
Multi-Target Effect of Aloeresin-A against Bacterial and Host Inflammatory Targets Benefits Contact Lens-Related Keratitis: A Multi-Omics and Quantum Chemical Investigation


Abstract
:1. Introduction
2. Results and Discussion
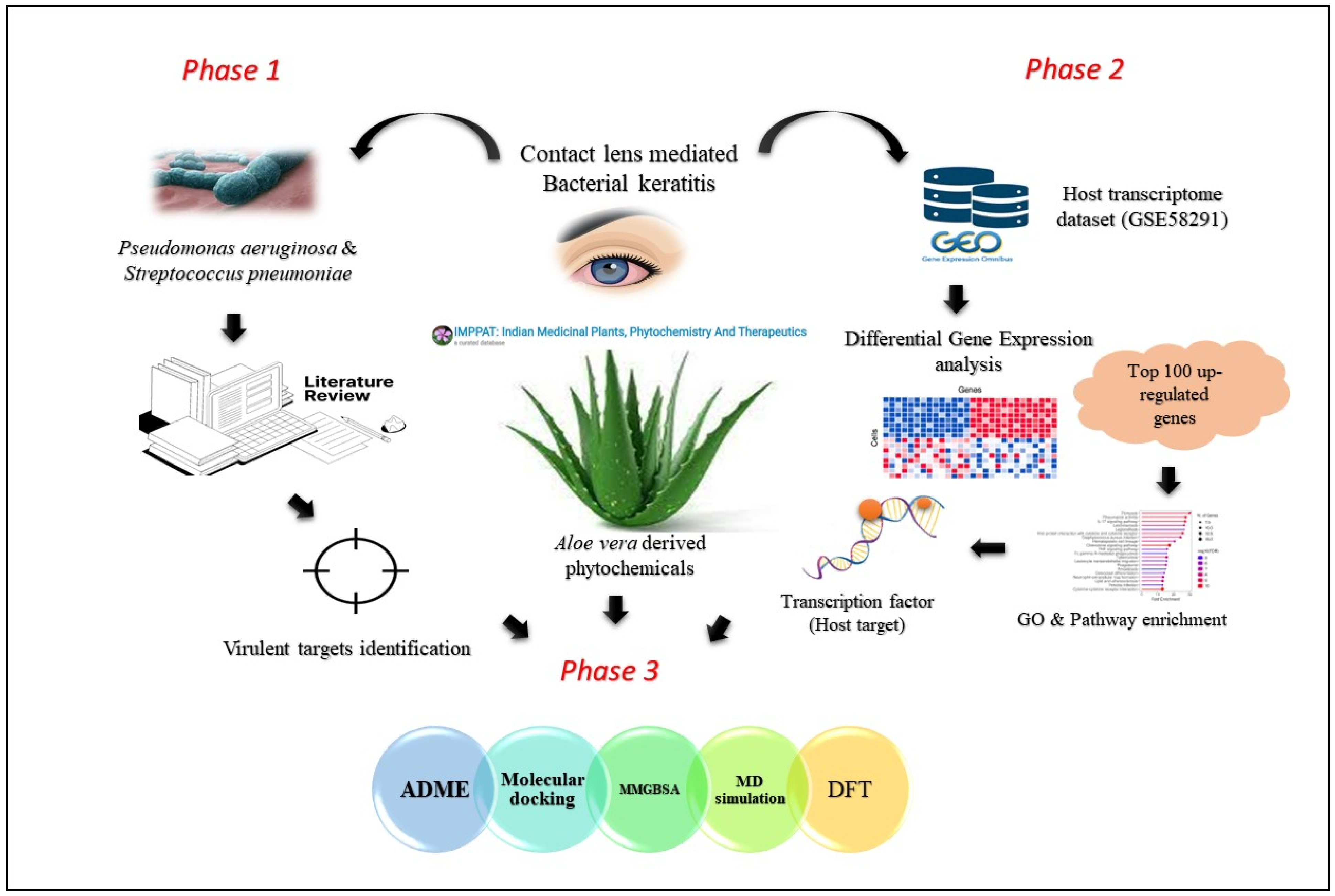
2.1. Literature Analysis and Bacterial Target Identification
2.2. ADME Profiling and Ligand Collection
2.3. Regulatory Network Construction and Host Target Discovery
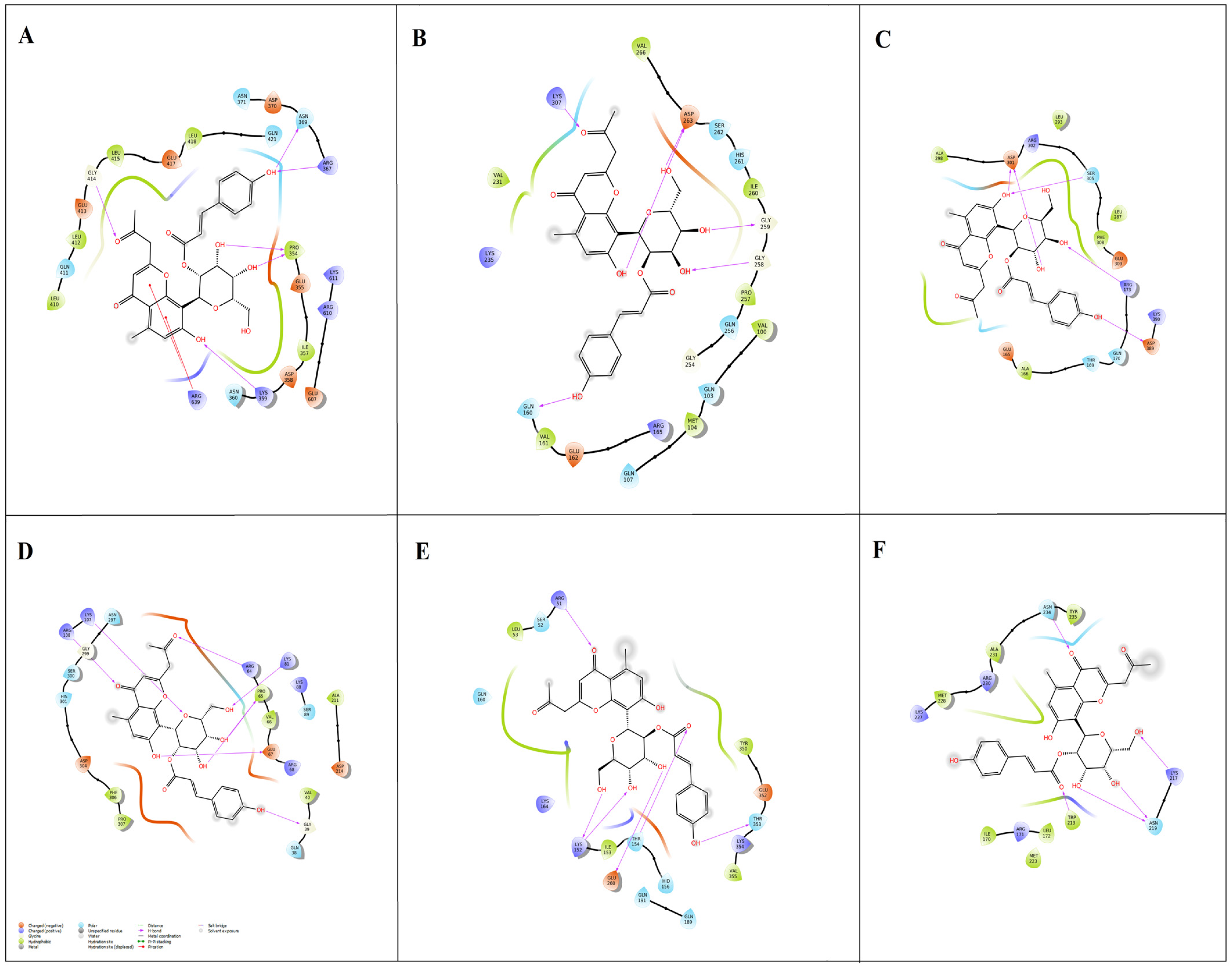
2.4. Inhibitory Potential of A. vera Phytochemicals against Keratitis Targets
2.5. Binding Free Energy Calculations
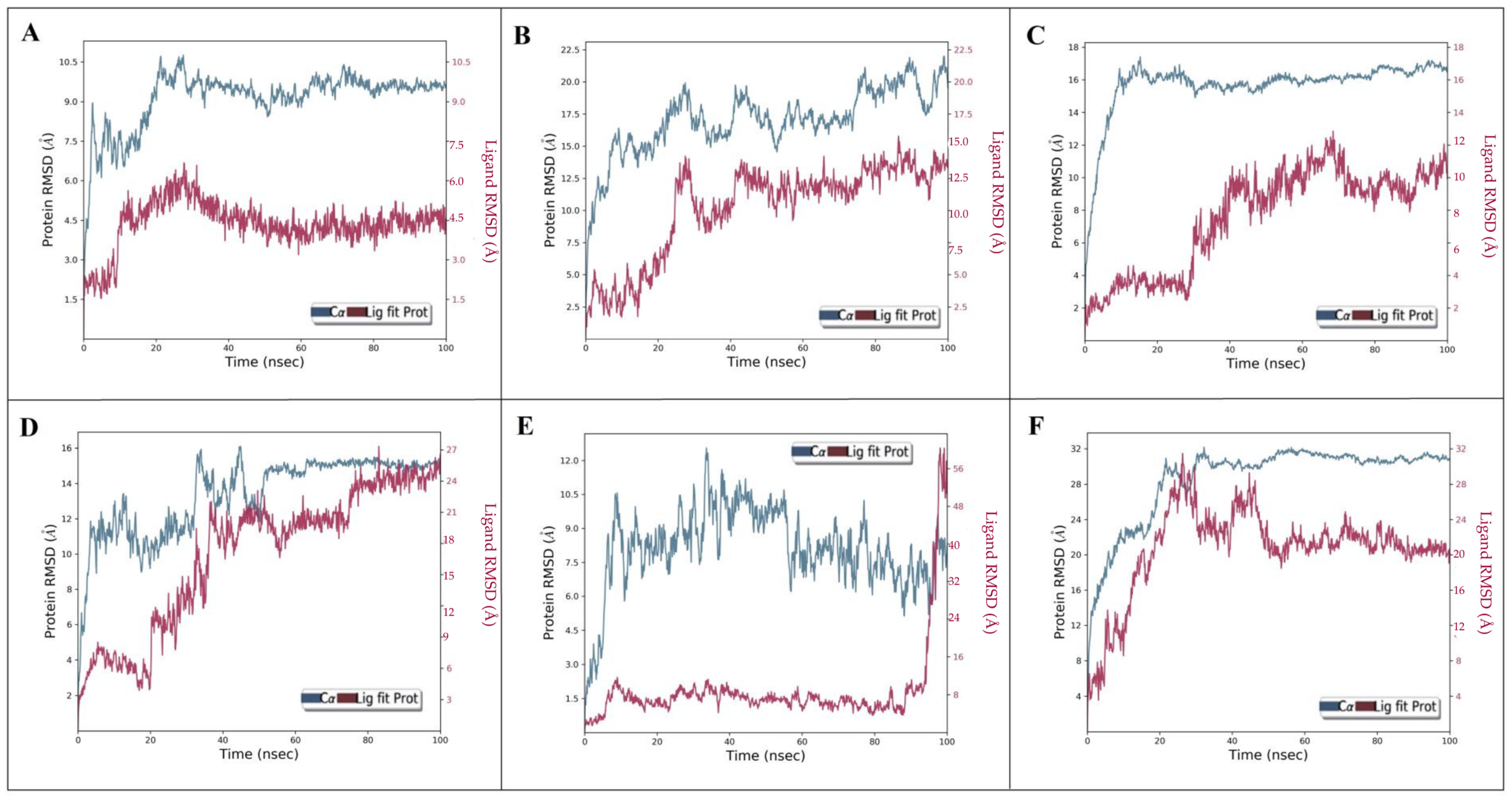
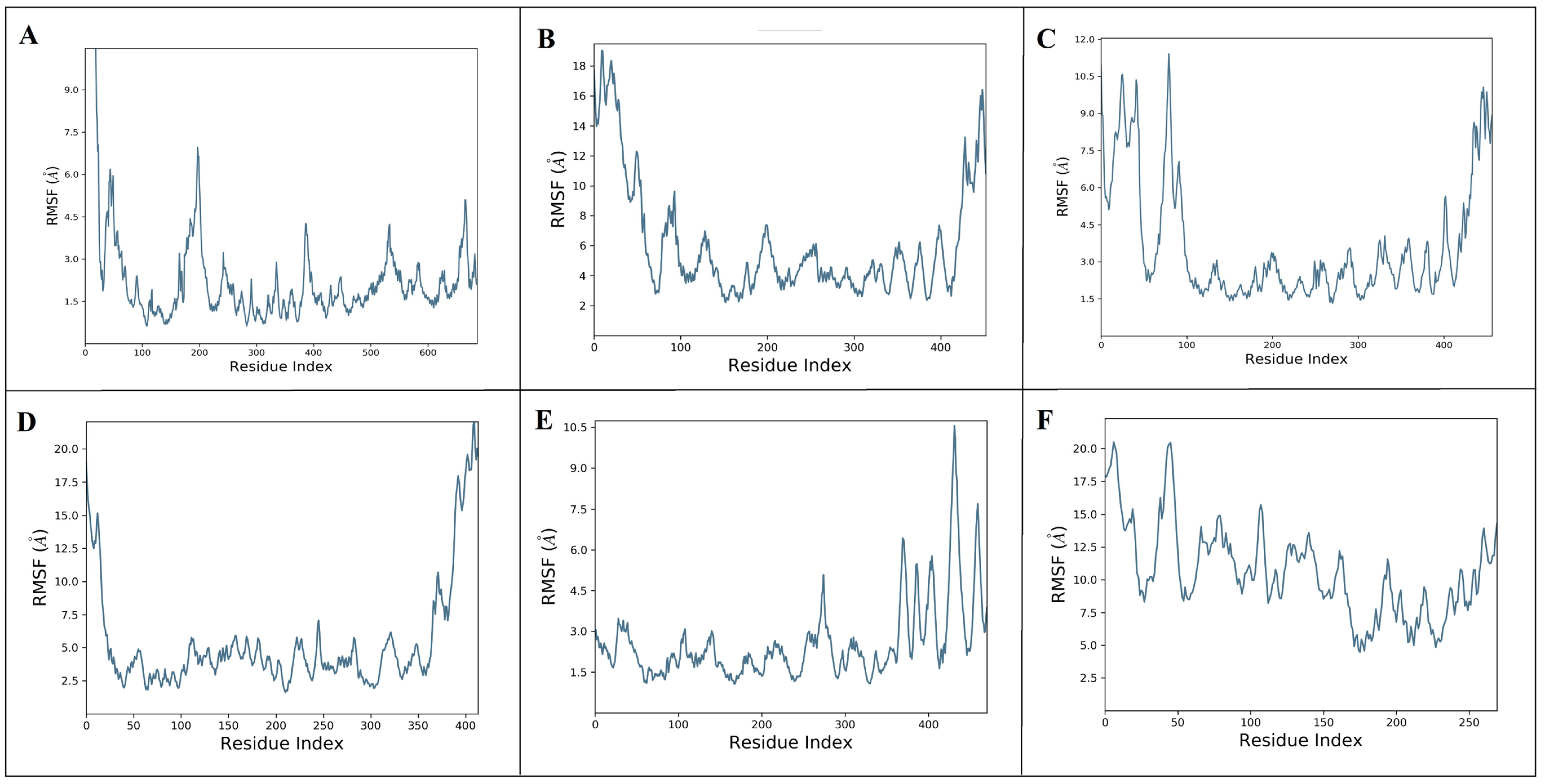
2.6. Dynamic Simulation of Aloeresin-A against Bacterial and Host Targets
2.7. Density Functional Theory Analysis
2.7.1. HOMO and LUMO Analysis
2.7.2. Quantum Chemical Descriptors
2.7.3. Molecular Electrostatic Potential (MEP)
2.7.4. Mulliken Charge Analysis
3. Materials and Methods
3.1. Phytochemicals Collection and ADME Evaluation
3.2. Bacterial Target Search from the Literature
3.3. Host Target Identification through Regulatory Network Analysis
3.4. Molecular Docking with Bacterial and Host Targets
3.5. MMGBSA Analysis
3.6. Molecular Dynamic Simulation
3.7. DFT Calculations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Moreddu, R.; Vigolo, D.; Yetisen, A.K. Contact Lens Technology: From Fundamentals to Applications. Adv. Healthc. Mater. 2019, 8, 15. [Google Scholar] [CrossRef]
- Rhee, M.K.; Jacobs, D.S.; Dhaliwal, D.K.; Szczotka-Flynn, L.; Prescott, C.R.; Jhanji, V.; Steinemann, T.L.; Koffler, B.H.; Jeng, B.H. Contact Lens Safety for the Correction of Refractive Error in Healthy Eyes. Eye Contact Lens 2022, 48, 449–454. [Google Scholar] [CrossRef]
- Alamillo-Velazquez, J.; Ruiz-Lozano, R.E.; Hernandez-Camarena, J.C.; Rodriguez-Garcia, A. Contact lens-associated infectious keratitis: Update on diagnosis and therapy. In Infectious Eye Diseases-Recent Advances in Diagnosis and Treatment; IntechOpen: London, UK, 2021. [Google Scholar]
- Eltis, M. Contact-lens-related microbial keratitis: Case report and review. J. Optom. 2011, 4, 122–127. [Google Scholar] [CrossRef]
- Gurnani, B.; Kaur, K. Bacterial Keratitis. In StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2023. [Google Scholar]
- Al-Mujaini, A.; Al-Kharusi, N.; Thakral, A.; Wali, U.K. Bacterial keratitis: Perspective on epidemiology, clinico-pathogenesis, diagnosis and treatment. Sultan Qaboos Univ. Med. J. 2009, 9, 184–195. [Google Scholar] [PubMed]
- Suzuki, T.; Inoue, H. Mechanisms Underlying Contact Lens-Related Keratitis Caused by Pseudomonas aeruginosa. Eye Contact Lens 2022, 48, 134–137. [Google Scholar] [CrossRef]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Norcross, E.W.; Sanders, M.E.; Moore, Q.C., 3rd; Marquart, M.E. Pathogenesis of A Clinical Ocular Strain of Streptococcus pneumoniae and the Interaction of Pneumolysin with Corneal Cells. J. Bacteriol. Parasitol. 2011, 2, 108. [Google Scholar] [CrossRef] [PubMed]
- Bouhenni, R.; Dunmire, J.; Rowe, T.; Bates, J. Proteomics in the Study of Bacterial Keratitis. Proteomes 2015, 3, 496–511. [Google Scholar] [CrossRef]
- Yoo, I.H.; Shin, H.S.; Kim, Y.J.; Kim, H.B.; Jin, S.; Ha, U.H. Role of pneumococcal pneumolysin in the induction of an inflammatory response in human epithelial cells. FEMS Immunol. Med. Microbiol. 2010, 60, 28–35. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, F.B.M.; Morais, V.C.; de Oliveira, B.T.M.; de Lima, K.Y.G.; Gomes, V.T.; do Amaral, I.P.G.; Vasconcelos, U. Multi-purpose Disinfecting Solutions only Partially Inhibit the Development of Ocular Microbes Biofilms in Contact Lens Storage Cases. Middle East Afr. J. Ophthalmol. 2021, 28, 116–122. [Google Scholar] [PubMed]
- Pinto-Fraga, J.; Blázquez Arauzo, F.; Urbano Rodríguez, R.; González-García, M.J. Evaluation of safety and efficacy of a new multipurpose disinfecting solution on silicone hydrogel contact lenses. J. Optom. 2015, 8, 40–47. [Google Scholar] [CrossRef]
- Rhone, M.; Basu, A. Phytochemicals and age-related eye diseases. Nutr. Rev. 2008, 66, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, M.; Fazeli, M.; Azad, M.; Seyedjavadi, S.S.; Mousavi, R. Aloe vera Gel: Effective Therapeutic Agent against Multidrug-Resistant Pseudomonas aeruginosa Isolates Recovered from Burn Wound Infections. Chemother. Res. Pract. 2015, 2015, 639806. [Google Scholar] [CrossRef] [PubMed]
- Naini, M.A.; Zargari-Samadnejad, A.; Mehrvarz, S.; Tanideh, R.; Ghorbani, M.; Dehghanian, A.; Hasanzarrini, M.; Banaee, F.; Koohi-Hosseinabadi, O.; Tanideh, N.; et al. Anti-Inflammatory, Antioxidant, and Healing-Promoting Effects of Aloe vera Extract in the Experimental Colitis in Rats. Evid.-Based Complement. Altern. Med. Ecam 2021, 2021, 9945244. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, S.; Sivagnanam, K.; Subramanian, S. Modulatory effects of Aloe vera leaf gel extract on oxidative stress in rats treated with streptozotocin. J. Pharm. Pharmacol. 2005, 57, 241–246. [Google Scholar] [CrossRef]
- Jain, S.; Rathod, N.; Nagi, R.; Sur, J.; Laheji, A.; Gupta, N.; Agrawal, P.; Prasad, S. Antibacterial Effect of Aloe Vera Gel against Oral Pathogens: An In-vitro Study. J. Clin. Diagn. Res. JCDR 2016, 10, ZC41–ZC44. [Google Scholar] [CrossRef] [PubMed]
- Saks, Y.; Barkai-Golan, R. Aloe vera gel activity against plant pathogenic fungi. Postharvest Biol. Technol. 1995, 6, 159–165. [Google Scholar] [CrossRef]
- Ceravolo, I.; Mannino, F.; Irrera, N.; Squadrito, F.; Altavilla, D.; Ceravolo, G.; Pallio, G.; Minutoli, L. Health Potential of Aloe vera against Oxidative Stress Induced Corneal Damage: An “In Vitro” Study. Antioxidants 2021, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; González-Burgos, E.; Iglesias, I.; Gómez-Serranillos, M.P. Pharmacological Update Properties of Aloe Vera and its Major Active Constituents. Molecules 2020, 25, 1324. [Google Scholar] [CrossRef] [PubMed]
- Fleiszig, S.M.; Evans, D.J. Pathogenesis of contact lens-associated microbial keratitis. Optom. Vis. Sci. Off. Publ. Am. Acad. Optom. 2010, 87, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, R.S.; Priya, J.L.; Leal, S.M., Jr.; Toska, J.; Rietsch, A.; Prajna, V.; Pearlman, E.; Lalitha, P. Host response and bacterial virulence factor expression in Pseudomonas aeruginosa and Streptococcus pneumoniae corneal ulcers. PLoS ONE 2013, 8, e64867. [Google Scholar] [CrossRef]
- Mohanraj, K.; Karthikeyan, B.S.; Vivek-Ananth, R.P.; Chand, R.P.B.; Aparna, S.R.; Mangalapandi, P.; Samal, A. IMPPAT: A curated database of Indian Medicinal Plants, Phytochemistry and Therapeutics. Sci. Rep. 2018, 8, 4329. [Google Scholar] [CrossRef]
- Kulanthaivel, L.; Jeyaraman, J.; Biswas, A.; Subbaraj, G.K.; Santhoshkumar, S. Identification of potential inhibitors for Penicillinbinding protein (PBP) from Staphylococcus aureus. Bioinformation 2018, 14, 471–476. [Google Scholar] [CrossRef]
- Arbab, S.; Ullah, H.; Weiwei, W.; Wei, X.; Ahmad, S.U.; Wu, L.; Zhang, J. Comparative study of antimicrobial action of aloe vera and antibiotics against different bacterial isolates from skin infection. Vet. Med. Sci. 2021, 7, 2061–2067. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Janky, R.; Verfaillie, A.; Imrichová, H.; Van de Sande, B.; Standaert, L.; Christiaens, V.; Hulselmans, G.; Herten, K.; Naval Sanchez, M.; Potier, D.; et al. iRegulon: From a gene list to a gene regulatory network using large motif and track collections. PLoS Comput. Biol. 2014, 10, e1003731. [Google Scholar] [CrossRef]
- Han, R.; Yuan, T.; Yang, Z.; Zhang, Q.; Wang, W.W.; Lin, L.B.; Zhu, M.Q.; Gao, J.M. Ulmoidol, an unusual nortriterpenoid from Eucommia ulmoides Oliv. Leaves prevents neuroinflammation by targeting the PU.1 transcriptional signaling pathway. Bioorganic Chem. 2021, 116, 105345. [Google Scholar] [CrossRef]
- Xia, W.; Luo, P.; Hua, P.; Ding, P.; Li, C.; Xu, J.; Zhou, H.; Gu, Q. Discovery of a New Pterocarpan-Type Antineuroinflammatory Compound from Sophora tonkinensis through Suppression of the TLR4/NFκB/MAPK Signaling Pathway with PU.1 as a Potential Target. ACS Chem. Neurosci. 2019, 10, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Breaud, C.; Lallemand, L.; Mares, G.; Mabrouki, F.; Bertolotti, M.; Simmler, C.; Greff, S.; Mauduit, M.; Herbette, G.; Garayev, E.; et al. LC-MS Based Phytochemical Profiling towards the Identification of Antioxidant Markers in Some Endemic Aloe Species from Mascarene Islands. Antioxidants 2022, 12, 50. [Google Scholar] [CrossRef] [PubMed]
- Elekofehinti, O.O.; Iwaloye, O.; Josiah, S.S.; Lawal, A.O.; Akinjiyan, M.O.; Ariyo, E.O. Molecular docking studies, molecular dynamics and ADME/tox reveal therapeutic potentials of STOCK1N-69160 against papain-like protease of SARS-CoV-2. Mol. Divers. 2021, 25, 1761–1773. [Google Scholar] [CrossRef]
- Alghamdi, A.; Abouzied, A.S.; Alamri, A.; Anwar, S.; Ansari, M.; Khadra, I.; Zaki, Y.H.; Gomha, S.M. Synthesis, Molecular Docking, and Dynamic Simulation Targeting Main Protease (Mpro) of New, Thiazole Clubbed Pyridine Scaffolds as Potential COVID-19 Inhibitors. Curr. Issues Mol. Biol. 2023, 45, 1422–1442. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Su, N.Q.; Yang, W. Describing Chemical Reactivity with Frontier Molecular Orbitalets. JACS Au 2022, 2, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Abu-Melha, S. Pyridyl thiosemicarbazide: Synthesis, crystal structure, DFT/B3LYP, molecular docking studies and its biological investigations. Chem. Cent. J. 2018, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Pantsar, T.; Poso, A. Binding Affinity via Docking: Fact and Fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef]
- Deghady, A.M.; Hussein, R.K.; Alhamzani, A.G.; Mera, A. Density Functional Theory and Molecular Docking Investigations of the Chemical and Antibacterial Activities for 1-(4-Hydroxyphenyl)-3-phenylprop-2-en-1-one. Molecules 2021, 26, 3631. [Google Scholar] [CrossRef]
- El-Shamy, N.T.; Alkaoud, A.M.; Hussein, R.K.; Ibrahim, M.A.; Alhamzani, A.G.; Abou-Krisha, M.M. DFT, ADMET and Molecular Docking Investigations for the Antimicrobial Activity of 6, 6′-Diamino-1, 1′, 3, 3′-tetramethyl-5, 5′-(4-chlorobenzylidene) bis [pyrimidine-2, 4 (1H, 3H)-dione]. Molecules 2022, 27, 620. [Google Scholar] [CrossRef] [PubMed]
- Sasitha, T.; John, W.J. Design, docking, and DFT investigations of 2,6-bis(3,4-dihydroxyphenyl)-3-phenethylpiperidin-4-one. Heliyon 2021, 7, e06127. [Google Scholar] [CrossRef] [PubMed]
- Dlala, N.A.; Bouazizi, Y.; Ghalla, H.; Hamdi, N. DFT calculations and molecular docking studies on a chromene derivative. J. Chem. 2021, 2021, 6674261. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Hatami, S.; Sirous, H.; Mahnam, K.; Najafipour, A.; Fassihi, A. Preparing a database of corrected protein structures important in cell signaling pathways. Res. Pharm. Sci. 2022, 18, 67–77. [Google Scholar] [PubMed]
- Pandi, S.; Kulanthaivel, L.; Subbaraj, G.K.; Rajaram, S.; Subramanian, S. Screening of Potential Breast Cancer Inhibitors through Molecular Docking and Molecular Dynamics Simulation. BioMed Res. Int. 2022, 2022, 3338549. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SI.NO | Target | Phytochemicals | Docking Score (kcal/mol) |
|---|---|---|---|
| 1 | ExoU | Aloenin | −7.74 |
| Aloeresin-A | −7.59 | ||
| Isoaloesin | −7.38 | ||
| Aloin | −7.16 | ||
| Barbaloin | −7.16 | ||
| 2 | ExoS | Aloin | −7.22 |
| Barbaloin | −7.22 | ||
| 10-Hydroxyaloin B 6′-catate | −7.14 | ||
| Aloeresin-A | −7.09 | ||
| 10-Hydroxyaloin A | −7.01 | ||
| 3 | ExoT | Aloeresin-A | −6.56 |
| Isoaloesin | −6.35 | ||
| Aloenin | −6.07 | ||
| 10-Hydroxyaloin A | −6.04 | ||
| 7-Hydroxyaloin | −5.97 | ||
| 4 | ExoY | Aloeresin-A | −6.42 |
| Aloenin | −6.20 | ||
| Aloin | −5.98 | ||
| Barbaloin | −5.98 | ||
| 10-Hydroxyaloin B 6′-catate | −5.91 | ||
| 5 | PLY | Allantoin | −6.89 |
| Aloeresin-A | −6.20 | ||
| Aloesin | −6.15 | ||
| Anthracene-1,8-diol | −6.12 | ||
| Isoaloesin | −5.90 | ||
| 6 | SPI1 | Aloeresin-A | −5.62 |
| SI. NO | Target Protein | ΔGbind (kcal/mol) |
|---|---|---|
| 1 | ExoU | −57.36 |
| 2 | ExoS | −54.89 |
| 3 | ExoT | −56.98 |
| 4 | ExoY | −50.72 |
| 5 | PLY | −59.18 |
| 6 | SPI1 | −49.21 |
| SI. No | Descriptors | Definition | Value (eV) |
|---|---|---|---|
| 1 | EHOMO | - | −6.07 |
| 2 | ELUMO | - | −1.70 |
| 3 | Energy gap (ΔE) | ∆E = ELUMO − EHOMO | 4.37 |
| 4 | Electron affinity (A) | −ELUMO | 1.70 |
| 5 | Ionization potential (I) | −EHOMO | 6.07 |
| 6 | Electron accepting capacity (ω+) | (I + 3A)2/16(I − A) | 1.78 |
| 7 | Electron donating capacity (ω−) | (3I + A)2/16(I − A) | 5.66 |
| 8 | Chemical hardness (η) | (I-A)/2 | 2.18 |
| 9 | Global softness (S) | 1/2η | 0.22 |
| 10 | Chemical softness (σ) | 1/η | 0.45 |
| 11 | Electrophilicity index (ω) | μ2/2η | 3.45 |
| 12 | Nucleophilicity index (N) | EHOMO-EHOMO(TCE) | 3.11 |
| 13 | Electronegativity (χ) | (I + A)/2 | 3.88 |
| 14 | Electronic chemical potential (μ) | −(I + A)/2 | −3.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roshni, J.; Ahmad, S.F.; Wani, A.; Ahmed, S.S.S.J. Multi-Target Effect of Aloeresin-A against Bacterial and Host Inflammatory Targets Benefits Contact Lens-Related Keratitis: A Multi-Omics and Quantum Chemical Investigation. Molecules 2023, 28, 6955. https://doi.org/10.3390/molecules28196955
Roshni J, Ahmad SF, Wani A, Ahmed SSSJ. Multi-Target Effect of Aloeresin-A against Bacterial and Host Inflammatory Targets Benefits Contact Lens-Related Keratitis: A Multi-Omics and Quantum Chemical Investigation. Molecules. 2023; 28(19):6955. https://doi.org/10.3390/molecules28196955
Chicago/Turabian StyleRoshni, Jency, Sheikh F. Ahmad, Abubakar Wani, and Shiek S. S. J. Ahmed. 2023. "Multi-Target Effect of Aloeresin-A against Bacterial and Host Inflammatory Targets Benefits Contact Lens-Related Keratitis: A Multi-Omics and Quantum Chemical Investigation" Molecules 28, no. 19: 6955. https://doi.org/10.3390/molecules28196955
APA StyleRoshni, J., Ahmad, S. F., Wani, A., & Ahmed, S. S. S. J. (2023). Multi-Target Effect of Aloeresin-A against Bacterial and Host Inflammatory Targets Benefits Contact Lens-Related Keratitis: A Multi-Omics and Quantum Chemical Investigation. Molecules, 28(19), 6955. https://doi.org/10.3390/molecules28196955