
Biostimulant Formulations and Moringa oleifera Extracts to Improve Yield, Quality, and Storability of Hydroponic Lettuce

 , ,
, ,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
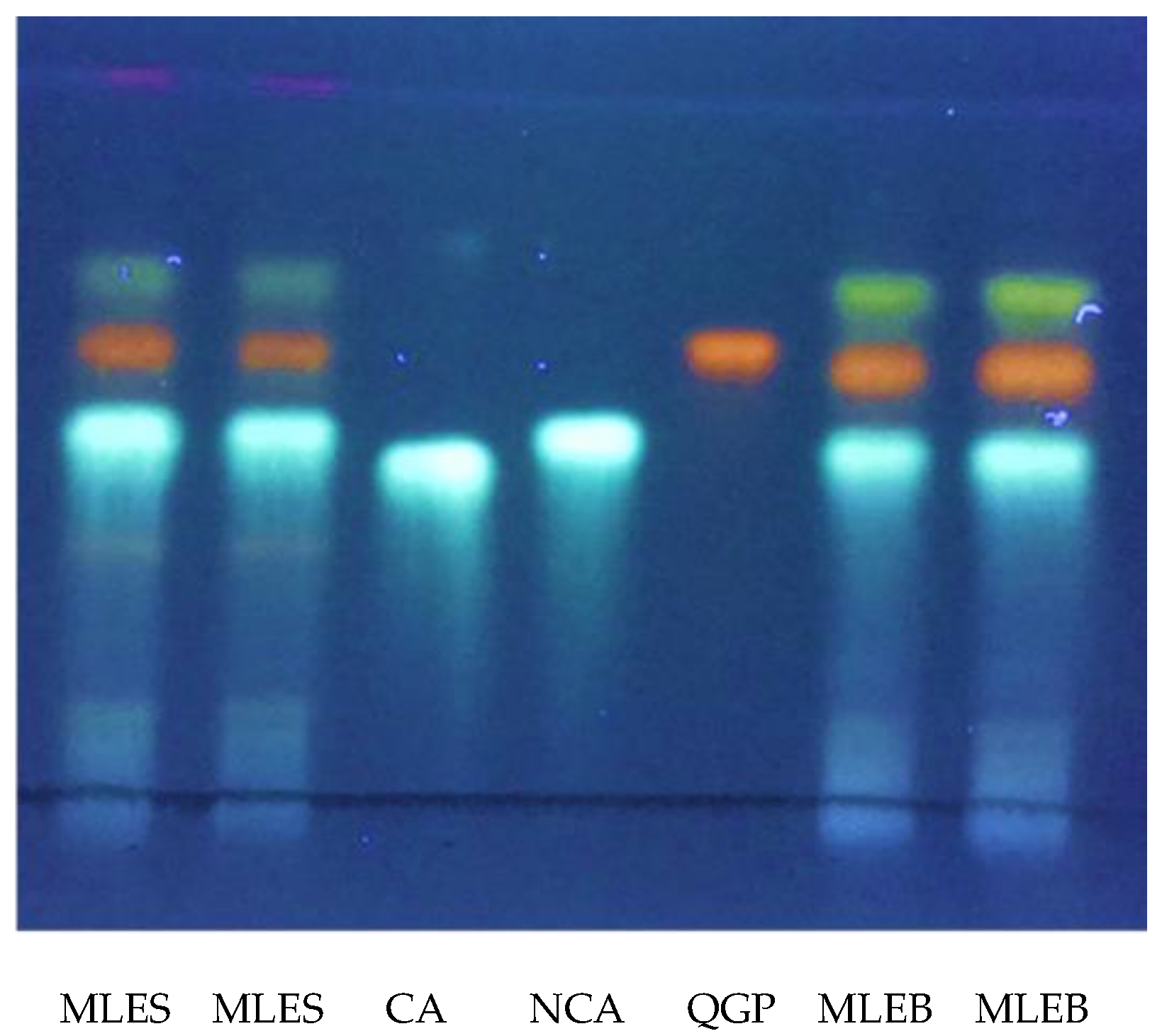
2.1. Qualitative and Quantitative Characterization of MLEs
2.2. Yield, Morphological Parameter Measurements and Quality Analysis Samplings
2.3. Leaf Color, SPAD Index, and Pigment Contents
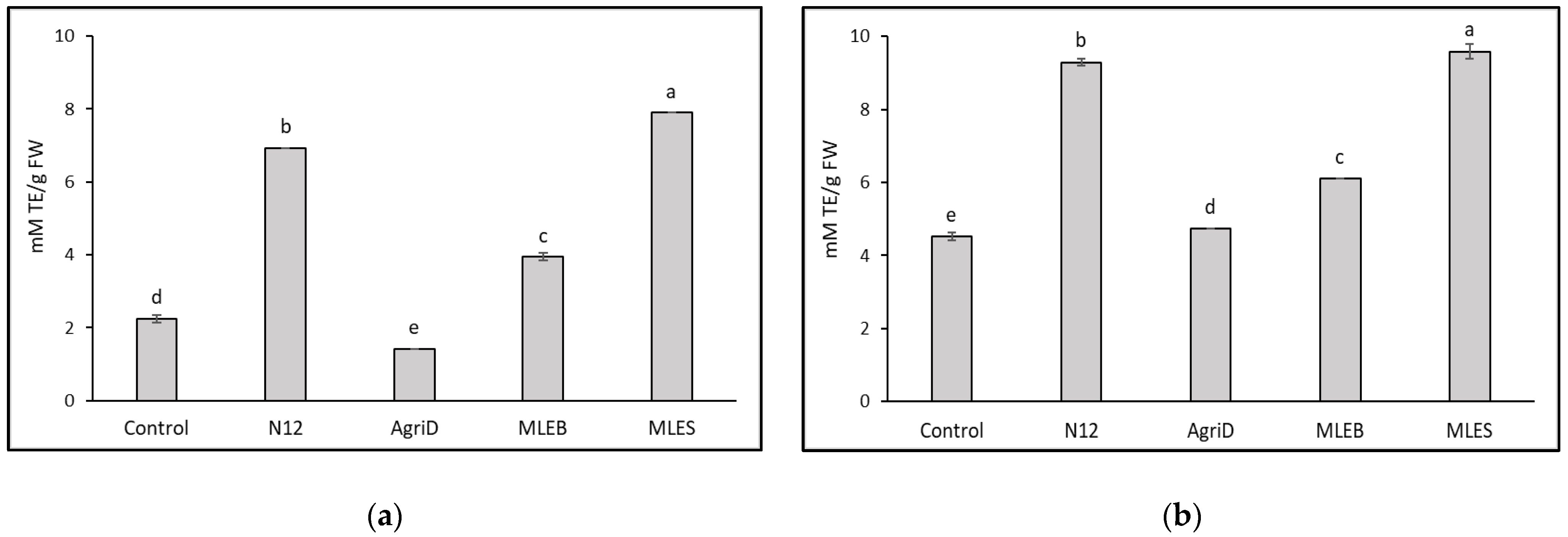
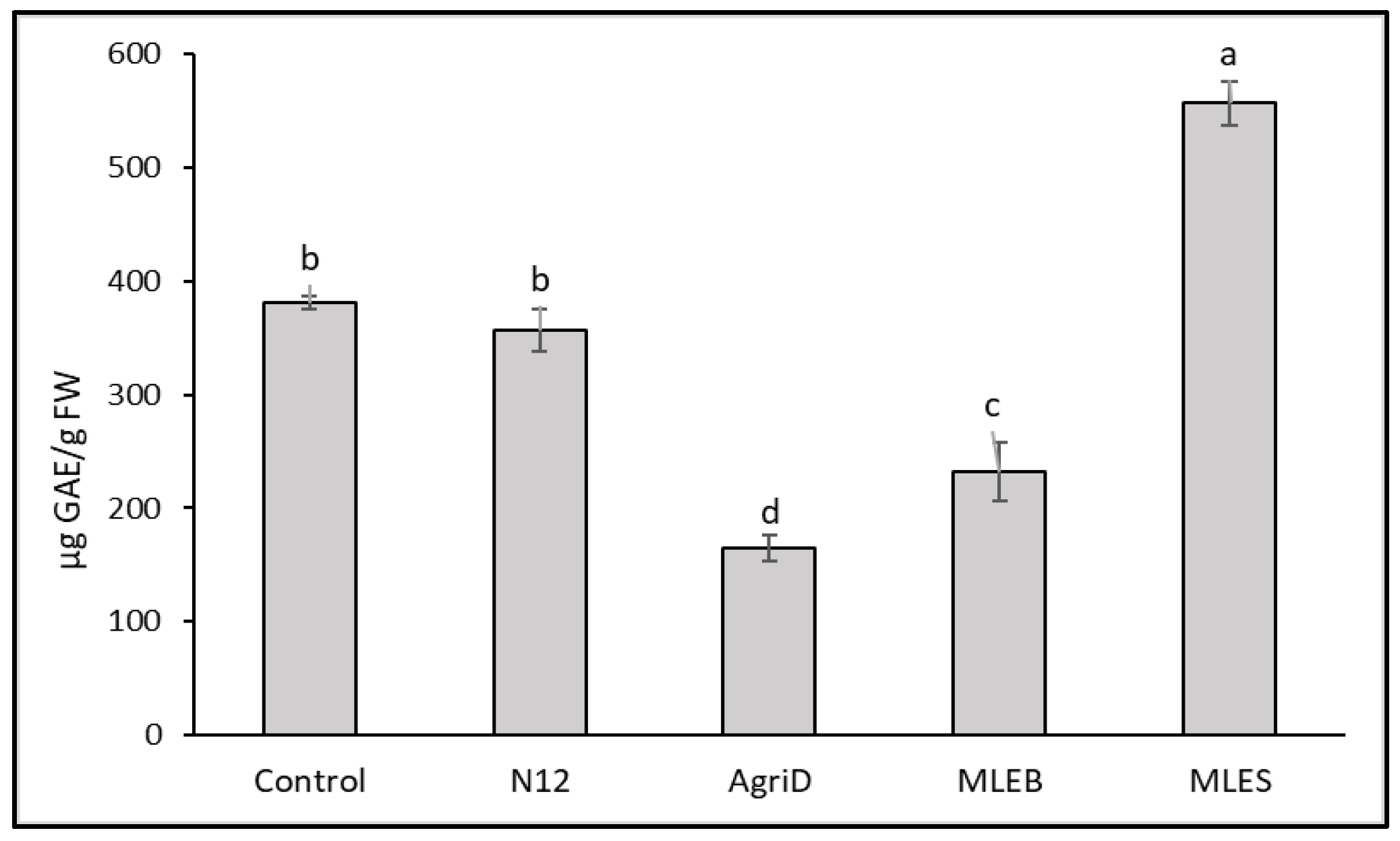
2.4. Total Phenolic Compounds and Antioxidant Activities
2.5. Resistance to Postharvest Grey Mold
3. Materials and Methods
3.1. Lettuce Growth Conditions, Experimental Design, and Treatments
3.2. Moringa Leaves Extract Preparation and Characterization
3.3. Yield and Agronomic Parameters
3.4. Leaf Colour, SPAD Index, and Pigment Content
3.5. Total Phenolic Compounds and Antioxidant Activities
3.6. Resistance to Postharvest Grey Mold
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Mir, S.A.; Shah, M.A.; Mir, M.M.; Dar, B.N.; Greiner, R.; Roohinejad, S. Microbiological contamination of ready-to-eat vegetable salads in developing countries and potential solutions in the supply chain to control microbial pathogens. Food Cont. 2018, 85, 235–244. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL/ (accessed on 28 December 2022).
- Kim, M.J.; Moon, Y.; Kopsell, D.A.; Park, S.; Tou, J.C.; Waterland, N.L. Nutritional value of crisphead ‘Iceberg’and romaine lettuces (Lactuca sativa L.). J. Agric. Sci. 2016, 8, 1–10. [Google Scholar]
- Casati, D.; Baldi, L. L’importanza economica del comparto della IV gamma. In Le Avversità Degli Ortaggi da Foglia Per la IV Gamma; Sannino, L., Espinosa, B., Eds.; TerraOrti: Eboli, Italy, 2016; pp. 19–31. [Google Scholar]
- Searchinger, T.D. Synergies and Tradeoffs for Small Farmers and Climate Mitigation; OXFAM America Research Background Series; OXFAM America: Boston, MA, USA, 2013; pp. 1–75. [Google Scholar]
- Wheeler, T.; Von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef]
- Al-Chalabi, M. Vertical farming: Skyscraper sustainability? Sustain. Cities Soc. 2015, 18, 74–77. [Google Scholar] [CrossRef]
- Corbo, M.; Campaniello, D.; Speranza, B.; Bevilacqua, A.; Sinigaglia, M. Non-conventional tools to preserve and prolong the quality of minimally-processed fruits and vegetables. Coatings 2015, 5, 931–961. [Google Scholar] [CrossRef]
- European Biostimulant Industry Council. Available online: www.biostimulants.eu (accessed on 20 October 2022).
- Mattner, S.W.; Wite, D.; Riches, D.A.; Porter, I.J.; Arioli, T. The effect of kelp extract on seedling establishment of broccoli on contrasting soil types in southern Victoria, Australia. Biol. Agric. Hort. 2013, 29, 258–270. [Google Scholar] [CrossRef]
- Dodd, I.C.; Ruiz-Lozano, J.M. Microbial enhancement of crop resource use efficiency. Curr. Opin. Biotechnol. 2012, 23, 236–242. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hort. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Barbosa, G.; Gadelha, F.; Kublik, N.; Proctor, A.; Reichelm, L.; Weissinger, E.; Wohelleb, G.M.; Halden, R. Comparison of land, water and energy requirement of lettuce grown using hydroponic vs. conventional agricultural methods. Int. J. Environ. Res. Public Health 2015, 12, 6879–6891. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Petropoulos, S.A.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; Colla, G.; Troise, A.D.; Vitaglione, P.; De Pascale, S.; Rouphael, Y. The bioactive profile of lettuce produced in a closed soilless system as configured by combinatorial effects of genotype and macrocation supply composition. Food Chem. 2020, 309, 125713. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, I.; La Bella, E.; Rovetto, E.I.; Lo Piero, A.R.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Lisiecka, J.; Knaflewski, M.; Spizewski, T.; Fraszczak, B.; Kaluzewicz, A.; Krzesinski, W. The effect of animal protein hydrolysate on quantity and quality of strawberry daughter plants cv. Elsanta. Acta Sci. Pol. Hortorum Cultus 2011, 10, 31–40. [Google Scholar]
- Kurepin, L.V.; Zaman, M.; Pharis, R.P. Phytohormonal basis for the plant growth promoting action of naturally occurring biostimulators. J. Sci. Food Agric. 2014, 94, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Gomiero, T. Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An overview of plant-based natural biostimulants for sustainable horticulture with a particular focus on moringa leaf extracts. Plant Sci. 2020, 295, 110194. [Google Scholar] [CrossRef]
- Younis, A.; Akhtar, M.S.; Riaz, A.; Zulfiqar, F.; Qasim, M.; Farooq, A.; Tariq, U.; Ahsan, M.; Bhatti, Z.M. Improved cut flower and corm production by exogenous moringa leaf extract application on gladiolus cultivars. Acta Sci. Pol. Hortorum Cultus 2018, 17, 25–38. [Google Scholar] [CrossRef]
- Merwad, A.R.M. Using Moringa oleifera extract as biostimulant enhancing the growth, yield and nutrients accumulation of pea plants. J. Plant Nutr. 2018, 41, 425–431. [Google Scholar] [CrossRef]
- Nasir, M.; Khan, A.S.; Basra, S.A.; Malik, A.U. Foliar application of moringa leaf extract, potassium and zinc influence yield and fruit quality of ‘Kinnow’ mandarin. Sci. Hort. 2016, 210, 227–235. [Google Scholar] [CrossRef]
- Saini, R.K.; Sivanesan, I.; Keum, Y.S. Phytochemicals of Moringa oleifera: A review of their nutritional, therapeutic and industrial significance. 3 Biotech 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Howladar, S.M. A novel Moringa oleifera leaf extract can mitigate the stress effects of salinity and cadmium in bean (Phaseolus vulgaris L.) plants. Ecotox. Environ. Saf. 2014, 100, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Latif, H.H.; Mohamed, H.I. Exogenous applications of moringa leaf extract effect on retrotransposon, ultrastructural and biochemical contents of common bean plants under environmental stresses. S. Afr. J. Bot. 2016, 106, 221–231. [Google Scholar] [CrossRef]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa oleifera: A review on nutritive importance and its medicinal application. Food Sci. Hum. Wellness 2016, 5, 49–56. [Google Scholar] [CrossRef]
- Yan, G.; Liping, S.; Yongliang, Z. UPLC-Q-Orbitrap-MS2 analysis of Moringa oleifera leaf extract and its antioxidant, antibacterial and anti-inflammatory activities. Nat. Prod. Res. 2020, 34, 2090–2094. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Quirantes-Piné, R.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Optimization of extraction method to obtain a phenolic compounds-rich extract from Moringa oleifera Lam leaves. Ind. Crop. Prod. 2015, 66, 246–254. [Google Scholar] [CrossRef]
- Xu, Y.-B.; Chen, G.-L.; Guo, M.-Q. Antioxidant and Anti-Inflammatory Activities of the Crude Extracts of Moringa oleifera from Kenya and Their Correlations with Flavonoids. Antioxidants 2019, 8, 296. [Google Scholar] [CrossRef]
- Gao, Q.; Wei, Z.; Liu, Y.; Wang, F.; Zhang, S.; Serrano, C.; Li, L.; Sun, B. Characterization, Large-Scale HSCCC Separation and Neuroprotective Effects of Polyphenols from Moringa oleifera Leaves. Molecules 2022, 27, 678. [Google Scholar] [CrossRef]
- Herrmann, K. On the occurrence of flavonol and flavone glycosides in vegetables. Z. Lebens. Unters. Forsch 1988, 186, 1–5. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Schena, L.; De Girolamo, A.; Ippolito, A.; González-Candelas, L. Characterization of genes associated with induced resistance against Penicillium expansum in apple fruit treated with quercetin. Postharvest Biol. Technol. 2010, 56, 1–11. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Dewang, S.P. Influence of Soil-application of Fish-protein Hydrolysate Liquid on Growth and Yield of Spinach (Spinacia oleracea L.). Asian J. Dairy Food Res. 2021, 40, 69–75. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hort. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Boselli, M.; Bahouaoui, M.A.; Lachhab, N.; Sanzani, S.M.; Ferrara, G.; Ippolito, A. Protein hydrolysates effects on grapevine (Vitis vinifera L.; cv. Corvina) performance and water stress tolerance. Sci. Hort. 2019, 258, 108784. [Google Scholar] [CrossRef]
- Taha, M.A.; AM, A.E.A.; El-Shennawy, M.Z. Effect of some Plant Aqueous Extracts on Lettuce Growth, Chemical Constituents, Yield and Downy Mildew Disease. J. Plant Prod. 2020, 11, 933–938. [Google Scholar]
- Lachhab, N.; Sanzani, S.M.; Bahouaoui, M.A.; Boselli, M.; Ippolito, A. Effect of some protein hydrolysates against gray mould of table and wine grapes. Eur. J. Plant Pathol. 2016, 144, 821–830. [Google Scholar] [CrossRef]
- Samanta, A.; Das, G.; Das, S.K. Roles of flavonoids in plants. Carbon 2011, 100, 12–35. [Google Scholar]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-based biostimulants influence the agronomical, physiological, and qualitative responses of baby rocket leaves under diverse nitrogen conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Drench application of fish-derived protein hydrolysates affects lettuce growth, chlorophyll content, and gas exchange. Horttechnology 2017, 27, 539–543. [Google Scholar] [CrossRef]
- Rizwan, A.; Bushra, S.; Munawar, I.; Muhammad, M. Variation in biochemical and antioxidant attributes of Raphanus sativus in response to foliar application of plant leaf extracts as plant growth regulator. J. Genetic Eng. Biotechnol. 2016, 14, 1–8. [Google Scholar]
- Lachhab, N.; Sanzani, S.M.; Adrian, M.; Chiltz, A.; Balacey, S.; Boselli, M.; Ippolito, A.; Poinssot, B. Soybean and casein hydrolysates induce grapevine immune responses and resistance against Plasmopara viticola. Front. Plant Sci. 2014, 5, 716. [Google Scholar] [CrossRef] [PubMed]
- Lachhab, N.; Sanzani, S.M.; Fallanaj, F.; Youssef, K.; Nigro, F.; Boselli, M.; Ippolito, A. Protein hydrolysates as resistance inducers for controlling green mould of citrus fruit. Acta Hortic. 2015, 1065, 1593–1598. [Google Scholar] [CrossRef]
- Hojati, M.; Modarres-Sanavy, S.A.M.; Ghanati, F.; Panahi, M. Hexaconazole induces antioxidant protection and apigenin-7-glucoside accumulation in Matricaria chamomilla plants subjected to drought stress. J. Plant Physiol. 2011, 168, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Vàsquez, H.; Ouhibi, C.; Lizzi, Y.; Azzouz, N.; Forges, M.; Bardin, M.; Nicot, P.; Urban, L.; Aarrouf, J. Pre-harvest hormetic doses of UV-C radiation can decrease susceptibility of lettuce leaves (Lactuca sativa L.) to Botrytis cinerea L. Sci. Hort. 2017, 222, 32–39. [Google Scholar] [CrossRef]
- Roselli, M.; Lovece, A.; Bruno, C.; Cavalluzzi, M.M.; Laghezza, A.; Mercurio, A.; Lentini, G.; Corbo, F.; la Forgia, F.; Fontana, S.; et al. Antioxidant activity of Uva di Troia Canosina: Comparison of two extraction methods. Clin. Immunol. Endocr. Metab. Drugs 2015, 2, 8–12. [Google Scholar] [CrossRef]
- Caputo, L.; Quintieri, L.; Cavalluzzi, M.M.; Lentini, G.; Habtemariam, S. Antimicrobial and antibiofilm activities of citrus water-extracts obtained by microwave-assisted and conventional methods. Biomedicines 2018, 6, 70. [Google Scholar] [CrossRef]
- Milani, G.; Curci, F.; Cavalluzzi, M.M.; Crupi, P.; Pisano, I.; Lentini, G.; Clodoveo, M.L.; Franchini, C.; Corbo, F. Optimization of microwave-assisted extraction of antioxidants from bamboo shoots of Phyllostachys pubescens. Molecules 2020, 25, 215. [Google Scholar] [CrossRef]
- Cavalluzzi, M.M.; Lamonaca, A.; Rotondo, N.P.; Miniero, D.V.; Muraglia, M.; Gabriele, P.; Corbo, F.; De Palma, A.; Budriesi, R.; De Angelis, E.; et al. Microwave-Assisted Extraction of Bioactive Compounds from Lentil Wastes: Antioxidant Activity Evaluation and Metabolomic Characterization. Molecules 2022, 27, 7471. [Google Scholar] [CrossRef]
- Cristofano, F.; El-Nakhel, C.; Pannico, A.; Giordano, M.; Colla, G.; Rouphael, Y. Foliar and root applications of vegetal-derived protein hydrolysates differentially enhance the yield and qualitative attributes of two lettuce cultivars grown in floating system. Agronomy 2021, 11, 1194. [Google Scholar] [CrossRef]
- Tarantino, A.; Difonzo, G.; Lopriore, G.; Disciglio, G.; Paradiso, V.M.; Gambacorta, G.; Caponio, F. Bioactive compounds and quality evaluation of ‘Wonderful’pomegranate fruit and juice as affected by deficit irrigation. J. Sci. Food Agri. 2020, 100, 5539–5545. [Google Scholar] [CrossRef] [PubMed]
- Difonzo, G.; Aresta, A.; Cotugno, P.; Ragni, R.; Squeo, G.; Summo, C.; Massari, F.; Pasqualone, A.; Faccia, M.; Zambonin, C.; et al. Supercritical CO2 extraction of phytocompounds from olive pomace subjected to different drying methods. Molecules 2021, 26, 598. [Google Scholar] [CrossRef] [PubMed]
- Bardin, M.; Comby, M.; Lenaerts, R.; Nicot, P.C. Diversity in susceptibility of Botrytis cinerea to biocontrol products inducing plant defence mechanisms. IOBC-WPRS Bull. 2013, 88, 45–49. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Experimental Mass | Calculated Mass | Key Fragments (m/z) | Error (ppm) a | Molecular Formula | Reference |
|---|---|---|---|---|---|---|
| Neochlorogenic acid | 353.0895 | 353.0878 | 135.0449 (100), 179.0346 (27.0), 191.0557 (80.9) | 3.4 | C16H18O9 | [30,31] |
| Quercetin 3-O-β-d-glucopyranoside | 463.0886 | 463.0880 | 271.0249 (22.3), 300.0300 (100), 301.0354 (52.5) | 1.3 | C21H20O12 | [30,31] |
| Kaempferol acetylglucoside | 489.1046 | 489.1037 | 284.0336 (100), 285.0356 (78.6) | 1.8 | C23H22O12 | [30] |
| Quercetin acetylglucoside | 505.100 | 505.0988 | 271.0260 (21.9), 300.0294 (100), 301.0358 (55.6) | 2.4 | C23H22O13 | [30,31] |
| Quercetin malonylglucoside | 549.0905 | 549.0886 | 300.0271 (100), 301.0335 (56.7) | 3.5 | C24H22O15 | [30] |
| Glucomoringin | 570.0972 | 570.0957 | 96.9612 (100), 259.0144 (6.8), 328.0879 (7.1) | 2.6 | C20H29NO14S2 | [30,31,32] |
| Acetyl glucomoringin | 612.1075 | 612.1062 | 96.9612 (100), 259.0144 (9.59) | 2.1 | C22H31NO15S2 | [31] |
| Compound | Experimental mass | Calculated mass | Key fragments (m/z) | Error (ppm) a | Molecular formula | Reference |
|---|---|---|---|---|---|---|
| Neochlorogenic acid | 353.089 | 353.0878 | 135.0465 (100), 179.0379 (27.57), 191.0578 (91.93) | 3.4 | C16H18O9 | [30,31] |
| Quercetin 3-O-β-d-glucopyranoside | 463.0870 | 463.0880 | 271.0244 (18.02), 300.0266 (100), 301.0330 (50.7), | –2.2 | C21H20O12 | [30,31] |
| Glucomoringin | 570.0955 | 570.0957 | 96.9604 (100), 259.0140 (10.33), 328.0866 (8.49 | –0.3 | C20H29NO14S2 | [30,31,32] |
| Acetyl glucomoringin | 612.1068 | 612.1062 | 96.9606 (100), 259.0118 (7.73) | 1.0 | C22H31NO15S2 | [31] |
| Treatments | Marketable Yield (g/m) | Head Fresh Weight (g) | Leaf Area (cm2) | Leaf Number | Water Content (%) | Dry Matter (%) |
|---|---|---|---|---|---|---|
| Control | 799.52 | 53.30 ± 26.07 b | 130.44 ± 40.15 c | 21.40 ± 2.19 b | 90.63 ± 0.37 ab | 9.37 ± 0.37 ab |
| N12 | 818.79 | 54.59 ± 28.73 b | 150.73 ± 44.49 ab | 20.80 ± 2.05 b | 89.35 ± 0.48 b | 10.65 ± 0.48 a |
| AgriD | 1280.18 | 85.30 ± 46.80 a | 168.21 ± 32.92 a | 23.40 ± 1.14 ab | 91.78 ± 1.31 a | 8.22 ± 1.31 b |
| MLEB | 1144.91 | 76.30 ± 43.10 ab | 151.60 ± 33.08 ab | 22.40 ± 1.82 b | 92.20 ± 0.44 a | 7.80 ± 0.44 b |
| MLES | 999.90 | 66.66 ± 24.19 ab | 134.59 ± 26.47 bc | 25.40 ± 2.70 a | 90.78 ± 1.82 ab | 9.22 ± 1.82 ab |
| Treatments | L* (lightness) | C* (Chroma Value) | h° (Hue Angle) | SPAD Index | Tot Chlorophyll (a + b) (µg/g FW) | Carotenoid Contents (µg/g FW) |
|---|---|---|---|---|---|---|
| Control | 57.91 ± 3.42 ab | 35.74 ± 2.64 b | 178.77 ± 0.03 b | 28.58 ± 0.63 b | 19.21 ± 0.18 d | 4.29 ± 0.06 c |
| N12 | 57.21 ± 2.61 c | 31.99 ± 2.69 d | 178.79 ± 0.02 a | 31.76 ± 1.96 a | 20.45 ± 0.06 c | 4.87 ± 0.02 b |
| AgriD | 56.34 ± 2.20 b | 34.51 ± 1.94 c | 178.78 ± 0.01 a | 29.84 ± 1.40 ab | 22.51 ± 0.04 a | 5.13 ± 0.03 a |
| MLEB | 56.09 ± 1.94 b | 34.68 ± 2.22 bc | 178.78 ± 0.01 a | 30.00 ± 1.84 ab | 21.40 ± 0.10 b | 4.83 ± 0.01 b |
| MLES | 53.30 ± 2.50 a | 36.96 ± 2.31 a | 178.76 ± 0.02 b | 30.32 ± 1.01 ab | 12.64 ± 0.14 e | 3.24 ± 0.17 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Admane, N.; Cavallo, G.; Hadjila, C.; Cavalluzzi, M.M.; Rotondo, N.P.; Salerno, A.; Cannillo, J.; Difonzo, G.; Caponio, F.; Ippolito, A.; et al. Biostimulant Formulations and Moringa oleifera Extracts to Improve Yield, Quality, and Storability of Hydroponic Lettuce. Molecules 2023, 28, 373. https://doi.org/10.3390/molecules28010373
Admane N, Cavallo G, Hadjila C, Cavalluzzi MM, Rotondo NP, Salerno A, Cannillo J, Difonzo G, Caponio F, Ippolito A, et al. Biostimulant Formulations and Moringa oleifera Extracts to Improve Yield, Quality, and Storability of Hydroponic Lettuce. Molecules. 2023; 28(1):373. https://doi.org/10.3390/molecules28010373
Chicago/Turabian StyleAdmane, Naouel, Giuseppe Cavallo, Chahinez Hadjila, Maria Maddalena Cavalluzzi, Natalie Paola Rotondo, Antonio Salerno, Joseph Cannillo, Graziana Difonzo, Francesco Caponio, Antonio Ippolito, and et al. 2023. "Biostimulant Formulations and Moringa oleifera Extracts to Improve Yield, Quality, and Storability of Hydroponic Lettuce" Molecules 28, no. 1: 373. https://doi.org/10.3390/molecules28010373
APA StyleAdmane, N., Cavallo, G., Hadjila, C., Cavalluzzi, M. M., Rotondo, N. P., Salerno, A., Cannillo, J., Difonzo, G., Caponio, F., Ippolito, A., Lentini, G., & Sanzani, S. M. (2023). Biostimulant Formulations and Moringa oleifera Extracts to Improve Yield, Quality, and Storability of Hydroponic Lettuce. Molecules, 28(1), 373. https://doi.org/10.3390/molecules28010373