
A Comprehensive Update of Various Attempts by Medicinal Chemists to Combat COVID-19 through Natural Products

 , , and
, , and
Abstract
1. Introduction
2. In Vivo Studies
3. In Vitro Studies
4. In Vitro and In Silico Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial No. | Compound | Binding Potential | Interactions at Enzyme Active Site * | References | ||
|---|---|---|---|---|---|---|
| Docking Score (kcal/mol) | Binding Free Energy (kcal/mol) | H-Bond Interactions | Hydrophobic Interactions | |||
| Main Protease (Mpro/3CLpro) | ||||||
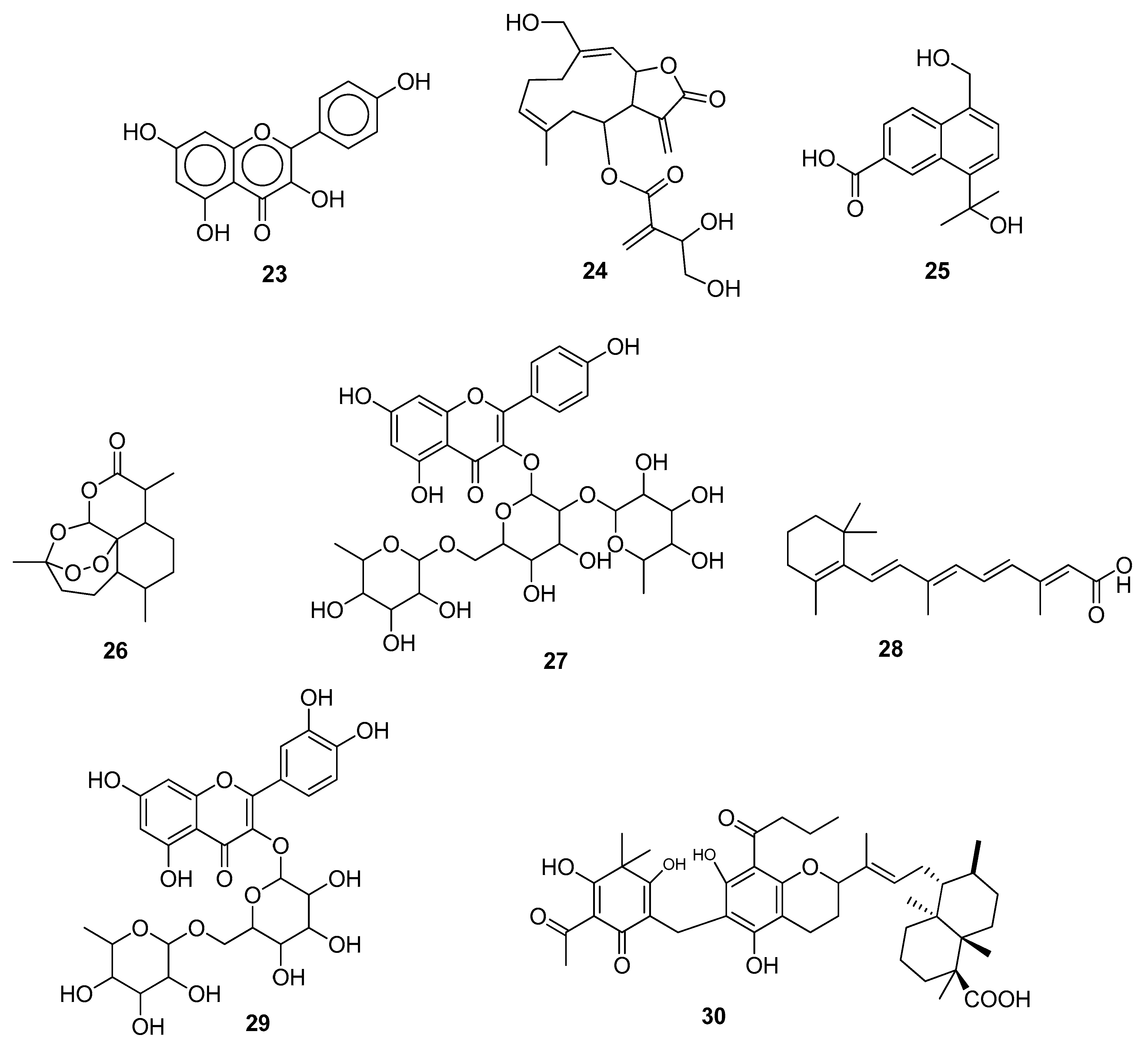
| 1 | Kaempferol (23) | −6.4 | −26.81 | PHE140, LEU141, ASN142, CYS145, ARG188 | - | [44] |
| 2 | Artemisinin (26) | −5.21 | −25.84 | ASN142 | LEU141, CYS145, VAL42, CYS44, MET49, LEU27, MET106 | [48] |
| 3 | Rutin (29) | −9.19 | - | HIS163, GLU166(2), PHE140, CYS145 | HIS41 | [51] |
| 4 | Myricetin (33) | <−8.0 | - | PHE140, GLU166, ASP187 | HIS41 | [55] |
| 5 | 5α-cholesta-4 (27), 24-dien-3β, 23 β-diol (37) | - | −24.68 | HIS41 | MET49, MET165 | [58] |
| 6 | Chlorogenic acid (39) | - | −24.9 | GLY143, CYS145 | MET165 | [60] |
| 7 | Kaempferol (23) | - | −36.5 | GLU166, MET49 | HIS41, CYS145 | [60] |
| 8 | Quercetin (40) | - | −39.66 | GLU166, GLY143, CYS145 | MET49 | [60] |
| 9 | Catechin (41) | - | −39.66 | GLU166, ASN142, HIS163, PHE140 | MET165, LEU141 | [60] |
| 10 | Amentoflavone (42) | −8.7 | - | - | CYS145 | [61] |
| Papain-Like Protease (PLpro) | ||||||
| 11 | Amentoflavone (42) | −7.7 | - | - | TYR264 | [61] |
| Spike Proteins | ||||||
| 12 | Candinone sesquiterpenoids (25) | - | - | ARG403, ARG405, ARG408, ARG393, HIS34 | GLU406, LYS417, ARG403, GLN409 | [46] |
| 13 | Artemisinin (26) | −5.06 | −30.61 | GLU23, LYS26, THR27, ASP30, LYS417, TYR421, PHE456, ARG457, TYR473 | [48] | |
| 14 | Mauritianin (27) | - | −9.4799 | ASP405, GLY496, LYS403, GLU37, ASP30 | HIS34, ALA387 | [49] |
| 15 | Kuwanon C (31) | −7.1 | - | ARG403 | TYR453, GLN493, SER494, TYR495, GLY496, PHE497, GLN498, ASN501, GLY502, TYR505 | [53] |
| 16 | Silibinin (38) | −7.78 | - | ASN907, LYS1038, ILE909, GLU1092, TYR904, ASN907, GLY904, TYR904 | TYR904, ASN907, GLY910, GLY908, LYS1038, GLU1092, TYR904 | [59] |
| 17 | Amentoflavone (42) | -8.7 | - | - | TYR449, GLN493 | [61] |
| 18 | Mulberrofuran G (45) | −8.4 | - | GLN493, TYR453 GLY496 | GLU406, LYS417, LEU455, SER494, TYR495, GLY496, GLN498, ASN501, TYR505 | [63] |
| (a) ACE2 | ||||||
| 19 | Amentoflavone (42) | −9.1 | - | - | ASP30, ASP32, GLN42, TYR83, LYS353 | [61] |
| 20 | Mulberrofuran G (45) | −7.4 | - | GLU23, THR27, ASN33, GLN96 | GLU23, GLN24, LYS26, THR27, LEU29, ASP30, VAL93, AND PRO389 | [63] |
| (b) TMPRSS2 | ||||||
| 21 | Withanoside V (43) | −7.96 | - | GLU299, TYR337, SER339 | HIS296, LYS300, TYR337, ASP338, LYS340, THR341, LYS342, ASP435, SER436, CYS437, GLN438, GLY439, SER441, THR459, SER460, TRP461, GLY462, GLY464, CYS465, GLY472, VAL473 | [62] |
| 22 | Withanoside IV (44) | −6.92 | - | ASP338, LYS342, GLU389, SER436, SER441 | HIS296, GLU299, TYR337, ASP435, CYS437, GLN438, GLY439, ASP440, THR459, SER460, TRP461, GLY462, GLY464, CYS465, ALA466, GLY472, VAL473 | [62] |
| RDRP | ||||||
| 23 | Cnicin (24) | −9.7 | −10.3 | LYS41, LEU49, LYS50, THR51, ASN52 | PHE35, ASP36, ILE37, TYR38, ASN39, VAL42, PHE48, HIS75, ASP208, ASP218, ASP221 | [45] |
| 24 | 5α-cholesta-4(27), 24-dien-3β, 23 β-diol (37) | - | −29.86 | TR680, CYS622 | URD20, ADE11, LYS545, URD10, VAL557, ALA547 | [58] |
| 25 | Silibinin (38) | −7.15 | - | U20, ASP618, ILE548, ASP618 | ARG836, ASP618, SER814, GLU811, LYS545, LYS551, ALA547, ASP760, ILE-548 | [59] |
| 26 | Amentoflavone (42) | −9.4 | - | - | ASP618 | [61] |
| Endoribonuclease | ||||||
| 27 | Cnicin (24) | −9.8 | −9.3 | GLN245, HIS250, LYS290 | ASN278, VAL292, SER294, THR341, TYR343, PRO344, LYS345, LEU346 | [45] |
| Adeno Diphosphate (ADP)-Ribose Phosphatase/ADPRP | ||||||
| 28 | Cnicin (24) | −9.2 | −10.1 | ALA38, VAL49, ALA50, LEU126, ALA129, ILE131, PHE132, ALA154, PHE156, ASP157 | ALA21, ILE23, GLY47, PRO125, SER128, LEU160 | [45] |
| Nonstructural proteins | ||||||
| 29 | 5α-cholesta-4(27),24-dien-3β,23-β-diol (37) | - | −23.47 | ASP6912 | LEU6898, PRO6932 | [58] |
5. In Silico Studies
5.1. Natural Compounds as SARS-CoV-2 Main Protease Inhibitors
5.2. Natural Compounds as SARS-CoV-2 Spike Protein Inhibitors
5.3. Natural Compounds as SARS-CoV-2 RdRp Inhibitors
5.4. Natural Compounds as SARS-CoV-2 Nucleocapsid Inhibitors
5.5. Natural Compounds as SARS-CoV-2 Endoribonuclease Inhibitors
5.6. Natural Compounds as SARS-CoV-2 Helicase Inhibitors
5.7. Natural Compounds as SARS-CoV-2 Methyltransferase Inhibitors
5.8. Natural Compounds as SARS-CoV-2 ADP-Ribose Phosphatase Inhibitors
5.9. Natural Compounds as SARS-CoV-2 Exoribonuclease Inhibitors
5.10. Natural Compounds as Inhibitors of Other SARS-CoV-2 NSPs
5.11. Natural Compounds as SARS-CoV-2 Envelope Protein Inhibitors
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asselah, T.; Durantel, D.; Pasmant, E.; Lau, G.; Schinazi, R.F. COVID-19: Discovery, Diagnostics and Drug Development. J. Hepatol. 2021, 74, 168–184. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, M.; Dinda, S.; Chakraborty, M. Some Natural Compounds and Their Analogues Having Potent Anti- SARS-CoV-2 and Anti-Proteases Activities as Lead Molecules in Drug Discovery for COVID-19. Eur. J. Med. Chem. Rep. 2022, 6, 100079. [Google Scholar] [CrossRef] [PubMed]
- Petersen, E.; Koopmans, M.; Go, U.; Hamer, D.H.; Petrosillo, N.; Castelli, F.; Storgaard, M.; Al Khalili, S.; Simonsen, L. Comparing SARS-CoV-2 with SARS-CoV and Influenza Pandemics. Lancet Infect. Dis. 2020, 20, e238–e244. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The Species Severe Acute Respiratory Syndrome-Related Coronavirus: Classifying 2019-NCoV and Naming It SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar]
- Souid, I.; Korchef, A.; Souid, S. In Silico Evaluation of Vitis Amurensis Rupr. Polyphenol Compounds for Their Inhibition Potency Against COVID-19 Main Enzymes Mpro and RdRp. Saudi Pharm. J. 2022, 30, 570–584. [Google Scholar] [CrossRef]
- Tirado-Kulieva, V.A.; Hernández-Martínez, E.; Choque-Rivera, T.J. Phenolic Compounds Versus SARS-CoV-2: An Update on the Main Findings Against COVID-19. Heliyon 2022, 8, e10702. [Google Scholar] [CrossRef]
- Yusuf, A.P.; Zhang, J.; Li, J.; Muhammad, A.; Abubakar, M.B. Herbal Medications and Natural Products for Patients with COVID-19 and Diabetes Mellitus: Potentials and Challenges. Phytomedicine Plus 2022, 2, 100280. [Google Scholar] [CrossRef]
- Prasansuklab, A.; Theerasri, A.; Rangsinth, P.; Sillapachaiyaporn, C.; Chuchawankul, S.; Tencomnao, T. Anti-COVID-19 Drug Candidates: A review on Potential Biological Activities of Natural Products in the Management of New Coronavirus Infection. J. Tradit. Complement. Med. 2021, 11, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Stobart, C.C.; Sexton, N.R.; Munjal, H.; Lu, X.; Molland, K.L.; Tomar, S.; Mesecar, A.D.; Denison, M.R. Chimeric Exchange of Coronavirus NSP5 proteases (3CLpro) Identifies Common and Divergent Regulatory Determinants of Protease Activity. J. Virol. 2013, 87, 12611–12618. [Google Scholar] [CrossRef]
- De Wit, E.; Van Doremalen, N.; Falzarano, D. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Trougakos, I.P.; Stamatelopoulos, K.; Terpos, E.; Tsitsilonis, O.E.; Aivalioti, E.; Paraskevis, D.; Kastritis, E.; Pavlakis, G.N.; Dimopoulos, M.A. Insights to SARS-CoV-2 Life Cycle, Pathophysiology, and Rationalized Treatments that Target COVID-19 Clinical Complications. J. Biomed. Sci. 2021, 28, 154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal Structure of SARS-CoV-2 Main Protease Provides a Basis for Design of Improved a-Ketoamide Inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Oladele, J.O.; Ajayi, E.I.; Oyeleke, O.M.; Oladele, O.T.; Olowookere, B.D.; Adeniyi, B.M.; Oyewole, O.I.; Oladiji, A.T. A Systematic Review on COVID-19 Pandemic with Special Emphasis on Curative Potentials of Nigeria Based Medicinal Plants. Heliyon 2020, 6, e04897. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Witek-Krowiak, A.; Skrzypczak, D.; Mikula, K.; Młynarz, P. Phytochemicals Containing Biologically Active Polyphenols as an Effective Agent Against COVID-19-Inducing Coronavirus. J. Funct. Foods 2020, 73, 104146. [Google Scholar] [CrossRef]
- Le Maréchal, M.; Morand, P.; Epaulard, O.; Némoz, B. COVID-19 in Clinical Practice: A Narrative Synthesis. Med. Mal. Infect. 2020, 50, 639–647. [Google Scholar] [CrossRef]
- Tiwari, V.; Beer, J.C.; Sankaranarayanan, N.V.; Swanson-Mungerson, M.; Desai, U.R. Discovering Small-Molecule Therapeutics Against SARS-CoV-2. Drug Discov. Today 2020, 25, 1535–1544. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Roles of Flavonoids Against Coronavirus Infection. Chem. Biol. Interact. 2020, 328, 109211. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Q.; Guo, D. Emerging Coronaviruses: Genome Structure, Replication, and Pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef]
- Dos Santos, O.V.; Pinaffi Langley, A.C.d.C.; Mota de Lima, A.J.; Vale Moraes, V.S.; Dias Soares, S.; Teixeira-Costa, B.E. Nutraceutical Potential of Amazonian Oilseeds in Modulating the Immune System Against COVID-19—A Narrative Review. J. Funct. Foods 2022, 94, 105123. [Google Scholar] [CrossRef]
- Benke, C.; Autenrieth, L.K.; Asselmann, E.; Pané-farré, C.A. Lockdown, Quarantine Measures, and Social Distancing: Associations with Depression, Anxiety and Distress at the Beginning of the COVID-19 Pandemic among Adults from Germany. Psychiatry Res. 2020, 293, 113462. [Google Scholar] [CrossRef]
- Fuzimoto, A.D.; Isidoro, C. The Antiviral and Coronavirus-Host Protein Pathways Inhibiting Properties of Herbs and Natural Compounds—Additional Weapons in the Fight Against the COVID-19 Pandemic? J. Tradit. Complement. Med. 2020, 10, 405–419. [Google Scholar] [CrossRef]
- Córdoba-Tovar, L.; Barón, P.A.R.; Marrugo-Negrete, J.; Roa-Fuentes, L.L.; Jonathan, M.P. Scientific Production on Medicinal Plants and Their Efficacy Against COVID-19: A Review and Scientometric Analysis Based on VOS viewer. Acta Ecol. Sin. 2022; in press. [Google Scholar] [CrossRef]
- Cai, Q.; Yang, M.; Liu, D.; Chen, J.; Shu, D.; Xia, J.; Liao, X.; Gu, Y.; Cai, Q.; Yang, Y.; et al. Experimental Treatment with Favipiravir for COVID-19: An Open-Label Control Study. Engineering 2020, 6, 1192–1198. [Google Scholar] [CrossRef]
- Sharma, S.; Suvalka, C.; Bharti; Joshi, M.; Bahuguna, A. Potential Inhibitors and Plant Based Technology: An Alternative Approach to Treat Corona Virus. Bioorg. Chem. 2021, 117, 105460. [Google Scholar] [CrossRef]
- Ceramella, J.; Iacopetta, D.; Sinicropi, M.S.; Andreu, I.; Mariconda, A.; Saturnino, C.; Giuzio, F.; Longo, P.; Aquaro, S.; Catalano, A. Drugs for COVID-19: An Update. Molecules 2022, 27, 8562. [Google Scholar] [CrossRef]
- Kashte, S.; Gulbake, A.; El-Amin, S.F.; Gupta, A. COVID-19 Vaccines: Rapid Development, Implications, Challenges and Future Prospects. Hum. Cell 2021, 34, 711–733. [Google Scholar] [CrossRef]
- de Albuquerque Veloso Machado, M.; Roberts, B.; Wong, B.L.H.; van Kessel, R.; Mossialos, E. The relationship Between the COVID-19 Pandemic and Vaccine Hesitancy: A Scoping Review of Literature until August 2021. Front. Public Health 2021, 9, 1370. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.I.; Ghany, S.; Gilkes, T.; Umakanthan, S. Review of COVID-19 Vaccine Subtypes, Efficacy and Geographical Distributions. Postgrad. Med. J. 2022, 98, 389–394. [Google Scholar] [CrossRef]
- Mistry, P.; Barmania, F.; Mellet, J.; Peta, K.; Strydom, A.; Viljoen, I.M.; James, W.; Gordon, S.; Pepper, M.S. SARS-CoV-2 Variants, Vaccines, and Host Immunity. Front. Immunol. 2022, 12, 5400. [Google Scholar] [CrossRef] [PubMed]
- Chhetri, B.K.; Tedbury, P.R.; Sweeney-jones, A.M.; Mani, L.; Soapi, K.; Manfredi, C.; Sorscher, E.; Sara, S.G.; Kubanek, J. Marine Natural Products as Leads Against SARS-CoV-2 Infection. J. Nat. Prod. 2022, 85, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Yao, S.; Zhao, W.; Li, M.; Liu, J.; Shang, W.J.; Xie, H.; Ke, C.; Gao, M.; Yu, K.; et al. Discovery of Baicalin and Baicalein as Novel, Natural Product Inhibitors of SARS-CoV-2 3CL Protease In Vitro. BioRxiv 2020. [Google Scholar] [CrossRef]
- Liu, H.; Ye, F.; Sun, Q.; Liang, H.; Li, C.; Li, S.; Lu, R.; Huang, B.; Tan, W.; Lai, L. Scutellaria Baicalensis Extract and Baicalein Inhibit Replication of SARS-CoV-2 and Its 3C-like Protease In Vitro. J. Enzym. Inhib. Med. Chem. 2021, 36, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Zandi, K.; Musall, K.; Oo, A.; Cao, D.; Liang, B.; Hassandarvish, P.; Lan, S.; Slack, R.L.; Kirby, K.A.; Bassit, L.; et al. Baicalein and Baicalin Inhibit SARS-CoV-2 RNA-dependent-RNA polymerase. Microorganisms 2021, 9, 893. [Google Scholar] [CrossRef] [PubMed]
- He, C.L.; Huang, L.Y.; Wang, K.; Gu, C.J.; Hu, J.; Zhang, G.J.; Xu, W.; Xie, Y.H.; Tang, N.; Huang, A.L. Identification of bis-Benzylisoquinoline Alkaloids as SARS-CoV-2 Entry Inhibitors from a Library of Natural Products. Signal Transduct. Target. Ther. 2021, 6, 131. [Google Scholar] [CrossRef]
- Sa-Ngiamsuntorn, K.; Suksatu, A.; Pewkliang, Y.; Thongsri, P.; Kanjanasirirat, P.; Manopwisedjaroen, S.; Charoensutthivarakul, S.; Wongtrakoongate, P.; Pitiporn, S.; Chaopreecha, J.; et al. Anti-SARS-CoV-2 Activity of Andrographis Paniculata Extract and Its Major Component Andrographolide in Human Lung Epithelial Cells and Cytotoxicity Evaluation in Major Organ Cell Representatives. J. Nat. Prod. 2021, 84, 1261–1270. [Google Scholar] [CrossRef]
- Saadh, M.J. Sauchinone with Zinc Sulphate Significantly Inhibits the Activity of SARS-CoV-2 3CL-Protease. Pharmacologyonline 2021, 2, 242–248. [Google Scholar]
- Brown, A.N.; Strobel, G.; Hanrahan, K.C.; Sears, J. Antiviral Activity of the Propylamylatintm Formula Against the Novel Coronavirus SARS-CoV-2 In Vitro Using Direct Injection and Gas Assays in Virus Suspensions. Viruses 2021, 13, 415. [Google Scholar] [CrossRef]
- Liu, J.; Bodnar, B.H.; Meng, F.; Khan, A.I.; Wang, X.; Saribas, S.; Wang, T.; Lohani, S.C.; Wang, P.; Wei, Z.; et al. Epigallocatechin Gallate from Green Tea Effectively Blocks Infection of SARS-CoV-2 and New Variants by Inhibiting Spike Binding to ACE2 Receptor. Cell Biosci. 2021, 11, 168. [Google Scholar] [CrossRef]
- Ngwe Tun, M.M.; Luvai, E.; Nwe, K.M.; Toume, K.; Mizukami, S.; Hirayama, K.; Komatsu, K.; Morita, K. Anti-SARS-CoV-2 Activity of Various PET-Bottled Japanese Green Teas and Tea Compounds In Vitro. Arch. Virol. 2022, 167, 1547–1557. [Google Scholar] [CrossRef]
- Liu, Q.; Kwan, K.Y.; Cao, T.; Yan, B.; Ganesan, K.; Jia, L.; Zhang, F.; Lim, C.; Wu, Y.; Feng, Y.; et al. Broad-Spectrum Antiviral Activity of Spatholobus Suberectus Dunn Against SARS-CoV-2, SARS-CoV-1, H5N1, and Other Enveloped Viruses. Phyther. Res. 2022, 36, 3232–3247. [Google Scholar] [CrossRef]
- Eggers, M.; Jungke, P.; Wolkinger, V.; Bauer, R.; Kessler, U.; Frank, B. Antiviral Activity of Plant Juices and Green Tea against SARS-CoV-2 and Influenza Virus. Phyther. Res. 2022, 36, 2109–2115. [Google Scholar] [CrossRef] [PubMed]
- Baeshen, M.N.; Attar, R.; Bouback, T.A.; Albeshri, A.O.; Baeshen, N.N.; Karkashan, A.; Abbas, B.; Aljaddawi, A.A.; Almulaiky, Y.Q.; Mahmoud, S.H.; et al. Assaying for Antiviral Activity of the Folkloric Medicinal Desert Plant Rhazya Stricta on Coronavirus SARS-CoV-2. Biotechnol. Biotechnol. Equip. 2022, 36, 67–73. [Google Scholar] [CrossRef]
- Shin, J.A.; Oh, S.; Jeong, J.M. The Potential of BEN815 as an Anti-Inflammatory, Antiviral and Antioxidant Agent for the Treatment of COVID-19. Phytomedicine Plus 2021, 1, 100058. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Heng, W.; Wang, Y.; Qiu, J.; Wei, X.; Peng, S.; Saleem, S.; Khan, M.; Ali, S.S.; Wei, D.Q. In Silico and In Vitro Evaluation of Kaempferol as a Potential Inhibitor of the SARS-CoV-2 Main Protease (3CLpro). Phyther. Res. 2021, 35, 2841–2845. [Google Scholar] [CrossRef]
- Alhadrami, H.A.; Sayed, A.M.; Hassan, H.M.; Youssif, K.A.; Gaber, Y.; Moatasim, Y.; Kutkat, O.; Mostafa, A.; Ali, M.A.; Rateb, M.E.; et al. Cnicin as an Anti-SARS-CoV-2: An Integrated in Silico and In Vitro Approach for the Rapid Identification of Potential COVID-19 Therapeutics. Antibiotics 2021, 10, 542. [Google Scholar] [CrossRef]
- Li, X.; Gao, J.; Li, M.; Cui, H.; Jiang, W.; Tu, Z.C.; Yuan, T. Aromatic Cadinane Sesquiterpenoids from the Fruiting Bodies of Phellinus Pini Block SARS-CoV-2 Spike-ACE2 Interaction. J. Nat. Prod. 2021, 84, 2385–2389. [Google Scholar] [CrossRef]
- Hijikata, A.; Shionyu-Mitsuyama, C.; Nakae, S.; Shionyu, M.; Ota, M.; Kanaya, S.; Hirokawa, T.; Nakajima, S.; Watashi, K.; Shirai, T. Evaluating Cepharanthine Analogues as Natural Drugs against SARS-CoV-2. FEBS Open Bio 2022, 12, 285–294. [Google Scholar] [CrossRef]
- Dogan, K.; Erol, E.; Didem Orhan, M.; Degirmenci, Z.; Kan, T.; Gungor, A.; Yasa, B.; Avsar, T.; Cetin, Y.; Durdagi, S.; et al. Instant Determination of the Artemisinin from Various Artemisia annua L. Extracts by LC-ESI-MS/MS and Their In-Silico Modelling and In Vitro Antiviral Activity Studies Against SARS-CoV-2. Phytochem. Anal. 2022, 33, 303–319. [Google Scholar] [CrossRef]
- Owis, A.I.; El-Hawary, M.S.; El Amir, D.; Refaat, H.; Alaaeldin, E.; Aly, O.M.; Elrehany, M.A.; Kamel, M.S. Flavonoids of Salvadora persica L. (Meswak) and Its Liposomal Formulation as a Potential Inhibitor of SARS-CoV-2. RSC Adv. 2021, 11, 13537–13544. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Miyakawa, K.; Jeremiah, S.S.; Yamaoka, Y.; Sada, M.; Kuniyoshi, T.; Yang, J.; Kimura, H.; Ryo, A. All-Trans Retinoic Acid Exhibits Antiviral Effect Against SARS-CoV-2 by Inhibiting 3CLpro Activity. Viruses 2021, 13, 1669. [Google Scholar] [CrossRef] [PubMed]
- Alsfouk, A.A.; Taher, A.T.; Al-karmalawy, A.A. Pimenta dioica (L.) Merr. Bioactive Constituents Exert Anti-SARS-CoV-2 and Anti-Inflammatory Activities: Molecular Docking and Dynamics, In Vitro, and In Vivo Studies. Molecules 2021, 26, 5844. [Google Scholar]
- Hou, B.; Zhang, Y.; Liao, H.; Fu, L.; Li, D.; Zhao, X.; Qi, J.; Yang, W.; Xiao, G.; Yang, L.; et al. Target-Based Virtual Screening and LC/MS-Guided Isolation Procedure for Identifying Phloroglucinol-Terpenoid Inhibitors of SARS-CoV-2. J. Nat. Prod. 2022, 85, 327–336. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kwon, E.B.; Kim, B.; Chung, H.S.; Choi, G.; Kim, Y.H.; Choi, J.G. Mulberry Component Kuwanon C Exerts Potent Therapeutic Efficacy In Vitro against COVID-19 by Blocking the SARS-CoV-2 Spike S1 RBD: ACE2 Receptor Interaction. Int. J. Mol. Sci. 2022, 23, 12516. [Google Scholar] [CrossRef]
- Corona, A.; Wycisk, K.; Talarico, C.; Manelfi, C.; Milia, J.; Cannalire, R.; Esposito, F.; Gribbon, P.; Zaliani, A.; Iaconis, D.; et al. Natural Compounds Inhibit SARS-CoV-2 Nsp13 Unwinding and ATPase Enzyme Activities. ACS Pharmacol. Transl. Sci. 2022, 5, 226–239. [Google Scholar] [CrossRef]
- Xiao, T.; Cui, M.; Zheng, C.; Wang, M.; Sun, R.; Gao, D.; Bao, J.; Ren, S.; Yang, B.; Lin, J.; et al. Myricetin Inhibits SARS-CoV-2 Viral Replication by Targeting Mpro and Ameliorates Pulmonary Inflammation. Front. Pharmacol. 2021, 12, 669642. [Google Scholar] [CrossRef]
- Nawrot-Hadzik, I.; Zmudzinski, M.; Matkowski, A.; Preissner, R.; Kęsik-Brodacka, M.; Hadzik, J.; Drag, M.; Abel, R. Reynoutria Rhizomes as a Natural Source of SARS-CoV-2 Mpro Inhibitors–Molecular Docking and In Vitro Study. Pharmaceuticals 2021, 14, 742. [Google Scholar] [CrossRef]
- Chaves, O.A.; Fintelman-rodrigues, N.; Wang, X.; Sacramento, C.Q.; Temerozo, J.R.; Ferreira, A.C.; Mattos, M.; Pereira-dutra, F.; Bozza, T.; Castro-faria-neto, H.C.; et al. Commercialy available flavonols are better SARS-CoV-2 inhibitors than isoflavone and flavones. Viruses 2022, 14, 1458. [Google Scholar] [CrossRef] [PubMed]
- Abd El Hafez, M.S.M.; AbdEl-Wahab, M.G.; Seadawy, M.G.; El-Hosseny, M.F.; Beskales, O.; Saber Ali Abdel-Hamid, A.; El Demellawy, M.A.; Ghareeb, D.A. Characterization, In-Silico, and In-Vitro Study of a New Steroid Derivative from Ophiocoma Dentata as a Potential Treatment for COVID-19. Sci. Rep. 2022, 12, 5846. [Google Scholar] [CrossRef] [PubMed]
- Hamdy, R.; Mostafa, A.; Abo Shama, N.M.; Soliman, S.S.M.; Fayed, B. Comparative Evaluation of Flavonoids Reveals the Superiority and Promising Inhibition Activity of Silibinin against SARS-CoV-2. Phyther. Res. 2022, 36, 2921–2939. [Google Scholar] [CrossRef] [PubMed]
- Elhusseiny, S.M.; El-mahdy, T.S.; Elleboudy, N.S.; Yahia, I.S. In Vitro Anti SARS-CoV-2 Activity and Docking In Vitro Anti SARS-CoV-2 Activity and Docking Analysis of Pleurotus ostreatus, Lentinula edodes and Agaricus bisporus Edible Mushrooms. Infect. Drug Resist. 2022, 15, 3459–3475. [Google Scholar]
- Jorge, A.; Lopes, O.; Calado, G.P.; Nascimento, Y.; Alves, S.; Ara, D.; França, L.M.; Marcus, A.; Paes, D.A. Plant Metabolites as SARS-CoV-2 Inhibitors Candidates: In Silico and In Vitro Studies. Pharmaceutics 2022, 15, 1045. [Google Scholar]
- Kaur, J.; Kumar, V.; Garg, S.; Subramani, C.; Agarwal, S.; Wang, J.; Zhang, H.; Kaul, A.; Singh, R.; Kaul, S.C.; et al. Molecular Mechanism of Anti-SARS-CoV2 Activity of Ashwagandha-Derived Withanolides. Int. J. Biol. Macromol. 2021, 184, 297–312. [Google Scholar]
- Kim, Y.S.; Kim, B.; Kwon, E.B.; Chung, H.S.; Choi, J.G. Mulberrofuran G, a Mulberry Component, Prevents SARS-CoV-2 Infection by Blocking the Interaction between SARS-CoV-2 Spike Protein S1 Receptor-Binding Domain and Human Angiotensin-Converting Enzyme 2 Receptor. Nutrients 2022, 14, 4170. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, M.; Ghaani, M.R.; Bayazeid, O. Phenylethanoid Glycosides as a Possible COVID-19 Protease Inhibitor: A Virtual Screening Approach. J. Mol. Model. 2021, 27, 341. [Google Scholar] [CrossRef] [PubMed]
- Chtita, S.; Fouedjou, R.T.; Belaidi, S.; Djoumbissie, L.A.; Ouassaf, M.; Qais, F.A.; Bakhouch, M.; Efendi, M.; Tok, T.T.; Bouachrine, M.; et al. In Silico Investigation of Phytoconstituents from Cameroonian Medicinal Plants towards COVID-19 Treatment. Struct. Chem. 2022, 33, 1799–1813. [Google Scholar] [CrossRef]
- Hajji, H.; Alaqarbeh, M.; Lakhlifi, T.; Ajana, M.A.; Alsakhen, N.; Bouachrine, M. Computational Approach Investigation Bioactive Molecules from SaussureaCostus Plant as SARS-CoV-2 Main Protease Inhibitors Using Reverse Docking, Molecular Dynamics Simulation, and Pharmacokinetic ADMET Parameters. Comput. Biol. Med. 2022, 150, 106209. [Google Scholar] [CrossRef]
- Parida, P.K.; Paul, D.; Chakravorty, D. The Natural Way Forward: Molecular Dynamics Simulation Analysis of Phytochemicals from Indian Medicinal Plants as Potential Inhibitors of SARS-CoV-2 Targets. Phyther. Res. 2020, 34, 3420–3433. [Google Scholar] [CrossRef]
- Adem, S.; Eyupoglu, V.; Ibrahim, I.M.; Sarfraz, I.; Rasul, A.; Ali, M.; Elfiky, A.A. Multidimensional in silico strategy for identification of natural polyphenols-based SARS-CoV-2 main protease (Mpro) inhibitors to unveil a hope against COVID-19. Comput. Biol. Med. 2022, 145, 105452. [Google Scholar] [CrossRef]
- Elmi, A.; Sayem, S.A.J.; Ahmed, M.; Abdoul-Latif, F. Natural Compounds from Djiboutian Medicinal Plants as Inhibitors of COVID-19 by In Silico Investigations. Int. J. Curr. Pharm. Res. 2020, 12, 52–57. [Google Scholar] [CrossRef]
- Ebada, S.S.; Al-Jawabri, N.A.; Youssef, F.S.; El-Kashef, D.H.; Knedel, T.O.; Albohy, A.; Korinek, M.; Hwang, T.L.; Chen, B.H.; Lin, G.H.; et al. Anti-Inflammatory, Antiallergic and COVID-19 Protease Inhibitory Activities of Phytochemicals from the Jordanian Hawksbeard: Identification, Structure-Activity Relationships, Molecular Modeling and Impact on Its Folk Medicinal Uses. RSC Adv. 2020, 10, 38128–38141. [Google Scholar] [CrossRef]
- Rakib, A.; Paul, A.; Chy, N.U.; Sami, S.A.; Baral, S.K.; Majumder, M.; Tareq, A.M.; Amin, M.N. Biochemical and Computational Approach of Selected Phytocompounds from Tinospora Crispa in the Management of COVID-19. Molecules 2020, 25, 3936. [Google Scholar] [CrossRef]
- Verma, S.; Patel, C.N.; Chandra, M. Identification of Novel Inhibitors of SARS-CoV-2 Main Protease (Mpro) from Withania Sp. by Molecular Docking and Molecular Dynamics Simulation. J. Comput. Chem. 2021, 42, 1861–1872. [Google Scholar] [CrossRef]
- Mujwar, S.; Sun, L.; Fidan, O. In Silico Evaluation of Food-Derived Carotenoids against 2 Drug Targets: Crocin Is a Promising Dietary Supplement Candidate for COVID-19. J. Food Biochem. 2022, 46, e14219. [Google Scholar] [CrossRef]
- Johnson, T.O. A Computational Approach to Elucidate the Interactions of Chemicals from Artemisia Annua Targeted Toward SARS-CoV-2 Main Protease Inhibition for COVID-19 Treatment. Front. Med. 2022, 9, 907483. [Google Scholar] [CrossRef]
- Prajapati, J.; Patel, R.; Goswami, D.; Saraf, M.; Rawal, R.M. Sterenin M as a Potential Inhibitor of SARS-CoV-2 Main Protease Identified from MeFSAT Database Using Molecular Docking, Molecular Dynamics Simulation and Binding Free Energy Calculation. Comput. Biol. Med. 2021, 135, 104568. [Google Scholar] [CrossRef]
- Zarei, A.; Ramazani, A.; Pourmand, S.; Sattari, A.; Rezaei, A. In Silico Evaluation of COVID-19 Main Protease Interactions with Honeybee Natural Products for Discovery of High Potential Antiviral Compounds. Nat. Prod. Res. 2022, 36, 4254–4260. [Google Scholar] [CrossRef]
- Vijayakumar, B.G.; Ramesh, D.; Joji, A.; Jayachandraprakasan, J.; Kannan, T. In Silico Pharmacokinetic and Molecular Docking Studies of Natural Flavonoids and Synthetic Indole Chalcones Against Essential Proteins of SARS-CoV-2. Eur. J. Pharmacol. 2020, 886, 173448. [Google Scholar] [CrossRef] [PubMed]
- Tung, B.T.; Nhung, N.H.; Thi, T.; Hang, T.; Linh, V.K.; Ngoc, L.M. Evaluating Natural Product Compound Inhibitors of SARS-CoV-2 Main Protease and Spike Protein Target by Molecular Docking Approach. Vietnam J. Chem. 2021, 59, 846–861. [Google Scholar]
- De Leon, V.N.O.; Manzano, J.A.H.; Pilapil, D.Y.H.; Fernandez, R.A.T.; Ching, J.K.A.R.; Quimque, M.T.J.; Agbay, J.C.M.; Notarte, K.I.R.; Macabeo, A.P.G. Anti-HIV Reverse Transcriptase Plant Polyphenolic Natural Products with In Silico Inhibitory Properties on Seven Non-Structural Proteins Vital in SARS-CoV-2 Pathogenesis. J. Genet. Eng. Biotechnol. 2021, 19, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mpiana, P.T.; Ngbolua, K.t.N.; Tshibangu, D.S.T.; Kilembe, J.T.; Gbolo, B.Z.; Mwanangombo, D.T.; Inkoto, C.L.; Lengbiye, E.M.; Mbadiko, C.M.; Matondo, A.; et al. Identification of Potential Inhibitors of SARS-CoV-2 Main Protease from Aloe Vera Compounds: A Molecular Docking Study. Chem. Phys. Lett. 2020, 754, 137751. [Google Scholar] [CrossRef]
- Ullah, S.; Munir, B.; Al-Sehemi, A.G.; Muhammad, S.; Haq, I.U.; Aziz, A.; Ahmed, B.; Ghaffar, A. Identification of Phytochemical Inhibitors of SARS-CoV-2 Protease 3CLpro from Selected Medicinal Plants as per Molecular Docking, Bond Energies and Amino Acid Binding Energies. Saudi J. Biol. Sci. 2022, 29, 103274. [Google Scholar] [CrossRef] [PubMed]
- Khanh, N.T.; Hoa, T.T. In Silico Studies of Natural Products from Medicinal Plants to Identify Potential Inhibitors for SARS-CoV-2 3C-like Protease. Vietnam J. Chem. 2021, 59, 557–562. [Google Scholar]
- Sisakht, M.; Mahmoodzadeh, A.; Darabian, M. Plant-Derived Chemicals as Potential Inhibitors of SARS-CoV-2 Main Protease (6LU7), A Virtual Screening Study. Phyther. Res. 2021, 35, 3262–3274. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, P.; Pathak, P. Neuroprotective Immunity by Essential Nutrient “Choline” for the Prevention of SARS-CoV-2 Infections: An In Silico Study by Molecular Dynamics Approach. Chem. Phys. Lett. 2020, 761, 138057. [Google Scholar] [CrossRef] [PubMed]
- Jamhour, R.M.A.Q.; Al-Nadaf, A.H.; Wedian, F.; Al-Mazaideh, G.M.; Mustafa, M.; Huneif, M.A.; Mahmoud, S.Y.; Farrag, E.S.; Al-Rimawi, F.; Salman, H.A.; et al. Phytochemicals As a Potential Inhibitor of COVID-19: An In-Silico Perspective. Russ. J. Phys. Chem. A 2022, 96, 1589–1597. [Google Scholar] [CrossRef]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Fatimawali; Kepel, B.J.; Idroes, R.; Effendi, Y.; Sakib, S.A.; Emran, T.B. Potential of Plant Bioactive Compounds as SARS-CoV-2 Main Protease (Mpro) and Spike (S) Glycoprotein Inhibitors: A Molecular Docking Study. Scientifica 2020, 2020, 6307457. [Google Scholar] [CrossRef]
- Shady, N.H.; Hayallah, A.M.; Mohamed, M.F.A.; Ghoneim, M.M.; Chilingaryan, G.; Al-Sanea, M.M.; Fouad, M.A.; Kamel, M.S.; Abdelmohsen, U.R. Targeting 3CLPRO and SARS-CoV-2 RDRP by Amphimedon sp. Metabolites: A Computational Study. Molecules 2021, 26, 3775. [Google Scholar] [CrossRef]
- Kumar, V.; Parate, S.; Yoon, S.; Lee, G.; Lee, K.W. Computational Simulations Identified Marine-Derived Natural Bioactive Compounds as Replication Inhibitors of SARS-CoV-2. Front. Microbiol. 2021, 12, 647295. [Google Scholar] [CrossRef]
- Gyebi, G.A.; Ogunyemi, O.M.; Ibrahim, I.M.; Afolabi, S.O.; Adebayo, J.O. Dual Targeting of Cytokine Storm and Viral Replication in COVID-19 by Plant-Derived Steroidal Pregnanes: An In Silico Perspective. Comput. Biol. Med. 2021, 134, 104406. [Google Scholar] [CrossRef]
- Mehmood, A.; Khan, S.; Khan, S.; Ahmed, S.; Ali, A.; Xue, M.; Ali, L.; Hamza, M.; Munir, A.; ur Rehman, S.; et al. In Silico Analysis of Quranic and Prophetic Medicinals Plants for the Treatment of Infectious Viral Diseases Including Corona Virus. Saudi J. Biol. Sci. 2021, 28, 3137–3151. [Google Scholar] [CrossRef]
- Elsbaey, M.; Ibrahim, M.A.A.; Bar, F.A.; Elgazar, A.A. Chemical Constituents from Coconut Waste and Their In-Silico Evaluation as Potential Antiviral Agents against SARS-CoV-2. S. Afr. J. Bot. 2021, 141, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Irfan, A.; Imran, M.; Khalid, N.; Hussain, R.; Basra, M.A.R.; Khaliq, T.; Shahzad, M.; Hussien, M.; Shah, A.T.; Qayyum, M.A.; et al. Isolation of Phytochemicals from Malva neglectaWallr and their Quantum Chemical, Molecular Docking Exploration as Active Drugs against COVID-19. J. Saudi Chem. Soc. 2021, 25, 101358. [Google Scholar] [CrossRef]
- Panagiotopoulos, A.A.; Karakasiliotis, I.; Kotzampasi, D.M.; Dimitriou, M.; Sourvinos, G.; Kampa, M.; Pirintsos, S.; Castanas, E.; Daskalakis, V. Natural Polyphenols Inhibit the Dimerization of the SARS-CoV-2 Main Protease: The Case of Fortunellin and Its Structural Analogs. Molecules 2021, 26, 6068. [Google Scholar] [CrossRef] [PubMed]
- Abodunrin, O.P.; Onifade, O.F.; Adegboyega, A.E. Therapeutic Capability of Five Active Compounds in Typical African Medicinal Plants Against Main Proteases of SARS-CoV-2 by Computational Approach. Inform. Med. Unlocked 2022, 31, 100964. [Google Scholar] [CrossRef] [PubMed]
- Yalçın, S.; Yalçınkaya, S.; Ercan, F. In Silico Detection of Inhibitor Potential of Passiflora Compounds Against SARS-CoV-2(COVID-19) Main Protease by Using Molecular Docking and Dynamic Analyses. J. Mol. Struct. 2021, 1240, 130556. [Google Scholar] [CrossRef]
- Rauf, A.; Rashid, U.; Khalil, A.A.; Khan, S.A.; Anwar, S.; Alafnan, A.; Alamri, A.; Rengasamy, K.R. Docking-Based Virtual Screening and Identification of Potential COVID-19 Main Protease Inhibitors from Brown Algae. S. Afr. J. Bot. 2021, 143, 428–434. [Google Scholar] [CrossRef]
- Ibrahim, M.A.A.; Abdelrahman, A.H.M.; Atia, M.A.M.; Mohamed, T.A.; Moustafa, M.F.; Hakami, A.R.; Khalifa, S.A.M.; Alhumaydhi, F.A.; Alrumaihi, F.; Abidi, S.H.; et al. Blue Biotechnology: Computational Screening of Sarcophyton Cembranoid Diterpenes for SARS-CoV-2 Main. Mar. Drugs 2021, 19, 391. [Google Scholar] [CrossRef]
- Panikar, S.; Shoba, G.; Arun, M.; Sahayarayan, J.J.; Usha Raja Nanthini, A.; Chinnathambi, A.; Alharbi, S.A.; Nasif, O.; Kim, H.J. Essential Oils as an Effective Alternative for the Treatment of COVID-19: Molecular Interaction Analysis of Protease (Mpro) with Pharmacokinetics and Toxicological Properties. J. Infect. Public Health 2021, 14, 601–610. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Kumar, A.; Srivastava, H.; Misra, N. The Role of Herbal Plants in the Inhibition of SARS-CoV-2 Main Protease: A Computational Approach. J. Indian Chem. Soc. 2022, 99, 100640. [Google Scholar] [CrossRef]
- Vijayakumar, M.; Janani, B.; Kannappan, P.; Renganathan, S.; Al-ghamdi, S.; Alsaidan, M.; Abdelaziz, M.A.; Peer, A.; Shahid, M.; Ramesh, T. In Silico Identification of Potential Inhibitors Against Main Protease of SARS-CoV-2 6LU7 from Andrographis Panniculata via Molecular Docking, Binding Energy Calculations and Molecular Dynamics Simulation Studies. Saudi J. Biol. Sci. 2022, 29, 18–29. [Google Scholar] [CrossRef]
- Bora, A.; Pacureanu, L.; Crisan, L. In Silico Study of Some Natural Flavonoids as Potential Agents against COVID-19: Preliminary Results. Chem. Proc. 2021, 3, 25. [Google Scholar]
- Imran, M.; Iqbal, S.; Hussain, A.; Uddin, J.; Kashif, M.; Mahmood, K. In Silico Screening, SAR and Kinetic Studies of Naturally Occurring Flavonoids Against SARS-CoV-2 Main Protease. Arab. J. Chem. 2022, 15, 103473. [Google Scholar] [CrossRef]
- Muhammad, I.; Rahman, N.; Niaz, S.; Basharat, Z.; Rastrelli, L.; Jayanthi, S.; Efferth, T.; Khan, H. Screening of Potent Phytochemical Inhibitors against SARS-CoV-2 Protease and Its Two Asian Mutants. Comput. Biol. Med. 2021, 133, 104362. [Google Scholar] [CrossRef]
- Baildya, N.; Ashik, A.; Nath, N.; Dutta, T.; Chattopadhyay, A.P. Screening of Potential Drug from Azadirachta Indica (Neem) Extracts for SARS-CoV-2: An Insight from Molecular Docking and MD-Simulation Studies. J. Mol. Struct. 2021, 1227, 129390. [Google Scholar] [CrossRef] [PubMed]
- Joshi, T.; Joshi, T.; Pundir, H.; Sharma, P.; Chandra, S. Predictive Modeling by Deep Learning, Virtual Screening and Molecular Dynamics Study of Natural Compounds against SARS-CoV-2 Main Protease. J. Biomol. Struct. Dyn. 2021, 39, 6728–6746. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, S.; El-kafrawy, S.A.; Alandijany, T.A.; Bajrai, L.H.; Shah, A.A.; Dubey, A.; Sahoo, A.K.; Yadava, U.; Kamal, M.A.; Azhar, E.I.; et al. Structure-Based Identification of Natural Products as SARS-CoV-2 Mpro Antagonist from Echinacea angustifolia Using Computational Approaches. Viruses 2021, 13, 305. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, P. In Silico Investigation of Phytoconstituents from Indian Medicinal Herb ‘Tinospora Cordifolia (Giloy)’ against SARS-CoV-2 (COVID-19) by Molecular Dynamics Approach. J. Biomol. Struct. Dyn. 2021, 39, 6792–6809. [Google Scholar] [CrossRef]
- Shree, P.; Mishra, P.; Selvaraj, C.; Singh, S.K.; Chaube, R.; Garg, N.; Tripathi, Y.B. Targeting COVID-19 (SARS-CoV-2) Main Protease through Active Phytochemicals of Ayurvedic Medicinal Plants—Withania somnifera (Ashwagandha), Tinospora cordifolia (Giloy) and Ocimum sanctum (Tulsi)—A Molecular Docking Study. J. Biomol. Struct. Dyn. 2022, 40, 190–203. [Google Scholar] [CrossRef]
- Mohammadi, M.; Rupa, F.H.; Khan, M.F.; Rashid, R.B.; Rashid, M.A. Identification of Natural Compounds with Anti-SARS-CoV-2 Activity using Machine Learning, Molecular Docking and Molecular Dynamics Simulation Studies. Dhaka Univ. J. Pharm. Sci. 2022, 21, 1–13. [Google Scholar]
- Bui, T.Q.; Thi, N.; Hai, T.; Van Chen, T.; Quy, P.T.; Nguyen, L.; Du, H. Theoretical Study on Inhibitability of Some Natural Alkaloids against Influenza Virus Hemagglutinin and SARS-CoV-2 Main Protease. Vietnam J. Chem. 2022, 60, 502–517. [Google Scholar]
- Durgam, L.; Guruprasad, L. Computational Studies on the Design of NCI Natural Products as Inhibitors to SARS-CoV-2 Main Protease. J. Biomol. Struct. Dyn. 2022, 41, 3741–3751. [Google Scholar] [CrossRef]
- El-hawary, S.S.; Issa, M.Y.; Ebrahim, H.S.; Mohammed, A.F.; Hayallah, A.M.; El-kadder, E.M.A.; Sayed, A.M. Potential of (Citrus Nobilis Lour × Citrus Deliciosa Tenora) Metabolites on COVID-19 Virus Main Protease Supported by In Silico Analysis Potential of (Citrus Nobilis Lour  Citrus Deliciosa Tenora) Metabolites on COVID-19 Virus Main Protease Supported. Nat. Prod. Res. 2022, 36, 2843–2847. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Shukla, A.; Parmar, P.; Rawal, R.M.; Patel, B.; Saraf, M.; Goswami, D. Reckoning a Fungal Metabolite, Pyranonigrin A as a Potential Main protease (Mpro) Inhibitor of Novel SARS-CoV-2 Virus Identified Using Docking and Molecular Dynamics Simulation. Biophys. Chem. 2020, 264, 106425. [Google Scholar] [CrossRef] [PubMed]
- Roshni, J.; Vaishali, R.; Ganesh, K.S.; Dharani, N.; Alzahrani, K.J.; Banjer, H.J.; Alghamdi, A.H.; Theyab, A.; Ahmed, S.S.S.J.; Patil, S. Multi-Target Potential of Indian Phytochemicals Against SARS-CoV-2 a Docking, Molecular Dynamics and MM-GBSA Approach Extended to Omicron B. 1. 1. 529. J. Infect. Public Health 2022, 15, 662–669. [Google Scholar] [CrossRef]
- Moharana, M.; Das, A.; Narayan, S.; Kumar, S.; Khan, F. Evaluation of Binding Performance of Bioactive Compounds Against Main Protease and Mutant Model Spike Receptor Binding Domain of SARS-CoV-2: Docking, ADMET Properties and Molecular Dynamics Simulation Study. J. Indian Chem. Soc. 2022, 99, 100417. [Google Scholar] [CrossRef]
- Thuy, B.T.P.; Nhan, V.D.; Quang, N.M.; Duoc, N.T.; Tat, P.V. Evaluation of SARS-CoV-2 Inhibition of Some Compounds in Cymbopogon Citratus Oil Combining Docking and Molecular Dynamics Simulations. Vietnam J. Chem. 2021, 59, 790–799. [Google Scholar]
- Muhseen, Z.T.; Hameed, A.R.; Al-Hasani, H.M.; Ahmad, S.; Li, G. Computational Determination of Potential Multiprotein Targeting Natural Compounds for Rational Drug Design Against SARS-CoV-2. Molecules 2021, 26, 674. [Google Scholar] [CrossRef]
- Goyzueta-Mamani, L.D.; Barazorda-Ccahuana, H.L.; Mena-Ulecia, K.; Chávez-Fumagalli, M.A. Antiviral Activity of Metabolites from Peruvian Plants Against SARS-CoV-2: An In-Silico Approach. Molecules 2021, 26, 3882. [Google Scholar] [CrossRef]
- Africa, J.G.G.; Arturo, H.C.P.; Bernardo, L.J.M.; Kyle, J.; Ching, A.R.; Casandra, O.; De Cruz, E.; Hernandez, J.B.E.; Magsipoc, R.J.Y.; Sales, C.T.C.; et al. In Silico Triple Targeting of SARS-CoV-2 3CL Pro, PL pro, and RdRp by Philippine Antitubercular Natural Products Libraries. Philipp. J. Sci. 2022, 151, 35–58. [Google Scholar]
- Hang, T.T.T.; Khanh, D.T.H.; Tung, B.T. In Silico Screening of Natural Antivirals as Potential Inhibitors of SARS-CoV-2 Virus. Vietnam J. Chem. 2022, 60, 211–222. [Google Scholar]
- Shaldam, M.A.; Yahya, G.; Mohamed, N.H.; Abdel-daim, M.M.; Naggar, Y. In Silico Screening of Potent Bioactive Compounds from Honeybee Products Against COVID-19 Target Enzymes. Environ. Sci. Pollut. Res. 2021, 28, 40507–40514. [Google Scholar] [CrossRef]
- Melinda, Y.N.; Widada, J.; Dwi, T.; Febriansah, R.; Damayanti, E.; Mustofa, M. Heliyon Metabologenomics Approach to the Discovery of Novel Compounds from Streptomyces sp. GMR22 as Anti-SARS-CoV-2 Drugs. Heliyon 2021, 7, 08308. [Google Scholar] [CrossRef]
- Dutta, M.; Nezam, M.; Chowdhury, S.; Rakib, A.; Paul, A. Appraisals of the Bangladeshi Medicinal Plant Calotropis Gigantea Used by Folk Medicine Practitioners in the Management of COVID-19: A Biochemical and Computational Approach. Front. Mol. Biosci. 2021, 8, 625391. [Google Scholar] [CrossRef]
- Kumar, M.; Ghosh, S.K.; Das, A. Natural Product Compounds in Alpinia Officinarum and Ginger are Potent SARS-CoV-2 Papain-like Protease Inhibitors. ChemRxiv 2020, 1, 12071997. [Google Scholar]
- Suleimen, Y.M.; Jose, R.A.; Suleimen, R.N.; Arenz, C.; Ishmuratova, M.; Toppet, S.; Dehaen, W.; Alsfouk, A.A.; Elkaeed, E.B.; Eissa, I.H. Isolation and In Silico Anti-SARS-CoV-2 Papain-Like Protease Potentialities of Two Rare 2-Phenoxychromone Derivatives from Artemisia spp. Molecules 2022, 27, 1216. [Google Scholar] [CrossRef] [PubMed]
- Jose, S.; Gupta, M.; Sharma, U.; Quintero-Saumeth, J.; Dwivedi, M. Potential of Phytocompounds from Brassica Oleracea Targeting S2-Domain of SARS-CoV-2 Spike Glycoproteins: Structural and Molecular Insights. J. Mol. Struct. 2022, 1254, 132369. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Karmakar, A.; Mallick, T.; Begum, N.A. Exploring the Efficacy of Naturally Occurring Biflavone Based Antioxidants towards the Inhibition of the SARS-CoV-2 Spike Glycoprotein Mediated Membrane Fusion. Virology 2021, 556, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Chung, H.S.; Noh, S.G.; Lee, B.; Chung, H.Y.; Choi, J.G. Geraniin Inhibits the Entry of SARS-CoV-2 by Blocking the Interaction between Spike Protein RBD and Human ACE2 Receptor. Int. J. Mol. Sci. 2021, 22, 8604. [Google Scholar] [CrossRef] [PubMed]
- Kar, P.; Saleh-E-In, M.M.; Jaishee, N.; Anandraj, A.; Kormuth, E.; Vellingiri, B.; Angione, C.; Rahman, P.K.S.M.; Pillay, S.; Sen, A.; et al. Computational Profiling of Natural Compounds as Promising Inhibitors against the Spike Proteins of SARS-CoV-2 Wild-Type and the Variants of Concern, Viral Cell-Entry Process, and Cytokine Storm in COVID-19. J. Cell. Biochem. 2022, 123, 964–986. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, A.; Biswas, S.; Hajra, S.; Saha, P. In Silico Validation of Potent Phytochemical Orientin as Inhibitor of SARS-CoV-2 Spike and Host Cell Receptor GRP78 Binding. Heliyon 2021, 7, 05923. [Google Scholar] [CrossRef]
- Kiran, G.; Karthik, L.; Devi, M.S.S.; Sathiyarajeswaran, P.; Kanakavalli, K.; Kumar, K.M.; Kumar, D.R. In Silico Computational Screening of KabasuraKudineer-of Fi Cial Siddha Formulation and JACOM against SARS-CoV-2 Spike Protein. J. Ayurveda Integr. Med. 2022, 13, 100324. [Google Scholar] [CrossRef]
- Mhatre, S.; Gurav, N.; Shah, M.; Patravale, V. Entry-Inhibitory Role of Catechins Against SARS-CoV-2 and Its UK Variant. Comput. Biol. Med. 2021, 135, 104560. [Google Scholar] [CrossRef]
- Shadrack, D.M.; Vuai, S.A.H.; Sahini, M.G.; Onoka, I. In Silico Study of the Inhibition of SARS-CoV-2 Viral Cell Entry by Neem Tree Extracts. RSC Adv. 2021, 11, 26524–26533. [Google Scholar] [CrossRef] [PubMed]
- Cheke, R.S.; Jain, G.; Narkhede, R.; Shinde, S.D.; Ambhore, J.P. Natural Product Emerging as Potential SARS Spike Glycoproteins-ACE2 Inhibitors to Combat COVID-19 Attributed by In-Silico Investigations. Biointerface Res. Appl. Chem. 2021, 11, 10628–10639. [Google Scholar]
- Prasad, S.K.; Pradeep, S.; Shimavallu, C.; Kollur, S.P.; Syed, A.; Marraiki, N.; Egbuna, C.; Gaman, M.; Kosakowska, O.; Cho, W.C.; et al. Evaluation of Annona Muricata Acetogenins as Potential Anti-SARS-CoV-2 Agents through Computational Approaches. Front. Chem. 2021, 8, 1281. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, J.; Li, Y.; Wang, Z.; Chu, M.; Wang, Y. Tuftsin: A Natural Molecule Against SARS-CoV-2 Infection. Front. Mol. Biosci. 2022, 9, 859162. [Google Scholar] [CrossRef] [PubMed]
- Tallei, T.E.; Yelnetty, A.; Idroes, R.; Kusumawaty, D. An Analysis Based on Molecular Docking and Molecular Dynamics Simulation Study of Bromelain as Anti-SARS-CoV-2 Variants. Front. Pharmacol. 2021, 12, 2192. [Google Scholar] [CrossRef] [PubMed]
- Dharmashekara, C.; Pradeep, S.; Prasad, S.K.; Jain, A.S.; Syed, A.; Shiva, K.; Patil, S.S.; Beelagi, M.S.; Srinivasa, C.; Shivamallu, C. Virtual Screening of Potential Phyto-Candidates as Therapeutic Leads against SARS-CoV-2 Infection. Environ. Chall. 2021, 4, 100136. [Google Scholar] [CrossRef]
- Zhang, Y.; Yao, Y.; Yang, Y.; Wu, H. Investigation of Anti-SARS, MERS, and COVID-19 Effect of Jinhua Qinggan Granules Based on a Network Pharmacology and Molecular Docking Approach. Nat. Prod. Commun. 2021, 16, 1–9. [Google Scholar] [CrossRef]
- Kashyap, D.; Jakhmola, S.; Tiwari, D.; Kumar, R.; Moorthy, N.H.N.; Elangovan, M.; Brás, N.F.; Jha, H.C. Plant Derived Active Compounds as Potential Anti SARS-CoV-2 Agents: An In-Silico Study. J. Biomol. Struct. Dyn. 2021, 40, 10629–10650. [Google Scholar] [CrossRef]
- Siddiqui, S.; Ahmad, R.; Alaidarous, M.; Zia, Q.; Ahmad, S.; Bader, M.; Aditi, A.; Anchal, S. Phytoconstituents from Moringa Oleifera Fruits Target ACE2 and Open Spike Glycoprotein to Combat SARS-2: An Integrative Phytochemical and Computational Approach. J. Food Biochem. 2022, 46, 14062. [Google Scholar] [CrossRef] [PubMed]
- Mir, S.A.; Firoz, A.; Alaidarous, M.; Alshehri, B.; Bin Dukhyil, A.A.; Banawas, S.; Alsagaby, S.A.; Alturaiki, W.; Bhat, G.A.; Kashoo, F.; et al. Identification of SARS-CoV-2 RNA-Dependent RNA Polymerase Inhibitors from the Major Phytochemicals of Nigella Sativa: An In Silico Approach. Saudi J. Biol. Sci. 2022, 29, 394–401. [Google Scholar] [CrossRef]
- Husain, I.; Ahmad, R.; Siddiqui, S.; Chandra, A.; Misra, A.; Srivastava, A.; Ahamad, T.; Faheem, M.; Siddiqi, Z.; Trivedi, A.; et al. Structural interactions of phytoconstituent(s) from cinnamon, bay leaf, oregano, and parsley with SARS-CoV-2 nucleocapsid protein: A comparative assessment for development of potential antiviral nutraceuticals. J. Food Biochem. 2022, 46, 14262. [Google Scholar] [CrossRef]
- Suravajhala, R.; Parashar, A.; Choudhir, G.; Kumar, A.; Malik, B. Molecular Docking and Dynamics Studies of Curcumin with COVID-19 Proteins. Netw. Model. Anal. Health Inform. Bioinform. 2021, 10, 44. [Google Scholar] [CrossRef]
- Muhseen, Z.T.; Hameed, A.R.; Al-Hasani, H.M.H.; Tahir ul Qamar, M.; Li, G. Promising Terpenes as SARS-CoV-2 Spike Receptor-Binding Domain (RBD) Attachment Inhibitors to the Human ACE2 Receptor: Integrated Computational Approach. J. Mol. Liq. 2020, 320, 114493. [Google Scholar] [CrossRef]
- Hu, L.C.; Ding, C.H.; Li, H.Y.; Li, Z.Z.; Chen, Y.; Li, L.P.; Li, W.Z.; Liu, W.S. Identification of Potential Target Endoribonuclease NSP15 Inhibitors of SARS-CoV-2 from Natural Products through High-Throughput Virtual Screening and Molecular Dynamics Simulation. J. Food Biochem. 2022, 46, 14085. [Google Scholar] [CrossRef]
- Naik, B.; Gupta, N.; Ojha, R.; Singh, S.; Prajapati, V.K.; Prusty, D. High Throughput Virtual Screening Reveals SARS-CoV-2 Multi-Target Binding Natural Compounds to Lead Instant Therapy for COVID-19 Treatment. Int. J. Biol. Macromol. 2020, 160, 1–17. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Abiodun, O.A.; Ogboo, C.; Kola-mustapha, A.T.; Ifeanyi, E.; Edemhanria, L.; Kumari, A.; Jaganathan, R.; Niyi, S. Polypharmacology of Some Medicinal Plant Metabolites Against SARS-CoV-2 and Host Targets: Molecular Dynamics Evaluation of NSP9 RNA Binding Protein. J. Biomol. Struct. Dyn. 2021, 40, 11467–11483. [Google Scholar] [CrossRef]
- Khan, M.T.; Ali, A.; Wang, Q.; Irfan, M.; Khan, A.; Zeb, M.T.; Zhang, Y.; Chinnasamy, S.; Wei, D. Marine natural compounds as potent inhibitors against the main protease of SARS-CoV-2. A molecular dynamic study. J. Biomol. Struct. Dyn. 2020, 39, 3627–3637. [Google Scholar] [CrossRef] [PubMed]
- Sigman, D.S.; Mooser, G. Chemical Studies of Enzyme Active Sites. Annu. Rev. Biochem. 1975, 40, 889–931. [Google Scholar] [CrossRef] [PubMed]
- Parida, P.K.; Paul, D.; Chakravorty, D. Nature’s Therapy for COVID-19: Targeting the Vital Non-Structural Proteins (NSP) from SARS-CoV-2 with Phytochemicals from Indian Medicinal Plants. Phytomedicine Plus 2021, 1, 100002. [Google Scholar] [CrossRef] [PubMed]





















| Serial No. | Compound Name | Binding Potential | Interactions at the Enzyme Active Site * | References | ||
|---|---|---|---|---|---|---|
| Docking Score | Binding Free Energy | H-Bond Interactions | Hydrophobic and Other Interactions | |||
| Major Protease (Mpro/3CLpro) | ||||||
| 1 | Forsythiaside A (46) | −8.08 kcal/mol | −121 ± 19 kJ/mol | THR190, PRO168 | HIS41, ASP187 | [64] |
| 2 | Pycnanthuquinone C (47) | - | −7.8 kcal/mol | HIS41 | GLU166, HIS164, CYS145, LEU27 | [65] |
| 3 | Pycnanthuquinone B (48) | - | −8.3 kcal/mol | GLY143, SER144 | HIS41, HIS164 | [65] |
| 4 | Cyanopyrin (49) | −4.78 kcal/mol | −7.4 kcal/mol | THR111, ASP153, SER158 | PHE294, ILE106 | [66] |
| 5 | Withanolide R (50) | −9.63 kcal/mol | −141.96 kJ/mol | GLN189, HIS41 | MET165, PRO168, ASP187, ALA191 | [67] |
| 6 | Hesperidin (51) | −178.5910 kJ/mol | - | GLU166, GLN192, ARG 188, GLN189, MET49, ASP187, TYR54, LEU141, SER144, HIS163, THR26 | - | [68] |
| 7 | Rutin (29) | −176.2740 kJ/mol −9.09 kcal/mol | - - | CYS145, ASN142, PHE140, GLU166, GLN192, THR190, ASP187, TYR54, HIS164, GLU166, LEU8 | - - | [68] [69] |
| 8 | Diosmin (52) | −174.126 kJ/mol | - | GLN192, THR190, ARG188, HIS164, GLN189, GLU166, GLY143, SER144, CYS145 | - | [68] |
| 9 | Apiin (53) | −171.008 kJ/mol | - | THR190, SER144, LEU141, HIS163, CYS145, THR26 | - | [68] |
| 10 | Jaceidin (54) | −7.3 kcal/mol | - | LEU141, GLY143, SER144, CYS145, ARG188, | - | [70] |
| 11 | Pachypodol (55) | −7.1 kcal/mol | - | GLY143, SER144, CYS145 | - | [70] |
| 12 | Chrysosplenetin (56) | −7.1 kcal/mol | - | LEU141, GLY143, SER144, CYS145 | - | [70] |
| 13 | Imidazoline-4-one-2-imino-1-(4-methoxy-6-dimethylamino-1,3,5-triazin-2-yl) (57) | −7.013 kcal/mol | - | GLU166, GLN192 | - | [71] |
| 14 | Spiro[4,5]dec-6-en-1-ol, 2,6,10,10-tetramethyl (58) | −6.369 kcal/mol | - | - | MET165, HIS41, MET49, CYS145 | [71] |
| 15 | 3-hydroxy-5-cholen-24-oic acid (59) | −6.251 kcal/mol | - | THR25, THR190, GLN192 | MET49, HIS41, MET165 | [71] |
| 16 | Withacoagulin H (60) | −11.1 kcal/mol | −63.463 kJ/mol | GLY143, ARG188, THR190, GLN192 | HIS41, CYS145, THR25, THR26, LEU27, MET49, ASN142, SER144, HIS164, MET165, GLN166, GLN189 | [72] |
| 17 | Ajugin E (61) | −11.5 kcal/mol | −56.14 kJ/mol | GLN192, THR190 | HIS41, CYS145, THR25, MET49, ASN142, GLY143, HIS164, MET165, GLU166, PRO168, ARG188, GLN189 | [72] |
| 18 | Withacoagulin (62) | −10.8 kcal/mol | −44.496 kJ/mol | THR190, GLN192, GLY143 | HIS41, CYS145, THR25, THR26, LEU27, MET49, ASN142, SER144, HIS164, MET165, ARG188, GLN189 | [72] |
| 19 | Crocin (63) | −8.0 kcal/mol | - | ALA116, SER123, SER139, PHE140, LEU141, ASN142, GLU166, HIS172, GLN189 | [73] | |
| 20 | Rhamnocitrin (64) | −7.83 kcal/mol | −49.53 kcal/mol | GLU166, THR190 | HIS41, CYS145, CYS44, TYR54, PRO52, MET49, ASP187, ARG188, GLN189, ALA191, GLN192, PRO168, LEU167, MET165, HIS164 | [74] |
| 21 | Sterenin M (65) | −8.431 kcal/mol | −49.57 kcal/mol | GLU166, PHE140, HIS163, GLY143, ASN142, THR26, HIS41 | LEU141, HIS172, SER144, LEU27, THR25, THR24, CYS44, TYR54, ASP187, ARG188, MET49, GLN189, HIS164, MET165 | [75] |
| 22 | 3,4,5-tricaffeoylquinic acid (66) | −9.0 kcal/mol | - | THR190, ARG188, THR26, ASP187, GLY143, ASN142 | MET165, MET49, PRO168, HIS172 | [76] |
| 23 | Quercetin (40) | −9.2 kcal/mol | - | GLU290, GLU288, THR199 | LYS137, ARG131, LEU272, TYR239, LEU287 | [77] |
| 24 | Peonidin (67) | −8.4 kcal/mol | - | GLU290, ASP289, LYS5GLU288 LEU287, LEU271, GLY27, THR199, LEU272, GLU290 | [77] | |
| 25 | Catechin (41) | −7.67 kcal/mol | - | GLU166, HIE164 | - | [69] |
| 26 | Kaempferol (23) | −7.215 kcal/mol | - | GLU166, GLN189, HIE164 | - | [69] |
| 27 | Amentoflavone (42) | −9.8 kcal/mol −8.6 kcal/mol | - - | THR25, GLY143, GLN189, GLU166 CYS44, VAL186, ARG188, GLU166 | MET49, HIS41, CYS145, THR24, THR45, MET165, TYR54, ARG188, ASP187, LEU167, PRO168, ASN142 THR25, HIS41, ASN142, CYS145 | [78] [79] |
| 28 | Ferolide (68) | −7.9 kcal/mol | - | HIS163, CYS145, LEU141, ASN142 | CYS145, HIS41, GLU166, SER144, PHE140, HIS164, MET165, GLN189, CYS144, MET49, THR25, GLY143 | [80] |
| 29 | Ginkgolide A (69) | −65.412 kcal/mol | −63 kcal/mol | VAL72, LYS73, TYR135, GLY151, CYS144, HIS41 | [81] | |
| 30 | Gracillin (70) | −9.2 kcal/mol | - | GLU166, LEU141, SER144, CYS145, GLY143, ASN142 | - | [82] |
| 31 | Proanthocyanidin (71) | −9.2 kcal/mol | - | GLU166, HIS164, HIS163, TYR54 | HIS41, ASP187, MET165 | [82] |
| 32 | Ginkgolide M (72) | −11.2 kcal/mol | - | ASN142, CYS145, GLU166, GLY143, HIS163, PHE140 | [83] | |
| 33 | Mezerein (73) | −11 kcal/mol | - | ASN142, CYS145, GLU166, GLY143, HIS163, HIS172, PHE140 | [83] | |
| 34 | Tubocuraine (74) | −10.9 kcal/mol | - | ASN142, CYS145, GLU166, GLY14 | [83] | |
| 35 | Choline (75) | −3.7 kcal/mol | - | GLY143, LEU141 | ASN142, PHE140, HIS163, SER144, GLU166 | [84] |
| 36 | Volkensiflavone (76) | −8.6 kcal/mol | - | CYS145, GLU166 | THR25, HIS41 | [79] |
| 37 | Stigmasterol (77) | −6.30 kcal/mol | - | PRO39 | CYS145, LEU27, VAL42, HIS41 | [85] |
| 38 | Hesperidin (51) | −8.3 kcal/mol | - | PHE140, GLU166, SER144, CYS145 | MET49, LEU141, HIS163, THR26 | [86] |
| 39 | Amphimedoside C (78) | - | −127.3 kJ/mol | ASN142, PHE140, LEU141, HIS163, SER144, CYS145 | ASN142, HIS172, GLN189 | [87] |
| 40 | Fasciospongide A (79) | - | −104.37 kJ/mol | HIS41, THR190, GLN192, VAL186 | MET165, MET49, PRO168, ALA193, LEU167, PHE185, ARG188, VAL42, THR25, CYS145 | [88] |
| 41 | Glaucogenin D (81) | −7.9 kcal/mol | - | GLY143, HIS41 | MET165, THR24 | [89] |
| 42 | Calcium elenolate (83) | −7.0 kcal/mol | - | GLU291 | LYS152, PHE111 | [90] |
| 43 | Maackin A (164) | −8.4 kcal/mol | −43.1 kcal/mol | THR26, HIS41, MET49, GLN192 | THR25, GLY143, ASN142, MET165, PRO168 | [91] |
| 44 | Anthracene dione (165) | −5.73 kcal/mol | - | GLY143 | - | [92] |
| 45 | Fortunellin (166) | −13.9 kcal/mol | - | LEU32, ASP33, ASP34, VAL35, TYR37, GLN83, LYS88, TYR101, LYS102, PHE103, VAL104, ARG105, ASP108, PHE159, CYS160, ASP176, LEU177, GLU178 | [93] | |
| 46 | Curcumin (86) | −8.62 kcal/mol | - | THR26, GLY143, GLN189, THR190 | - | [94] |
| 47 | Kolaviron (87) | −7.027 kcal/mol | - | GLU166, GLY143 | - | |
| 48 | Bisdemethoxycurcumin (88) | −5.641 kcal/mol | - | THR26, THR190, GLN189, GLU166 | - | [94] |
| 49 | 6-gingerol (89) | −4.975 kcal/mol | - | THR190, GLN189 | - | [94] |
| 50 | Artemisinin (26) | −4.252 kcal/mol | - | GLN 189, GLU 166 | - | [94] |
| 51 | Luteolin (90) | −8.3 kcal/mol | - | GLN110, THR111 ASN151 | - | [95] |
| 52 | Lucenin (91) | −10.7 kcal/mol | - | PHE219, LEU220, ARG222, ASN274 | - | [95] |
| 53 | Olealonic acid (92) | −9.5 kcal/mol | - | ASP289 | - | [95] |
| 54 | Isoorientin (93) | −9.2 kcal/mol | - | PRO52, TYR54, PHE181 | - | [95] |
| 55 | Isochaphoside (94) | −10.5 kcal/mol | - | ARG222, ASN274, PHE219 | - | [95] |
| 56 | Saponarin (95) | −10.6 kcal/mol | - | ARG40, PHE181 | - | [95] |
| 57 | Schaftoside (96) | −10.2 kcal/mol | - | ASP197, ASN238, ARG131, LYS137 | - | [95] |
| 58 | 7,2″-bieckol (98) | −10.7855 kcal/mol | - | THR24, THR26, GLY143, GLU189 | - | [96] |
| 59 | Sarelengan B (105) | −9.8 kcal/mol | - | HIS41, CYS145, GLU166 | - | [97] |
| 60 | Bislatumlide A (106) | −9.6 kcal/mol | −34.8 kcal/mol | GLY143, GLU166 | - | [97] |
| 61 | Eucalyptol (107) | −5.86 kcal/mol | - | - | MET49, MET165, HIS164, ARG188, HIS41, PRO52, ASP187, GLN189, TYR54, PHE18 | [98] |
| 62 | Nictoflorin (114) | −9.18 kcal/mol | THR190, GLY143 | - | [99] | |
| 63 | Aloenin (115) | −9.13 kcal/mol | PHE140 | - | [99] | |
| 64 | Andrographolide (116) | −7.06 kcal/mol | - | PHE140, SER144 | - | [100] |
| 65 | Galangin (117) | −8.066 kcal/mol | - | CYS145, PHE140, HIS164, HIS41 | HIS41, MET165, MET49, GLU166 | [101] |
| 65 | Amentoflavone (42) | −9.9 kcal/mol | - | HIS163, THR26, GLU166 | GLN189, ARG188, ASP187, TYR54, MET49, HIS164, HIS172, PHE140, SER144, LEU141, GLY143, LEU27, THR25, THR26, ASN142 | [102] |
| 67 | Kazinol T (123) | −14.355008 kcal/mol | - | HIS41 | GLY143, THR190, GLY143, HIS42, CYS145, GLY143, HIS164 | [103] |
| 68 | Desacetyl gedunin (125) | −7.3 kcal/mol | - | TYR 207, SER245, MET206, ARG166, VAL202, SER170, GLU203, LEU199, THR197, LEUA85, LYS232, MET208 | [104] | |
| 69 | Palmatine (126) | −8.7 kcal/mol | −71.47 kJ/mol | GLN189 | ASN142, GLY143, THR25, LEU27, ARG188,GLN192, PRO168, CYS145, MET165, GLN189, HIS41 | [105] |
| 70 | Sauchinone (127) | −8.9 kcal/mol | −71.68 kJ/mol | MET49, HIS41 | ARG188, ASP187, PRO52, CYS44, HIS41, THR25, LEU141, GLU166, GLN189, CYS145, MET165 | [105] |
| 71 | Quercetagetin 7-glucoside (130) | −15.20 kcal/mol | - | CYS44, LEU141, CYS145, GLU166, GLN189, HIS41, CYS44 | LEU27, CYS44, MET49, PHE140, LEU141, CYS145, MET165, LEU167, PRO168 | [106] |
| 72 | Berberine (134) | −7.3 kcal/mol | - | THR25, SER46 | HIS163 | [107] |
| 73 | Withanoside V (43) | −10.32 kcal/mol | - | ASN84, ARG40, MET 82 | CYS85, ARG105, PHE134 | [108] |
| 74 | Lignoid (140) | −9.0 kcal/mol | - | THR25, HIS41, LEU141, SER144, CYS145, HIS163, GLN189 | LEU27, MET49, HIS163, MET165, ASP187, GLN189 | [109] |
| 75 | Lycorine (141) | −11.9 kcal/mol | - | GLU166, HIS41 | ASP188, GLN192, MET 165, GLN 189, HIS 164, HIS163, PHE 140, LEU 141, CYS 146, SER 144, ARG 188 | [110] |
| 76 | Hemanthamine (142) | −11.4 kcal/mol | - | MET165, HIS163, HIS41 | CYS145, PHE140, ASN142, SER144, LEU141, ARG188, HIS164, GLU166, ASP187, GLN189 | [110] |
| 77 | NSC36398 (143) | −8.1 kcal/mol | - | PHE140, LEU141, SER144, MET165, GLU166, GLN189 | LEU27, HIS41, LEU50, PHE140, LEU141, GLY143, SER144, CYS145, HIS163, HIS164 MET165, GLU166, LEU167, PRO168, GLN189 | [111] |
| 78 | Quercetin 7-O-glucoside-3-O-rutinoside (147) | −9.47 kcal/mol | - | PRO168, GLU166, CYS145, MET165, MET49, GLN189 | MET165, GLU166, GLN189, ASN142, MET49, PRO168, HIS41 | [112] |
| 79 | Pyranonigrin A (151) | −7.3 kcal/mol | - | LEU141, GLY143, SER144, CYS145, HIS163, GLU166 GLN189, ASN142 | MET165 | [113] |
| 80 | Arboreol (153) | −8.2 kcal/mol | - | - | - | [114] |
| 81 | Acacetin (154) | −7.77 kcal/mol | - | MET49, TYR54, CYS145 | MET49 | [115] |
| 82 | Epoxy-linalool oxide (155) | −12.80 kcal/mol | - | LEU141, CYS145, GLY143 | MET165, ASN142, HIS163, HIS41, GLU166, HIS164, SER144, LEU27, THR25, MET49, THR26 | [116] |
| 83 | Glycyrrhizin (156) | −8.7 kcal/mol | - | ASP289, ASN238 | TYR239, LYS137, ASP197, THR198, GLU290, SER284, TYR237, LEU287, LEU286, LEU272, GLY275, LEU271, GLN273, ASN274, MET276, TYR239 | [117] |
| 84 | Rutin (29) | - | −40.293 kcal/mol | ASN142, MET49, HIS41, ASP187 | [118] | |
| 85 | Vobtusine lactone (157) | −8.3 kcal/mol | - | ASN119, SER46, THR25 | CYS44, HIS41, MET49 | [119] |
| 86 | Sotetsuflavone (159) | −9.6 kcal/mol | - | GLU166, GLN189, THR25 | PRO168, LEU167, GLY170, MET165, ASP187, ARG188, TYR54, THR45, THR24, ASN142 | [120] |
| 87 | Kaempferol (23) | −7.8 kcal/mol | - | SER144, LEU141, ASP187, TYR54, GLN189 | HIS163, HIS164, CYS145, GLU166, MET165, MET49, HIS41, ARG188 | [121] |
| 88 | Echoside A (168) | −8.4 kcal/mol | - | THR25, HIS41, CYS145, GLU 166 | MET165, MET49 | [122] |
| 89 | Echoside B (169) | −9.4 kcal/mol | - | ARG4, SER 284, and LYS5 | LYS5, GLU288, LEU282 | [122] |
| 90 | Juniper camphor (172) | −6.06 kcal/mol | - | MET165 | MET49, MET165, PRO168, ASP147 | [123] |
| Papain-Like Protease | ||||||
| 91 | Amentoflavone (42) | −10.8 kcal/mol | - | HIS342, LYS711, ARG712 | LYS711, ASP339, ARG558, ILE310, ILE580, ALA579, LEU742 | [79] |
| 92 | Jezonofol (167) | −9.0 kcal/mol | −60.7 kcal/mol | ARG166, GLU16, GLN174, TYR264 | SER170. GLY163, MET208, GLN203, LYS157, VAL202, MET206,LEU199, TYR207, LEU185, GLU161, LEU162, GLN269, TYR268 | [91] |
| 93 | Constanolactone B (80) | - | −92.57 kJ/mol | TYR268, ASP164 | ASN109, TYR11, GLY271, LEY162, CYS270, GLY163, GLN269, THR301, ARG166, PRO248, MET208, PRO247, ALA246, SER245 | [88] |
| 94 | Glaucogenin D (81) | −6.4 kcal/mol | - | HIS272 | TRP106, HIS272 | [89] |
| 95 | Glaucogenin A (82) | −6.4 kcal/mol | - | HIS272, ASP286 | TRP106, HIS272, TRP270 | [89] |
| 96 | (E)-7-(4-hydroxy-3-methoxyphenyl)-1-phenylhept-4-en-3-one (84) | - | −47 kJ/mol (closed conformation) −28 kJ/mol (open conformation) | ARG166, ASP164, GLN269, TYR264, LEU163, GLY163, MET208 - | [124] | |
| 97 | 8-gingerol (85) | −43 kJ/mol (closed conformation) −15 kJ/mol (open conformation) | ARG166, ASP164, TYR268, GLN269, TYR264, PRO247, GLY163, PRO248 - | [124] | ||
| 98 | 4,8-dihydroxysesamin (152) | −10.3 kcal/mol | - | - | - | [114] |
| 99 | Glycyrrhizin (156) | −7.9 kcal/mol | - | ASN128, ASP179, GLN174 | ASP76, ASN156, ARG82, THR74, PHE79, TYR154, HIS175, HIS73, PHE69, VAL202, PHE173, LEU178, ALA176, ASN177 | [117] |
| 100 | Rutin (29) | - | −21.713 kcal/mol | TYR268, GLN269 | [118] | |
| 101 | Deoxyvobtusine lactone (158) | −10.8 kcal/mol | - | ARG558, ARG712 | ILE310, PHE735, LEU742, ASP339 THR583 | [119] |
| 102 | 6-demethoxy-4′-O-capillarsine (170) | −18.86 kcal/mol | - | GLN270, PRO249 | TYR269, ASP165, PRO248, PRO249 | [125] |
| 103 | Tenuflorin C (171) | −18.37 kcal/mol | - | GLN270, TYR274, ALA247, LEU163 | TYR269, ASP165, PRO248, ASP165, MET209 | [125] |
| Spike Proteins | ||||||
| 104 | Terpene NPACT01552 (173) | −11 kcal/mol | - | GLY496, TYR453, GLN493, SER494, GLU484 | LYS417, LEU452, TYR489, PHE456, LEU455 | [126] |
| 105 | Hinokiflavone (175) | −9.60 kcal/mol | - | GLN1201, GLN926, GLU1195, LEU1197, ASN928, GLU918, ASN919 | GLU1195, LEU1197, GLU918 | [127] |
| 106 | Robustaflavone (176) | −9.40 kcal/mol | - | ASP936, GLN926, LYS1191, GLU1195, ASN928 | GLU1195, ASN928 | [127] |
| 107 | Abietatriene (178) | −9.8 ± 0.02 kcal/mol | - | - | TYR453, TYR495, ASN501, TYR505 | [128] |
| 108 | Quercetin (40) | −7.8 kcal/mol | - | ASN501, TYR505, GLY496 | GLN493, LYS417, GLU406, LEU455, TYR495, PHE497, GLN506 | [77] |
| 109 | Hesperidin (51) | - | −10.4 kcal/mol | ASN1023, SER1030, LHR1027 | ARG1039, ALA1026, LEU1024, PHE1042, ARG1039, THR1027, ARG1039, LEU1024, THR1027, ALA1020, ASN1023, GLN784, GLU780 | [86] |
| 110 | Nabiximols (179) | - | −10.2 kcal/mol | GLN762, LYS776, GLU1017 | ARG765, ARG1014, ALA958, ALA766, LEU1012, GLY769 | [86] |
| 111 | Amentoflavone (42) | −9.9 kcal/mol | - | LEU861, LYS733, SER730, THR732 | ALA956, PRO862, HIS1058, ASP867, VAL860, PRO1057, MET731, VAL952, ASN955, PHE823 | [78] |
| 112 | Crocin (63) | −7.6 kcal/mol | - | ARG346, PHE347, SER349, TYR351, LEU441, LYS444, VAL445, GLY447, ASN448, TYR449, ASN450, PHE490 | [73] | |
| 113 | Pelargonidin-3-galactoside (180) | −8.6 kcal/mol | - | GLN166, PHE83 | SER80, LYS103, PRO40, ASP165 | [90] |
| 114 | Orientin (181) | −6.2 kcal/mol | - | LYS333, ASP429, THR431, ASN435, ASN437, TYR438, ARG495 | ASN437 | [129] |
| 115 | Chrysoeriol (182) | −11.478 kcal/mol | - | CYS336, GLY339, ASP364 | PHE338, PHE342, PHE374, LEU335, VAL367, SER373 | [130] |
| 116 | Luteolin (183) | −11.392 kcal/mol | - | ASP364, VAL367, SER371, SER373, CYS336, VAL362 | PHE338, GLY339, PHE374, PHE342 | [130] |
| 117 | Acacetin (154) | −7.75 kcal/mol | - | THR26, THR24, HIS164, GLU166, LEU141, SER144, CYS145, GLT143 | HIS41 | [115] |
| 118 | Epigallocatechin gallate (21) | −6.3 kcal/mol | - | TYR1110, PHE1109, GLN1071, GLU918, THR716 | - | [131] |
| 119 | Geraniin (177) | −8.1 kcal/mol | - | ARG403, TYR449, TYR453, GLN493, SER494, GLN498 | TYR495, GLY496 | [132] |
| 120 | cis-Annonacin (192) | −7.7 kcal/mol | - | PHE390, ARG393, GLN409, GLY496, TYR505, ARG403, TYR453 | ASN33, GLU37, PRO389, ARG403, GLU406, LYS417, TYR495, PHE497 | [133] |
| 121 | Trigoneoside IB (195) | −8.5 kcal/mol | - | THR260, ALA618, SER289, LYS946, SER279, THR261, THR616, GLY288, THR302, THE285, CYS288 | - | [134] |
| 122 | Arctiin (196) | 137.043 (LiDock score) | - | SER142 | THR193, ALA146, GLY144, SER196, GLY143, PRO195, PHE118, THR141, VAL194 | [135] |
| 123 | Linarin (197) | 162.676 (LiDock score) | - | GLY42, VAL105, GLN168 | ARG44, GLY103, GLN102, TYR88, GLN43, GLN39, ALA85, GLU107, PRO41, SER170, GLU167, THR104, GLY103 | [135] |
| 124 | Withanolide D (198) | −9.8 kcal/mol | - | GLN954 | ASP950, VAL951, LYS947, PRO728, GLU1017, GLU773, ILE770, LEU1012, ILE1013, GLN1010, AGR1014, ALA766, GLY769 | [136] |
| 125 | Morellic acid (199) | −10.3 kcal/mol | - | THR549, THR547 | MET740, GLY744, ASN856, PHE855, PHE541, ILE587, THR573, LEU546, ASP571, LEU977, ARG1000, SER975, ASN978, ASN540, ASP745, GLY548 | [120] |
| 126 | Echoside A (168) | −7.9 kcal/mol | - | GLU1195, LEU1200, GLN1201, ASP1199 | LEU1197 | [122] |
| 127 | Echoside B (169) | −7.8 kcal/mol | - | SER943, ASP936, ARG1185 | LYS1191, ALA1190 | [122] |
| 128 | Pyrrolidinone (200) | −5.97 kcal/mol | - | SER730 | THR778, PHE782, VAL729, ILE870, ALA1056, AND GLY1059 | [137] |
| 129 | Scutellarein (118) | −8.9 kcal/mol | - | - | - | [114] |
| (a) RBD-ACE2 | ||||||
| 130 | 3-p-coumaroylquinic acid (174) | −8.9 kcal/mol | −6.71 kcal/mol | THR1006, GLN1005 | ALA766, LEU763, VAL1008, GLN1010, GLN1002, THR1009, THR1006 | [138] |
| 131 | Rutin (29) | −7.601 kcal/mol | - | ASN388, ASP389, ALA363, CYS361, SER359, ILE332, ASN331 | - | [69] |
| 132 | Catechin (41) | −6.470 kcal/mol | - | SER359, ASN331, CYS 361, ILE332 | - | [69] |
| 133 | Kaempferol (23) | −6.743 kcal/mol | - | ILE358, ASN388 | - | [69] |
| 134 | Azadirachtin H (184) | −8.18 kcal/mol | - | ARG393, ARG408, LYS417, GLN409, ASP30, ASN33, HIS34, LYS31, ASP30, GLU406, LYS455, ARG403 | [139] | |
| 135 | Indigo blue (186) | −11.2 kcal/mol | - | GLN 947, GLN 744 | PHE 741, THR 943 | [140] |
| 136 | Echoside A (168) | −7.5 kcal/mol | - | ARG393, PHE390, ASN394 | ALA99, LEU100, LEU73, LEU391, PHE40 | [122] |
| 137 | Echoside B (169) | −8.2 kcal/mol | - | SER47 | TRP349 | [122] |
| (b) ACE2 | ||||||
| 138 | Geraniin (177) | −7.0 kcal/mol | - | ARG403, TYR449, TYR453, GLN493, SER494, GLN498 | TYR495, GLY496 | [132] |
| 139 | Tuftsin (193) | −6.9 kcal/mol | - | SER47 | ASN51, HIS345, ASP67 | [141] |
| 140 | Epoxy-linalool oxide (155) | −13.13 kcal/mol | - | GLN101, ASN103 | GLN81, GLN98, LEU85, HIS195, ASN194, ALA193, AND GLN102 | [116] |
| 141 | Withanolide D (198) | −8.1 kcal/mol | - | - | GLY352, ARG393, TRP349, ALA348, THR347, GLU402, HIS378, HIS401, ASP382, ASP350, PHE40 | [136] |
| 142 | Morellic acid (199) | −12.1 kcal/mol | - | THR371, ASP368, LYS363, ASP367 | CYS361, MET360, ASN149, TRP271, ASP269, THR276, THR445, HIS374, GLU406, GLU375, HIS505, TYR515 | [120] |
| 143 | Pyrrolidinone (200) | −5.24 kcal/mol | - | THR434 | ILE291, PRO415, PHE438 | [137] |
| (c) GRP78 | ||||||
| 144 | Orientin (181) | −7.2 kcal/mol | - | MET433, LYS435, ARG439, GLU469 | LYS435, PRO438, PRO471, LYS556 | [129] |
| (d) NRP1 | ||||||
| 145 | Tuftsin (193) | −8.1 kcal/mol | - | ASN544 | GLU550, LYS397, PRO398, LEU551 | [141] |
| (e) TMPRSS2 | ||||||
| 146 | Withanolide D (198) | −9.7 kcal/mol (TMPRSS2) | - | ARG257, ARG241, TYR238 | GLU253, THR396, TYR250, ASN249, THR246, GLU243, MET239, TYR250 | [136] |
| RNA-dependent RNA polymerase | ||||||
| 147 | α-hederin (201) | −8.6 kcal/mol | - | ASP760, CYS622, ARG553 | THR556, ALA688, LYS500, ASP623, SER682 | [142] |
| 148 | Kaempferol (23) | −7.0 kcal/mol | - | MET87 | ASN414, ASN416, GLN18, GLN19, SER15, ASP846, LYS411, PRQ412, TYR546 | [90] |
| 149 | Ellagitannin punicalin (202) | −9.5 kcal/mol | - | ASN497, GLY590, ASP684, TYR689 | ILE494, LYS577, ASP684, ALA685 | [79] |
| 150 | Cyanidin (203) | −7.7 kcal/mol | - | ASP761, TRP617, TRP800, GLU811 | CYS622, TYR619, ASP760, ASP618, ALA762, GLY616, PHE812 | [77] |
| 151 | Amphimedoside C (78) | - | −47.9 kJ/mol | ASP623, LYS621, ALA554, ASP452, ARG553 | ARG553 | [87] |
| 152 | 14-debromoaraplysilin I (204) | - | −111.52 kJ/mol | ALA762, ASP623, ARG555, ARG553 | TRP617, CYS622, PHE694, VAL764, GLY616, VAL763, ASN695, ASP618, TYR619, ASP760 | [88] |
| 153 | Glaucogenin D (81) | −7.3 kcal/mol | - | TRP619, ASP760 | - | [89] |
| 154 | Glaucogenin C (205) | −7.3 kcal/mol | - | GLU811, ASP760 | LYS798 | [89] |
| 155 | Crocin (63) | −10.5 kcal/mol | - | ASN497, ASP499, LYS500, LYS545, ILE548, ARG836, ASP845, ARG858 | [73] | |
| 156 | Jezonofol (167) | −10.5 kcal/mol | −30.7 kcal/mol | ARG836, A19, A11, U9, LEU544, ASP845, U18, U17 | ALA574, ILE548, ILE847, TYE546, LYS545, VAL557, U20 | [91] |
| 157 | Ellagic acid (206) | −6.4 kcal/mol | - | GLY808, THR817, PRO809, HIS816, TYR831 | HIS810, LYS807, GLU802, | [121] |
| 158 | Vobtusine lactone (157) | −8.7 kcal/mol | - | SER682, ALA688 | THR687, ASP684, ASN497, ALA685, TYR689, ARG569, GLN573, ILE494, LEU576, LYS577, ALA580, SER759, GLY590 | [119] |
| 159 | Echoside A (168) | −7.3 kcal/mol | - | ASN781, SER709, LYS47 | LYS780, GLY774, ALA706 | [122] |
| 160 | Echoside B (169) | −8.0 kcal/mol | - | LYS47, SER709, LYS714, THR710 | LYS 780 | [122] |
| 161 | Arboreol (153) | −8.9 kcal/mol | - | - | - | [114] |
| Nucleocapsid | ||||||
| 162 | 4,8-dihydroxysesamin (152) | −10.7 kcal/mol | - | PRO163, THR166, LEU162, GLY70 | GLU137, GLY165, GLN164, THR167, THR77, ASN76, GLN84, SER79, PRO81, PRO163, THR136 | [114] |
| 163 | Arboreol (153) | −10.6 kcal/mol | - | GLN84, PRO163, GLY70 | ILE75, GLN164, GLY165, THR166, GLU137, LEU162, PRO81, PRO163, THR136 | [114] |
| 164 | Glycyrrhizin (156) | −7.9 kcal/mol | −30.05 kcal/mol | THR92, ARG94, ARG89, TYR110, ARG150 | ARG90, ALA91, ASN49, THR50, ALS51, PHE54, THR55, TYR112, PRO118, PRO152, ALA157 | [117] |
| 165 | Rutin (29) | −34.342 kcal/mol | - | - | [118] | |
| 166 | Trigoneoside IB (195) | −7.6 kcal/mol | - | ARG71, ARG70, THR60, SER29, ALA28, THR27, GLY125, ILE124, TYR90 | - | [134] |
| 167 | Apigenin (120) | −8.11 kcal/mol | - | PHE67, PRO68, ARG69, GLY70, GLN71, TYR124, TRP133, VAL134, THR136, GLY138, ALA139 | [143] | |
| 168 | Curcumin (86) | −8.75 kcal/mol | - | LEU161, GLN163, ALA173 | [144] | |
| Endoribonuclease | ||||||
| 169 | Hinokiflavone (175) | −8.6 kcal/mol | - | MET243 | MET243, TYR262, GLU258, HIS362, ALA256 | [79] |
| 170 | Rutin (29) | −8.68 kcal/mol | - | LEU246, GLY248, GLN245, CYS291, THR341, HIS250 | LYS345, LEU346, TYR343 | [85] |
| 171 | Glucopyranoside derivative (211) | - | −263.640 kJ/mol | GLU340, GLN245, HIS235, LYS290, GLY248, VAL292, SER294 | TRP333 MET331 LYS345, PRO344, VAL318, TYR343, GLN347, GLY247, HIS243, ASP240, LEU246, THR341, HIS250, CYS293 | [145] |
| 172 | 4-((2S,3R)-3-(hydroxymethyl)-5-((E)-3-hydroxyprop-1-en-1-yl)-7-methoxy-2,3-dihydrobenzofuran-2-yl)-2-methoxyphenol (212) | −6.26 kcal/mol | −87.52 kcal/mol | - | - | [146] |
| Helicase | ||||||
| 173 | Crocin (63) | −9.5 kcal/mol | - | ALA18, ILE20, CYS112, ASP113, TRP114, THR141, PHE145, GLY415, HIS482, ASP483, VAL484, SER485, TYR515, THR552, HIS554 | [73] | |
| 174 | Rhusflavanone (213) | −9.2 kcal/mol | - | GLU341, ASP534 | ALA312, ALA313, VAL340 | [79] |
| 175 | Morelloflavone (214) | −9.2 kcal/mol | - | LYS288, ALA316, ARG443 | THR286, ALA316, LYS320, GLY538, SER539 | [79] |
| 176 | Chromone (215) | −6.24 kcal/mol | −90.99 kcal/mol | - | - | [146] |
| 177 | Chromone (216) | −6.24 kcal/mol | −90.99 kcal/mol | - | - | [146] |
| 178 | Scirpusin A (217) | −8.9 kcal/mol | −41.9 kcal/mol | PRO514, ASN516 | ARG560, SP534, ASN177, GLU201, LYS202 | [91] |
| Methyl transferase (NSP16–NSP10 complex) | ||||||
| 179 | Robustaflavone (176) | −10.6 kcal/mol (NSP16) −7.7 kcal/mol (NSP10) | - - | ASP6897, ASP6928 ASP4335 | CYS6913, CYS6914, MET6929, ASP6931, PHE6947, GLY6869, LEU6898 ARG4331, ILE4334, LYS4346 | [79] [79] |
| 180 | Michellamine B (218) | −10.6 kcal/mol (NSP16) | - | LYS6844, CYS6913, ASP6928, ASP6928, ASN6996 | ASN6841, ASP6897, GLY6869, MET6929, LEU6898, GLU7001 | [79] |
| 181 | Chromone (219) | −6.20 kcal/mol | −75.24 kcal/mol | - | - | [146] |
| ADP phosphatase/ADPRP | ||||||
| 182 | Crocin (63) | −8.2 kcal/mol | - | ASP22, LYS44, GLY48, ALA154, PHE156 | [73] | |
| Exoribonuclease | ||||||
| 183 | Chromone (220) | −7.09 kcal/mol | −81.16 kcal/mol | - | - | [146] |
| Other Nonstructural proteins | ||||||
| 184 | Hispaglabridin B (221) | −8.5 kcal/mol | −42.88 kcal/mol | AASP61, GLY62, THR63, LEU45, ARG11, LEU10 | ALA55, ARG100, VAL103, ILE66, VAL42 | [147] |
| 185 | Licoflavone B (222) | −8.1 kcal/mol | −42.76 kcal/mol | MET13, GLY62 | GLY64, ILE66, LYS93, LEU95, ALA31, ARG11 | [147] |
| 186 | Ochnaflavone (223) | −9.1 kcal/mol | −41.43 kcal/mol | GLY94, ARG40, ASP301 | THR68, ILE92, GLY39, GLY38, PHE57, LYS59, MET13, LYS93, LEU95, SER60, PHE41, VAL42, ILE66, ARG40, GLU331 | [147] |
| 187 | 2,3-dehydrosomnifericin (224) | −12.3 kcal/mol | - | LEU126 | PHE132, ILE131, VAL49 | [148] |
| Envelope proteins | ||||||
| 188 | Trigoneoside IB (195) | −7.5 kcal/mol | - | ILE124, TYR90, ARG70, THR60, SER29, ALA28, THR27, GLY125 | - | [134] |
| 189 | Dicumarol (225) | −7.4 kcal/mol | - | - | - | [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rafiq, A.; Jabeen, T.; Aslam, S.; Ahmad, M.; Ashfaq, U.A.; Mohsin, N.u.A.; Zaki, M.E.A.; Al-Hussain, S.A. A Comprehensive Update of Various Attempts by Medicinal Chemists to Combat COVID-19 through Natural Products. Molecules 2023, 28, 4860. https://doi.org/10.3390/molecules28124860
Rafiq A, Jabeen T, Aslam S, Ahmad M, Ashfaq UA, Mohsin NuA, Zaki MEA, Al-Hussain SA. A Comprehensive Update of Various Attempts by Medicinal Chemists to Combat COVID-19 through Natural Products. Molecules. 2023; 28(12):4860. https://doi.org/10.3390/molecules28124860
Chicago/Turabian StyleRafiq, Ayesha, Tooba Jabeen, Sana Aslam, Matloob Ahmad, Usman Ali Ashfaq, Noor ul Amin Mohsin, Magdi E. A. Zaki, and Sami A. Al-Hussain. 2023. "A Comprehensive Update of Various Attempts by Medicinal Chemists to Combat COVID-19 through Natural Products" Molecules 28, no. 12: 4860. https://doi.org/10.3390/molecules28124860
APA StyleRafiq, A., Jabeen, T., Aslam, S., Ahmad, M., Ashfaq, U. A., Mohsin, N. u. A., Zaki, M. E. A., & Al-Hussain, S. A. (2023). A Comprehensive Update of Various Attempts by Medicinal Chemists to Combat COVID-19 through Natural Products. Molecules, 28(12), 4860. https://doi.org/10.3390/molecules28124860