
Influence of the Enzymatic Hydrolysis Using Flavourzyme Enzyme on Functional, Secondary Structure, and Antioxidant Characteristics of Protein Hydrolysates Produced from Bighead Carp (Hypophthalmichthys nobilis)


Abstract
1. Introduction
2. Results and Discussion
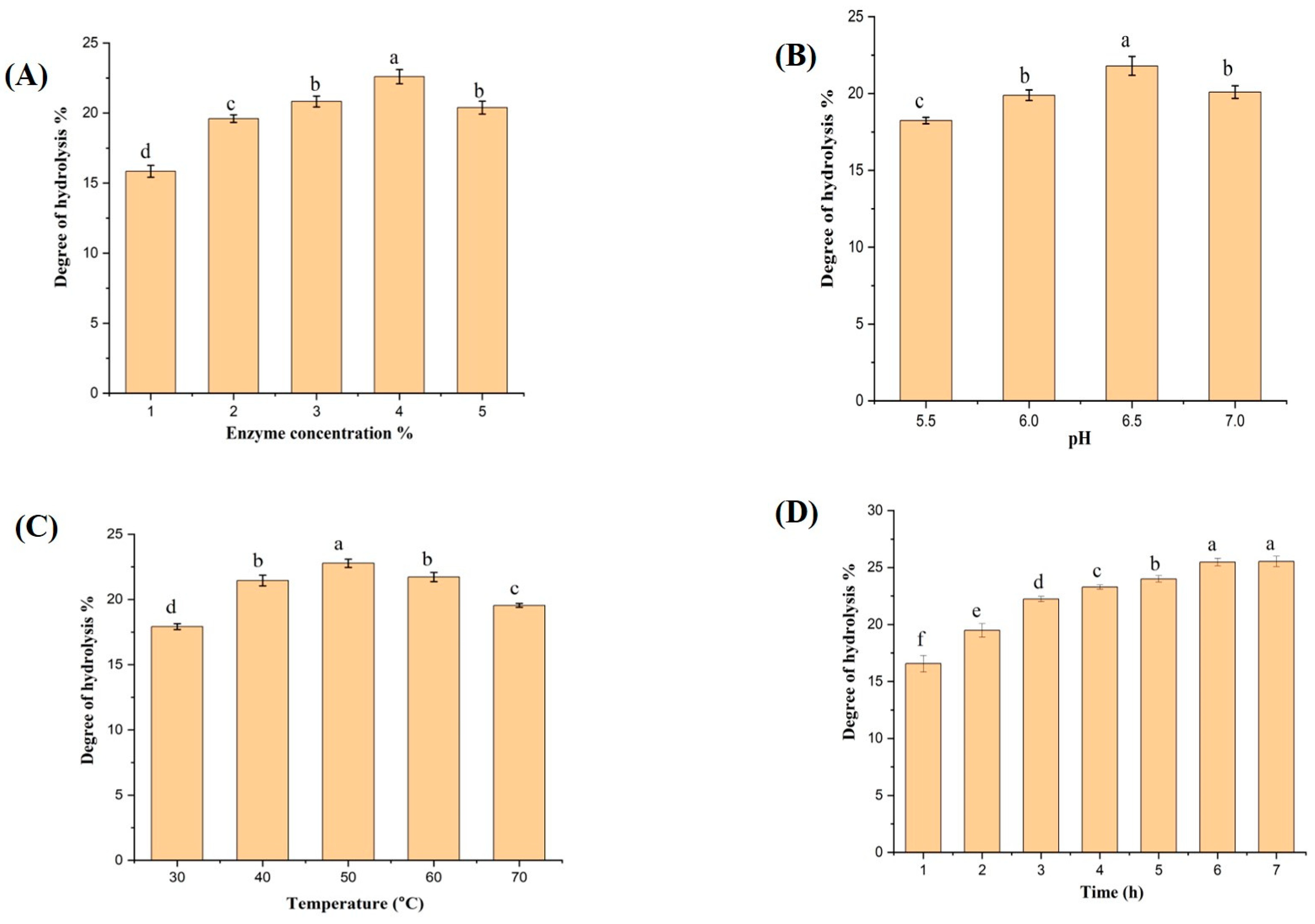
2.1. Optimization of Enzymatic Hydrolysis Factors
2.2. Proximate Analysis of Protein Hydrolysates
2.3. Yield Measurement
2.4. Color Determination
2.5. Water Activity
2.6. Amino Acid Composition
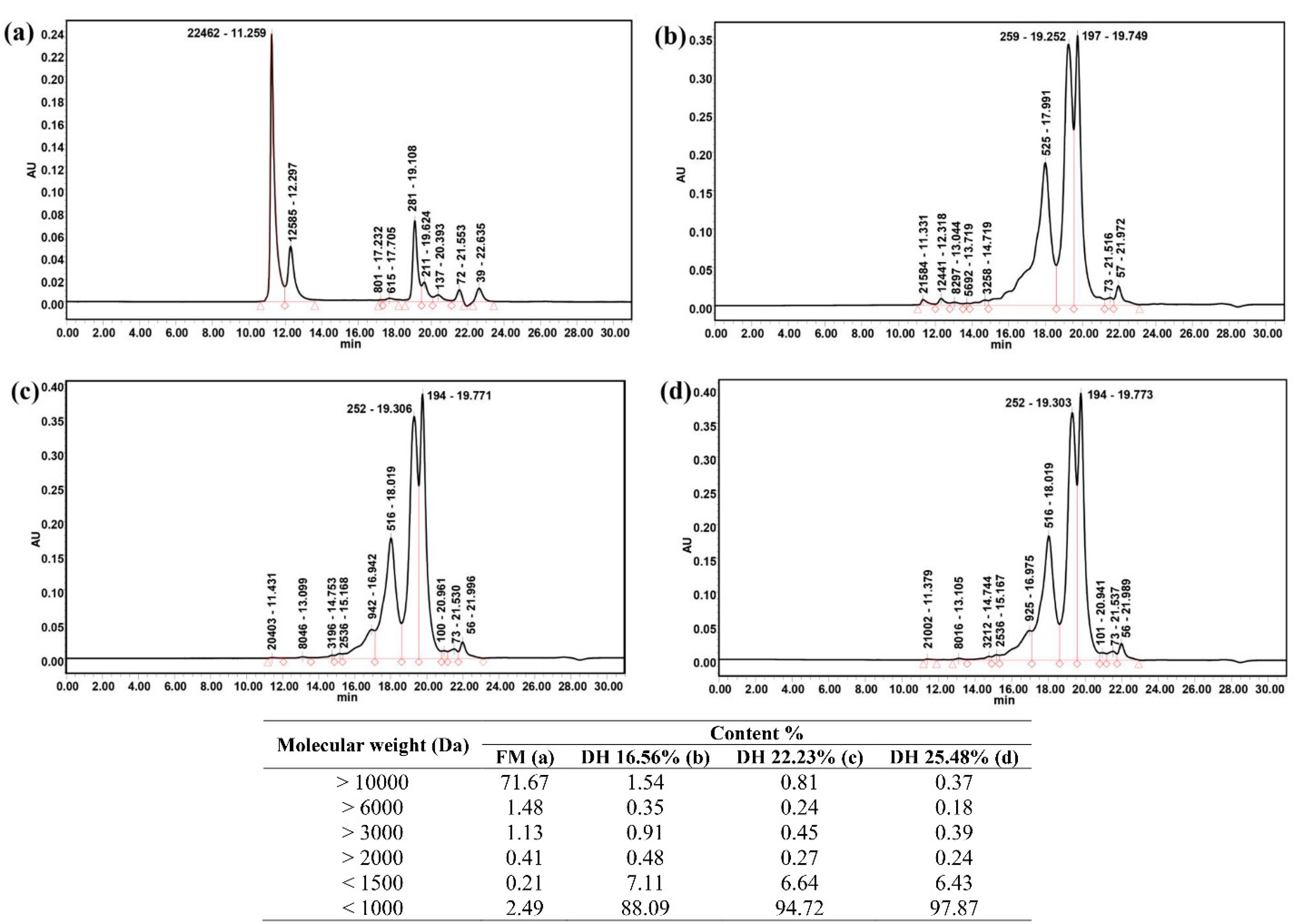
2.7. Molecular Mass Distribution
2.8. Scanning Electron Microscopy Evaluation
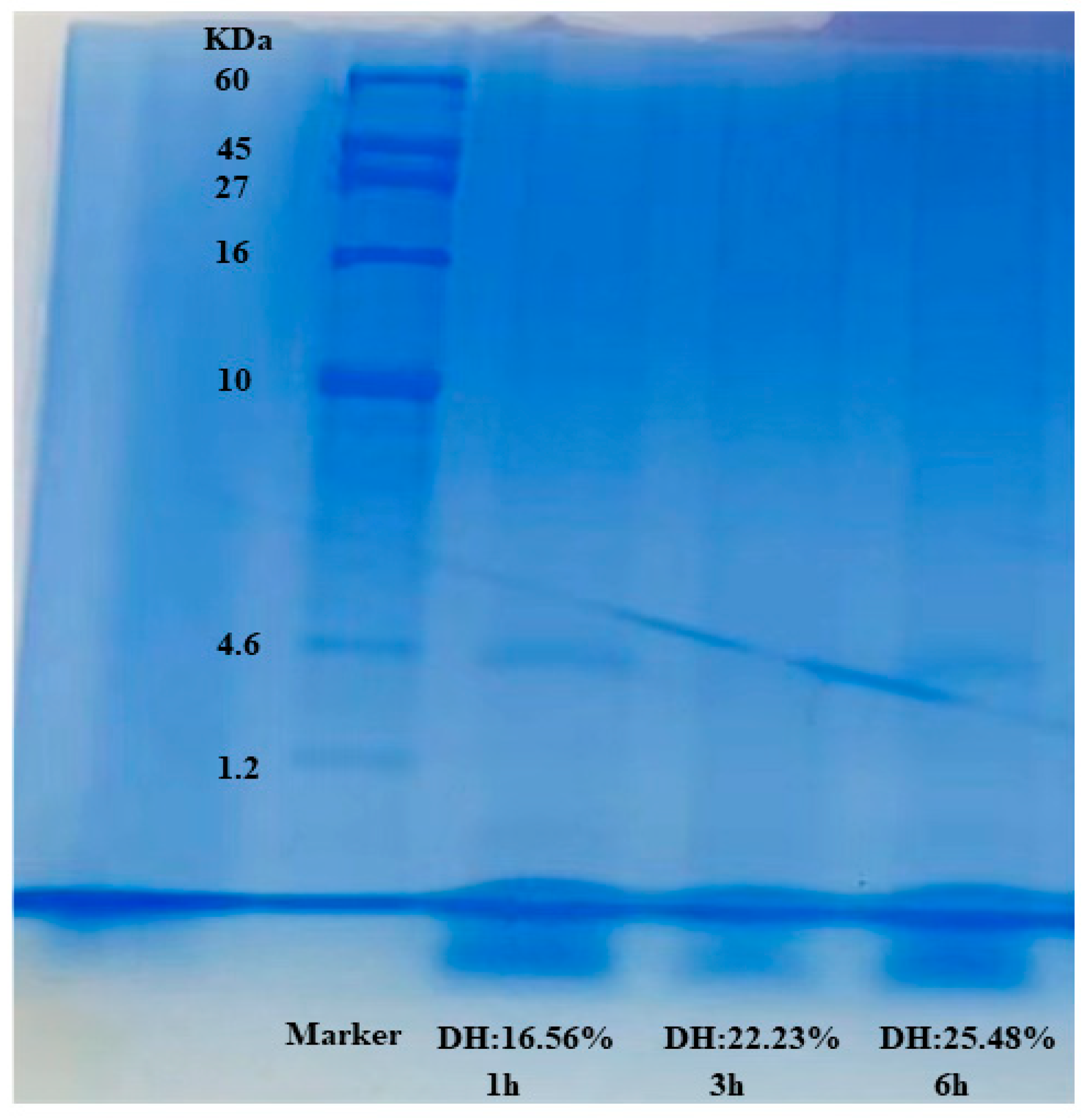
2.9. Electrophoretic Profile Analysis
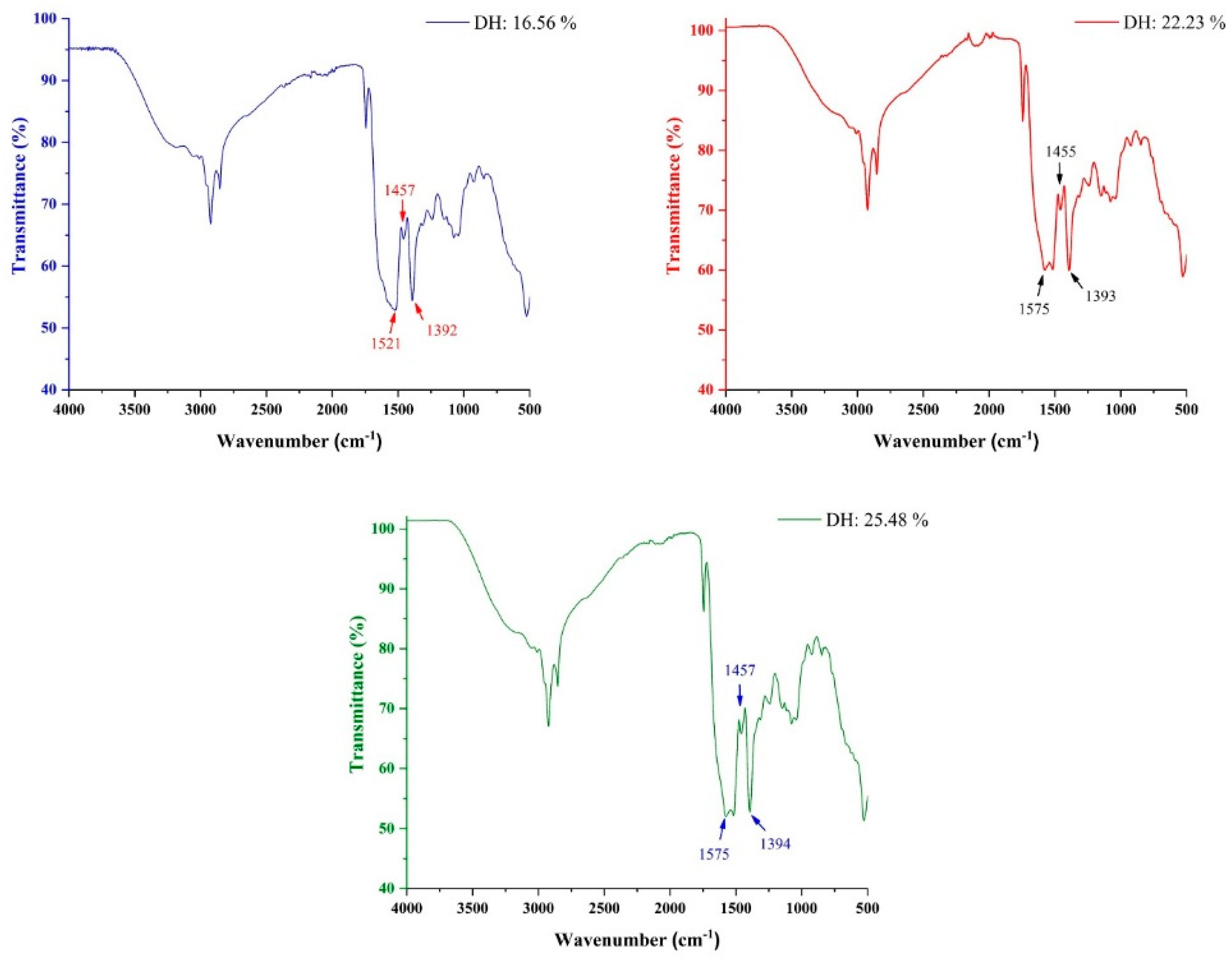
2.10. FTIR Spectra of Hydrolystes
2.11. Functional Properties of FPH
2.11.1. Protein Hydrolysate Solubility
2.11.2. Water- and Oil-Holding Capacity
2.12. Circular Dichroism Spectra
2.13. Antioxidant Activities of Fish Protein Hydrolysates
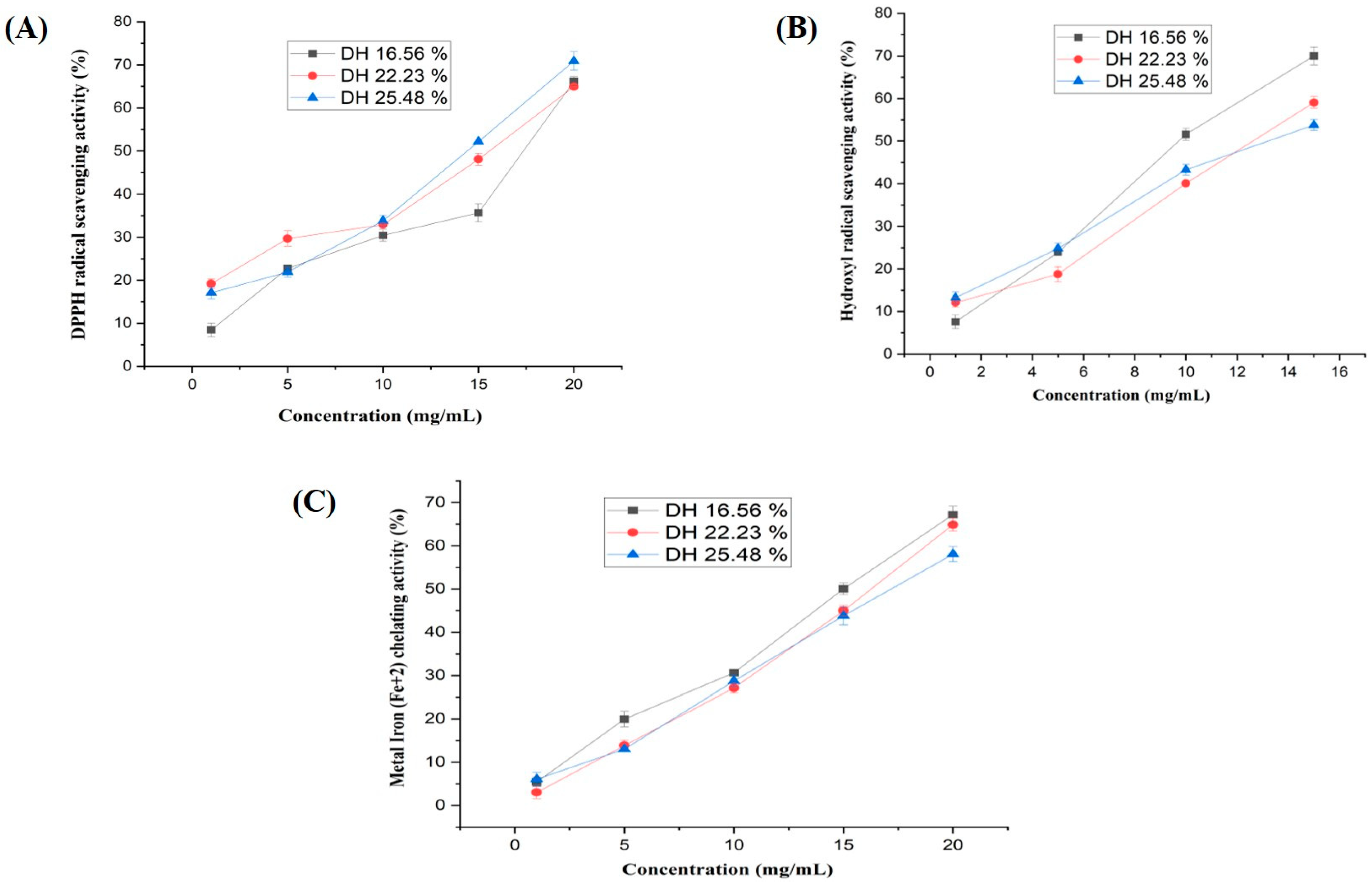
2.13.1. DPPH Radical-Scavenging Activity
2.13.2. Hydroxyl Radical-Scavenging Activity
2.13.3. Metal Iron (Fe+2) Chelating Activity
3. Materials and Methods
3.1. Preparation of Raw Materials
3.2. Enzyme and Chemicals
3.3. The Production Process of Protein Hydrolysates
3.4. Determination of Degree of Hydrolysis
3.5. Yield
3.6. Chemical Composition Analysis
3.7. Water Activity
3.8. Color Measurement
3.9. Amino Acid Determination
3.10. Molecular Mass Distribution Analysis
3.11. Electrophoretic Profile Using SDS-Page
3.12. Scanning Electron Microscopy
3.13. Fourier Transform Infrared Spectroscopy
3.14. Functional Characteristics of FPH
3.14.1. Solubility
3.14.2. Water-Holding Capacity
3.14.3. Oil-Holding Capacity
3.15. Protein Secondary Structure
3.16. Antioxidant Properties of FPH
3.16.1. DPPH Radical-Scavenging Activity
3.16.2. (Fe2+) Chelating Activity
3.16.3. Hydroxyl Radical Scavenging Assay
3.17. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- FAO. Fishery and Aquaculture Statistics; Food and Agriculture Organization of the United Nations: Rome, Italy, 2016; p. 11. [Google Scholar]
- Shi, X.; Zhang, J.; Shi, C.; Tan, Y.; Hong, H.; Luo, Y. Nondestructive prediction of freshness for bighead carp (Hypophthalmichthys nobilis) head by Excitation-Emission Matrix (EEM) analysis based on fish eye fluid: Comparison of BPNNs and RBFNNs. Food Chem. 2022, 382, 132341. [Google Scholar] [CrossRef]
- Cupp, A.R.; Erickson, R.A.; Fredricks, K.T.; Swyers, N.M.; Hatton, T.W.; Amberg, J.J. Responses of Invasive Silver and Bighead Carp to a Carbon Dioxide Barrier in Outdoor Ponds. Can. J. Fish. Aquat. Sci. 2017, 74, 297–305. [Google Scholar] [CrossRef]
- Ashraf, S.A.; Adnan, M.; Patel, M.; Siddiqui, A.J.; Sachidanandan, M.; Snoussi, M.; Hadi, S. Fish-based bioactives as potent nutraceuticals: Exploring the therapeutic perspective of sustainable food from the sea. Mar. Drugs 2020, 18, 265. [Google Scholar] [CrossRef] [PubMed]
- Chukwu, O. Influences of drying methods on nutritional properties of tilapia fish (Oreochromis nilotieus). World J. Agric. Sci. 2009, 5, 256–258. [Google Scholar]
- Kristinsson, H.G.; Rasco, B.A. Fish protein hydrolysates: Production, biochemical, and functional properties. Crit. Rev. Food Sci. Nutr. 2000, 40, 43–81. [Google Scholar] [CrossRef]
- Alahmad, K.; Xia, W.; Jiang, Q.; Xu, Y. Effect of the Degree of Hydrolysis on Nutritional, Functional, and Morphological Characteristics of Protein Hydrolysate Produced from Bighead Carp (Hypophthalmichthys nobilis) Using Ficin Enzyme. Foods 2022, 11, 1320. [Google Scholar] [CrossRef]
- Noman, A.; Xu, Y.; AL-Bukhaiti, W.Q.; Abed, S.M.; Ali, A.H.; Ramadhan, A.H.; Xia, W. Influence of enzymatic hydrolysis conditions on the degree of hydrolysis and functional properties of protein hydrolysate obtained from Chinese sturgeon (Acipenser sinensis) by using papain enzyme. Process Biochem. 2018, 67, 19–28. [Google Scholar] [CrossRef]
- Petreu, T.; Cotrutz, C.E.; Neamtu, M.; Maria Filioreanu, A.; Badescu, L.; Neamtu, A. Bioinformatic lookup for metalloproteinases and their natural inhibitors. Structural relationships. Ann. Rom. Soc. Cell Biol. 2011, 16, 175–180. [Google Scholar]
- Ismail, B.; Mohammed, H.; Nair, A.J. Influence of proteases on functional properties of food. In Green Bio-Processes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 31–53. [Google Scholar]
- Fadimu, G.J.; Gill, H.; Farahnaky, A.; Truong, T. Investigating the impact of ultrasound pretreatment on the physicochemical, structural, and antioxidant properties of lupin protein hydrolysates. Food Bioprocess Technol. 2021, 14, 2004–2019. [Google Scholar] [CrossRef]
- Wasswa, J.; Tang, J.; Gu, X.H.; Yuan, X.Q. Influence of the extent of enzymatic hydrolysis on the functional properties of protein hydrolysate from grass carp (Ctenopharyngodon idella) skin. Food Chem. 2007, 104, 1698–1704. [Google Scholar] [CrossRef]
- Fadimu, G.J.; Farahnaky, A.; Gill, H.; Truong, T. Influence of ultrasonic pretreatment on structural properties and biological activities of lupin protein hydrolysate. Int. J. Food Sci. Technol. 2022, 57, 1729–1738. [Google Scholar] [CrossRef]
- He, S.; Franco, C.; Zhang, W. Functions, applications and production of protein hydrolysates from fish processing co-products (FPCP). Food Res. Int. 2013, 50, 289–297. [Google Scholar] [CrossRef]
- Islam, M.S.; Hongxin, W.; Admassu, H.; Noman, A.; Ma, C.; An Wei, F. Degree of hydrolysis, functional and antioxidant properties of protein hydrolysates from Grass Turtle (Chinemys reevesii) as influenced by enzymatic hydrolysis conditions. Food Sci. Nutr. 2021, 9, 4031–4047. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Wang, F.; Ning, Z.; Yang, B.; Wang, Y. Autolysis of anchovy (Engraulis japonicus) protein: Evaluation of antioxidant properties and nutritional quality of resulting hydrolysates. J. Aquat. Food Prod. Technol. 2015, 24, 417–428. [Google Scholar] [CrossRef]
- Islam, M.; Hongxin, W.; Admassu, H.; Mahdi, A.A.; Chaoyang, M.; Wei, F.A. In vitro Antioxidant, Cytotoxic and Antidiabetic Activities of Protein Hydrolysates Prepared from Chinese Pond Turtle (Chinemys reevesii). Food Technol. Biotechnol. 2021, 59, 360–375. [Google Scholar] [CrossRef]
- Noman, A.; Wang, Y.; Zhang, C.; Yin, L.; Abed, S.M. Antioxidant Activities of Optimized Enzymatic Protein Hydrolysates from Hybrid Sturgeon (Huso dauricus × Acipenser schrenckii) Prepared Using Two Proteases. J. Aquat. Food Prod. Technol. 2022, 31, 909–923. [Google Scholar] [CrossRef]
- Gao, R.; Shen, Y.; Shu, W.; Bai, F.; Jin, W.; Wang, J.; Yuan, L. Optimization of enzymatic conditions of sturgeon muscles and their anti-inflammatory potential. J. Food Qual. 2020, 2020, 9698134. [Google Scholar] [CrossRef]
- Ovissipour, M.; Abedian, A.; Motamedzadegan, A.; Rasco, B.; Safari, R.; Shahiri, H. The effect of enzymatic hydrolysis time and temperature on the properties of protein hydrolysates from Persian sturgeon (Acipenser persicus) viscera. Food Chem. 2009, 115, 238–242. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Rao, G.N.; Rao, D.G.; Jyothirmayi, T. Protein hydrolysates from meriga (Cirrhinus mrigala) egg and evaluation of their functional properties. Food Chem. 2010, 120, 652–657. [Google Scholar] [CrossRef]
- Noman, A.; Qixing, J.; Xu, Y.; Ali, A.H.; Al-Bukhaiti, W.Q.; Abed, S.M.; Xia, W. Influence of degree of hydrolysis on chemical composition, functional properties, and antioxidant activities of chinese sturgeon (Acipenser sinensis) hydrolysates obtained by using alcalase 2.4 L. J. Aquat. Food Prod. Technol. 2019, 28, 583–597. [Google Scholar] [CrossRef]
- García-Moreno, P.J.; Batista, I.; Pires, C.; Bandarra, N.M.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Antioxidant activity of protein hydrolysates obtained from discarded Mediterranean fish species. Food Res. Int. 2014, 65, 469–476. [Google Scholar] [CrossRef]
- Ktari, N.; Jridi, M.; Bkhairia, I.; Sayari, N.; Salah, R.B.; Nasri, M. Functionalities and antioxidant properties of protein hydrolysates from muscle of zebra blenny (Salaria basilisca) obtained with different crude protease extracts. Food Res. Int. 2012, 49, 747–756. [Google Scholar] [CrossRef]
- Rao, Q.; Kamdar, A.K.; Labuza, T.P. Storage stability of food protein hydrolysates—A review. Crit. Rev. Food Sci. Nutr. 2016, 56, 1169–1192. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Jyothirmayi, T.; Bhaskarachary, K.; Vajreswari, A.; Hemalatha, R.; Kumar, B.D. Chemical composition, molecular mass distribution and antioxidant capacity of rohu (Labeo rohita) roe (egg) protein hydrolysates prepared by gastrointestinal proteases. Food Res. Int. 2013, 52, 221–229. [Google Scholar] [CrossRef]
- Slizyte, R.; Rommi, K.; Mozuraityte, R.; Eck, P.; Five, K.; Rustad, T. Bioactivities of fish protein hydrolysates from defatted salmon backbones. Biotechnol. Rep. 2016, 11, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Elavarasan, K.; Shamasundar, B.A. Effect of oven drying and freeze drying on the antioxidant and functional properties of protein hydrolysates derived from freshwater fish (Cirrhinus mrigala) using papain enzyme. J. Food Sci. Technol. 2016, 53, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Prihanto, A.A.; Nurdiani, R.; Sari, L.W. Production and characteristics of sailfin catfish (Pterygoplichthys pardalis) protein hydrolysate. F1000Research 2021, 10, 1089. [Google Scholar] [CrossRef]
- Noman, A.; Ali, A.H.; AL-Bukhaiti, W.Q.; Mahdi, A.A.; Xia, W. Structural and physicochemical characteristics of lyophilized Chinese sturgeon protein hydrolysates prepared by using two different enzymes. J. Food Sci. 2020, 85, 3313–3322. [Google Scholar] [CrossRef]
- Kristoffersen, K.A.; Liland, K.H.; Böcker, U.; Wubshet, S.G.; Lindberg, D.; Horn, S.J.; Afseth, N.K. FTIR-based hierarchical modeling for prediction of average molecular weights of protein hydrolysates. Talanta 2019, 205, 120084. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Jiang, L.; Qi, B.; Zhou, L. Relationship between secondary structure and surface hydrophobicity of soybean protein isolate subjected to heat treatment. J. Chem. 2014, 2014, 475389. [Google Scholar] [CrossRef]
- Wubshet, S.G.; Måge, I.; Böcker, U.; Lindberg, D.; Knutsen, S.H.; Rieder, A.; Rodriguez, D.A.; Afseth, N.K. FTIR as a rapid tool for monitoring molecular weight distribution during enzymatic protein hydrolysis of food processing by-products. Anal. Methods 2017, 9, 4247–4254. [Google Scholar] [CrossRef]
- Yang, H.; Yang, S.; Kong, J.; Dong, A.; Yu, S. Obtaining information about protein secondary structures in aqueous solution using Fourier transform IR spectroscopy. Nat. Protoc. 2015, 10, 382–396. [Google Scholar] [CrossRef]
- Cai, S.; Singh, B.R. A distinct utility of the amide III infrared band for secondary structure estimation of aqueous protein solutions using partial least squares methods. Biochemistry 2004, 43, 2541–2549. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [PubMed]
- Thiansilakul, Y.; Benjakul, S.; Shahidi, F. Compositions, functional properties and antioxidative activity of protein hydrolysates prepared from round scad (Decapterus maruadsi). Food Chem. 2007, 103, 1385–1394. [Google Scholar] [CrossRef]
- Gbogouri, G.; Linder, M.; Fanni, J.; Parmentier, M. Influence of hydrolysis degree on the functional properties of salmon byproducts hydrolysates. J. Food Sci. 2004, 69, C615–C622. [Google Scholar] [CrossRef]
- Foh, M.B.K.; Amadou, I.; Foh, B.M.; Kamara, M.T.; Xia, W. Functionality and antioxidant properties of tilapia (Oreochromis niloticus) as influenced by the degree of hydrolysis. Int. J. Mol. Sci. 2010, 11, 1851–1869. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Luo, Y.; Shen, H.; You, J. Antioxidant activities and functional properties of grass carp (Ctenopharyngodon idellus) protein hydrolysates. J. Sci. Food Agric. 2012, 92, 292–298. [Google Scholar] [CrossRef]
- Nalinanon, S.; Benjakul, S.; Kishimura, H.; Shahidi, F. Functionalities and antioxidant properties of protein hydrolysates from the muscle of ornate threadfin bream treated with pepsin from skipjack tuna. Food Chem. 2011, 124, 1354–1362. [Google Scholar] [CrossRef]
- Zhao, J.; Xiong, Y.L.; McNear, D.H. Changes in structural characteristics of antioxidative soy protein hydrolysates resulting from scavenging of hydroxyl radicals. J. Food Sci. 2013, 78, C152–C159. [Google Scholar] [CrossRef]
- Wali, A.; Ma, H.; Shahnawaz, M.; Hayat, K.; Xiaong, J.; Jing, L. Impact of power ultrasound on antihypertensive activity, functional properties, and thermal stability of rapeseed protein hydrolysates. J. Chem. 2017, 2017, 4373859. [Google Scholar] [CrossRef]
- Gromiha, M.M. Protein Bioinformatics: From Sequence to Function; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Chalamaiah, M.; Jyothirmayi, T.; Diwan, P.V.; Dinesh Kumar, B. Antioxidant activity and functional properties of enzymatic protein hydrolysates from common carp (Cyprinus carpio) roe (egg). J. Food Sci. Technol. 2015, 52, 5817–5825. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Wang, F.; Yuan, L.; Liu, J.; Sun, D.; Li, X. Physicochemical property, antioxidant activity, and cytoprotective effect of the germinated soybean proteins. Food Sci. Nutr. 2019, 7, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.S.; Andersen, L.L.; Nielsen, H.H.; Jacobsen, C.; Jakobsen, G.; Johansson, I.; Jessen, F. Antioxidant activity of Cod (Gadus morhua) protein hydrolysates: In vitro assays and evaluation in 5% fish oil-in-water emulsion. Food Chem. 2014, 149, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, G. The protein digestibility-corrected amino acid score (PDCAAS)—A concept for describing protein quality in foods and food ingredients: A critical review. J. AOAC Int. 2005, 88, 988–994. [Google Scholar] [CrossRef]
- Romadhoni, A.R.; Afrianto, E.; Pratama, R.I.; Grandiosa, R. Extraction of snakehead fish [Ophiocephalus striatus (Bloch, 1793)] into fish protein concentrate as albumin source using various solvent. Aquat. Procedia 2016, 7, 4–11. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemist; AOAC International: Gaithersburg, MD, USA, 1999. [Google Scholar]
- Liu, W.; Gu, R.; Lin, F.; Lu, J.; Yi, W.; Ma, Y.; Dong, Z.; Cai, M. Isolation and identification of antioxidative peptides from pilot-scale black-bone silky fowl (Gallus gallus domesticus Brisson) muscle oligopeptides. J. Sci. Food Agric. 2013, 93, 2782–2788. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Jain, S.; Anal, A.K. Optimization of extraction of functional protein hydrolysates from chicken egg shell membrane (ESM) by ultrasonic assisted extraction (UAE) and enzymatic hydrolysis. LWT-Food Sci. Technol. 2016, 69, 295–302. [Google Scholar] [CrossRef]
- Jemil, I.; Jridi, M.; Nasri, R.; Ktari, N.; Salem, R.B.S.B.; Mehiri, M.; Hajji, M.; Nasri, M. Functional, antioxidant and antibacterial properties of protein hydrolysates prepared from fish meat fermented by Bacillus subtilis A26. Process Biochem. 2014, 49, 963–972. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Dang, X.; Zheng, X.; Zhang, W. Housefly larvae hydrolysate: Orthogonal optimization of hydrolysis, antioxidant activity, amino acid composition and functional properties. BMC Res. Notes 2013, 6, 197. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Degree of Hydrolysis | |||
|---|---|---|---|
| Parameters | 16.56% | 22.23% | 25.48% |
| Hydrolysate yield | 13.08 ± 0.58 c | 17.10 ± 0.36 b | 17.83 ± 0.67 a |
| Moisture | 3.72 ± 0.09 a | 3.64 ± 0.05 a | 3.77 ± 0.09 a |
| Protein | 83.61 ± 1.25 c | 85.94 ± 0.77 b | 88.19 ± 2.02 a |
| Fat | 2.98 ± 0.09 a | 3.34 ± 0.12 a | 2.04 ± 0.05 b |
| Ash | 6.21 ± 0.12 a | 5.92 ± 0.10 a | 4.85 ± 0.23 b |
| Color measurement | |||
| L* | 86.89 ± 0.52 a | 84.06 ± 0.24 b | 83.98 ± 0.16 b |
| a* | 1.39 ± 0.08 b | 1.43 ± 0.05 b | 1.92 ± 0.11 a |
| b* | 15.81 ± 0.37 b | 17.42 ± 0.18 a | 17.97 ± 0.21 a |
| ∆E | 18.11 ± 0.09 a | 17.28 ± 0.05 b | 18.51 ± 0.13 a |
| Water activity | 0.36 ± 0.00 a | 0.34 ± 0.00 a | 0.27 ± 0.01 b |
| WHC (g/g FPH) | 1.97 ± 0.09 c | 2.64 ± 0.17 b | 3.49 ± 0.25 a |
| OHC (g/g FPH) | 2.85 ± 0.07 a | 2.31 ± 0.15 b | 2.19 ± 0.11 b |
| CD Values | Hydrolysates | ||
|---|---|---|---|
| 16.56% | 22.23% | 25.48% | |
| α-Helix (%) | 5.3 ± 0.18 a | 4.1 ± 0.11 b | 4.15 ± 0.25 b |
| β-Sheets (%) | 40.1 ± 0.64 b | 42.5 ± 1.82 a | 42.4 ± 1.12 a |
| β-Turn (%) | 20.2 ± 0.48 b | 21.4 ± 1.09 a | 21.3 ± 0.51 a |
| Unordered (%) | 34.4 ± 1.30 a | 32.1 ± 1.44 b | 32.1 ± 0.60 b |
| Amino Acids | Hydrolysates | ||
|---|---|---|---|
| Essential amino acids (EAAs) | DH 16.56% (1 h) | DH 22.23% (3 h) | DH 25.48% (6 h) |
| Histidine | 2.87 ± 0.08 b | 2.89 ± 0.07 b | 3.12 ± 0.02 a |
| Threonine | 4.10 ± 0.04 b | 4.90 ± 0.04 a | 4.88 ± 0.02 a |
| Valine | 4.67 ± 0.06 b | 4.68 ± 0.03 b | 5.13 ± 0.05 a |
| Methionine | 2.83 ± 0.04 b | 2.90 ± 0.06 b | 3.11 ± 0.08 a |
| Phenylalanine | 3.63 ± 0.04 c | 3.78 ± 0.02 b | 4.12 ± 0.03 a |
| Isoleucine | 4.46 ± 0.02 b | 4.51 ± 0.01 b | 4.90 ± 0.09 a |
| Leucine | 6.59 ± 0.02 b | 6.66 ± 0.04 b | 6.84 ± 0.05 a |
| Lysine | 7.11 ± 0.03 b | 7.20 ± 0.08 b | 7.88 ± 0.04 a |
| Arginine | 6.38 ± 0.02 b | 6.41 ± 0.01 b | 6.91 ± 0.02 a |
| Tyrosine | 2.67 ± 0.03 b | 2.92 ± 0.05 a | 2.94 ± 0.09 a |
| Non-essential amino acids (NAAs) | |||
| Cysteine | 0.21 ± 0.03 a | 0.22 ± 0.02 a | 0.26 ± 0.02 a |
| Aspartic acid | 9.11 ± 0.02 b | 9.20 ± 0.07 b | 9.44 ± 0.09 a |
| Glutamic acid | 12.47 ± 0.04 a | 12.45 ± 0.02 a | 12.29 ± 0.07 b |
| Serine | 2.37 ± 0.03 a | 2.38 ± 0.08 a | 2.32 ± 0.05 a |
| Glycine | 3.93 ± 0.05 b | 4.20 ± 0.01 a | 3.89 ± 0.04 b |
| Proline | 2.41 ± 0.03 b | 2.67 ± 0.06 a | 2.61 ± 0.07 a |
| Alanine | 5.30 ± 0.02 b | 5.34 ± 0.01 a | 5.26 ± 0.02 b |
| TEAA | 45.34 ± 0.58 c | 46.93 ± 0.28 b | 49.81 ± 0.19 a |
| TNAA | 35.81 ± 0.16 b | 36.11 ± 0.21 ab | 36.22 ± 0.29 a |
| TAA | 81.15 ± 0.41 c | 83.05 ± 0.54 b | 86.03 ± 0.34 a |
| TEAA/TAA% | 55.87 ± 0.33 | 56.50 ± 0.45 | 57.91 ± 0.52 |
| Parameters | Levels |
|---|---|
| pH | 5.5, 6, 6.5, and 7 |
| Temperature (°C) | 30, 40, 50, 60, and 70 |
| E/S ratio (%) | 1, 2, 3, 4, and 5 |
| Hydrolysis time (h) | 1, 2, 3, 4, 5, 6, and 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alahmad, K.; Noman, A.; Xia, W.; Jiang, Q.; Xu, Y. Influence of the Enzymatic Hydrolysis Using Flavourzyme Enzyme on Functional, Secondary Structure, and Antioxidant Characteristics of Protein Hydrolysates Produced from Bighead Carp (Hypophthalmichthys nobilis). Molecules 2023, 28, 519. https://doi.org/10.3390/molecules28020519
Alahmad K, Noman A, Xia W, Jiang Q, Xu Y. Influence of the Enzymatic Hydrolysis Using Flavourzyme Enzyme on Functional, Secondary Structure, and Antioxidant Characteristics of Protein Hydrolysates Produced from Bighead Carp (Hypophthalmichthys nobilis). Molecules. 2023; 28(2):519. https://doi.org/10.3390/molecules28020519
Chicago/Turabian StyleAlahmad, Kamal, Anwar Noman, Wenshui Xia, Qixing Jiang, and Yanshun Xu. 2023. "Influence of the Enzymatic Hydrolysis Using Flavourzyme Enzyme on Functional, Secondary Structure, and Antioxidant Characteristics of Protein Hydrolysates Produced from Bighead Carp (Hypophthalmichthys nobilis)" Molecules 28, no. 2: 519. https://doi.org/10.3390/molecules28020519
APA StyleAlahmad, K., Noman, A., Xia, W., Jiang, Q., & Xu, Y. (2023). Influence of the Enzymatic Hydrolysis Using Flavourzyme Enzyme on Functional, Secondary Structure, and Antioxidant Characteristics of Protein Hydrolysates Produced from Bighead Carp (Hypophthalmichthys nobilis). Molecules, 28(2), 519. https://doi.org/10.3390/molecules28020519