
Functional Study and Efficient Catalytic Element Mining of CYP76AHs in Salvia Plants
 ,
,
Abstract
1. Introduction
2. Results
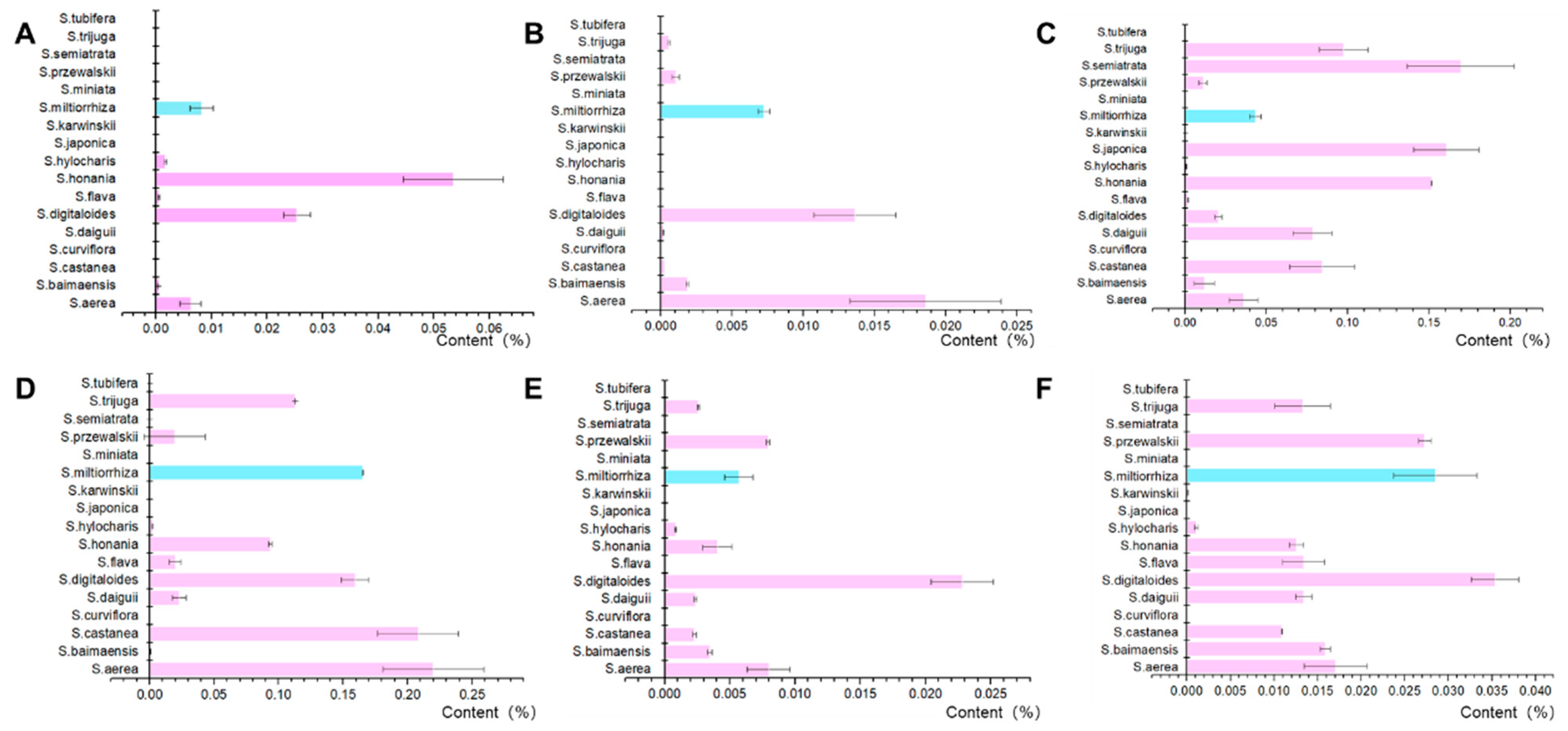
2.1. Chemical Constituents of Salvia
2.2. Phylogenetic Analysis of Candidate CYP76AH Subfamily Genes
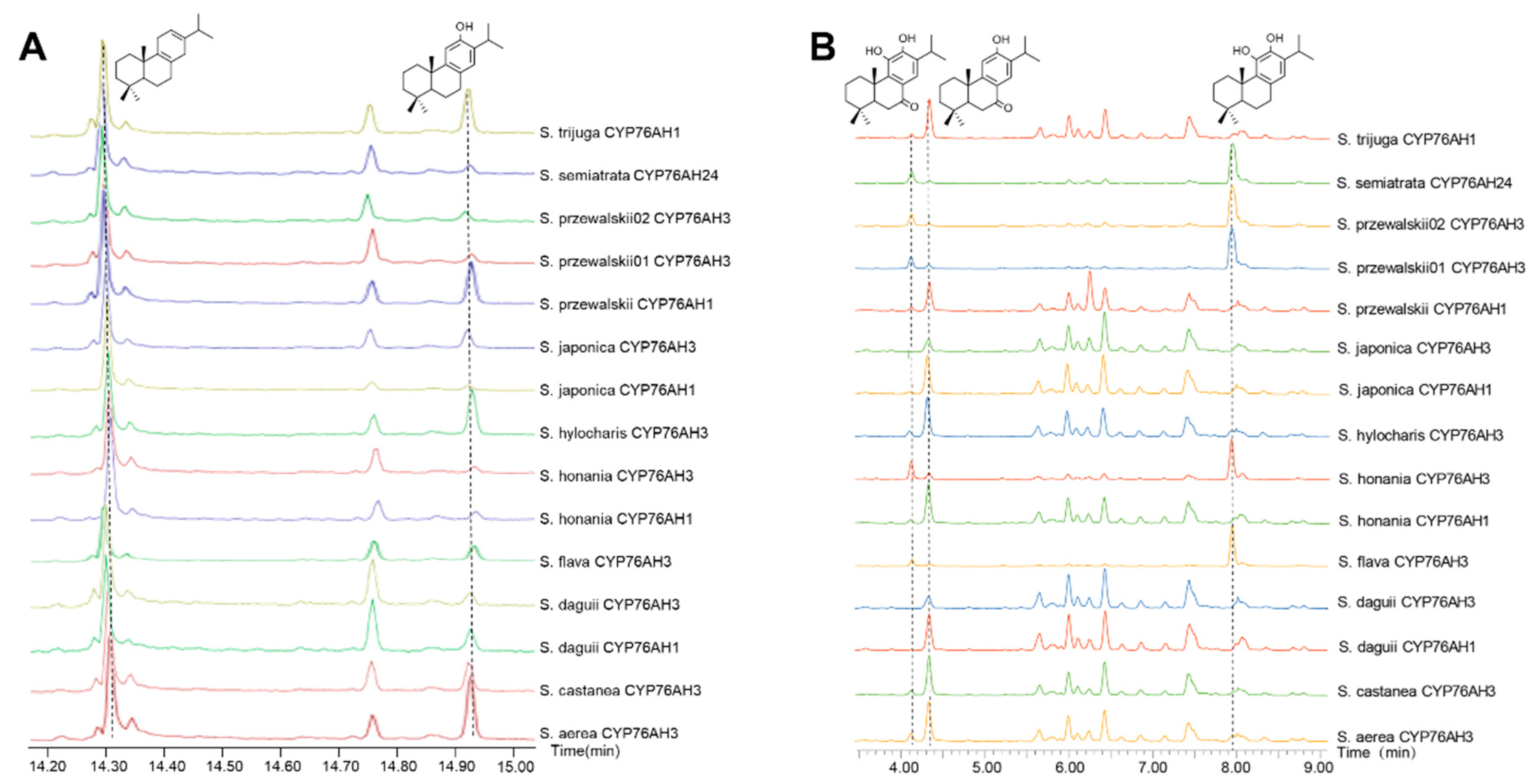
2.3. Biochemical Characterization of CYP76Ahs
2.4. Correlation Analysis of CYP76AH Protein Structure and Activity
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. Chemical Constituents of Salvia
4.3. Identification of CYP76AH Genes
4.4. Gene Diversity of CYP76AH Genes
4.5. Heterologous Expression of CYP76AH in Yeast and In Vitro Activity Assays
4.6. Chemical Analysis of the Enzyme Reaction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wu, Y.B.; Ni, Z.Y.; Shi, Q.W.; Dong, M.; Kiyota, H.; Gu, Y.C. Constituents from Salvia species and their biological activities. Chem. Rev. 2012, 11, 5967–6026. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.R.; Bhat, W.W.; Bibik, J.; Turmo, A.; Hamberger, B.; Evolutionary Mint Genomics Consortium; Hamberger, B. A database-driven approach identifies additional diterpene synthase activities in the mint family (Lamiaceae). J. Biol. Chem. 2019, 294, 1349–1362. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Gaire, B.P. Tanshinone IIA: A phytochemical as a promising drug candidate for neurodegenerative diseases. Pharmacol. Res. 2021, 169, 105661. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Zhang, M.; Liu, J.-N.; Zhao, X.; Zhang, Y.-Q.; Fang, L. Tanshinone IIA: A Review of its Anticancer Effects. Front. Pharmacol. 2021, 11, 611087. [Google Scholar] [CrossRef] [PubMed]
- Meunier, B.; Visser, S.; Shaik, S. Mechanism of Oxidation Reactions Catalyzed by Cytochrome P450 Enzyme. Chem. Rev. 2004, 9, 3947–3980. [Google Scholar] [CrossRef] [PubMed]
- Bathe, U.; Frolov, A.; Porzel, A.; Tissier, A. CYP76 Oxidation Network of Abietane Diterpenes in Lamiaceae Reconstituted in Yeast. J. Agric. Food Chem. 2019, 67, 13437–13450. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhou, Y.J.; Hillwig, M.L.; Shen, Y.; Yang, L.; Wang, Y.; Zhang, X.; Liu, W.; Peters, R.J.; Chen, X.; et al. CYP76AH1 catalyzes turnover of miltiradiene in tanshinones biosynthesis and enables heterologous production of ferruginol in yeasts. Proc. Natl. Acad. Sci. USA 2013, 110, 12108–12113. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Ma, X.; Cai, Y.; Ma, Y.; Zhan, Z.; Zhou, Y.J.; Liu, W.; Guan, M.; Yang, J.; Cui, G.; et al. Cytochrome P450 promiscuity leads to a bifurcating biosynthetic pathway for tanshinones. New Phytol. 2015, 210, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Scheler, U.; Brandt, W.; Porzel, A.; Rothe, K.; Manzano, D.; Božić, D.; Papaefthimiou, D.; Balcke, G.U.; Henning, A.; Lohse, S.; et al. Elucidation of the biosynthesis of carnosic acid and its reconstitution in yeast. Nat. Commun. 2016, 7, 12942. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Y.; Yang, L.; Liu, Y.; Xu, Z.-G.; Gao, J.; Huang, Y.-B.; Xu, J.-J.; Fan, H.; Kong, Y.; Wei, Y.-K.; et al. The sage genome provides insight into the evolutionary dynamics of diterpene biosynthesis gene cluster in plants. Cell Rep. 2022, 40, 111236. [Google Scholar] [CrossRef] [PubMed]
- Pateraki, I.; Andersen-Ranberg, J.; Jensen, N.B.; Wubshet, S.G.; Heskes, A.M.; Forman, V.; Hallström, B.; Hamberger, B.; Motawia, M.S.; Olsen, C.E.; et al. Total biosynthesis of the cyclic AMP booster forskolin from Coleus forskohlii. eLife 2017, 6, e23001. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.C.; Su, P.; Zhang, R.Z.; Gao, L.H.; Wang, J.D.; Hu, T.Y.; Zhou, J.W.; Zhang, Y.F.; Zhao, Y.; Liu, Y.; et al. Genome of Tripterygium wilfordii and identification of cytochrome P450 involved in triptolide biosynthesis. Nat. Commun. 2020, 11, 971. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, J.; Ma, L.; Tu, L.; Hu, T.; Wu, X.; Su, P.; Zhao, Y.; Liu, Y.; Li, D.; et al. Tandemly duplicated CYP82Ds catalyze 14-hydroxylation in triptolide biosynthesis and precursor production in Saccharomyces cerevisiae. Nat. Commun. 2023, 14, 875. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Cui, G.; Chen, T.; Ma, X.; Wang, R.; Jin, B.; Yang, J.; Kang, L.; Tang, J.; Lai, C.; et al. Expansion within the CYP71D subfamily drives the heterocyclization of tanshinones synthesis in Salvia miltiorrhiza. Nat. Commun. 2021, 12, 685. [Google Scholar] [CrossRef] [PubMed]
- Urban, P.; Mignotte, C.; Kazmaier, M.; Delorme, F.; Pompon, D. Cloning, Yeast Expression, and Characterization of the Coupling of Two Distantly Related Arabidopsis thaliana NADPH-Cytochrome P450 Reductases with P450 CYP73A5. J. Biol. Chem. 1997, 272, 19176–19186. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Ma, Y.; Chen, T.; Ma, X.; Xu, Y.; Bo, J.; Li, Q.; Jin, B.; Wang, Y.; Li, Y.; et al. Functional Integration of Two CYP450 Genes Involved in Biosynthesis of Tanshinones for Improved Diterpenoid Production by Synthetic Biology. ACS Synth. Biol. 2020, 9, 1763–1770. [Google Scholar] [CrossRef]
- Forman, V.; Bjerg-Jensen, N.; Dyekjær, J.D.; Møller, B.L.; Pateraki, I. Engineering of CYP76AH15 can improve activity and specificity towards forskolin biosynthesis in yeast. Microb. Cell Factor. 2018, 17, 181. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Utomo, J.C.; Park, K.; Pascoe, C.A.; Chiorean, S.; Ngo, I.; Pelot, K.A.; Pan, C.-H.; Kim, S.-W.; Zerbe, P.; et al. Cytochrome P450-Catalyzed Biosynthesis of a Dihydrofuran Neoclerodane in Magic Mint (Salvia divinorum). ACS Catal. 2021, 12, 777–782. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid Similarity (%) | Sequence Differences | |||||
|---|---|---|---|---|---|---|
| SmCYP76AH1 | SmCYP76AH3 | SpCYP76AH22 | SpCYP76AH24 | SmCYP76AH1 | SmCYP76AH3 | |
| SaeCYP76AH3 | 99.60 | 79.60 | 78.26 | 77.73 | S3Y, K329R | |
| ScaCYP76AH3 | 95.91 | 77.00 | 75.76 | 75.24 | S3Y, KS494NPRIRNTTHYRARASTWNRS | |
| SdaCYP76AH1 | 98.99 | 78.99 | 77.67 | 77.14 | ||
| SdaCYP76AH3 | 80.00 | 99.39 | 86.93 | 86.85 | S5P, Q161E, K320Q | |
| SflCYP76AH3 | 79.39 | 96.76 | 87.33 | 86.85 | S5P, F12L, S14T, S21F, S22F, R157K, Q161K, A258T, D261N, N277Q, N279S, V315I, A322G, A394G, D466N, R488K | |
| ShoCYP76AH1 | 99.39 | 79.60 | 78.26 | 77.53 | F4S, T18I, K384M | |
| ShoCYP76AH3 | 79.80 | 99.39 | 86.73 | 86.65 | S3N, S5P, K320E | |
| ShyCYP76AH3 | 99.80 | 79.80 | 78.46 | 77.93 | S3I | |
| SjaCYP76AH1 | 98.99 | 79.60 | 77.43 | 76.49 | D2E, F4S, I13T, T18I, K156R | |
| SjaCYP76AH3 | 80.00 | 98.58 | 86.93 | 86.65 | S5P, H156D, R157K, D319H, K320Q, R345S, Q473G | |
| SprCYP76AH1 | 99.19 | 79.39 | 78.06 | 77.53 | D2E, F4S, N279S | |
| SprCYP76AH3-01 | 80.30 | 96.76 | 87.52 | 87.65 | S3Y, S5P, S14T, S21F, S22F, R157K, Q161K, D261N, N277Q, N279S, A322G, D406E, A407S, Q409E, D466N, R488K | |
| SprCYP76AH3-02 | 76.67 | 85.49 | 83.88 | 83.23 | ||
| SseCYP76AH24 | 79.39 | 79.60 | 86.93 | 86.25 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Yang, D.; Guo, J.; Liu, X.; Li, Q.; Su, P.; Wang, J.; Ma, Y.; Huang, L. Functional Study and Efficient Catalytic Element Mining of CYP76AHs in Salvia Plants. Molecules 2023, 28, 4711. https://doi.org/10.3390/molecules28124711
Zhao Z, Yang D, Guo J, Liu X, Li Q, Su P, Wang J, Ma Y, Huang L. Functional Study and Efficient Catalytic Element Mining of CYP76AHs in Salvia Plants. Molecules. 2023; 28(12):4711. https://doi.org/10.3390/molecules28124711
Chicago/Turabian StyleZhao, Zhenyu, Dongfeng Yang, Juan Guo, Xiuyu Liu, Qishuang Li, Ping Su, Jian Wang, Ying Ma, and Luqi Huang. 2023. "Functional Study and Efficient Catalytic Element Mining of CYP76AHs in Salvia Plants" Molecules 28, no. 12: 4711. https://doi.org/10.3390/molecules28124711
APA StyleZhao, Z., Yang, D., Guo, J., Liu, X., Li, Q., Su, P., Wang, J., Ma, Y., & Huang, L. (2023). Functional Study and Efficient Catalytic Element Mining of CYP76AHs in Salvia Plants. Molecules, 28(12), 4711. https://doi.org/10.3390/molecules28124711