
Effects of Microplastic (MP) Exposure at Environmentally Relevant Doses on the Structure, Function, and Transcriptome of the Kidney in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of PS-MP Exposure on Weight Gain and Kidney Coefficient and Function
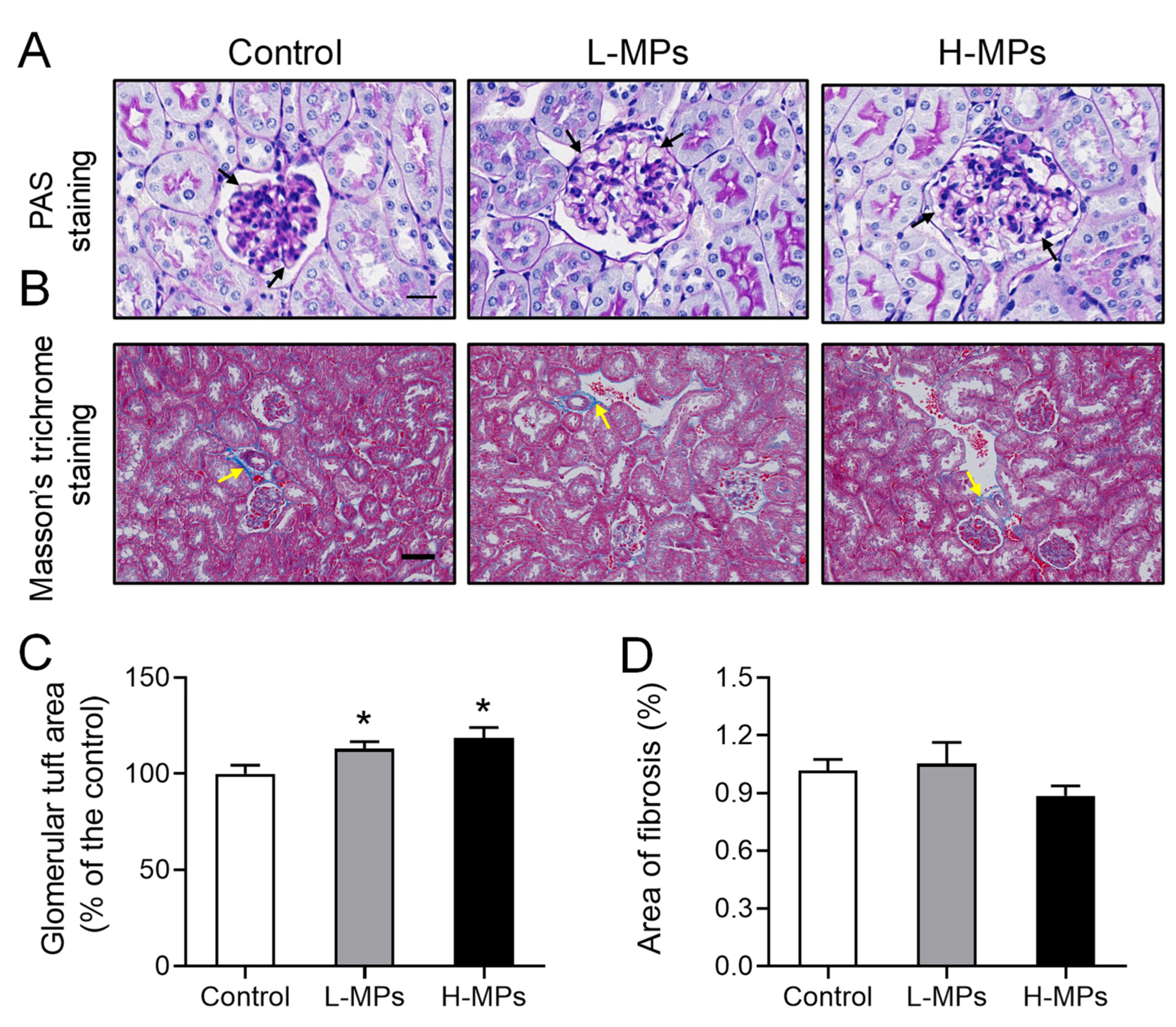
2.2. Effects of PS-MP Exposure on Kidney Histopathology
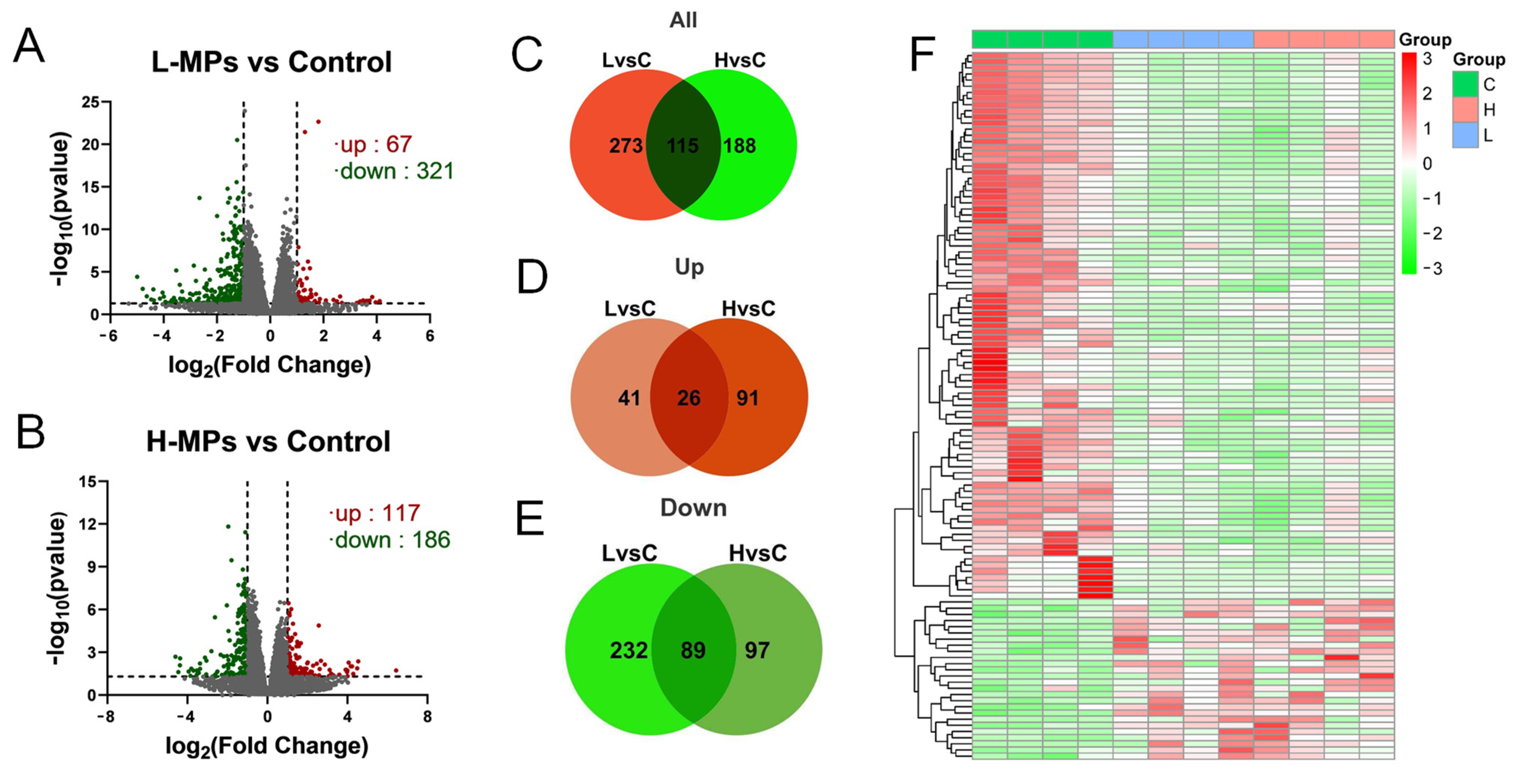
2.3. Effects of PS-MP Exposure on Kidney Transcriptome
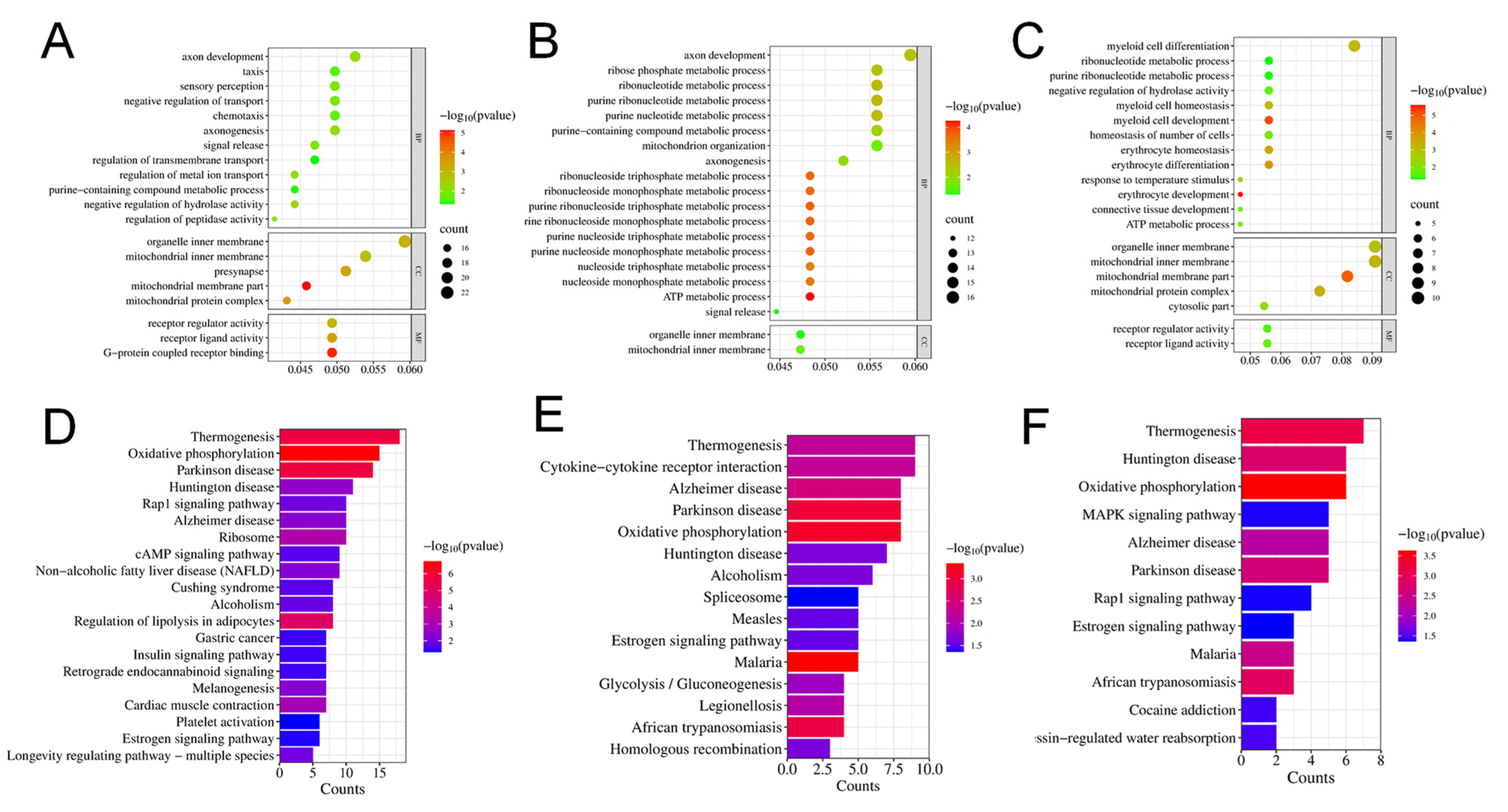
2.4. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis of Enrichment of Differentially Expressed Genes
2.5. Protein–Protein Interaction (PPI) Network Analysis and Hub Gene Identification
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Mice and Treatments
4.3. Routine Kidney Function Blood Test
4.4. Kidney Histopathology
4.5. RNA Extraction and Sequencing
4.6. Sequencing Data Analysis
4.7. Enrichment Analysis of Differentially Expressed Genes
4.8. PPI Analysis of Differentially Expressed Genes
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Statista Research Department. Global Plastic Production 1950–2021. Available online: https://www.statista.com/statistics/282732/global-production-of-plastics-since-1950/ (accessed on 16 May 2023).
- Wang, C.; Zhao, J.; Xing, B. Environmental source, fate, and toxicity of microplastics. J. Hazard. Mater. 2021, 407, 124357. [Google Scholar] [CrossRef] [PubMed]
- Hashmi, M.Z. Microplastic Pollution: Environmental Occurrence and Treatment Technologies; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Akdogan, Z.; Guven, B. Microplastics in the environment: A critical review of current understanding and identification of future research needs. Environ. Pollut. 2019, 254, 113011. [Google Scholar] [CrossRef] [PubMed]
- Law, K.L.; Thompson, R.C. Microplastics in the seas. Science 2014, 345, 144–145. [Google Scholar] [CrossRef]
- Sarijan, S.; Azman, S.; Said, M.; Jamal, M.H. Microplastics in freshwater ecosystems: A recent review of occurrence, analysis, potential impacts, and research needs. Environ. Sci. Pollut. Res. Int. 2021, 28, 1341–1356. [Google Scholar] [CrossRef]
- Muhib, M.I.; Uddin, M.K.; Rahman, M.M.; Malafaia, G. Occurrence of microplastics in tap and bottled water, and food packaging: A narrative review on current knowledge. Sci. Total Environ. 2023, 865, 161274. [Google Scholar] [CrossRef]
- Li, Z.; Yang, Y.; Chen, X.; He, Y.; Bolan, N.; Rinklebe, J.; Lam, S.S.; Peng, W.; Sonne, C. A discussion of microplastics in soil and risks for ecosystems and food chains. Chemosphere 2023, 313, 137637. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Simnani, F.Z.; Singh, D.; Patel, P.; Sinha, A.; Nandi, A.; Ghosh, A.; Saha, U.; Kumari, K.; Jaganathan, S.K.; et al. Atmospheric microplastic and nanoplastic: The toxicological paradigm on the cellular system. Ecotoxicol. Environ. Saf. 2023, 259, 115018. [Google Scholar] [CrossRef]
- Materic, D.; Kjaer, H.A.; Vallelonga, P.; Tison, J.L.; Rockmann, T.; Holzinger, R. Nanoplastics measurements in northern and southern polar ice. Environ. Res. 2022, 208, 112741. [Google Scholar] [CrossRef]
- Guzzetti, E.; Sureda, A.; Tejada, S.; Faggio, C. Microplastic in marine organism: Environmental and toxicological effects. Environ. Toxicol. Phar. 2018, 64, 164–171. [Google Scholar] [CrossRef]
- Wang, C.; Hou, M.; Shang, K.; Wang, H.; Wang, J. Microplastics (polystyrene) exposure induces metabolic changes in the liver of rare minnow (Gobiocypris rarus). Molecules 2022, 27, 584. [Google Scholar] [CrossRef]
- Cox, K.D.; Covernton, G.A.; Davies, H.L.; Dower, J.F.; Juanes, F.; Dudas, S.E. Human consumption of microplastics. Environ. Sci. Technol. 2019, 53, 7068–7074. [Google Scholar] [CrossRef] [PubMed]
- Prata, J.C.; Da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef] [PubMed]
- Yee, M.S.; Hii, L.W.; Looi, C.K.; Lim, W.M.; Wong, S.F.; Kok, Y.Y.; Tan, B.K.; Wong, C.Y.; Leong, C.O. Impact of microplastics and nanoplastics on human health. Nanomaterials 2021, 11, 496. [Google Scholar] [CrossRef]
- Leslie, H.A.; van Velzen, M.J.M.; Brandsma, S.H.; Vethaak, A.D.; Garcia-Vallejo, J.J.; Lamoree, M.H. Discovery and quantification of plastic particle pollution in human blood. Environ. Int. 2022, 163, 107199. [Google Scholar] [CrossRef] [PubMed]
- Amato-Lourenco, L.F.; Carvalho-Oliveira, R.; Ribeiro Junior, G.; Galvao, L.D.S.; Ando, R.A.; Mauad, T. Presence of airborne microplastics in human lung tissue. J. Hazard. Mater. 2021, 416, 126124. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, M.C.A.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef]
- Horvatits, T.; Tamminga, M.; Liu, B.; Sebode, M.; Carambia, A.; Fischer, L.; Puschel, K.; Huber, S.; Fischer, E.K. Microplastics detected in cirrhotic liver tissue. EBioMedicine 2022, 82, 104147. [Google Scholar] [CrossRef]
- Nugrahapraja, H.; Sugiyo, P.W.W.; Putri, B.Q.; Matuzahroh, N.; Fatimah; Huang, L.; Hafza, N.; Götz, F.; Santoso, H.; Wibowo, A.T.; et al. Effects of microplastic on human gut microbiome: Detection of plastic-degrading genes in human gut exposed to microplastics—preliminary study. Environments 2022, 9, 140. [Google Scholar]
- Meng, X.; Zhang, J.; Wang, W.; Gonzalez-Gil, G.; Vrouwenvelder, J.S.; Li, Z. Effects of nano- and microplastics on kidney: Physicochemical properties, bioaccumulation, oxidative stress and immunoreaction. Chemosphere 2022, 288, 132631. [Google Scholar] [CrossRef]
- Wang, Y.; Lee, Y.; Hsu, Y.; Chiu, I.; Huang, C.C.; Huang, C.; Chia, Z.; Lee, C.; Lin, Y.; Chiu, H. The kidney-related effects of polystyrene microplastics on human kidney proximal tubular epithelial cells hk-2 and male c57bl/6 mice. Environ. Health Persp. 2021, 129, 057003. [Google Scholar] [CrossRef]
- Wang, W.; Guan, J.; Feng, Y.; Nie, L.; Xu, Y.; Xu, H.; Fu, F. Polystyrene microplastics induced nephrotoxicity associated with oxidative stress, inflammation, and endoplasmic reticulum stress in juvenile rats. Front. Nutr. 2022, 9, 1059660. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Wang, Y.; Wang, M.; Zhang, D.; Liu, M. Identification of reliable reference genes for gene expression studies in mouse models under microplastics stress. Ecotoxicol. Environ. Saf. 2023, 252, 114569. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Gao, L.; Chen, C.; Zhu, K.; Luo, P.; Li, L. The microplastics exposure induce the kidney injury in mice revealed by rna-seq. Ecotoxicol. Environ. Saf. 2023, 256, 114821. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wan, Z.; Luo, T.; Fu, Z.; Jin, Y. Polystyrene microplastics induce gut microbiota dysbiosis and hepatic lipid metabolism disorder in mice. Sci. Total Environ. 2018, 631–632, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Giacoman, S.; Madero, M. Biomarkers in chronic kidney disease, from kidney function to kidney damage. World J. Nephrol. 2015, 4, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Deng, H.; Zhang, M. Whole transcriptome sequencing analysis revealed key rna profiles and toxicity in mice after chronic exposure to microplastics. Chemosphere 2022, 304, 135321. [Google Scholar] [CrossRef]
- Che, R.; Yuan, Y.; Huang, S.; Zhang, A. Mitochondrial dysfunction in the pathophysiology of renal diseases. Am. J. Physiol. Renal Physiol. 2014, 306, F367–F378. [Google Scholar] [CrossRef]
- Duann, P.; Lin, P.H. Mitochondria damage and kidney disease. Adv. Exp. Med. Biol. 2017, 982, 529–551. [Google Scholar]
- Granata, S.; Dalla, G.A.; Tomei, P.; Lupo, A.; Zaza, G. Mitochondria: A new therapeutic target in chronic kidney disease. Nutr. Metab. 2015, 12, 49. [Google Scholar] [CrossRef]
- Tanriover, C.; Copur, S.; Ucku, D.; Cakir, A.B.; Hasbal, N.B.; Soler, M.J.; Kanbay, M. The mitochondrion: A promising target for kidney disease. Pharmaceutics 2023, 15, 570. [Google Scholar] [CrossRef]
- Han, N.; Luo, R.; Liu, J.; Guo, T.; Feng, J.; Peng, X. Transcriptomic and proteomic analysis reveals mechanisms of patulin-induced cell toxicity in human embryonic kidney cells. Toxins 2020, 12, 681. [Google Scholar] [CrossRef] [PubMed]
- Varanasi, L.; Hosler, J.P. Subunit iii-depleted cytochrome c oxidase provides insight into the process of proton uptake by proteins. Biochim. Biophys. Acta 2012, 1817, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhong, H.; Chen, S.Y.; Yao, Y.G.; Liu, Y.P. Association between mt-co3 haplotypes and high-altitude adaptation in tibetan chicken. Gene 2013, 529, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wei, X.; Hu, H.; Zhang, B.; Yang, D.; Du, H.; Zhu, R.; Sun, X.; Oh, Y.; Gu, N. Effects of oral administration of polystyrene nanoplastics on plasma glucose metabolism in mice. Chemosphere 2022, 288, 132607. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, T.; Zhang, W.; Wang, Y.; Li, H.; Wu, J.; Wang, Q.; Qin, L.; Zhang, L.; Liu, C.; Li, R. Effects of Microplastic (MP) Exposure at Environmentally Relevant Doses on the Structure, Function, and Transcriptome of the Kidney in Mice. Molecules 2023, 28, 7104. https://doi.org/10.3390/molecules28207104
Shen T, Zhang W, Wang Y, Li H, Wu J, Wang Q, Qin L, Zhang L, Liu C, Li R. Effects of Microplastic (MP) Exposure at Environmentally Relevant Doses on the Structure, Function, and Transcriptome of the Kidney in Mice. Molecules. 2023; 28(20):7104. https://doi.org/10.3390/molecules28207104
Chicago/Turabian StyleShen, Ting, Wenjing Zhang, Yirun Wang, Haizhu Li, Jingwei Wu, Qian Wang, Li Qin, Lu Zhang, Cuiqing Liu, and Ran Li. 2023. "Effects of Microplastic (MP) Exposure at Environmentally Relevant Doses on the Structure, Function, and Transcriptome of the Kidney in Mice" Molecules 28, no. 20: 7104. https://doi.org/10.3390/molecules28207104
APA StyleShen, T., Zhang, W., Wang, Y., Li, H., Wu, J., Wang, Q., Qin, L., Zhang, L., Liu, C., & Li, R. (2023). Effects of Microplastic (MP) Exposure at Environmentally Relevant Doses on the Structure, Function, and Transcriptome of the Kidney in Mice. Molecules, 28(20), 7104. https://doi.org/10.3390/molecules28207104