

A Potential Anti-Glioblastoma Compound LH20 Induces Apoptosis and Arrest of Human Glioblastoma Cells via CDK4/6 Inhibition
 ,
,  and
and
Abstract
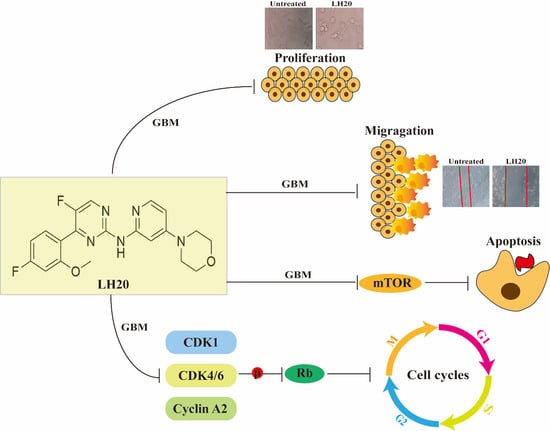
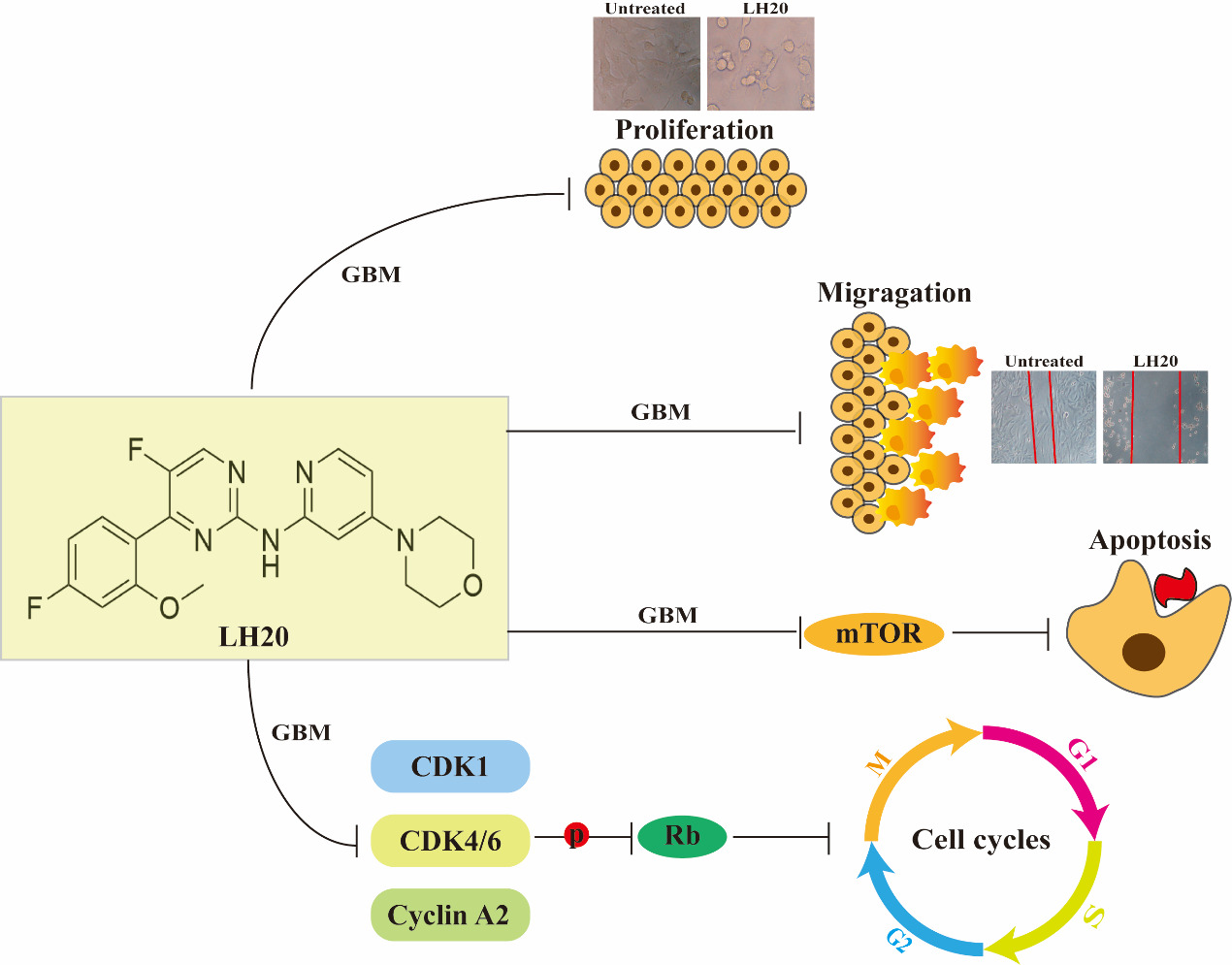
1. Introduction
2. Results
2.1. LH20 Inhibits GBM Cell Proliferation
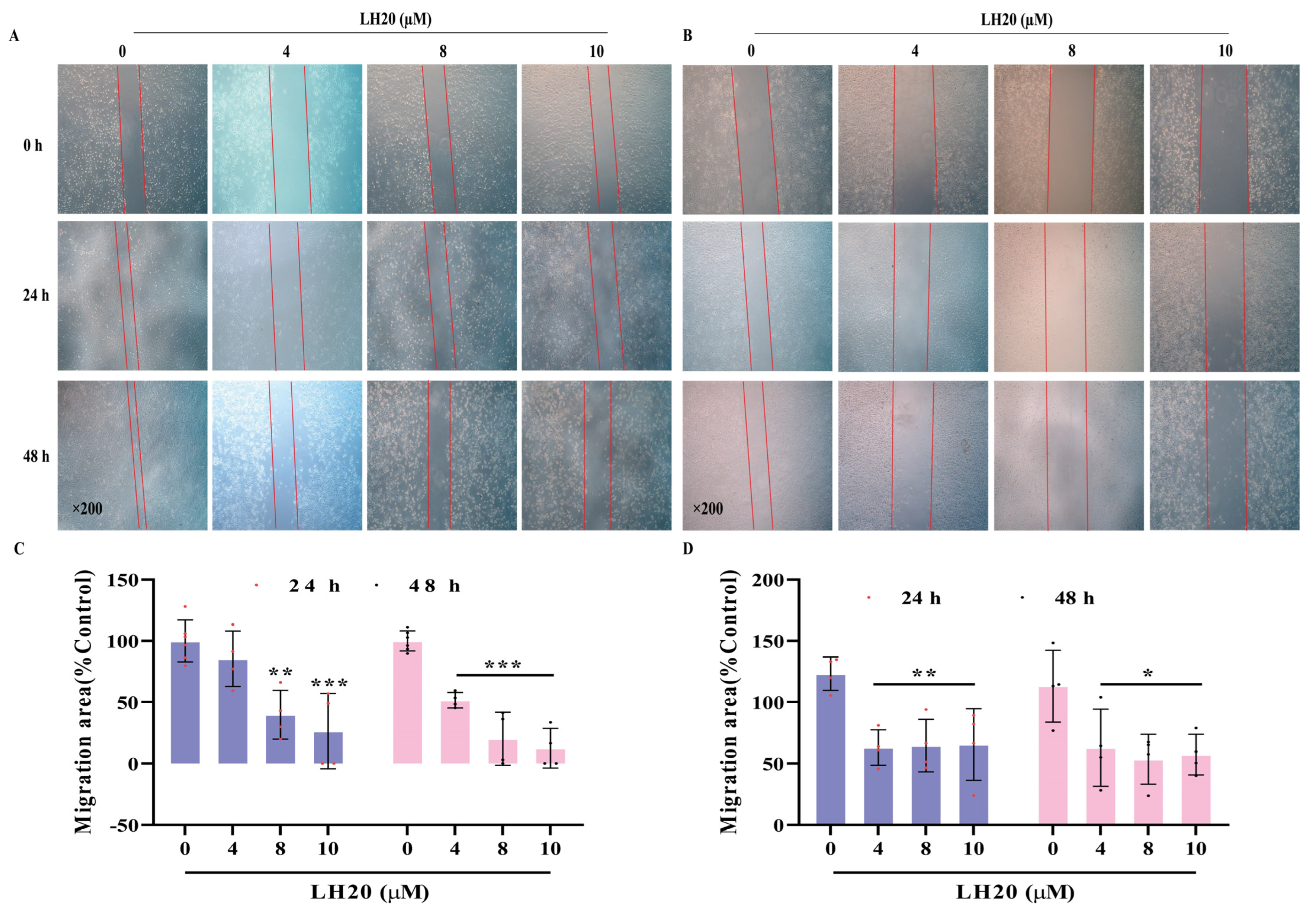
2.2. LH20 Suppresses GBM Cell Migration
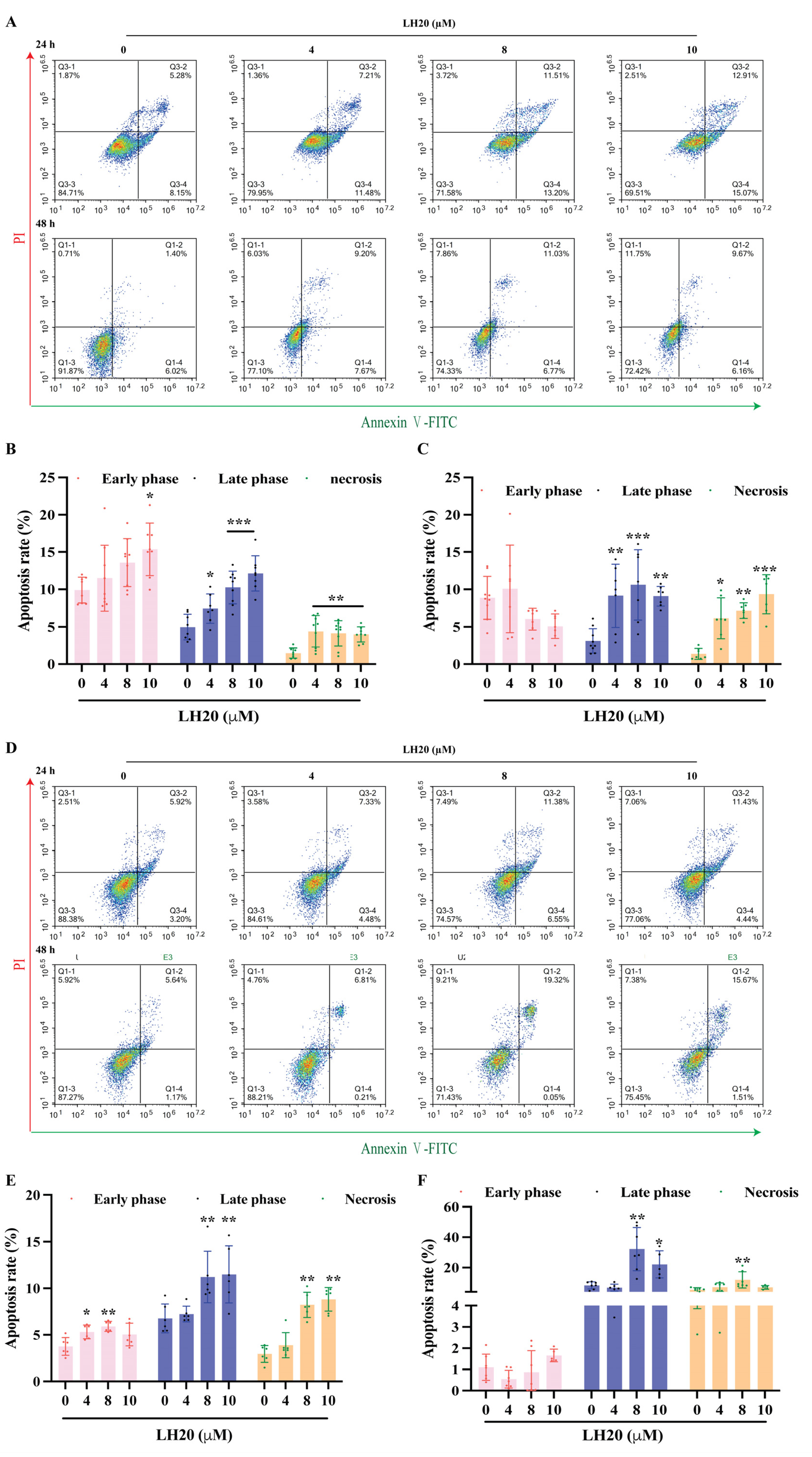
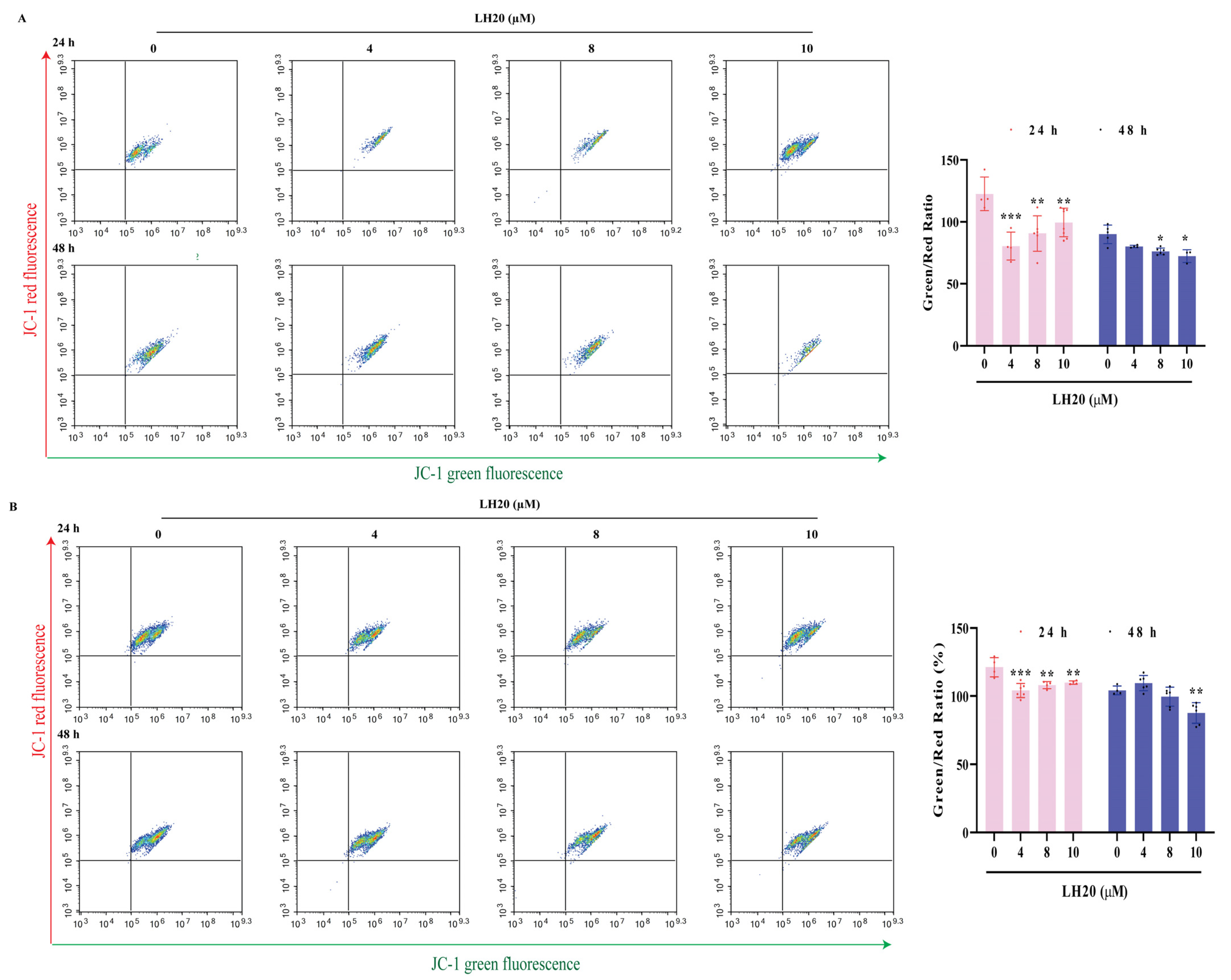
2.3. LH20 Induces GBM Cell Apoptosis and Mitochondria Membrane Potential (MMP) Drop
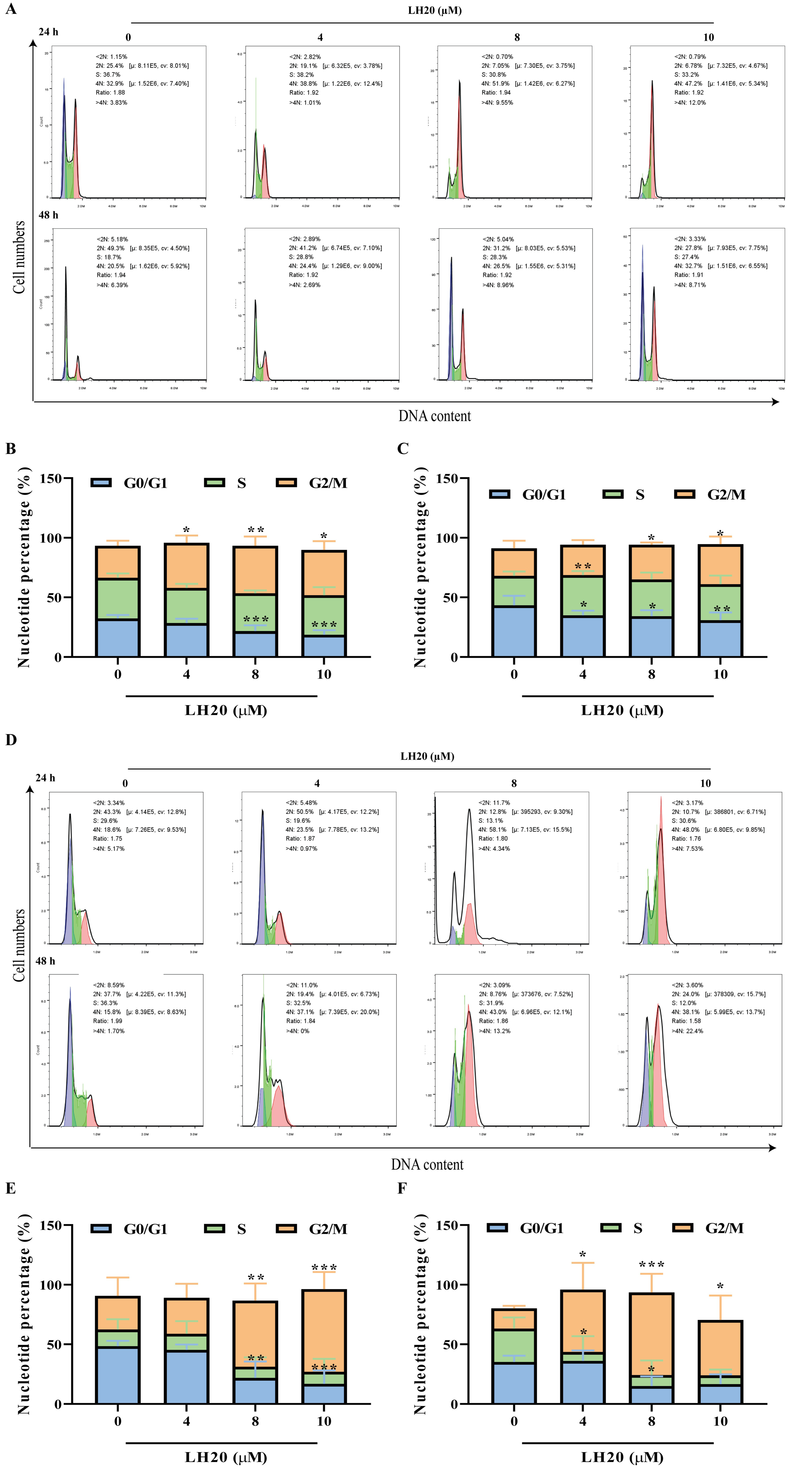
2.4. LH20 Arrested the GBM Cell Cycle at G2/M Phase
2.5. CDK4/6 Were Predicted to Be the Targets of LH20
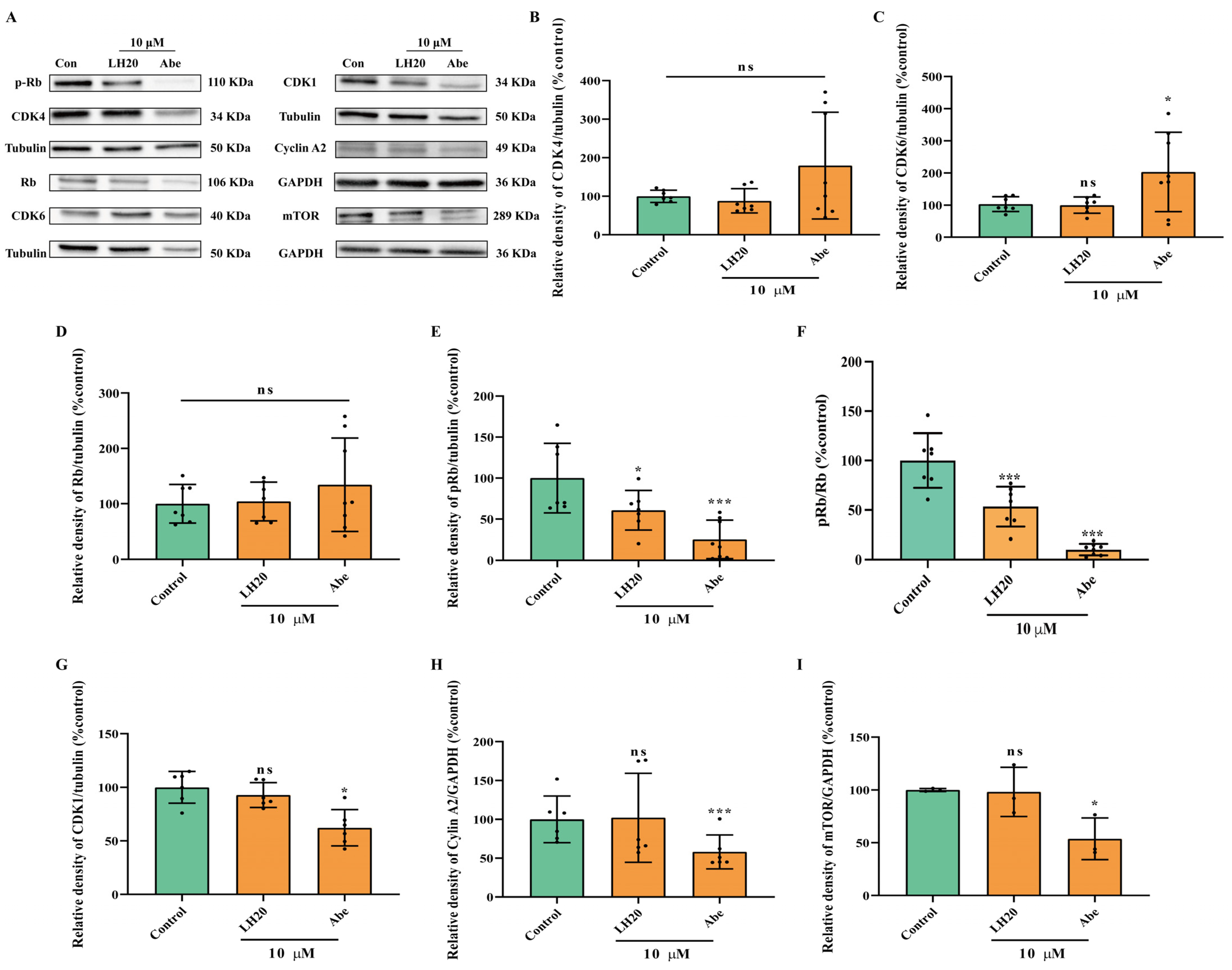
2.6. LH20 Inactivates Phosphorylation of Retinoblastoma (Rb), the Downstream Protein of CDK4/6
3. Discussion
4. Materials and Methods
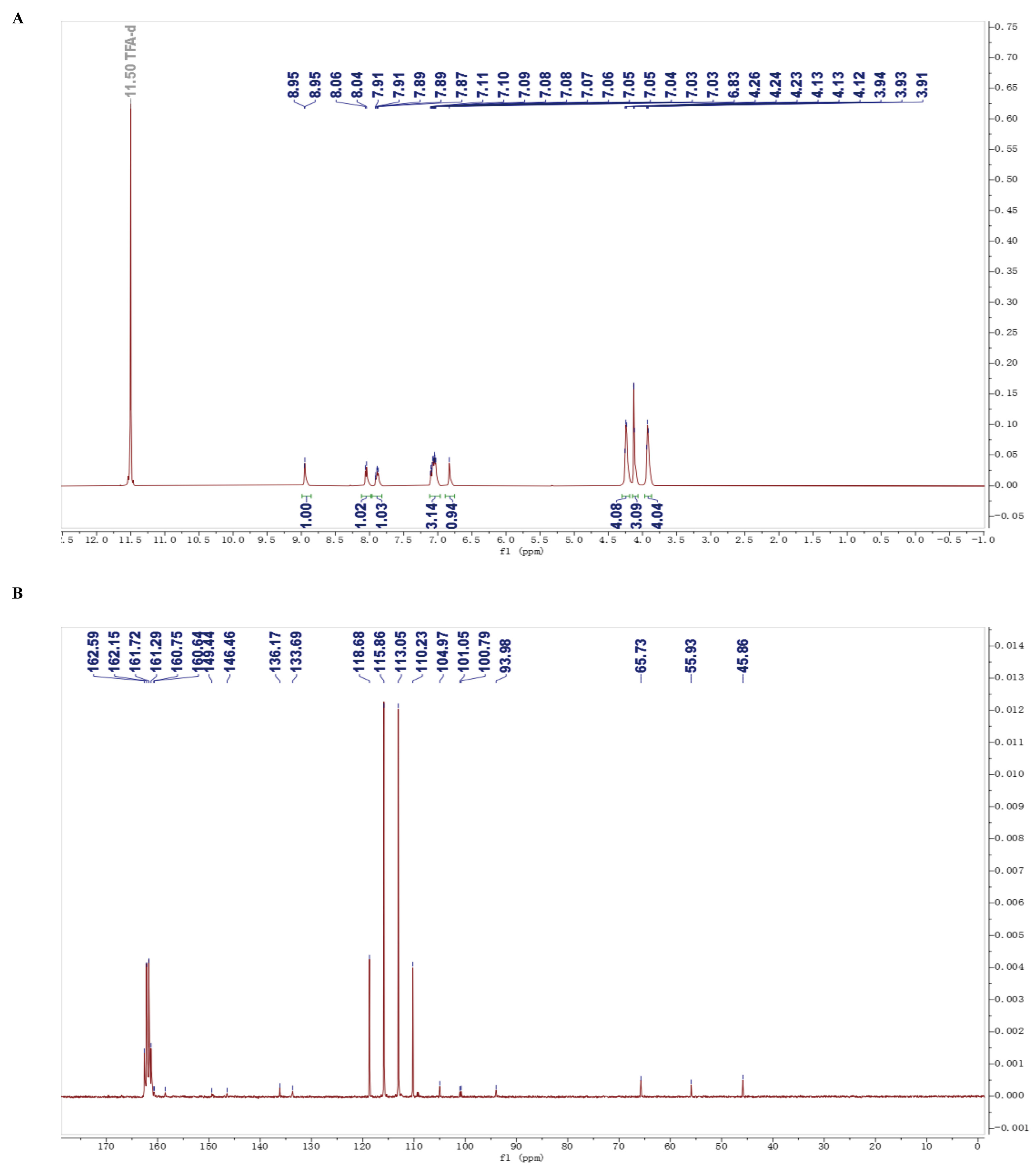
4.1. Preparation of LH20
4.2. Cell Culture
4.3. Cell Viability
4.4. LDH Assay
4.5. Wound Healing Assay
4.6. Annexin V-FITC/PI Apoptosis Assay
4.7. Determination of MMP
4.8. Cell Cycle Analysis
4.9. Predicting Targets and Pathways of LH20
4.10. Molecular Docking
4.11. Western Blotting Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ostrom, Q.T.; Price, M.; Neff, C.; Cioffi, G.; Waite, K.A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2015–2019. Neuro-Oncol. 2022, 24, v1–v95. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Faujdar, C.; Sharma, R.; Sharma, S.; Malik, B.; Nepali, K.; Liou, J.P. Glioblastoma: Current Status, Emerging Targets, and Recent Advances. J. Med. Chem. 2022, 65, 8596–8685. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Costa, A.; Osório, L.; Lago, R.C.; Linhares, P.; Carvalho, B.; Caeiro, C. Current Standards of Care in Glioblastoma Therapy. In Glioblastoma; Exon Publications: Brisbane City, Australia, 2017; pp. 197–241. [Google Scholar] [CrossRef]
- Malumbres, M.; Barbacid, M. Cell cycle, CDKs and cancer: A changing paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Lubanska, D.; Porter, L. Revisiting CDK Inhibitors for Treatment of Glioblastoma Multiforme. Drugs R&D 2017, 17, 255–263. [Google Scholar] [CrossRef]
- Bury, M.; Le Calvé, B.; Ferbeyre, G.; Blank, V.; Lessard, F. New Insights into CDK Regulators: Novel Opportunities for Cancer Therapy. Trends Cell Biol. 2021, 31, 331–344. [Google Scholar] [CrossRef]
- Brennan, C.W.; Verhaak, R.G.W.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The Somatic Genomic Landscape of Glioblastoma. Cell 2013, 155, 462–477, Erratum in Cell 2014, 157, 753. [Google Scholar] [CrossRef]
- Molinaro, A.M.; Taylor, J.W.; Wiencke, J.K.; Wrensch, M.R. Genetic and molecular epidemiology of adult diffuse glioma. Nat. Rev. Neurol. 2019, 15, 405–417. [Google Scholar] [CrossRef]
- Olmez, I.; Brenneman, B.; Xiao, A.; Serbulea, V.; Benamar, M.; Zhang, Y.; Manigat, L.; Abbas, T.; Lee, J.; Nakano, I.; et al. Combined Cdk4/6 and Mtor Inhibition Is Synergistic against Glioblastoma Via Multiple Mechanisms. Clin. Cancer Res. 2017, 23, 6958–6968. [Google Scholar] [CrossRef]
- Patnaik, A.; Rosen, L.S.; Tolaney, S.M.; Tolcher, A.W.; Goldman, J.W.; Gandhi, L.; Papadopoulos, K.P.; Beeram, M.; Rasco, D.W.; Hilton, J.F.; et al. Efficacy and Safety of Abemaciclib, an Inhibitor of CDK4 and CDK6, for Patients with Breast Cancer, Non-Small Cell Lung Cancer, and Other Solid Tumors. Cancer Discov. 2016, 6, 740–753. [Google Scholar] [CrossRef]
- Turner, N.C.; Slamon, D.J.; Ro, J.; Bondarenko, I.; Im, S.-A.; Masuda, N.; Colleoni, M.; DeMichele, A.; Loi, S.; Verma, S.; et al. Overall Survival with Palbociclib and Fulvestrant in Advanced Breast Cancer. N. Engl. J. Med. 2018, 379, 1926–1936. [Google Scholar] [CrossRef]
- Xiao, J.; Liang, J.; Fan, J.; Hou, P.; Li, X.; Zhang, H.; Li, K.; Bu, L.; Li, P.; He, M.; et al. Cdk4/6 Inhibition Enhances Oncolytic Virus Efficacy by Potentiating Tumor-Selective Cell Killing and T-Cell Activation in Refractory Glioblastoma. Cancer Res. 2022, 82, 3359–3374. [Google Scholar] [CrossRef]
- Zhao, Y.; He, J.; Li, Y.; Lv, S.; Cui, H. Nusap1 Potentiates Chemoresistance in Glioblastoma through Its Sap Domain to Stabilize ATR. Signal Transduct. Target. Ther. 2020, 5, 44. [Google Scholar] [CrossRef]
- Liu, C.A.; Chang, C.Y.; Hsueh, K.W.; Su, H.L.; Chiou, T.W.; Lin, S.Z.; Harn, H.J. Migration/Invasion of Malignant Gliomas and Implications for Therapeutic Treatment. Int. J. Mol. Sci. 2018, 19, 1115. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.W.; Li, W.; Zheng, X.J.; Liu, J.Y.; Yang, Y.H.; Li, S.; Zhang, S.; Fu, W.Q.; Xiao, B.; Wang, J.H.; et al. Benzimidazoles Induce Concurrent Apoptosis and Pyroptosis of Human Glioblastoma Cells Via Arresting Cell Cycle. Acta Pharmacol. Sin. 2022, 43, 194–208. [Google Scholar] [CrossRef]
- Riess, C.; del Moral, K.; Fiebig, A.; Kaps, P.; Linke, C.; Hinz, B.; Rupprecht, A.; Frank, M.; Fiedler, T.; Koczan, D.; et al. Implementation of a combined CDK inhibition and arginine-deprivation approach to target arginine-auxotrophic glioblastoma multiforme cells. Cell Death Dis. 2022, 13, 555. [Google Scholar] [CrossRef] [PubMed]
- Skowron, M.A.; Vermeulen, M.; Winkelhausen, A.; Becker, T.K.; Bremmer, F.; Petzsch, P.; Schonberger, S.; Calaminus, G.; Kohrer, K.; Albers, P.; et al. Cdk4/6 Inhibition Presents as a Therapeutic Option for Paediatric and Adult Germ Cell Tumours and Induces Cell Cycle Arrest and Apoptosis Via Canonical and Non-Canonical Mechanisms. Br. J. Cancer 2020, 123, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. Cyclin-dependent protein serine/threonine kinase inhibitors as anticancer drugs. Pharmacol. Res. 2019, 139, 471–488. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research, N. Comprehensive Genomic Characterization Defines Human Glioblastoma Genes and Core Pathways. Nature 2008, 455, 1061–1068. [Google Scholar] [CrossRef]
- Gonzalez-Tablas, M.; Arandia, D.; Jara-Acevedo, M.; Otero, A.; Vital, A.L.; Prieto, C.; Gonzalez-Garcia, N.; Nieto-Librero, A.B.; Tao, H.; Pascual, D.; et al. Heterogeneous EGFR, CDK4, MDM4, and PDGFRA Gene Expression Profiles in Primary GBM: No Association with Patient Survival. Cancers 2020, 12, 231. [Google Scholar] [CrossRef]
- Lam, P.Y.; Di Tomaso, E.; Ng, H.K.; Pang, J.C.; Roussel, M.F.; Hjelm, N.M. Expression of p19 INK4d, CDK4, CDK6 in glioblastoma multiforme. Br. J. Neurosurg. 2000, 14, 28–32. [Google Scholar]
- Sherr, C.J.; Roberts, J.M. Living with or without cyclins and cyclin-dependent kinases. Genes Dev. 2004, 18, 2699–2711. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A BOILED-Egg To Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef]
- Rashid, M. Design, synthesis and ADMET prediction of bis-benzimidazole as anticancer agent. Bioorganic Chem. 2020, 96, 103576. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, H.; Wang, X.; Huang, J.; Li, S.; Liu, C.; Dong, R.; Zhu, G.; Duan, C.; Jiang, F.; et al. Discovery of Coumarin Derivatives as Potent and Selective Cyclin-Dependent Kinase 9 (Cdk9) Inhibitors with High Antitumour Activity. Eur. J. Med. Chem. 2020, 200, 112424. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, Y.; Zhang, X.; Cai, H.; Zhang, C.; Qu, H.; Liu, L.; Zhang, M.; Fu, J.; Zhang, J.; et al. Oxidative Stress Activates Norad Expression by H3K27ac and Promotes Oxaliplatin Resistance in Gastric Cancer by Enhancing Autophagy Flux Via Targeting the miR-433-3p. Cell Death Dis. 2021, 12, 90. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Wang, G.; Lu, J.; Li, Z. Overexpression of Homer1b/C Induces Valproic Acid Resistance in Epilepsy. CNS Neurosci. Ther. 2023, 29, 331–343. [Google Scholar] [CrossRef]
- Lee, S.; Kwon, M.C.; Jang, J.P.; Sohng, J.K.; Jung, H.J. The Ginsenoside Metabolite Compound K Inhibits Growth, Migration and Stemness of Glioblastoma Cells. Int. J. Oncol. 2017, 51, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zhang, N.; Zhao, Q.; Xie, X.; Li, X.; Zhu, H.P.; Peng, C.; Huang, W.; Han, B. Evodiamine Alleviates Lipopolysaccharide-Induced Pulmonary Inflammation and Fibrosis by Activating Apelin Pathway. Phytother. Res. 2021, 35, 3406–3417. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhang, H.; Liu, Y.; Liu, Y.X.; Huang, L. Evenn: Easy to Create Repeatable and Editable Venn Diagrams and Venn Networks Online. J. Genet. Genom. 2021, 48, 863–866. [Google Scholar] [CrossRef]
- Tang, Y.; Li, M.; Wang, J.; Pan, Y.; Wu, F.X. Cytonca: A Cytoscape Plugin for Centrality Analysis and Evaluation of Protein Interaction Networks. Biosystems 2015, 127, 67–72. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Source | Identifier | Dilution Rate |
|---|---|---|---|
| Rabbit anti-CDK1 | Proteintech | Cat# 19532-1-AP | 1:1000 |
| Rabbit anti-CDK4 | Abcam | Cat# ab199728 | 1:2000 |
| Mouse anti-CDK6 | Santa Cruz Biotechnology | Cat# sc-7961 | 1:200 |
| Mouse anti-Rb | Santa Cruz Biotechnology | Cat# sc-102 | 1:200 |
| Rabbit anti-pRb (Ser807/811) | Cell Signaling Technology | Cat# 8516 | 1:1000 |
| Mouse anti-CylinA2 | Proteintech | Cat# 66391-1-lg | 1:5000 |
| Rabbit anti-GAPDH | Proteintech | Cat# 10494-1-AP | 1:6000 |
| Mouse anti-Tubulin | Proteintech | Cat# 66031-1-lg | 1:20,000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, Y.; Liu, D.; Zheng, D.; Li, X.; Li, C.; Huang, C.; Wang, Y.; Wang, X.; Li, Q.; et al. A Potential Anti-Glioblastoma Compound LH20 Induces Apoptosis and Arrest of Human Glioblastoma Cells via CDK4/6 Inhibition. Molecules 2023, 28, 5047. https://doi.org/10.3390/molecules28135047
Wang Y, Li Y, Liu D, Zheng D, Li X, Li C, Huang C, Wang Y, Wang X, Li Q, et al. A Potential Anti-Glioblastoma Compound LH20 Induces Apoptosis and Arrest of Human Glioblastoma Cells via CDK4/6 Inhibition. Molecules. 2023; 28(13):5047. https://doi.org/10.3390/molecules28135047
Chicago/Turabian StyleWang, Yan, Youbin Li, Dong Liu, Danyang Zheng, Xiaogang Li, Chang Li, Caihui Huang, Yun Wang, Xuesong Wang, Qifu Li, and et al. 2023. "A Potential Anti-Glioblastoma Compound LH20 Induces Apoptosis and Arrest of Human Glioblastoma Cells via CDK4/6 Inhibition" Molecules 28, no. 13: 5047. https://doi.org/10.3390/molecules28135047
APA StyleWang, Y., Li, Y., Liu, D., Zheng, D., Li, X., Li, C., Huang, C., Wang, Y., Wang, X., Li, Q., & Xu, J. (2023). A Potential Anti-Glioblastoma Compound LH20 Induces Apoptosis and Arrest of Human Glioblastoma Cells via CDK4/6 Inhibition. Molecules, 28(13), 5047. https://doi.org/10.3390/molecules28135047