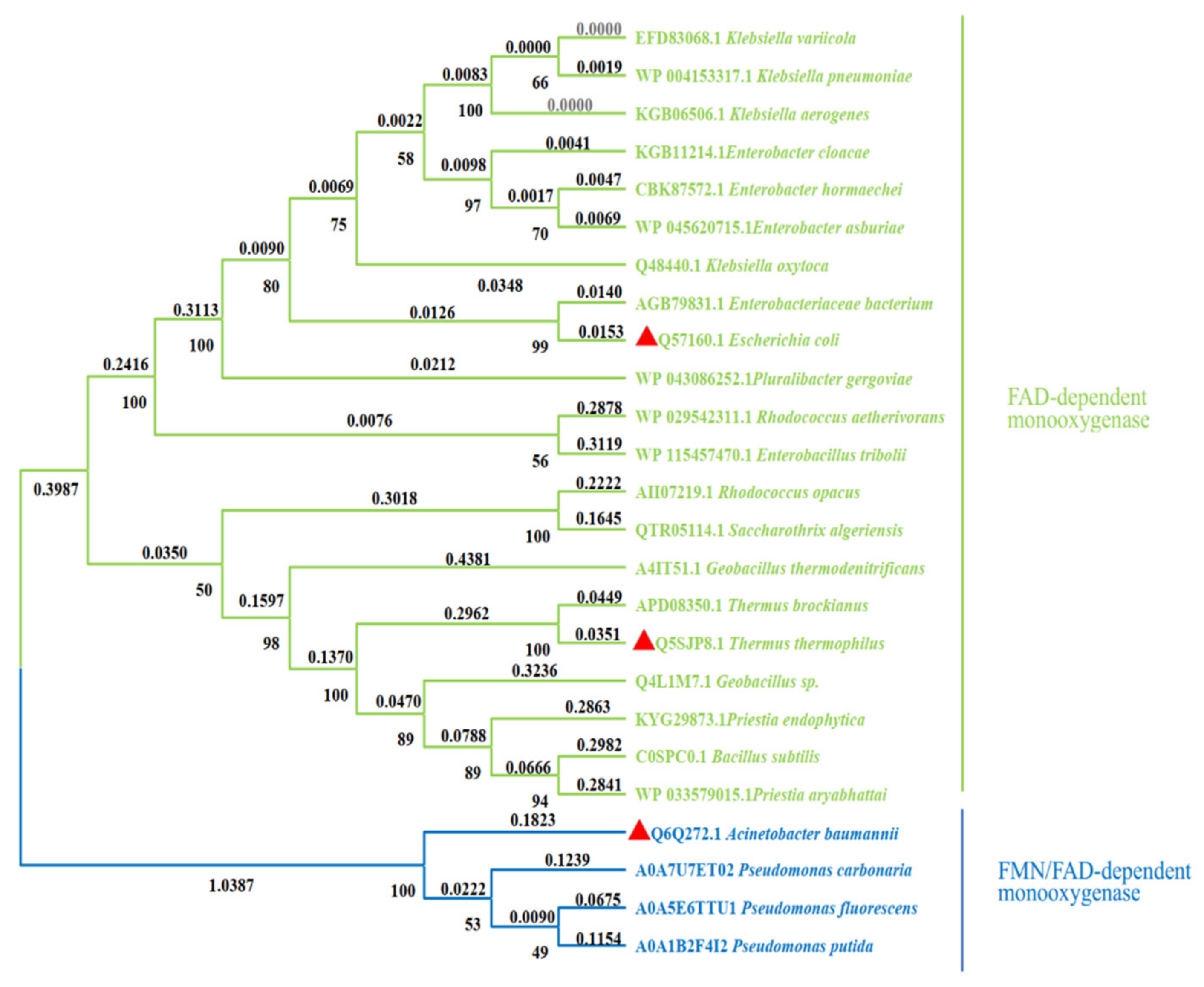
2. Classification of HpaB
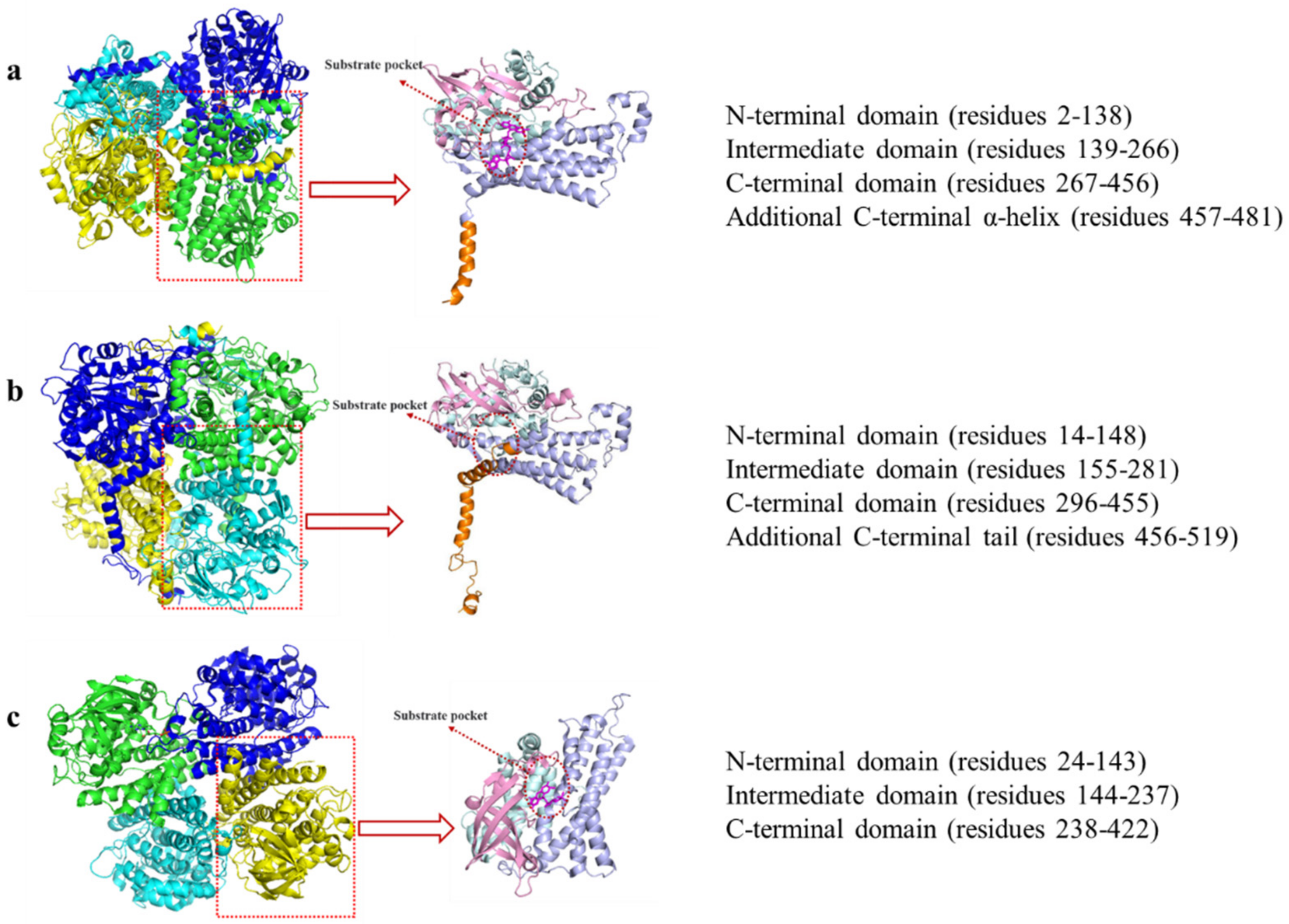
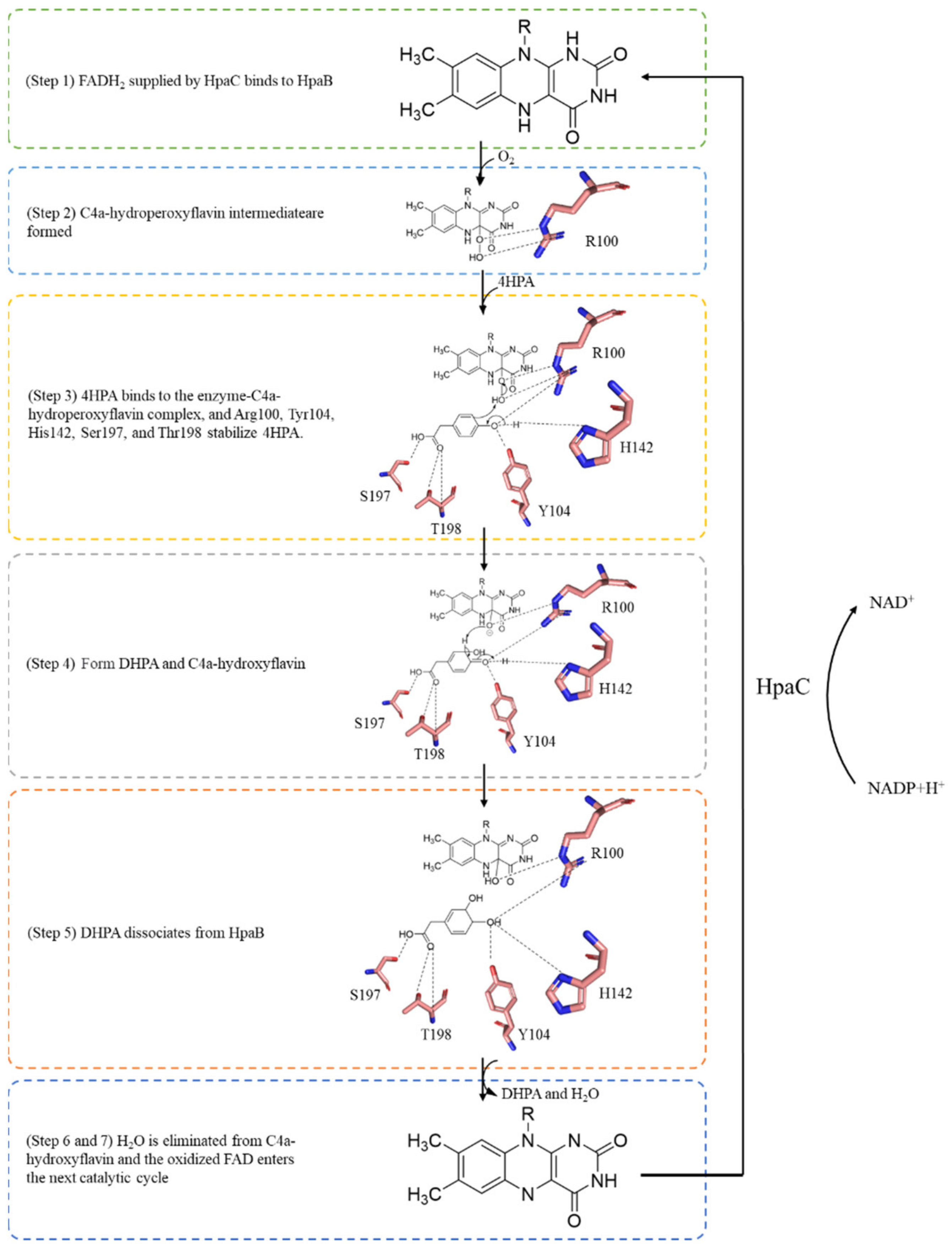
3. HpaB Structure and Catalytic Mechanism

4. Molecular Modification of HpaB
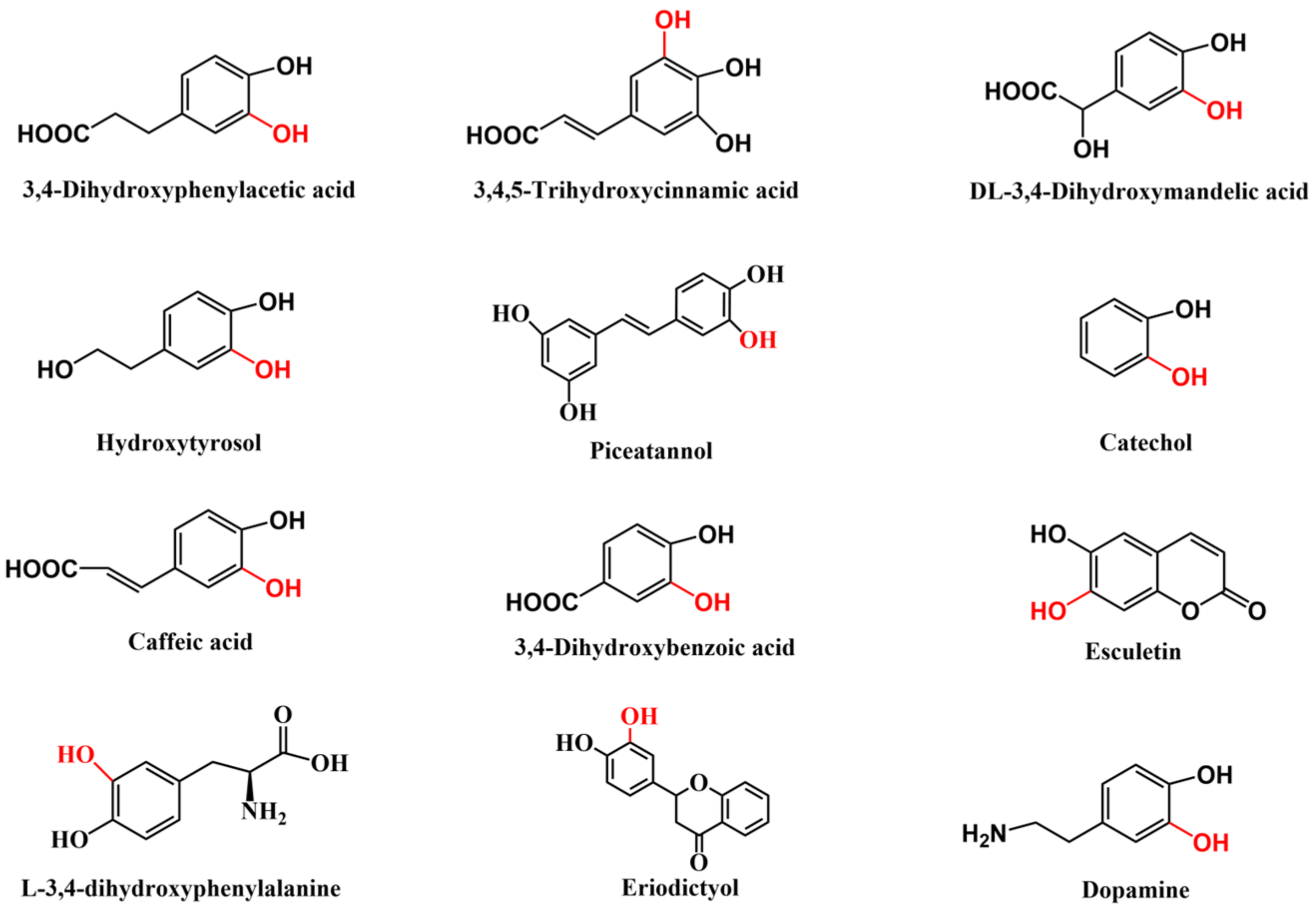
5. Applications of HpaB in the Synthesis of Catechol-Containing Compounds
5.1. Synthesis of Catechol-Containing Compounds Using HpaB-Involved Synthetic Molecular Pathways
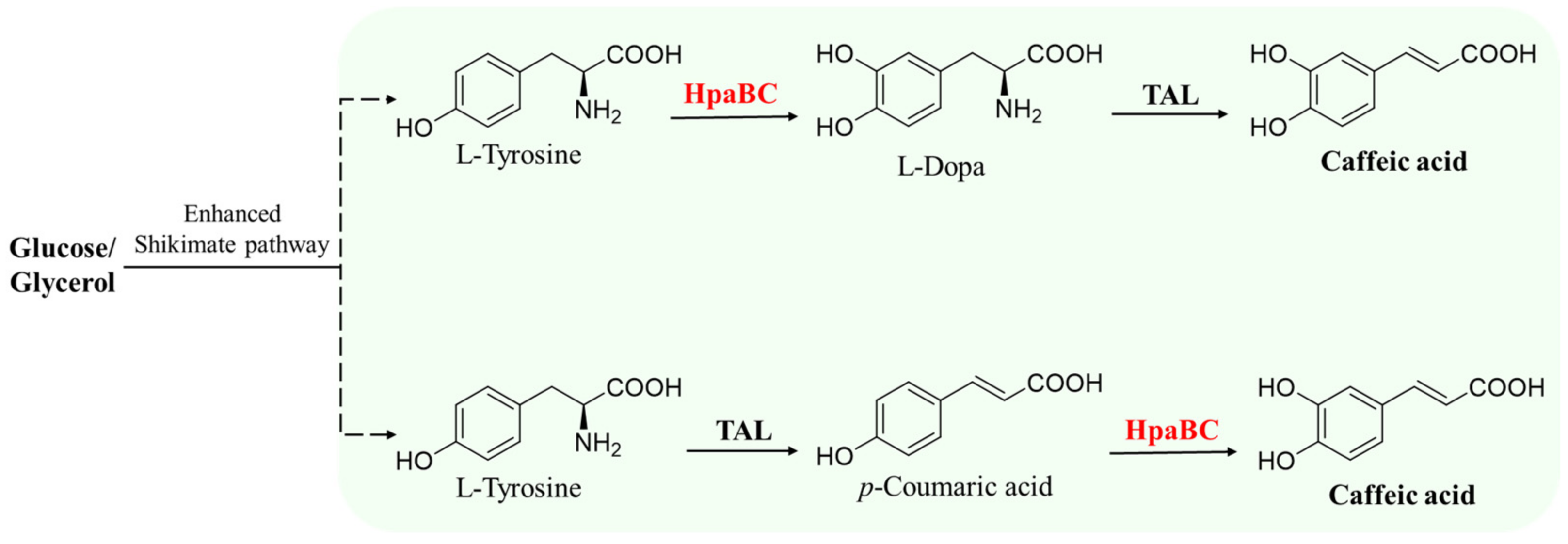
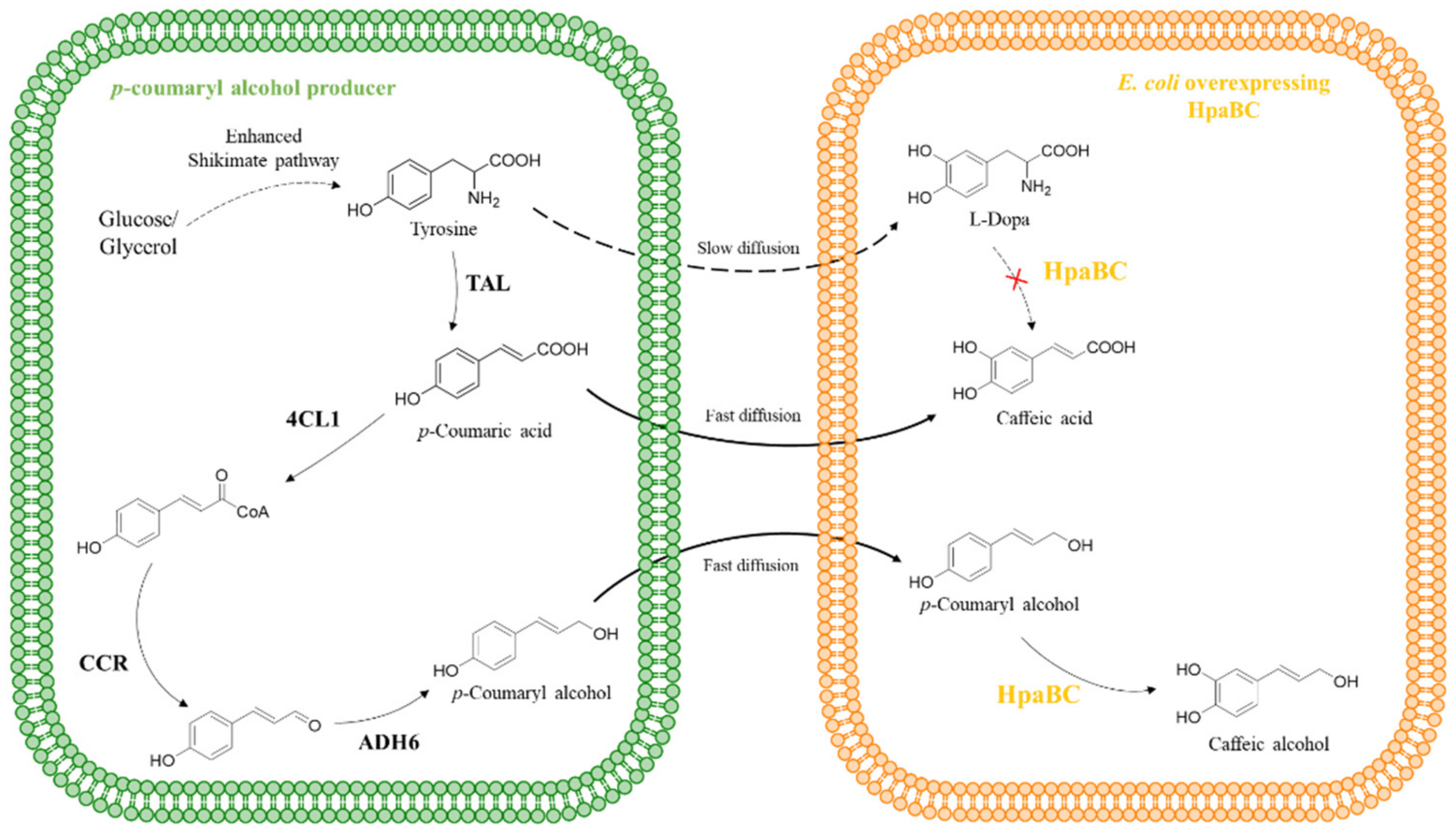
5.1.1. Synthesis of Caffeic Acid and Its Derivatives
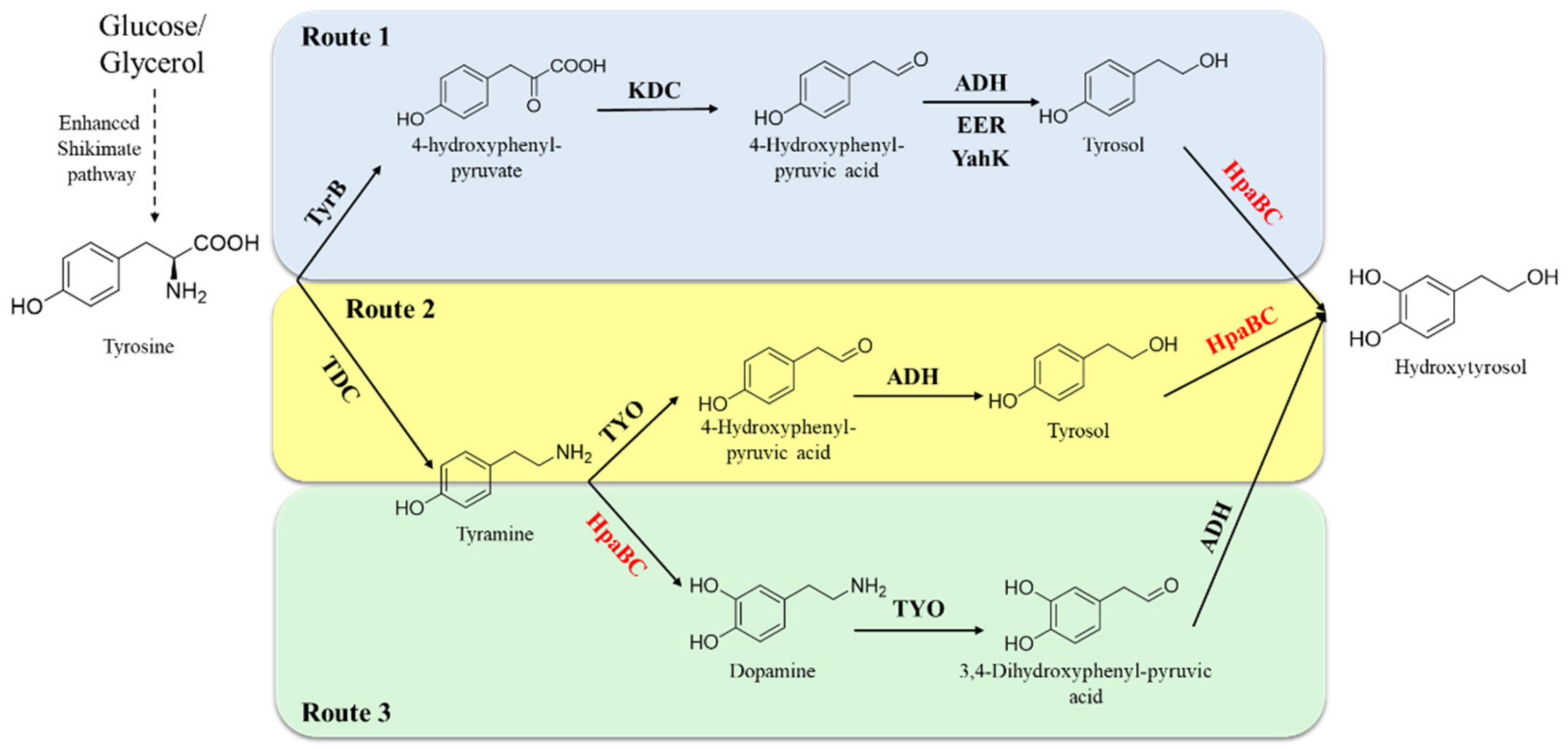
5.1.2. Synthesis of Hydroxytyrosol Acid and Its Derivatives
5.1.3. Synthesis of Other Catechol-Containing Compounds
5.2. One-Step Biotransformation to Catechol-Containing Compounds Using Direct Phenol Precursors
6. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Goldstein, D.S.; Cheshire, W.P. Roles of Catechol Neurochemistry in Autonomic Function Testing. Clin. Auton. Res. 2018, 28, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Tang, Y.; Liu, L. Adsorption Kinetics of Tea Waste to Catechins. J. Zhejiang Univ. (Agric. Life Sci.) 2014, 40, 679–687. [Google Scholar]
- Zheng, L.T.; Ryu, G.-M.; Kwon, B.-M.; Lee, W.-H.; Suk, K. Anti-Inflammatory Effects of Catechols in Lipopolysaccharide-Stimulated Microglia Cells: Inhibition of Microglial Neurotoxicity. Eur. J. Pharmacol. 2008, 588, 106–113. [Google Scholar] [CrossRef]
- Furushima, D.; Otake, Y.; Koike, N.; Onishi, S.; Mori, T.; Ota, N.; Yamada, H. Investigation of the Oral Retention of Tea Catechins in Humans: An Exploratory Interventional Study. Nutrients 2021, 13, 3024. [Google Scholar] [CrossRef]
- Li, Z.; Li, P.; Deng, C.; Luo, H. Piceatannol Inhibits Prostate Cancer Cell Proliferation, Migration and Invasion. Chin. J. Pathophysiol. 2017, 12, 1130–1133. [Google Scholar]
- Khalesi, S.; Sun, J.; Buys, N.; Jamshidi, A.; Nikbakht-Nasrabadi, E.; Khosravi-Boroujeni, H. Green Tea Catechins and Blood Pressure: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Eur. J. Nutr. 2014, 53, 1299–1311. [Google Scholar] [CrossRef]
- Liang, Y.-F.; Li, X.; Wang, X.; Zou, M.; Tang, C.; Liang, Y.; Song, S.; Jiao, N. Conversion of Simple Cyclohexanones into Catechols. J. Am. Chem. Soc. 2016, 138, 12271–12277. [Google Scholar] [CrossRef]
- Brisdelli, F.; D’Andrea, G.; Bozzi, A. Resveratrol: A Natural Polyphenol with Multiple Chemopreventive Properties. Curr. Drug Metab. 2009, 10, 530–546. [Google Scholar] [CrossRef]
- Maier, G.P.; Bernt, C.M.; Butler, A. Catechol Oxidation: Considerations in the Design of Wet Adhesive Materials. Biomater. Sci. 2018, 6, 332–339. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, D.; Geng, F.; Dong, J.; Su, L.; Zhou, Y.; Yin, S. Metal-Free Oxidative Condensation of Catechols, Aldehydes and NH4OAc towards Benzoxazoles. Adv. Synth. Catal. 2021, 363, 3607–3614. [Google Scholar] [CrossRef]
- Deng, Y.; Faivre, B.; Back, O.; Lombard, M.; Pecqueur, L.; Fontecave, M. Structural and Functional Characterization of 4-Hydroxyphenylacetate 3-Hydroxylase from Escherichia coli. ChemBioChem 2020, 21, 163–170. [Google Scholar] [CrossRef]
- Zambrano, J.L.; Dorta, R. Improving the Dakin Reactionby Using an Ionic Liquid Solvent. Synlett 2003, 2003, 1545–1546. [Google Scholar] [CrossRef]
- Saha, D.; Das, P.; Biswas, P.; Guin, J. Synthesis of Phenolic Compounds via Palladium Catalyzed C-H Functionalization of Arenes. Chem. Asian J. 2019, 14, 4534–4548. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yan, D.; Chen, Y.; Wang, T.; Xiong, F.; Wei, W.; Lu, Y.; Sun, W.-Y.; Li, J.J.; Zhao, J. A Redox-Neutral Catechol Synthesis. Nat. Commun. 2017, 8, 14227. [Google Scholar] [CrossRef]
- Lin, Y.; Yan, Y. Biotechnological Production of Plant-Specific Hydroxylated Phenylpropanoids. Biotechnol. Bioeng. 2014, 111, 1895–1899. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Lin, Y.; Yan, Y. Caffeic Acid Production Enhancement by Engineering a Phenylalanine Over-Producing Escherichia coli Strain. Biotechnol. Bioeng. 2013, 110, 3188–3196. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Guo, H.Y.; Zhou, Q.F. Construction of 4-Hydroxyphenylacetate-3-hydroxylase A Expression Strain and Its Biotransformation Effect on Hydroxytyrosol. China Biotechnol. 2015, 35, 56–60. [Google Scholar]
- Lee, N.; Kim, E.J.; Kim, B.-G. Regioselective Hydroxylation of Trans-Resveratrol via Inhibition of Tyrosinase from Streptomyces Avermitilis MA4680. ACS Chem. Biol. 2012, 7, 1687–1692. [Google Scholar] [CrossRef]
- Pandey, B.P.; Lee, N.; Choi, K.-Y.; Jung, E.; Jeong, D.-H.; Kim, B.-G. Screening of Bacterial Cytochrome P450s Responsible for Regiospecific Hydroxylation of (Iso)Flavonoids. Enzym. Microb. Technol. 2011, 48, 386–392. [Google Scholar] [CrossRef]
- Fayad, N.; Marchal, L.; Billaud, C.; Nicolas, J. Comparison of Beta-Cyclodextrin Effect on Polyphenol Oxidation Catalyzed by Purified Polyphenol Oxidase from Different Sources. J. Agric. Food Chem. 1997, 45, 2442–2446. [Google Scholar] [CrossRef]
- Lee, P.-G.; Lee, S.-H.; Hong, E.Y.; Lutz, S.; Kim, B.-G. Circular Permutation of a Bacterial Tyrosinase Enables Efficient Polyphenol-Specific Oxidation and Quantitative Preparation of Orobol. Biotechnol. Bioeng. 2019, 116, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Xun, L.; Sandvik, E.R. Characterization of 4-Hydroxyphenylacetate 3-Hydroxylase (HpaB) of Escherichia coli as a Reduced Flavin Adenine Dinucleotide-Utilizing Monooxygenase. Appl. Environ. Microbiol. 2000, 66, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Wu, C.-Z.; Kang, S.Y.; Ahn, J.S.; Uhm, T.-B.; Hong, Y.-S. Biosynthesis of Plant-Specific Phenylpropanoids by Construction of an Artificial Biosynthetic Pathway in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2011, 38, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, W.; Takahashi, H.; Kojima, M. Production of Diphenols by Polyphenol Oxidase from Mushroom and Mung Bean (Vigna mungo). Nippon. Shokuhin Kagaku Kogaku Kaishi 1996, 43, 875–879. [Google Scholar] [CrossRef]
- Selinheimo, E.; Gasparetti, C.; Mattinen, M.-L.; Steffensen, C.L.; Buchert, J.; Kruus, K. Comparison of Substrate Specificity of Tyrosinases from Trichoderma reesei and Agaricus bisporus. Enzym. Microb. Technol. 2009, 44, 1–10. [Google Scholar] [CrossRef]
- Selinheimo, E.; NiEidhin, D.; Steffensen, C.; Nielsen, J.; Lomascolo, A.; Halaouli, S.; Record, E.; O’Beirne, D.; Buchert, J.; Kruus, K. Comparison of the Characteristics of Fungal and Plant Tyrosinases. J. Biotechnol. 2007, 130, 471–480. [Google Scholar] [CrossRef]
- Fernández-Medarde, A.; Luengo, J.M. Purification and Characterization of the 4-Hydroxyphenylacetic Acid-3-Hydroxylase from Pseudomonas Putida U. FEMS Microbiol. Lett. 1997, 157, 47–53. [Google Scholar] [CrossRef]
- Zhang, H.; Lin, J.; Hu, D.; Liu, G.; Sun, L. High-density Fermentation of Escherichia coli to Express 4-Hydroxyphenylacetate 3-Hydroxylase and Efficient Biosynthesis of Caffeic Acid. Chin. J. Biotechnol. 2022, 38, 3466–3477. [Google Scholar]
- Shen, X.; Zhou, D.; Lin, Y.; Wang, J.; Gao, S.; Kandavelu, P.; Zhang, H.; Zhang, R.; Wang, B.-C.; Rose, J.; et al. Structural Insights into Catalytic Versatility of the Flavin-Dependent Hydroxylase (HpaB) from Escherichia coli. Sci. Rep. 2019, 9, 7087. [Google Scholar] [CrossRef]
- Dhammaraj, T.; Phintha, A.; Pinthong, C.; Medhanavyn, D.; Tinikul, R.; Chenprakhon, P.; Sucharitakul, J.; Vardhanabhuti, N.; Jiarpinitnun, C.; Chaiyen, P. p-Hydroxyphenylacetate 3-Hydroxylase as a Biocatalyst for the Synthesis of Trihydroxyphenolic Acids. ACS Catal. 2015, 5, 4492–4502. [Google Scholar] [CrossRef]
- Coulombel, L.; Nolan, L.C.; Nikodinovic, J.; Doyle, E.M.; O’Connor, K.E. Biotransformation of 4-Halophenols to 4-Halocatechols Using Escherichia coli Expressing 4-Hydroxyphenylacetate 3-Hydroxylase. Appl. Microbiol. Biotechnol. 2011, 89, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, X.; Ruan, H.; Chen, Y.; Gao, L.; Lei, T.; Li, Y.; Gui, L.; Guo, L.; Xia, T.; et al. Optimization of the Biosynthesis of B-Ring Ortho-Hydroxy Lated Flavonoids Using the 4-Hydroxyphenylacetate 3-Hydroxylase Complex (HpaBC) of Escherichia coli. Molecules 2021, 26, 2919. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yao, J.; Meng, J.; Han, W.; Tao, Y.; Chen, Y.; Guo, Y.; Shi, G.; He, Y.; Jin, J.-M.; et al. Promiscuous Enzymatic Activity-Aided Multiple-Pathway Network Design for Metabolic Flux Rearrangement in Hydroxytyrosol Biosynthesis. Nat. Commun. 2019, 10, 960. [Google Scholar] [CrossRef]
- Prieto, M.A.; Garcia, J.L. Molecular Characterization of 4-Hydroxyphenylacetate 3-Hydroxylase of Escherichia coli. A Two-Protein Component Enzyme. J. Biol. Chem. 1994, 269, 22823–22829. [Google Scholar] [CrossRef] [PubMed]
- Chaiyen, P.; Suadee, C.; Wilairat, P. A Novel Two-Protein Component Flavoprotein Hydroxylase. Eur. J. Biochem. 2001, 268, 5550–5561. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, S.; Dippe, M.; Pecher, P.; Funke, E.; Pietzsch, M.; Wessjohann, L.A. Engineered Bacterial Flavin-Dependent Monooxygenases for the Regiospecific Hydroxylation of Polycyclic Phenols. Chembiochem 2022, 23, e202100480. [Google Scholar] [CrossRef]
- Qi, L.; Liu, C.; Peplowski, L.; Shen, W.; Yang, H.; Xia, Y.; Chen, X. Efficient Production of Hydroxytyrosol by Directed Evolution of HpaB in Escherichia coli. Biochem. Biophys. Res. Commun. 2023, 663, 16–24. [Google Scholar] [CrossRef]
- Thotsaporn, K.; Sucharitakul, J.; Wongratana, J.; Suadee, C.; Chaiyen, P. Cloning and Expression of p-Hydroxyphenylacetate 3-Hydroxylase from Acinetobacter Baumannii: Evidence of the Divergence of Enzymes in the Class of Two-Protein Component Aromatic Hydroxylases. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 2004, 1680, 60–66. [Google Scholar] [CrossRef]
- Yao, J.; He, Y.; Su, N.; Bharath, S.R.; Tao, Y.; Jin, J.-M.; Chen, W.; Song, H.; Tang, S.-Y. Developing a Highly Efficient Hydroxytyrosol Whole-Cell Catalyst by de-Bottlenecking Rate-Limiting Steps. Nat. Commun. 2020, 11, 1515. [Google Scholar] [CrossRef]
- Imagawa, T.; Tsurumura, T.; Sugimoto, Y.; Aki, K.; Ishidoh, K.; Kuramitsu, S.; Tsuge, H. Structural Basis of Free Reduced Flavin Generation by Flavin Reductase from Thermus thermophilus HB8. J. Biol. Chem. 2011, 286, 44078–44085. [Google Scholar] [CrossRef]
- Chakraborty, S.; Ortiz-Maldonado, M.; Entsch, B.; Ballou, D.P. Studies on the Mechanism of p-Hydroxyphenylacetate 3-Hydroxylase from Pseudomonas Aeruginosa: A System Composed of a Small Flavin Reductase and a Large Flavin-Dependent Oxygenase. Biochemistry 2010, 49, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, S.; Wang, J.; Shen, X.; Feng, X.; Yuan, S.; Sun, X.; Yuan, Q. Engineering a Prokaryotic Non-P450 Hydroxylase for 3′-Hydroxylation of Flavonoids. ACS Synth. Biol. 2022, 11, 3865–3873. [Google Scholar] [CrossRef] [PubMed]
- Hawumba, J.F.; Brözel, V.S.; Theron, J. Cloning and Characterization of a 4-Hydroxyphenylacetate 3-Hydroxylase from the Thermophile geobacillus sp. PA-9. Curr. Microbiol. 2007, 55, 480–484. [Google Scholar] [CrossRef]
- Gibello, A.; Suárez, M.; Allende, J.L.; Martín, M. Molecular Cloning and Analysis of the Genes Encoding the 4-Hydroxyphenylacetate Hydroxylase from Klebsiella Pneumoniae. Arch. Microbiol. 1997, 167, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Hisano, T.; Takeda, K.; Iwasaki, W.; Ebihara, A.; Miki, K. Crystal Structure of the Oxygenase Component (HpaB) of the 4-Hydroxyphenylacetate 3-Monooxygenase from Thermus thermophilus HB8. J. Biol. Chem. 2007, 282, 33107–33117. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, A.; Fersini, F.; Ruangchan, N.; Prongjit, M.; Chaiyen, P.; Mattevi, A. Structure of the Monooxygenase Component of a Two-Component Flavoprotein Monooxygenase. Proc. Natl. Acad. Sci. USA 2007, 104, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Heine, T.; van Berkel, W.J.H.; Gassner, G.; van Pée, K.-H.; Tischler, D. Two-Component FAD-Dependent Monooxygenases: Current Knowledge and Biotechnological Opportunities. Biology 2018, 7, 42. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Oonanant, W.; Sucharitakul, J.; Chaiyen, P.; Yuvaniyama, J. Crystallization and Preliminary X-Ray Analysis of the Reductase Component of p-Hydroxyphenylacetate 3-Hydroxylase from Acinetobacter Baumannii. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012, 68, 720–723. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering Key Features in Protein Structures with the New ENDscript Server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Sucharitakul, J.; Chaiyen, P.; Entsch, B.; Ballou, D.P. Kinetic Mechanisms of the Oxygenase from a Two-Component Enzyme, p-Hydroxyphenylacetate 3-Hydroxylase from Acinetobacter Baumannii. J. Biol. Chem. 2006, 281, 17044–17053. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jin, Y.; Yang, K.; Hu, S.; Lv, C.; Huang, J.; Mei, J.; Zhao, W.; Mei, L. Modification of the 4-Hydroxyphenylacetate-3-Hydroxylase Substrate Pocket to Increase Activity towards Resveratrol. Molecules 2023, 28, 5602. [Google Scholar] [CrossRef] [PubMed]
- Chenprakhon, P.; Dhammaraj, T.; Chantiwas, R.; Chaiyen, P. Hydroxylation of 4-Hydroxyphenylethylamine Derivatives by R263 Variants of the Oxygenase Component of p-Hydroxyphenylacetate-3-Hydroxylase. Arch. Biochem. Biophys. 2017, 620, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dhammaraj, T.; Pinthong, C.; Visitsatthawong, S.; Tongsook, C.; Surawatanawong, P.; Chaiyen, P. A Single-Site Mutation at Ser146 Expands the Reactivity of the Oxygenase Component of p-Hydroxyphenylacetate 3-Hydroxylase. ACS Chem. Biol. 2016, 11, 2889–2896. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yan, Y. Biosynthesis of Caffeic Acid in Escherichia coli Using Its Endogenous Hydroxylase Complex. Microb. Cell Fact. 2012, 11, 42. [Google Scholar] [CrossRef]
- Liu, L.; Liu, H.; Zhang, W.; Yao, M.; Li, B.; Liu, D.; Yuan, Y. Engineering the Biosynthesis of Caffeic Acid in Saccharomyces cerevisiae with Heterologous Enzyme Combinations. Engineering 2019, 5, 287–295. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, X.; Li, Y.; Yan, Y.; Yuan, Q. Metabolic Engineering of Escherichia coli for Microbial Synthesis of Monolignols. Metab. Eng. 2017, 39, 102–109. [Google Scholar] [CrossRef]
- Li, X.; Chen, Z.; Wu, Y.; Yan, Y.; Sun, X.; Yuan, Q. Establishing an Artificial Pathway for Efficient Biosynthesis of Hydroxytyrosol. ACS Synth. Biol. 2018, 7, 647–654. [Google Scholar] [CrossRef]
- Choo, H.J.; Kim, E.J.; Kim, S.Y.; Lee, Y.; Kim, B.-G.; Ahn, J.-H. Microbial Synthesis of Hydroxytyrosol and Hydroxysalidroside. Appl. Biol. Chem. 2018, 61, 295–301. [Google Scholar] [CrossRef]
- Guo, D.; Fu, X.; Sun, Y.; Li, X.; Pan, H. De Novo Biosynthesis of Tyrosol Acetate and Hydroxytyrosol Acetate from Glucose in Engineered Escherichia coli. Enzym. Microb. Technol. 2021, 150, 109886. [Google Scholar] [CrossRef]
- Fordjour, E.; Adipah, F.K.; Zhou, S.; Du, G.; Zhou, J. Metabolic Engineering of Escherichia coli BL21 (DE3) for de Novo Production of l-DOPA from d-Glucose. Microb. Cell Fact. 2019, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Harnessing Probiotics to Deliver Parkinson’s Disease Treatment-Neuroscience News. Available online: https://neurosciencenews.com/parkinsons-probiotic-20287/ (accessed on 27 August 2023).
- Li, X.; Zhou, W.; Zhuang, Y.; Lu, F.; Yin, H. Biosynthesis of 3, 4-dihydroxymandelic Acid in an Engineered Escherichia coli Strain. Biotechnol. Bulletie 2017, 33, 135–140. [Google Scholar]
- Yao, Y.-F.; Wang, C.-S.; Qiao, J.; Zhao, G.-R. Metabolic Engineering of Escherichia coli for Production of Salvianic Acid A via an Artificial Biosynthetic Pathway. Metab. Eng. 2013, 19, 79–87. [Google Scholar] [CrossRef]
- Wu, X.; Liu, J.; Liu, D.; Yuwen, M.; Koffas, M.A.G.; Zha, J. Biosynthesis of Eriodictyol from Tyrosine by Corynebacterium Glutamicum. Microb. Cell Fact. 2022, 21, 86. [Google Scholar] [CrossRef]
- Jones, J.A.; Collins, S.M.; Vernacchio, V.R.; Lachance, D.M.; Koffas, M.A.G. Optimization of Naringenin and P-Coumaric Acid Hydroxylation Using the Native E. Coli Hydroxylase Complex, HpaBC. Biotechnol. Prog. 2016, 32, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Kino, K. Regioselective Synthesis of Piceatannol from Resveratrol: Catalysis by Two-Component Flavin-Dependent Monooxygenase HpaBC in Whole Cells. Tetrahedron Lett. 2014, 55, 2853–2855. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, K.; Zhang, Q.; Zhao, W.; Hu, S.; Lv, C.; Huang, J.; Mei, J.; Mei, L. Advances in 4-Hydroxyphenylacetate-3-hydroxylase Monooxygenase. Molecules 2023, 28, 6699. https://doi.org/10.3390/molecules28186699
Yang K, Zhang Q, Zhao W, Hu S, Lv C, Huang J, Mei J, Mei L. Advances in 4-Hydroxyphenylacetate-3-hydroxylase Monooxygenase. Molecules. 2023; 28(18):6699. https://doi.org/10.3390/molecules28186699
Chicago/Turabian StyleYang, Kai, Qianchao Zhang, Weirui Zhao, Sheng Hu, Changjiang Lv, Jun Huang, Jiaqi Mei, and Lehe Mei. 2023. "Advances in 4-Hydroxyphenylacetate-3-hydroxylase Monooxygenase" Molecules 28, no. 18: 6699. https://doi.org/10.3390/molecules28186699
APA StyleYang, K., Zhang, Q., Zhao, W., Hu, S., Lv, C., Huang, J., Mei, J., & Mei, L. (2023). Advances in 4-Hydroxyphenylacetate-3-hydroxylase Monooxygenase. Molecules, 28(18), 6699. https://doi.org/10.3390/molecules28186699