
Chemical Characterization and Cytotoxic and Antioxidant Activity Evaluation of the Ethanol Extract from the Bulbs of Pancratium maritimun Collected in Sicily


 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Chemical Characterization
2.1.1. Nuclear Magnetic Resonance Analysis of Bulbs’ Extract from P. maritimum
2.1.2. LC-DAD-MSn Analysis of Bulbs’ Extract from P. maritimum
2.2. Absence of Cytotoxic Effect by P. maritimum Bulbs’ Extract on Human Intestinal Cells
2.3. Down-Regulation of Reactive Oxygen Species (ROS) Production in LPS-Induced Cells by P. maritimum Bulbs’ Extract
3. Materials and Methods
3.1. Plant Material
3.2. Extraction of Plant Material
3.3. Chemical Analysis
3.4. Cell Culture, Differentiation and Treatments
3.5. Trypan Blue Exclusion Assay
3.6. Assay for Cellular Antioxidant Activity
3.7. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meerow, A.D.; Snijman, D. The Families and Genera of Vascular Plants III. Flowering Plants Monocotyledons; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; pp. 83–110. [Google Scholar]
- Meerow, A.; Francisco-Ortega, J.; Kuhn, D.; Schnell, R. Phylogenetic relationships and biogeography within the Eurasian Clade of Amaryllidaceae based on plastid ndhF and nrDNA ITS sequences: Lineage sorting in a reticulate area? Syst. Bot. 2006, 31, 42–60. [Google Scholar] [CrossRef]
- Plants of the World Online. Available online: https://powo.science.kew.org/ (accessed on 12 January 2023).
- Davis, P.H. Flora of Turkey and the East Aegean Islands; Edinburgh University Press: Edinburgh, UK, 1984; Volume 8, pp. 1–632. [Google Scholar]
- Press Syndicate of the University of Cambridge. Flora Europaea; Tutin, T.G., Ed.; Cambridge University Press: Cambridge, UK, 1980; Volume 5, pp. 1–452. [Google Scholar]
- Perrone, B.; Salmeri, C.; Brullo, S.; Colombo, P.; De Castro, O. What do leaf anatomy and micro-morphology tell us about the psammophilous P. maritimum L. (Amaryllidaceae) in response to sand dune conditions? Flora 2015, 213, 20–31. [Google Scholar] [CrossRef]
- Du Merac, M.L. Systematics and biochemistry of the Amaryllidaceae—Alkaloid content of P. maritimum. Compt. Rend. 1954, 239, 300–302. [Google Scholar]
- Cedrón, J.C.; Del Arco-Aguilar, M.; Estévez-Braun, A.; Ravelo, Á.G. Chemistry and biology of Pancratium Alkaloids. In The Alkaloids: Chemistry and Biology; Cordell, G.A., Ed.; Elesevier: Amsterdam, The Netherlands, 2010; Volume 68, pp. 1–37. [Google Scholar]
- Abou-Donia, A.H.; De Giulio, A.; Evidente, A.; Gaber, M.; Habib, A.A.; Lanzetta, R.; El Din, A.A.S. Narciclasine-4-O-β-D-glucopyranoside, a glucosyloxy amidic phenanthridone derivative from P. maritimum. Phytochemistry 1991, 30, 3445–3448. [Google Scholar] [CrossRef]
- Abou-Donia, A.H.; Abib, A.A.; El Din, A.S.; Evidente, A.; Gaber, M.; Scopa, A. Two betaine-type alkaloids from Egyptian P. maritimum. Phytochemistry 1992, 31, 2139–2141. [Google Scholar] [CrossRef]
- Ibrahim, S.R.; Mohamed, G.A.; Shaala, L.A.; Youssef, D.T.; El Sayed, K.A. New alkaloids from P. maritimum. Planta Med. 2013, 79, 1480–1484. [Google Scholar] [CrossRef]
- Youssef, D.T.A.; Frahm, A.W. Alkaloids of the flowers of P. maritimum. Planta Med. 1998, 64, 669–670. [Google Scholar] [CrossRef]
- Youssef, D.T.A. Further alkaloids from the flowers of P. maritimum. Pharmazie 1999, 54, 535–537. [Google Scholar]
- Hetta, M.H.; Shafei, A.A. Comparative cytotoxic and antimicrobial activities of the alkaloid content of Egyptian P. maritimum L. fruits and flowers. J. Am. Sci. 2013, 9, 104–109. [Google Scholar]
- Schrader, K.K.; Avolio, F.; Andolfi, A.; Cimmino, A.; Evidente, A. Ungeremine and its hemisynthesized analogues as bactericides against Flavobacterium columnare. J. Agric. Food Chem. 2013, 61, 1179–1183. [Google Scholar] [CrossRef]
- Youssef, D.T.A.; Shaala, L.A.; Altyar, A.E. Cytotoxic phenylpropanoid derivatives and alkaloids from the flowers of P. maritimum L. Plants 2022, 11, 476. [Google Scholar] [CrossRef] [PubMed]
- Berkov, S.; Evstatieva, L.; Popov, S. Alkaloids in Bulgarian P. maritimum L. Z. Naturforsch. C 2004, 59, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, B.; Kaya, G.I.; Somer, N.U. Chemical composition and enzyme inhibitory activities of Turkish P. maritimum bulbs. Nat. Prod. Commun. 2019, 14, 1–4. [Google Scholar] [CrossRef]
- Şener, B.; Konukol, S.; Kruk, C.; Pandit, U.K. New crinine-type alkaloids from P. maritimum L- growing in Turkey. Nat. Prod. Lett. 1993, 1, 287–291. [Google Scholar] [CrossRef]
- Şener, B.; Konukol, S.; Kruk, C.; Pandit, U.K. Alkaloids from Amaryllidaceae: IV. Alkaloids from the aerial parts of P. maritimum L. Gazi Univ. Eczaci. Fak. Derg. 1993, 10, 83–86. [Google Scholar]
- Şener, B.; Konukol, S.; Kruk, C.; Pandit, U.K. Alkaloids from Amaryllidaceae III. Alkaloids from the bulbs of P. maritimum. Nat. Prod. Sci. 1998, 4, 148–152. [Google Scholar]
- Şener, B.; Orhan, I.; Satayavivad, J. Antimalarial activity screening of some alkaloids and the plant extracts from Amaryllidaceae. Phytother. Res. 2003, 17, 1220–1223. [Google Scholar] [CrossRef]
- Sanaa, A.; Boussaid, M.; Ben Fadhel, N. Alkaloids and sodium alginate in Tunisian P. maritimum L. Acta Hortic. 2010, 853, 215–218. [Google Scholar] [CrossRef]
- Sancha, S.A.R.; Gomes, V.A.; Loureiro, J.B.; Saraiva, L.; Ferreira, M.J.U. Amaryllidaceae-type alkaloids from Pancratium maritimum: Apoptosis-inducing effect and cell cycle arrest on triple-negative breast cancer cells. Molecules 2022, 27, 5759. [Google Scholar] [CrossRef]
- Suau, R.; Gómez, A.I.; Rico, R.; Tato, M.P.V.; Castedo, L.; Riguera, R. Alkaloid N-oxides of Amaryllidaceae. Phytochemistry 1998, 27, 3285–3287. [Google Scholar] [CrossRef]
- Tato, M.P.V.; Castedo, L.; Riguera, R. New alkaloids from P. maritimum L. Heterocycles 1988, 27, 2833–2838. [Google Scholar] [CrossRef]
- Masi, M.; Di Lecce, R.; Mérindol, N.; Girard, M.P.; Berthoux, L.; Desgagné-Penix, I.; Calabrò, V.; Evidente, A. Cytotoxicity and antiviral properties of alkaloids isolated from P. maritimum. Toxins 2022, 14, 262. [Google Scholar] [CrossRef] [PubMed]
- Boit, H.G.; Ehmke, H. Alkaloide von Nerine bowdenii, Crinum powellii, Amaryllis belladonna und P. maritimum (XII. Mitteil. Über Amaryllidaceen-Alkaloide). Chem. Ber. 1956, 89, 2093–2097. [Google Scholar] [CrossRef]
- Petit, G.R.; Petit, G.R., III; Groszek, G.; Backhaus, R.A.; Doubek, D.L.; Barr, R.J. Antineoplastic agents, 301. An investigation of the Amaryllidaceae genus Hymenocallis. J. Nat. Prod. 1995, 58, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.A.; Makboul, M.A.; Attia, A.A.; Ali, D.T. Chromones and flavans from P. maritimum. Phytochemistry 1990, 29, 625–627. [Google Scholar] [CrossRef]
- Youssef, D.T.A.; Ramadan, M.A.; Khalifa, A.A. Acetophenones, a chalcone, a chromone and flavonoids from P. maritimum. Phytochemistry 1998, 49, 2579–2583. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Shaala, L.A.; Youssef, D.T.A. Non-alkaloidal compounds from the bulbs of the Egyptian plant P. maritimum. Z. Naturforsch. C 2014, 69, 92–98. [Google Scholar] [CrossRef]
- Rokbeni, N.; M’rabet, Y.; Cluzet, S.; Richard, T.; Krisa, S.; Boussaid, M.; Boulila, A. Determination of phenolic composition and antioxidant activities of P. maritimum L. from Tunisia. Ind. Crop. Prod. 2016, 94, 505–513. [Google Scholar] [CrossRef]
- Nikolova, M.; Gevrenova, R. Determination of phenolic acids in Amaryllidaceae species by high performance liquid chromatography. Pharm. Biol. 2005, 43, 289–291. [Google Scholar] [CrossRef]
- Sanaa, A.; Boulila, A.; Boussaid, M.; Fadhel, N.B. Alginic acid and derivatives, new polymers from the endangered P. maritimum L. Ind. Crop. Prod. 2013, 44, 290–293. [Google Scholar] [CrossRef]
- Kaya, G.I.; Sarikaya, B.; Çiçek, D.; Somer, N.U. In vitro cytotoxic activity of Sternbergia sicula, S. lutea and P. maritimum extracts. Hacet. Univ. J. Fac. Pharm. 2010, 30, 41–48. [Google Scholar]
- Calderón-Montaño, J.M.; Martínez-Sánchez, S.M.; Jiménez-González, V.; Burgos-Morón, E.; Guillén-Mancina, E.; Jiménez-Alonso, J.J.; Díaz-Ortega, P.; García, F.; Aparicio, A.; López-Lázaro, M. Screening for selective anticancer activity of 65 extracts of plants collected in western Andalusia, Spain. Plants 2021, 10, 2193. [Google Scholar] [CrossRef] [PubMed]
- Leporini, M.; Catinella, G.; Bruno, M.; Falco, T.; Tundis, R.; Loizzo, M.R. Investigating the antiproliferative and antioxidant properties of P. maritimum L. (Amaryllidaceae) stems, flowers, bulbs, and fruits extracts. Evid. Based Complem. Altern. Med. 2018, 2018, 9301247. [Google Scholar] [CrossRef] [PubMed]
- Tayoub, G.; Al-Odat, M.; Amer, A.; Aljapawe, A.; Ekhtiar, A. Antiproliferative effects of P. maritimum extracts on normal and cancerous cells. Iran. J. Med. Sci. 2018, 43, 52–64. [Google Scholar]
- Sür-Altiner, D.; Gürkan, E.; Mutlu, G.; Tuzlaci, E.; Ang, O. The antifungal activity of P. maritimum. Fitoterapia 1999, 70, 187–189. [Google Scholar] [CrossRef]
- Tosun, F.; Kizilay, C.A.; Şener, B.; Vural, M.; Palittapongarnpim, P. Antimycobacterial activity of some Turkish plants. Pharm. Biol. 2004, 42, 39–43. [Google Scholar] [CrossRef]
- Çakici, I.; Ulug, H.Y.; Inci, S.; Tunçtan, B.; Abacioglu, N.; Kanzik, I.; Sener, B. Antinociceptive effect of some Amaryllidaceae plants in mice. J. Pharm. Pharmacol. 1997, 49, 828–830. [Google Scholar] [CrossRef]
- El-Sayed, N.M.; Ismail, K.A.; Ahmed, S.A.-E.-G.; Hetta, M.H. In vitro amoebicidal activity of ethanol extracts of Arachis hypogaea L., Curcuma longa L. and P. maritimum L. on Acanthamoeba castellanii cysts. Parasitol. Res. 2012, 110, 1985–1992. [Google Scholar] [CrossRef]
- Soltan, M.M.; Hamed, A.R.; Hetta, M.H.; Hussein, A.A. Egyptian P. maritimum L. flowers as a source of anti-Alzheimer’s agents. Bull. Fac. Pharm. Cairo Univ. 2015, 53, 19–22. [Google Scholar] [CrossRef]
- Orhan, I.; Sener, B. Bioactivity-directed fractionation of alkaloids from some Amaryllidaceae plants and their anticholinesterase activity. Chem. Nat. Comp. 2003, 39, 383–386. [Google Scholar] [CrossRef]
- Badalamenti, N.; Ilardi, V.; Rosselli, S.; Bruno, M.; Maggi, F.; Leporini, M.; Falco, T.; Loizzo, M.R.; Tundis, R. Ferulago nodosa subsp. geniculata (Guss.) Troia & Raimondo from Sicily (Italy): Isolation of essential oil and evaluation of its bioactivity. Molecules 2020, 25, 3249. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, N.; Modica, A.; Ilardi, V.; Bruno, M.; Maresca, V.; Zanfardino, A.; Di Napoli, M.; Castagliuolo, G.; Varcamonti, M.; Basile, A. Daucus carota subsp. maximus (Desf.) Ball from Pantelleria, Sicily (Italy): Isolation of essential oils and evaluation of their bioactivity. Nat. Prod. Res. 2021, 19, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Cicio, A.; Serio, R.; Zizzo, M.G. Anti-inflammatory potential of Brassicaceae-derived phytochemicals: In vitro and in vivo evidence for a putative role in the prevention and treatment of IBD. Nutrients 2023, 15, 31. [Google Scholar] [CrossRef] [PubMed]
- Cicio, A.; Badalamenti, N.; Bruno, M. The ethnobotany, phytochemistry, and biological properties of genus Phagnalon (Asteraceae): A review. Nat. Prod. Res. 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; Di Napoli, M.; Castagliuolo, G.; Badalamenti, N.; Cicio, A.; Bruno, M.; Piacente, S.; Maresca, V.; Cianciullo, P.; Capasso, L.; et al. The chemical composition of the aerial parts of Stachys spreitzenhoferi (Lamiaceae) extract growing in Kythira Island (Greece) and its antioxidant, antimicrobial, anti-inflammatory and antiproliferative properties. Phytochemistry 2022, 203, 113373. [Google Scholar] [CrossRef] [PubMed]
- Ptak, A.; El Tahchy, A.; Dupire, F.; Boisbrun, M.; Henry, M.; Chapleur, Y.; Moś, M.; Laurain-Mattar, D. LCMS and GCMS for the screening of alkaloids in natural and in vitro extracts of Leucojum aestivum. J. Nat. Prod. 2009, 72, 142–147. [Google Scholar] [CrossRef]
- Ponce de León-Rodríguez, M.C.; Guyot, J.P.; Laurent-Babot, C. Intestinal in vitro cell culture models and their potential to study the effect of food components on intestinal inflammation. Crit. Rev. Food Sci. Nutr. 2019, 59, 3648–3666. [Google Scholar] [CrossRef]
- Evidente, A.; Cicala, M.R.; Giudicianni, I.; Randazzo, G.; Riccio, R. 1H and 13C NMR analysis of lycorine and α-dihydrolycorine. Phytochemistry 1983, 22, 581–584. [Google Scholar] [CrossRef]
- Berkov, S.; Reyes-Chilpa, R.; Codina, C.; Viladomat, F.; Batista, J. Revised NMR data for incartine: An alkaloid from Galanthus elwesii. Molecules 2007, 12, 1430–1435. [Google Scholar] [CrossRef]
- Şener, B.; Könükol, S.; Kruk, C.; Pandit, U.K. Alkaloids from Amaryllidaceae, I: Alkaloids of lycorine and lycorenine class from P. maritimum L. Arch. Pharm. 1993, 326, 61–62. [Google Scholar] [CrossRef]
- Bastida, J.; Codina, C.; Viladomat, F.; Rubiralta, M.; Quirion, J.C.; Husson, H.P.; Ma, G. Narcissus alkaloids, XIII. Complete assignment of the NMR Spectra of papyramine and 6-epi-papyramine by two-dimensional NMR Spectroscopy. J. Nat. Prod. 1990, 56, 1456–1462. [Google Scholar] [CrossRef]
- Codina, C.; Viladomat, F.; Bastida, J.; Rubiralta, M.; Quirion, J.-C. 2d Nmr studies of Lycorenine as a model for the structural assignment of Lycorenine-Type Alkaloids. Nat. Prod. Lett. 1992, 1, 85–92. [Google Scholar] [CrossRef]
- Makhkamova, A.U.; Safanova, E.V. Method for quantitative determination of lycorine in leaves of Ungernia sewerzowi. Chem. Nat. Comp. 1995, 30, 529–530. [Google Scholar] [CrossRef]
- Evidente, A.; Iasiello, I.; Randazzo, G. Rapid quantitative analysis of lycorine by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1983, 281, 362–366. [Google Scholar] [CrossRef]
- Konukol, S.; Sener, B. High-pressure liquid chromatographic analysis of some Amaryllidaceae alkaloids from P. maritimum L. J. Fac. Pharm. Gazi Univ. 1992, 9, 89–95. [Google Scholar]
- Kaya, G.I.; Fillik, A.; Hısıl, Y.; Unver, N. High pressure liquid chromatographic analysis of lycorine in four Galanthus species growing in Turkey. Turk. J. Pharm. Sci. 2004, 1, 105–114. [Google Scholar]
- Çitoğlu, G.S.; Yılmaz, B.S.; Bahadır, O. Quantitative analysis of lycorine in Sternbergia species growing in Turkey. Chem. Nat. Comp. 2008, 44, 826–828. [Google Scholar] [CrossRef]
- Kaya, G.I.; Cicek, D.; Sarıkaya, B.; Onur, M.A.; Somer, N.U. HPLC-DAD analysis of lycorine in Amaryllidaceae species. Nat. Prod. Commun. 2010, 5, 873–876. [Google Scholar] [CrossRef]
- Pham, L.H.; Döpke, W.; Wagner, J.; Mügge, C. Alkaloids from Crinum amabile. Phytochemistry 1998, 48, 371–376. [Google Scholar] [CrossRef]
- Thi Ngoc Tram, N.; Titorenkova, T.V.; Bankova, V.S.; Handjieva, N.V.; Popov, S.S. Crinum L. (Amaryllidaceae). Fitoterapia 2002, 73, 183–208. [Google Scholar] [CrossRef]
- Kreh, M.; Matusch, R. O-Methyloduline and N-demethylmasonine, alkaloids from Narcissus pseudonarcissus. Phytochemistry 1995, 38, 1533–1535. [Google Scholar] [CrossRef]
- Cahlíková, L.; Ločárek, M.; Benešová, N.; Kučera, R.; Chlebek, J.; Novák, Z.; Opletal, L. Isolation and cholinesterase inhibitory activity of Narcissus extracts and Amaryllidaceae alkaloid. Nat. Prod. Commun. 2013, 8, 781–785. [Google Scholar] [CrossRef]
- Liao, N.; Ao, M.; Zhang, P.; Yu, L. Extracts of Lycoris aurea induce apoptosis in murine sarcoma S180 cells. Molecules 2012, 17, 3723–3735. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-D.; Zhang, Y.; He, H.-P.; Li, S.-F.; Tang, G.-H.; Chen, D.-Z.; Cao, M.-M.; Di, Y.-T.; Hao, X.-J. A new amaryllidaceae alkaloid from the bulbs of Lycoris radiata. Chin. J. Nat. Med. 2013, 11, 406–410. [Google Scholar] [CrossRef]
- Kobayashi, S.; Tokumoto, T.; Kihara, M.; Imakura, Y.; Shingu, T.; Taira, Z. Alkaloidal constituents of Crinum latifolium and Crinum bulbispermum (Amaryllidaceae). Chem. Pharm. Bull. 1984, 32, 3015–3022. [Google Scholar] [CrossRef]
- Hanh, T.T.H.; Anh, D.H.; Huong, P.T.T.; Thanh, N.V.; Trung, N.Q.; Cuong, T.V.; Mai, N.T.; Cuong, N.T.; Cuong, N.X.; Nam, N.H.; et al. Crinane, augustamine, and β-carboline alkaloids from Crinum latifolium. Phytochem. Lett. 2018, 24, 27–30. [Google Scholar] [CrossRef]
- Nair, J.J.; Machocho, A.K.; Campbell, W.E.; Brun, R.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Crinum macowani. Phytochemistry 2000, 54, 945–950. [Google Scholar] [CrossRef]
- Lyle, R.E.; Kielar, E.A.; Crowder, J.R.; Wildman, W.C. The alkaloids of Nerine bowdenii W. Wats, and Crinum moorei J. D. Hook. J. Am. Chem. Soc. 1960, 82, 2620–2625. [Google Scholar] [CrossRef]
- Cahlíková, L.; Zavadil, S.; Macaḱová, K.; Valterová, I.; Kulhańková, A.; Hošt’álková, A.; Kuneš, J.; Opletal, L. Isolation and cholinesterase activity of amaryllidaceae alkaloids from Nerine bowdenii. Nat. Prod. Commun. 2011, 6, 1827–1830. [Google Scholar] [CrossRef]
- Fales, H.M.; Wildman, W.C. Alkaloids of the Amaryllidaceae. XIX. On the structures of crinamidine, flexinine, and nerbowdine. J. Org. Chem. 1961, 26, 181–187. [Google Scholar] [CrossRef]
- Queckenberg, O.R.; Frahm, A.W.; Muller-Doblies, D.; Muller-Doblies, U. Alkaloids from Brunsvigia gregari. Planta Med. 1995, 61, 581. [Google Scholar] [CrossRef] [PubMed]
- Viladomat, F.; Almanza, G.R.; Codina, C.; Bastida, J.; Campbell, W.E.; Mathee, S. Alkaloids from Brunsvigia orientalis. Phytochemistry 1996, 43, 1379–1384. [Google Scholar] [CrossRef]
- Machocho, A.; Chhabra, S.C.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Ammocharis tinneana. Phytochemistry 1999, 51, 1185–1191. [Google Scholar] [CrossRef]
- Katoch, D.; Kumar, S.; Kumar, N.; Singh, B. Simultaneous quantification of Amaryllidaceae alkaloids from Zephyranthes grandiflora by UPLC-DAD/ESI-MS/MS. J. Pharm. Biomed. Anal. 2012, 71, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Adedayo, B.C.; Oyeleye, S.I.; Okeke, B.M.; Oboh, G. Anti-cholinesterase and antioxidant properties of alkaloid and phenolic-rich extracts from pawpaw (Carica papaya) leaf: A comparative study. Flav. Fragr. J. 2021, 36, 47–54. [Google Scholar] [CrossRef]
- He, M.; Qu, C.; Gao, O.; Hu, X.; Hong, X. Biological and pharmacological activities of Amaryllidaceae alkaloids. RSC Adv. 2015, 5, 16562–16574. [Google Scholar] [CrossRef]
- Nair, J.J.; Van Staden, J. Cytotoxicity studies of lycorine alkaloids of the Amaryllidaceae. Nat. Prod. Commun. 2014, 9, 1193–1210. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive oxygen species in metabolic and inflammatory signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Taie, H.; Esawy, M.A.T.; Mohamed, M.A.E.M. Antioxidant and antimicrobial activities of different parts of P. maritimum. Planta Med. 2015, 81, 166–167. [Google Scholar] [CrossRef]
- Cao, Z.; Yang, P.; Zhou, Q. Multiple biological functions and pharmacological effects of lycorine. Sci. China Chem. 2013, 56, 1382–1391. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Liu, D.; Yu, X.; Sun, H.; Li, Y. A Caco-2 cell-based quantitative antioxidant activity assay for antioxidants. Food Chem. 2015, 175, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Elke, C.; Rosenberg, I.M.; Brandwein, S.L.; Beck, P.L.; Reinecker, H.C.; Podolsky, D.K. Lipopolysaccharide activates distinct signaling pathways in intestinal epithelial cell lines expressing toll-like receptors. J. Immunol. 2000, 164, 966–972. [Google Scholar] [CrossRef]
- Tunçer, S.; Banerjee, S. Determination of autophagy in the Caco-2 spontaneously differentiating model of intestinal epithelial cells. Methods Mol. Biol. 2019, 1854, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Luparello, C.; Asaro, D.M.L.; Cruciata, I.; Hassell-Hart, S.; Sansook, S.; Spencer, J.; Caradonna, F. Cytotoxic activity of the histone deacetylase 3-selective inhibitor pojamide on MDA-MB-231 triple-negative breast cancer Cells. Int. J. Mol. Sci. 2019, 20, 804. [Google Scholar] [CrossRef]
- Luparello, C.; Branni, R.; Abruscato, G.; Lazzara, V.; Drahos, L.; Arizza, V.; Mauro, M.; Di Stefano, V.; Vazzana, M. Cytotoxic capability and the associated proteomic profile of cell-free coelomic fluid extracts from the edible sea cucumber Holothuria tubulosa on HepG2 liver cancer cells. EXCLI J. 2022, 21, 722–743. [Google Scholar] [CrossRef]







{kind=link}
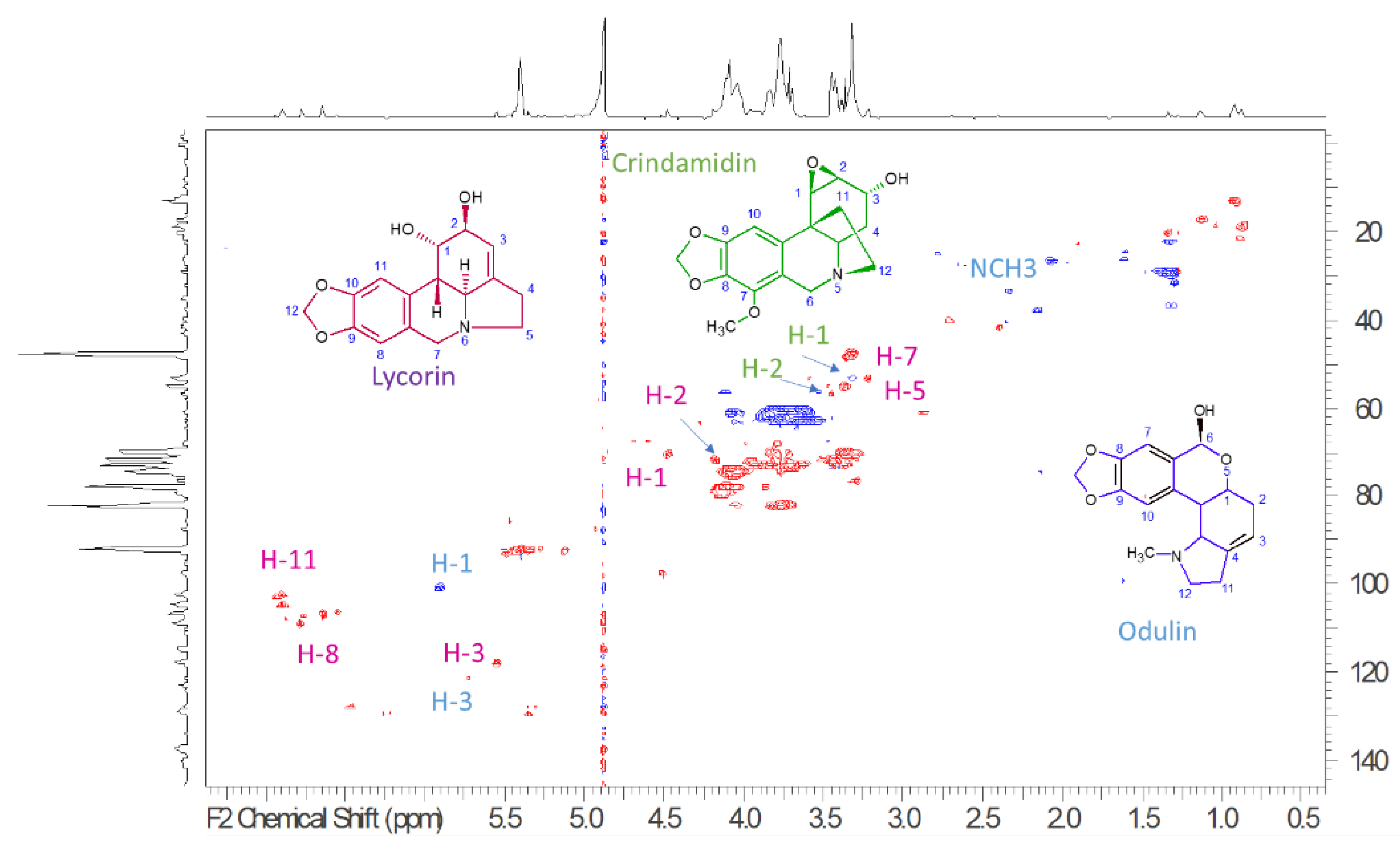{kind=link}
{kind=link}
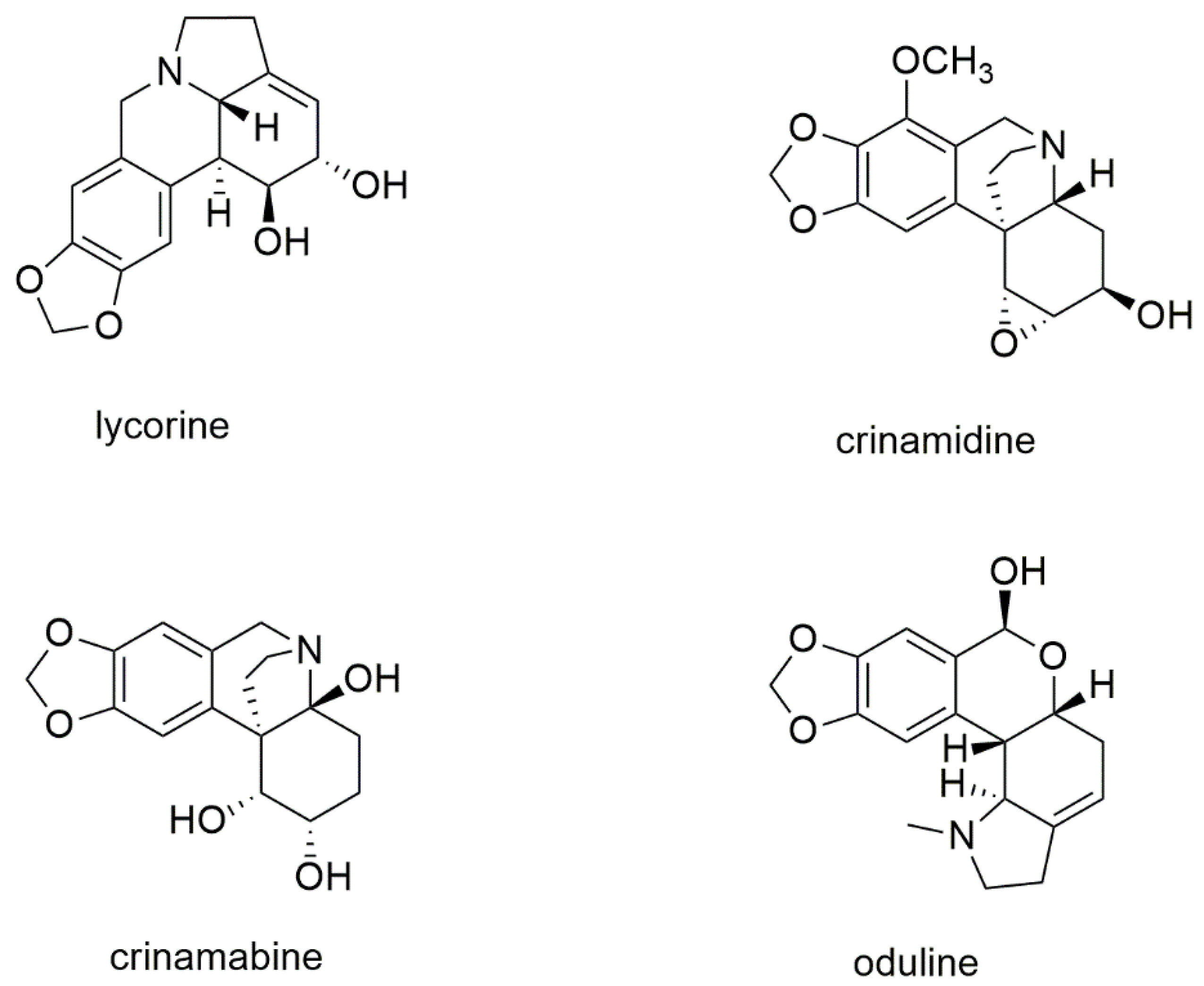{kind=link}
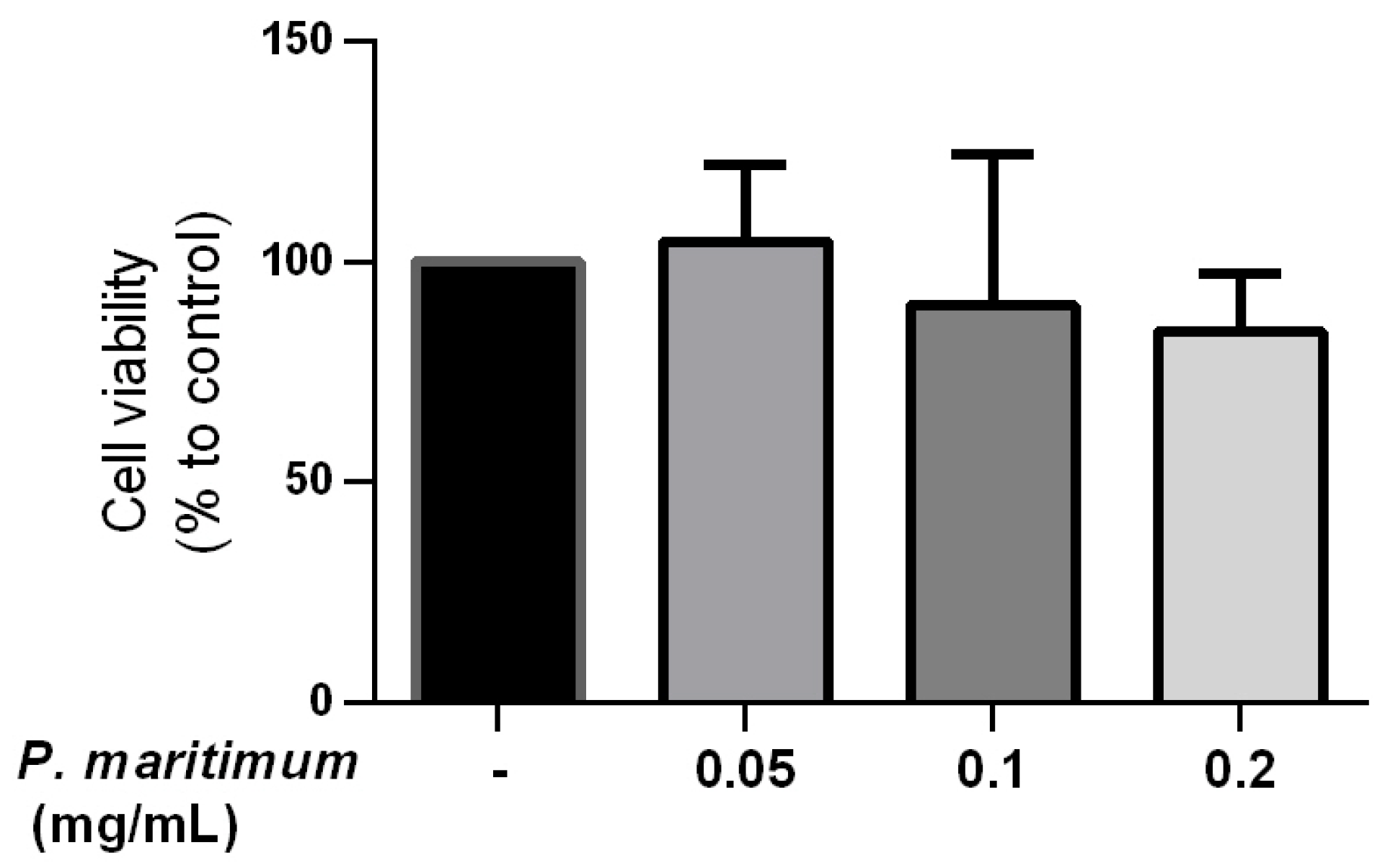{kind=link}
{kind=link}
| Origin | Alkaloids | Ref |
|---|---|---|
| Bulgaria | Trispheridine, graciline, galanthamine, buphanisine, N-demethylgalanthamine, crinine, demethylmarithidine, haemanthamine, tazettine, pancracine, lycorine | [17] |
| Caucasus | Tazettine, lycorine | [28] |
| Egypt | Narciclasine-4-O-β-glucopyranoside | [9] |
| Egypt | Ungeremine, zefbetaine, lycorine, tazettine, pancracine, lycorenine, galanthamine, sickenbergine, homolycorine, haemanthidine, hippadine, demethylhomolycorine, trispheridine, haemanthamine, pseudolycorine, 9-O-demethylhomolycorine, 11-hydroxyvittatine | [10] |
| Egypt | Ungeremine | [15] |
| Egypt | Pancrimatine A, pancrimatin B | [11] |
| Israel | Pancratistatin | [29] |
| Italy, Calabria | Lycorine, 9-O-demethyllycorine, haemanthidine, haemanthamine, 11-hydroxyvittatine, homolycorine, pancracine, obliquine, tazettine, vittatine. | [27] |
| Portugal | Haemanthidine, hippeastrine, lycorine, 11α-hydroxygalanthamine, 2α-10bα-dihydroxy-9-O-demethylhomolycorine, epi-galanthamine, 8-O-demethylhomolycorine, tazettine, haemanthamine | [24] |
| Spain | Ungiminorine N-oxide | [25] |
| Spain | 6-O-methylhaemanthidine, O,N-dimethylnorbelladine, lycorine, hippeastrine, galanthamine, haemanthamine, vittatine, 11-hidroxyvittatine, hordenine, 9-O-demethylhomolycorine, habranthine, ungiminorine, ungiminorine N-oxide | [26] |
| Tunisia | Narciclasine, lycorine, crinine (a), galanthamine, crinine (b), tazettine | [23] |
| Turkey | Lycorine, galanthamine, crinine, pancracine, 1-acetyl-β-carboline, 11,12-dehydrolycorene, galanthindole, 2,11-didehydro-2-dehydroxylycorine, assoanine, O-methylpretazettine, hordenine, 1-acetyl-β-carboline, 11,12-dehidrolycorene, buphanasine, 2,11-didehydro-2-dehydroxy-lycorine | [18] |
| Turkey | (−)-3β, 11α-dihroxy-1,2-dehydrocrinane, (−)-8-hydroxy-9-methoxycrinine | [19] |
| Turkey | Lycorine, (+)-demethylhomolycorine | [20] |
| Turkey | (−)-N-demethyl-galanthamine, (+)-tazetine, 2-O-demethylmontanine | [21] |
| tR (min) | M + H | Fragments | Identification | % a |
|---|---|---|---|---|
| 1.8 | 334 | 316 > 298–280 (265–221–249–237) | 2-hydroxy-3-dihydro-6-O-methyl-oduline | Traces |
| 1.9 | 288 | 270 > 252–222–237–178–149 | Lycorine | 3.18 ± 0.05 |
| 2.34 | 334 | 316 > 298–280 (265–221–249–237) | Isomer of the 2-hydroxy-3-dihydro-6-O-methyl-oduline | Traces |
| 2.4 | 288 | 270 > 252–222–237–178–149 | epi-Lycorine | 1.16 ± 0.02 |
| 3.33 | 302 | 284 266 255 193 | Oduline | 0.31 ± 0.02 |
| 3.9 | 288 | 270 > 252–222–237–178–149 | epi-Lycorine isomer 1 | Traces |
| 4.08 | 302 | 284 266 255 193 | Oduline isomer 1 | Traces |
| 4.4 | 288 | 270 > 252–222–237–178–149 | epi-Lycorine isomer 2 | Traces |
| 4.4 | 302 | 284 266 255 193 | Oduline isomer 2 | Traces |
| 4.85 | 494 | 462 492 360 314 300 227 211 | Unknown | - |
| 5.00 | 306 | 288 270 222 189 | epi-Crinamabine | 0.19 ± 0.01 |
| 5.6 | 318 | 300 286 268 250 227 | Crinamidine | 1.20 ± 0.03 |
| 6.05 | 302 | 270 266 252 227 211 182 | Methoxy lycorine | 1.86 ± 0.06 |
| 7.05 | 306 | 288 270 222 189 | Crinamabine | 0.96 ± 0.01 |
| 7.38 | 302 | 270 266 252 227 211 182 | Methoxyl lycorine isomer 1 | Traces |
| 7.84 | 302 | 270 266 252 227 211 182 | Methoxyl lycorine isomer 2 | Traces |
| Total- | 10.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cicio, A.; Sut, S.; Dall’Acqua, S.; Bruno, M.; Luparello, C.; Serio, R.; Zizzo, M.G. Chemical Characterization and Cytotoxic and Antioxidant Activity Evaluation of the Ethanol Extract from the Bulbs of Pancratium maritimun Collected in Sicily. Molecules 2023, 28, 3986. https://doi.org/10.3390/molecules28103986
Cicio A, Sut S, Dall’Acqua S, Bruno M, Luparello C, Serio R, Zizzo MG. Chemical Characterization and Cytotoxic and Antioxidant Activity Evaluation of the Ethanol Extract from the Bulbs of Pancratium maritimun Collected in Sicily. Molecules. 2023; 28(10):3986. https://doi.org/10.3390/molecules28103986
Chicago/Turabian StyleCicio, Adele, Stefania Sut, Stefano Dall’Acqua, Maurizio Bruno, Claudio Luparello, Rosa Serio, and Maria Grazia Zizzo. 2023. "Chemical Characterization and Cytotoxic and Antioxidant Activity Evaluation of the Ethanol Extract from the Bulbs of Pancratium maritimun Collected in Sicily" Molecules 28, no. 10: 3986. https://doi.org/10.3390/molecules28103986
APA StyleCicio, A., Sut, S., Dall’Acqua, S., Bruno, M., Luparello, C., Serio, R., & Zizzo, M. G. (2023). Chemical Characterization and Cytotoxic and Antioxidant Activity Evaluation of the Ethanol Extract from the Bulbs of Pancratium maritimun Collected in Sicily. Molecules, 28(10), 3986. https://doi.org/10.3390/molecules28103986