Abstract
Candida albicans infections are related to biofilm formation. The increase in antifungal resistance and their adverse effects have led to the search for therapeutic options as plant derivatives. This scoping review aims to identify the current status of in vitro research on the cytotoxicity and inhibitory effects of plant derivatives on C. albicans biofilms. In this study, PRISMA items were followed. After recognition of the inclusion criteria, full texts were read and disagreements were resolved with a third party. A risk of bias assessment was performed, and information was summarized using Microsoft Office Excel. Thirty-nine papers fulfilling the selection criteria were included. The risk of bias analysis identified most of the studies as low risk. Studies evaluated plant derivatives such as extracts, essential oils, terpenes, alkaloids, flavonoids and polyphenols. Some studies evaluated the inhibition of C. albicans biofilm formation, inhibition on preformed biofilms or both. The derivatives at concentrations greater than or equal to those that have an inhibitory effect on C. albicans biofilms, without showing cytotoxicity, include magnoflorin, ellagic acid, myricetin and eucarobustol from Eucalyptus robusta and, as the works in which these derivatives were studied are of good quality, it is desirable to carry out study in other experimental phases, with methodologies that generate comparable information.
1. Introduction
Candida albicans is an opportunistic fungus that can cause superficial and systemic infections in individuals when mucosal barriers are disrupted, or when the immune system is compromised [1]. Some factors associated with candidemia in intensive care units (ICU) are long durations of central venous catheterization, urinary tract catheterization and mechanical ventilation [2,3]. At low levels, Candida albicans colonizes oral mucosal surfaces [4] as a normal inhabitant, but, under certain circumstances, it can cause a superficial candidiasis characterized by the appearance of white plaques on inflamed and red mucosa and by pain when eating or swallowing [4,5]. If C. albicans infection becomes invasive, it can cause septicemia [6]. C. albicans has also been reported to infect oral mucositis lesions [7,8], causing inflammation of the oropharyngeal mucosa [9,10]. Patients who suffer from cancer affecting the head and neck, and who receive chemotherapy and radiotherapy treatments, are almost all affected by oral mucositis [9,10].
C. albicans infections are related to several virulence factors, including biofilm formation (BF) on implanted medical devices and oral mucosa (biotic and abiotic surfaces) [4,5,6], which allow the initial adhesion to epithelial cells with subsequent tissue invasion, damage and antimicrobial resistance increased risks [5,11,12]. Further, C. albicans virulence factors include its ability to switch from the yeast form to an invasive hyphae morphotype, and to secrete proteolytic and lipolytic enzymes [13].
Given its ability to grow in biofilms, C. albicans can tolerate higher concentrations of antimicrobials, which has an important impact on public health [14], and has led to an increase in research on drug resistance [15] and therapeutic options. Essential oils, extracts and other plant derivatives are among the most evaluated alternatives, being an attractive option as they provide the possibility of achieving better therapeutic effects with less toxicity and due to their multiple possibilities of use: systemic, topical and as antiseptic in abiotic surfaces.
On the other hand, variations in the techniques for studying cytotoxicity and antifungal capacity make it difficult to identify, in the literature, the plant derivatives with the best level of progress and scientific evidence that supports a possible application.
In this scoping review, we provide an overview of the antifungal C. albicans biofilms and the cytotoxic effect of plant derivatives, such as essential oils, extracts and terpenes, placing a particular emphasis on the in vitro information with good quality, the most common used technics and evaluation protocols.
2. Results
2.1. Essential Oils and Composition
The flow diagram of the screened manuscripts (Figure 1) shows a total of 1049 potentially eligible studies following the electronic screening strategy search. Reviewer agreement led to the elimination of 851 articles that did not conduct an evaluation of the effect of plant derivatives on C. albicans biofilms. After removing duplicates, 181 articles were reviewed, resulting in 120 non-eligible studies being excluded at this stage for not having antifungal evaluation at the planktonic level or cytotoxicity assays. Two articles without access to the full text were removed and, finally, fifty-nine full-text articles were reviewed, out of which thirty-nine studies fulfilled all of the selection criteria and were included in the full data analysis. All the included studies were published over a twelve-year period (from 2010 to 2022), with the period from 2014 to 2017 having the highest number of publications (46%).
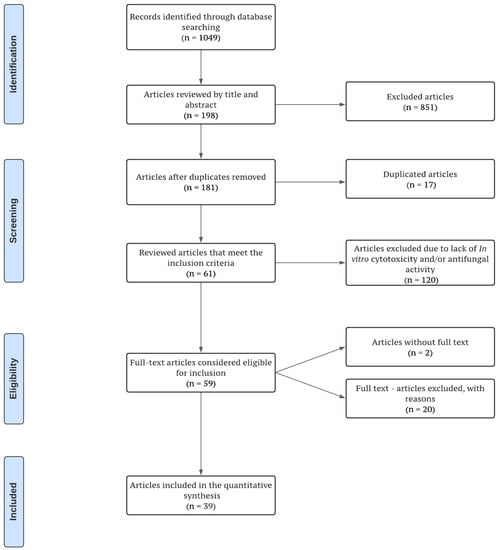
Figure 1.
Screening and selection process flow chart, according to the PRISMA statement.
Publications from five continents were identified; Brazil was the country with the most papers (36%). Two articles did not report funding entities, while thirty reported public funding and seven reported private funding. Some studies evaluated other aspects of C. albicans biofilms separately from the ones of the inclusion criteria, such as adhesion capacity (31%), yeast-hyphal transition (46%) and visualization by SEM/CLSM/fluorescence microscope (44%) (Table 1).

Table 1.
Additional study variables to those included in the inclusion criteria and funding sources.
2.2. Risk of Bias Assessment
All included studies were assessed for the risk of bias. Out of the thirty-nine articles, no study presented a high risk of bias, thirty articles showed low risk and nine studies had a medium risk of bias. Item 4 (presence of die control in cytotoxic activity) was the least reported (23%), while items 1 and 3 were reported by 97% and 92% of the studies, respectively. Thirty-five studies mentioned the number of replicates or repetitions of experiments, which is important to validate data. Complete scores of all items are described in Table 2, according to the parameters considered in the analysis.
Table 2.
Risk of bias assessment.
2.3. Type of Plant Derivatives
The studies evaluated different plant derivatives: extracts from plants were the most studied derivatives (thirteen articles), followed by essential oils (six studies) and terpenes (six studies). Other compounds evaluated were alkaloids, flavonoids, rich fractions, polyphenols and naphthols.
The majority of the studies reported the anti-C. albicans biofilm activity and cytotoxic effect of one plant derivative (26/39 articles), while thirteen studies evaluated at least two plant derivatives; one study evaluated seven extracts of Casearia sylvestris [19]. Three studies evaluated at least one sample of plant extracts and other compounds of a different chemical nature, such as essential oils [16], phenols [24] and fractions [54]. Additionally, one article evaluated flavonoids and diterpenes [46].
2.4. Cell Types in the Research Studies
Cytotoxic activity was performed on human cells in 67% of the studies, while 33% used cells derived from other animals. Most of the studies analyzed the plant derivatives’ activity in cell lines (twenty-six studies), whilst ten studies used primary cells and three studies examined bank and primary cells.
Epidermal cells were examined by seventeen studies, and eight studies evaluated cytotoxicity on macrophages. The most used cell line was murine macrophage cells (RAW 264.7) (6/40 articles), followed by human umbilical vein endothelial cells (HUVEC) (5/40 articles), human keratinocytes cells (HaCaT) (4/40 articles) and mouse fibroblast cells (L929) (4/40 articles). In one study, the hemolytic effect on human erythrocytes from a healthy person was evaluated [25].
Two studies [29,42], those of Ma et al. (2015) and Oliveira et al. (2017), analyzed the effect of plant derivatives on the viability of four different cell types, but most studies (28/39) evaluated the cytotoxicity over one cell line (Table 3).
Table 3.
Principal findings of the cytotoxic effect of plant derivatives.
2.5. C. albicans Strain
We can identify the use of thirteen different C. albicans reference strains in the thirty-nine studies included in this review. All studies examined at least one reference strain, the most used ones being SC 5314 (eleven studies), ATCC 10,231 (twelve studies), ATCC 90,028 (six studies) and ATCC 18,804 (four studies). Six studies used two or more strains, and four studies always jointly included clinical isolates with one or more reference strains (Table 4).
Table 4.
Principal findings of studies that investigated the effect of plant derivatives on C. albicans biofilm.
Sharma et al. (2019) and Kim et al. (2017) [39,44] included an experimental design with a sensitive and fluconazole-resistant strain. In the study of Souza et al. (2018) [28], nine clinical strains from kidney transplant patients were included, whereas the study of Sudjana et al. (2012) [37] included seven clinical isolates without data on the characteristics of the patients from which they were obtained. Ivanov et al. (2021) [51] used three clinical isolates from the oral cavities of patients from the Clinical Hospital Center Zvezdara (ENT clinic.), and Curvelo et al. (2014) [38] used one clinical strain from the oral mucosa of human-immunodeficiency-virus-positive pediatric patients resistant to fluconazole (Table 4).
2.6. Cell Cytotoxicity
Various assays were used to determine the cytotoxicity of plant derivatives. A total of 72% of the included articles used the tetrazolium assay (MTT), followed by Sulforhodamine B assay (1%). Other evaluation techniques consisted of a cytotoxicity detection kit, neutral red dye method, resazurin and Alamar blue®.
Different schemes of exposure times to plant derivatives were used, from 1 min to 72 h. A 24 h exposure was the most used (21/39), while nine studies used shorter exposure times, between 5 min and 2 h.
According to international standards (DIN EN ISO 10993-5:2009) guidelines for the cytotoxicity classification, plant derivatives are not cytotoxic when the inhibition of cell viability is less than 25%; slightly cytotoxic if inhibition is between 25 and 50%; moderately cytotoxic with inhibition between 50 and 75%; and strongly cytotoxic with inhibition higher than 75%, in comparison to the control group. According to this, plant derivatives in twelve studies did not display cytotoxic effects, two studies found slight toxicity and fourteen studies reported moderate and strong cytotoxicity.
Data from some studies did not allow researchers to classify cytotoxicity because the results were presented as CC50, IC50 or GI50. Moreover, three studies did not report values of CC50, IC50 or GI50 at the highest concentrations evaluated, and two studies indicated that the compounds did not display cytotoxicity, but it was not possible to extract information on the percentage of cellular viability after treatment.
In the study by de Oliveira et al. (2017) [42], four cell lines were treated with Rosmarinus officinalis extract, obtaining moderate cytotoxic effects in three cell lines, while in human breast carcinoma cells (MCF-7), the extract did not produce toxicity. Sudjana et al. (2012) [37] evaluated the essential oil of Melaleuca alternifolia in short exposure times, finding no cytotoxic effect in the three cell lines used.
Six of the studies that used 24 h of exposure did not identify cytotoxicity, two studies displayed results with slight cytotoxicity and eight studies reported plant derivatives with moderate and strong toxicity. Eight studies evaluated extracts on different cell lines, of which five reported no cytotoxicity or viability percentages >50%, even at concentrations of 25,000 μg/mL, as in the case of the extract of Arctium lappa [41]. Three studies described a cytotoxic effect in a concentration range from 100 μg/mL to 3125 μg/mL.
Exposure times lower than two hours were used in four studies, one of which reported that the essential oil of Melaleuca alternifolia [37] did not present cytotoxicity in different cell lines. Three studies showed a strong cytotoxicity of the extract of Eugenia uniflora [28], essential oil of Santolina impressa [49] and Allyl isothiocyanate of cruciferous plants [25] at concentrations between 200 μg/mL and 1050 μg/mL. Similarly, a study that evaluated seven extracts [19] reported results of slight and moderate toxicity with short treatment times.
Cytotoxicity curves were reported in three studies [34,50,53], evaluating 2, 3 and 7 different exposure times. Two of these studies reported similar results; one study did not identify any toxicity of the compound Hinokitiol [50] in the time range between 1 and 60 min, and, in another study, a strong cytotoxicity of the extracts of Cryptocarya mandioccana and Cryptocarya moschata [53] was found after exposure times of 6, 12 and 24 h.
Two studies evaluated Hinokitiol [44,50], finding similar results without a cytotoxic effect on the cells evaluated at concentrations of 1.6 μg/mL and 0.25 mM.
The compounds magnoflorine, ellagic acid, myricetin and eucarobustol from the leaves of Eucalyptus robusta, flavonoids and diterpenes of Prunus cerasoides, flavonoids from Moringa oleifera, an extract of Buchenavia tomentosa and an extract of Arctium lappa [20,24,30,35,41,46,48] should be noted because they did not present cytotoxicity at concentrations greater than or equal to those that have effect on C. albicans biofilms. A full description of cytotoxicity results can be found in Table 3.
2.7. Antibiofilm Activity
Seventeen of the included studies evaluated plant derivatives in models that allow the identification of whether inhibition occurs in C. albicans biofilm formation (BF); six studies evaluated the inhibition capacity on preformed biofilms (PB); and, in sixteen studies, both assessments were performed.
The most used techniques for the evaluation of biofilm formation (BF) inhibition were crystal violet (sixteen studies), XTT (sixteen studies) and viable counts (CFU/mL) (five studies). Other techniques used were dry weight measurement, alamar blue assay and MTT. In eight studies, the use of two techniques for the evaluation of BF was identified.
Twenty-three articles reported evaluations in PB, with the most used techniques being XTT (eleven studies), crystal violet (seven studies) and viable count (CFU/mL) (seven studies). Other techniques used in a smaller number of articles were MTT (three studies), SEM and fluorescence microscopy.
In the case of evaluations of BF, the most used exposure times were 24 h (twenty-five studies) and 48 h (seven studies). With regard to PB, the most used combination was 24 h of formation with 24 h of exposure (twelve studies). Other schemes, used in fewer studies, were 48 h of formation with 24 h of exposure (two studies) and 24 h of formation per 1 h of exposure (two studies).
Three BF studies used two exposure times, while, for PB, one study used four exposure times, another used five and the rest of the studies used one exposure time in different formations of the biofilm times, with 24 h being the most used exposure (fourteen studies).
Out of all the studies evaluating BF, 48.5% (15/33) showed inhibition results of the evaluated plant derivatives—lower than 60%—including two studies in which the evaluated derivatives did not display any antibiofilm activity. For PB, 39.1% (9/23 studies) of studies reporting this same level of inhibition were found.
As they show percentages of inhibition greater than 90% in BF models, we call attention to the flavonoids extracted from Moringa oleifera [30] seed coating (0.42 mg/mL), Cinnamomum burmannii extract, Cinnamon bark essential oil extracted from Cinnamomum verum [16], nepodin from Rumex crispus (Polygonaceae) [17], extract of Cymbopogon citratus (0.625 mg/mL) [32], myricetin and essential oil of Melaleuca alternifolia [37]. Out of these, in the PB evaluation, the flavonoids extracted from Moringa oleifera seed coating (0.42 mg/mL) and the extract of Cymbopogon citratus (0.625 mg/mL) showed reductions close to 80% with a 24 h exposure for flavonoids extracted from the Moringa oleifera seed coating in BF and PB, while, for Cymbopogon citratus, the treatment time was 8 h in PB.
Other compounds, such as dioscin [22] from herbs and vegetables of the Dioscorea genus (4 μg/mL) and dracorhodin perchlorate [23] from the exudates of the fruit of Daemonorops draco (32 μM), produced 89 and 80% inhibition in BF, respectively. In the first case (dioscin from herbs and vegetables of the Dioscorea genus), the evaluation in PB showed a reduction of 50% or more at 16 μg/mL, while dracorhodin perchlorate (64 μM) produced a reduction in PB of 20%, with treatments of 24 h.
Four studies included clinical isolates in their evaluations, of which three used BF evaluation and one used BF and PB evaluations. Three of these works identified that extracts from Eugenia spp. [28], essential oils extracted from Melaleuca alternifolia [37] and β-citronellol [39] displayed an inhibitory effect greater than 60% in the formation of biofilms of clinical C. albicans.
The concentrations at which these derivatives reach high percentages of inhibition are highly variable and, in some cases, range from micrograms to milligrams. The use of different measurement units was observed: molarity, p/v and v/v, used according to the nature of the evaluated derivative. A full description of plant derivatives’ activity can be found in Table 4.
3. Discussion
The increasing C. albicans antifungal resistance, partially attributed to its ability to form biofilms, has led to the search for therapeutic alternatives in plant derivatives, especially aiming to produce less cytotoxic effects.
This scoping review provided a good quality overview of in vitro studies on C. albicans biofilms and cytotoxic effects of plant derivatives, and included thirty-nine studies, thirty of which showed a low bias risk, ensuring an adequate quality, but also demonstrated some common limitations. Thirty studies did not clearly identify the death controls of the cytotoxic study techniques, eight studies did not provide completely defined outcome measures, seven studies did not clearly identify standard methods used to evaluate activity on planktonic cultures, six studies did not mention the used solvent to prepare the plant derivatives, five studies did not describe the process to define the concentrations of plant derivatives to be evaluated, four studies did not provide the number of replicates and repetitions of each test, four studies did not describe the statistical methods and three studies did not mention the control in the biofilm activity assessment.
A country with great biodiversity such as Brazil was expected to be the largest producer of this type of study (36%) and it was also expected that the public sector would show interest in financing them (twenty-nine studies), due to the possibility of developing bioeconomies that can be facilitated by the results.
Some studies evaluated other aspects of C. albicans that were not considered as inclusion criteria, but that complemented the knowledge of the plant derivatives’ effects. This is the case of the studies that evaluated inhibitory activity at the planktonic level and compared the results with activity in sessile cells, as in the studies by Yang et al. (2018) and Yang et al. (2018) [22,23], where they reported the inhibition of the adhesion of C. albicans to polystyrene surfaces in a dose-dependent manner by dioscin and dracorhodin perchlorate. In other studies, the morphosis of C. albicans was evaluated by derivatives such as Allyl Isothiocyanate, Sanguinarine and β-citronellol [25,39,40], which showed an effect in the yeast to hyphal transition and had effects on the structure of the biofilm through visualization by SEM/CLSM/fluorescence microscope (Table 1).
The literature analysis highlighted great variability among the selected studies, particularly regarding the cell type investigated, including twenty-six cell lines and ten primary cells. Variability also concerned C. albicans strains (thirteen reference strains were identified), the assays performed and the exposure conditions applied. Preparations from twenty-three plants demonstrated activity equal to or higher than 50% against C. albicans biofilms in BF or PB, seven of which were essential oils and eight were extracts. Some of the plant families were Myrtaceae (Eucalyptus robusta, Melaleuca alternifolia, Eugenia uniflora, Eugenia leitonii, Eugenia brasiliensis), Asteraceae (Santolina impressa, Artemisia judaica), Arecaceae (Daemonorops draco), Campanulaceae (Adenophora triphylla), Combretaceae (Buchenavia tomentosa), Dioscoreaceae (Dioscorea genus), Lamiaceae (Vitex gardneriana), Lauraceae (Cinnamomum burmannii, Cinnamomum verum, Cryptocarya mandioccana, Cryptocarya moschatta), Lythraceae (Punica granatum sarcotesta), Magnoliaceae (Magnolia officinalis), Menispermaceae (Fibraurea recisa), Piperaceae (Piper claussenianum), Poaceae (Cymbopogon citratus) and Papaveraceae.
We found that the in vitro cytotoxic effect was determined by different techniques, including different schemes of exposure times. Since 24 h of exposure was the most used scheme, the little variability allowed comparisons to be made. However, it is important to consider that each study adapted the methodology according to the possibility of clinical application expected for the derivative under study. Additionally, shorter exposure times naturally generate less cytotoxicity, as exemplified in the study of Nakamura et al. (2010) [50], in which hinokitiol did not present cytotoxicity in the time range between 1 and 60 min, while Oliveira et al. (2021) [53] evaluated Cryptocarya spp. extracts’ cytotoxicity at exposure times of 6, 12 and 24 h, and found strong toxicity at all times.
According to the analyzed publications, the predominant methodology was metabolic activity assays, whose results were expressed as CC50, CE50, CL50, IC50, DL50, GI50 and percentage cell viability. The difficulty in comparing the results between studies was one limitation due to the lack of consensus, since several factors, such as the origin and chemical composition of the oils, extracts and others, and particularly the selected cell lines, the technical conditions and the solvent used to dilute the plant derivatives, can influence the results of the in vitro tests [55].
As per the ISO 10993-5:2009 guidelines for cytotoxicity classification, the plant derivatives of twelve studies presented neither a cytotoxic effect nor slight toxicity, which is a key characteristic to consider along with the possible antifungal capacity. However, the derivatives of the fourteen studies showing moderate and strong cytotoxicity may have many other pharmacological properties, which label this as a future potential agent to treat many diseases, including cancer.
In the study by de Oliveira et al. (2017) [42], four cell lines were treated with Rosmarinus officinalis extract, obtaining moderate cytotoxic effects in three cell lines, while, in human breast carcinoma cells, MCF-7, the extract did not produce toxicity. Sudjana et al. (2012) [37] evaluated the essential oil of Melaleuca alternifolia at short exposure times. These findings reinforce the need to include different cell lines in studies and to establish cytotoxicity evaluation models consistent with the intended clinical applications.
Two of the included studies evaluated hinokitiol with similar results; no cytotoxic effect on the cells evaluated at the concentrations of 1.6 μg/mL and 0.25 mM. It is noteworthy that the evaluation methodologies used were different; Kim et al. (2017) [44] evaluated human-bone-marrow-derived mesenchymal stem cells, MSC, at prolonged exposure times (48 h) using the MTT assay, while Nakamura et al. (2010) [50] used epithelial cells, Ca9–22, in short exposure times (1 to 60 min) and a cytotoxicity detection kit.
According to the European Committee on Antimicrobial Susceptibility Testing (EUCAST), the antifungal clinical breakpoints are between 0.001 mg/L and 16 mg/L [56]. Using EUCAST guidelines in this review, the most active derivatives in planktonic culture were dioscin (4 μg/mL) [22], Eugenia leitonii extracts (15.62 μg/mL) [31], Eugenia brasiliensis extracts from seeds (15.62 μg/mL) [31], eucarobustol E [35] (16 μg/mL)./mL), licochalcone A [36] (6.25–12.5 μg/mL), glabridin [36] (12.5–6.25 μg/mL), sanguinarine [40] (3.2 μg/mL), pomegranate extract from Punica granatum [43] (3.9 μg/mL) and hinokitiol [50] (1.6 μg/mL). Although EUCAST does not have antifungal breakpoints for biofilms, if we compare the current breakpoints with the results on biofilms, the following plant derivatives could be considered to inhibit (≥50%) C. albicans biofilm either in BF or in PB: dioscin [22] (4–16 μg/mL), Adenophora triphylla var. japonica extract [52] (6.25 μg/mL), chitin-binding lectin (PgTeL) from Punica granatum sarcotesta [21] (0.39 μg/mL), magnolol and honokiol from Magnolia officinalis [27] (16 μg/mL), roemerine from Fibraurea recisa [29] (8 μg/mL), sanguinarine [40] (1.6–3.2 μg/mL), nepodin from Rumex crispus (2–5 μg/mL), Rumex japonicus extract (5 μg/mL) [17], licochalcone A (2 μg/mL) and hinokitiol (3.1 μg/mL) [36,44]. Among these, dioscin, sanguinarine and hinokitiol presented an inhibitory effect on planktonic cells and both biofilm models, with results expressed in p/v.
Since comparing the results in different concentration units is not possible, it is difficult to identify the derivatives with the best inhibitory characteristics, and it would be desirable to reach a consensus for future studies. Nevertheless, it is possible to identify the following as the plant derivatives displaying percentages of inhibition higher than 90% in BF models of C. albicans biofilms, regardless of their concentration or the strain evaluated: flavonoids from Moringa oleifera seed coating [30], Cinnamomum burmannii extract, cinnamon bark essential oil extracted from Cinnamomum verum, nepodin from Rumex crispus (Polygonaceae), extract of Cymbopogon citratus (0.625 mg/mL), myricetin and essential oil of Melaleuca alternifolia [16,32,37] (Table 4). These preparations included essential oils, extracts, naphthols and flavonoids. Most of the plant preparations acted on biofilm formation or mature biofilms above 50%, turning several of these derivatives into promising options for the control of C. albicans biofilms; for example, in abiotic surfaces.
Antibiofilm activity may vary between plants, even in the same family. Although few of the studies included in this review used the same plant derivatives, this can be observed in the case of the studies carried out with the plant derivatives of the Myrtaceae family [28,31,35,37], where there are differences in the inhibitory activity on C. albicans biofilms, perhaps due to the chemical nature of the derivatives, the genus of the plants or the different C. albicans strain used. Similarly, three or four studies reported that the derivatives did not present cytotoxic effects on the viability of animal cells.
Even when flavonoids extracted from the Moringa oleifera seed coating (0.42 mg/mL) and the extract of Cymbopogon citratus (0.625 mg/mL) [30,32] showed the best reductions (80%) in PB, taking into account the set of results of cytotoxicity and the inhibitory effect on C. albicans biofilms, it can be said that the following compounds have a special potential: magnoflorine, ellagic acid, roemerine, myricetin, camphor, licochalcone A, nepodin from Rumex crispus (Polygonaceae), eucarobustol from the leaves of Eucalyptus robusta, flavonoids and diterpenes of Prunus cerasoides, flavonoids from Moringa oleifera, Sanguinarine from Papaveraceae family, chitin-binding lectin (PgTeL) from P. granatum sarcotesta, cinnamon bark essential oil extracted from Cinnamomum verum, essential oil of Santolina impressa, extract of Rosmarinus officinalis, extract of Eugenia uniflora, Adenophora triphylla var. japonica extract, Cinnamomum burmanni extract, extract of Buchenavia tomentosa, extract of Punica granatum and extracts from Eugenia leitonii (seed) and Eugenia brasiliensis (seed and leaf). This conclusion is reached because they did not present cytotoxicity at concentrations higher than or equal to those that had an inhibitory effect on C. albicans biofilm.
4. Limitations of the Study
In this review, the investigation was limited to in vitro cell populations, both for cytotoxicity and inhibition C. albicans biofilm, with the aim of facilitating the selection of derivatives to study in the future and given the great variety reported in the literature.
As mentioned before, the numerous variables related to the technical aspects of obtaining plant derivatives and their subsequent preparation, toxicity evaluation and anti-biofilm capacity, as well as the variety of valid ways to express the results, constitute the main limitation for the analysis of the literature.
5. Conclusions
The results of this review show that, among a variety of plant derivatives that have been studied for their inhibitory effect on C. albicans biofilms and for their cytotoxic capacity, several can be considered as promising for the development of future research and bioproducts applicable to human health. Those capable of generating biofilm inhibition with short exposure times, as in the case of extract of Cymbopogon citratus, and those that combine an anti-biofilm effect with short exposure (<8 h) and a low cytotoxic effect, such as Buchenavia tomentosa extract, Rosmarinus Officinalis extract, cinnamon bark essential oil extracted from Cinnamomum verum and flavonoids extracted from Moringa oleifera seed coating, are particularly interesting for future studies in experimental models, including controlled clinical trials. In order to move forward in the study of these and other plant derivatives with promising results, it is important to consider the need for consensus when conducting in vitro experiments and reporting their results to produce comparable information.
6. Materials and Methods
This scoping review was conducted according to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) for Scoping Reviews. The aim was to identify the current state of in vitro research on the ability of plant derivatives to inhibit C. albicans biofilms, considering their possible cytotoxic effect on mammalian cells. The research team constructed the research question in accordance with the Population, Concept and Context (PCC) policy format [57] (Table 5).
Table 5.
Focused research questions presented using the PCC policy format.
6.1. Search Strategy
Two different reviewers carried out the search process. Specific search strategies were developed and implemented using the following electronic databases: Science Direct, PubMed, Scopus and Lilacs, which were limited to 28 February 2022. The observation period was of approximately twelve years, set between 2010 and 2022. A search strategy was developed using MeSH terms, and adjustments were made to match the same terms in different search engines across the four databases (Table 6), combined with database-specific filters.
Table 6.
Search strategy used in each database.
6.2. Criteria for the Eligibility of the Studies
Two authors assessed all papers. Eligibility criteria were defined based on the PCC for institutional methodology for scoping reviews. The following study criteria were considered for inclusion: (i) articles assessing the effects of natural compounds from plants on C. albicans anti-biofilm activity; (ii) articles providing a full description of the methods and results; (iii) articles describing the antifungal activity of single or combined compounds, as long as they were of natural origin; (iv) articles reporting cytotoxic activity assessed by in vitro experimental methods; and (v) articles in English and Spanish. Reviews, books, chapters and studies evaluating cytotoxicity in vivo and in situ were excluded.
6.3. Risk of Bias Assessment
Researchers assessed the risk of bias from individual studies. The assessment was adapted from previous systematic reviews [58]. We used nine parameters to evaluate the quality of each study: (1) description of the method for plant derivatives obtention, (2) solvent used, (3) mention of a control in the biofilm activity assessment, (4) the presence of a control in the cytotoxic activity assessment, (5) description of the number of replicates and repetitions of each test, (6) use of standard methods for activity determination on planktonic cultures, (7) description of the process to define the concentrations of plant derivatives to be evaluated, (8) description of statistical methods used, and (9) completely defined measures of outcome. Publications reporting fewer than four items were classified as having a high risk of bias, whereas those reporting more than six were classified as low risk.
6.4. Data Extraction Process, Synthesis and Analysis
All article titles initially found in the search were selected based on the eligibility criteria and duplicates were eliminated. Titles were read, and those that did not indicate relevance were excluded. The inclusion criteria for the abstract-based selection stage were in vitro studies that investigated plant derivatives’ inhibition of C. albicans biofilms.
The reviewers read the full texts of potentially eligible studies based on the inclusion/exclusion criteria, and any disagreements were resolved in consultation with another author. Only papers with all the eligibility criteria were included.
Scientific and technical information were extracted using a data table in Microsoft® Office Excel® (Version 2211, Redmond, WA, USA). The following data were tabulated, from the qualitative analyses, about what is currently known in the literature regarding the activity of plant derivatives in C. albicans biofilms and human cells: author(s), year of publication, strain, compound evaluated, solvent (for plant derivatives’ preparation), type of plant derivative (such as extract, essential oil, terpene and protein), planktonic methodology and results, evaluation technique on biofilm, time exposure in biofilm, result on biofilm formation or preformed biofilm, cell lines or animal cells evaluated and cytotoxicity technique and results.
Author Contributions
The authors M.L.-O. and L.A.-D. contributed equally to the literature search and information extraction. M.L.-O. and M.C.M.-P. were involved in table making and in writing and preparing the original draft. The conceptualization was performed mainly by M.C.M.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Ministry of Science, Technology and Innovation, the Ministry of Education, the Ministry of Industry, Commerce and Tourism, and ICETEX, Programme Ecosistema Científico-Colombia Científica, from the Francisco José de Caldas Fund, Grant RC-FP44842-212-2018.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors sincerely thanks Andrés Agudelo for his advice. The authors would like to thank David Nichols for the revision and English corrections.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cavalheiro, M.; Cacho-Teixeira, M. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Tadec, L.; Talarmin, J.; Gastinne, T. Epidemiology, risk factor, species distribution, antifungal resistance and outcome of candidemia at a single French hospital: A 7-year study. Mycoses 2016, 59, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xia, R.; Zhang, Q.; Bai, C.; Li, Z.; Zhang, P. Evaluation of candidemia in epidemiology and risk factors among cancer patients in a cancer center of China: An 8-year case-control study. BMC Infect. Dis. 2017, 17, 1–8. [Google Scholar] [CrossRef][Green Version]
- Rautemaa, R.; Ramage, G. Oral candidosis clinical challenges of a biofilm disease. Crit. Rev. Microbiol. 2011, 37, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Donnell, L.E.O.; Millhouse, E.; Sherry, L.; Kean, R.; Malcolm, J.; Nile, C.J.; Ramage, G. Polymicrobial Candida biofilms: Friends and foe in the oral cavity. FEMS Yeast Res. 2015, 15, 1–14. [Google Scholar] [CrossRef]
- Pappas, P.G. Invasive candidiasis. Infect. Dis. Clin. N. Am. 2006, 20, 485–506. [Google Scholar] [CrossRef]
- Stringer, A.M.; Logan, R.M. The role of oral flora in the development of chemotherapy-induced oral mucositis. J. Oral Pathol. Med. 2015, 44, 81–87. [Google Scholar] [CrossRef]
- Vanhoecke, B.; De Ryck, T.; Stringer, A.; Van De Wiele, T.; Keefe, D. Microbiota and their role in the pathogenesis of oral mucositis. Oral Dis. 2014, 21, 17–30. [Google Scholar] [CrossRef]
- Al-ansari, S.; Zecha, J.A.E.M.; Barasch, A.; Lange, J.D.; Rozema, F.R.; Raber-durlacher, J.E. Oral mucositis induced by anticancer therapies. Curr. Oral Heal. Rep. 2015, 2, 202–211. [Google Scholar] [CrossRef]
- Lalla, R.V.; Saunders, D.P.; Peterson, D.E.; Rcseda, F.D.S. Chemotherapy or radiation- induced oral mucositis oral mucositis stomatitis cancer chemotherapy radiation therapy. Dent. Clin. N. Am. 2014, 58, 341–349. [Google Scholar] [CrossRef]
- Douglas, L.J. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Höfs, S.; Mogavero, S.; Hube, B. Interaction of Candida albicans with host cells: Virulence factors, host defense, escape strategies, and the microbiota. J. Microbiol. 2016, 54, 149–169. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, S.; Mitchell, A.P. Mucosal biofilms of Candida albicans. Curr. Opin. Microbiol. 2012, 14, 380–385. [Google Scholar] [CrossRef]
- Tournu, H.; Van Dijck, P. Candida biofilms and the host: Models and new concepts for eradication. Int. J. Microbiol. 2012, 2012, 845352. [Google Scholar] [CrossRef]
- Veilleux, M.P.; Grenier, D. Determination of the effects of cinnamon bark fractions on Candida albicans and oral epithelial cells. BMC Complement Altern. Med. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.G.; Khadke, S.K.; Yamano, A.; Watanabe, A.; Lee, J. Inhibition of biofilm formation by Candida albicans and polymicrobial microorganisms by nepodin via hyphal-growth suppression. ACS Infect. Dis. 2019, 5, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, X.; Sui, Y.; Ma, Z.; Feng, X.; Wang, F.; Ma, T. Lycorine hydrochloride inhibits the virulence traits of Candida albicans. BioMed Res. Int. 2019, 2019, 1851740. [Google Scholar] [CrossRef]
- Ribeiro, S.M.; Fratucelli, É.D.O.; Bueno, P.C.P.; de Castro, M.K.V.; Francisco, A.A.; Cavalheiro, A.J.; Klein, M.I. Antimicrobial and antibiofilm activities of Casearia Sylvestris extracts from distinct brazilian biomes against Streptococcus mutans and Candida albicans. BMC Complement Altern. Med. 2019, 19, 308. [Google Scholar] [CrossRef]
- Kim, J.; Ha Quang Bao, T.; Shin, Y.K.; Kim, K.Y. Antifungal activity of magnoflorine against Candida Strains. World J. Microbiol. Biotechnol. 2018, 34, 1–7. [Google Scholar] [CrossRef]
- da Silva, P.M.; de Moura, M.C.; Gomes, F.S.; da Silva-Trentin, D.; Silva de Oliveira, A.P.; de Mello, G.S.V.; da Rocha-Pitta, M.G.; de Melo-Rego, M.J.B.; Coelho, L.C.B.B.; Macedo, A.J.; et al. PgTeL, the lectin found in Punica Granatum Juice, is an antifungal agent against Candida albicans and Candida krusei. Int. J. Biol. Macromol. 2018, 108, 391–400. [Google Scholar] [CrossRef]
- Yang, L.; Liu, X.; Zhong, L.; Sui, Y.; Quan, G.; Huang, Y.; Wang, F.; Ma, T. Dioscin inhibits virulence factors of Candida albicans. BioMed Res. Int. 2018, 2018, 4651726. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.F.; Liu, X.; Lv, L.L.; Ma, Z.M.; Feng, X.C.; Ma, T.H. Dracorhodin perchlorate inhibits biofilm formation and virulence factors of Candida albicans. J. Mycol. Med. 2018, 28, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Lourenção-Brighenti, F.; Salvador, M.J.; Gontijo, A.V.L.; Delbem, A.C.B.; Delbem, Á.C.B.; Soares, C.P.; De Oliveira, M.A.C.; Girondi, C.M.; Koga-Ito, C.Y. Plant extracts: Initial screening, identification of bioactive compounds and effect against Candida albicans biofilms. Future Microbiol. 2017, 12, 15–27. [Google Scholar] [CrossRef]
- Raut, J.S.; Bansode, B.S.; Jadhav, A.K.; Karuppayil, S.M. Activity of Allyl isothiocyanate and its synergy with fluconazole against Candida albicans biofilms. J. Microbiol. Biotechnol. 2017, 27, 685–693. [Google Scholar] [CrossRef]
- Sadowska, B.; Budzyńska, A.; Stochmal, A.; Żuchowski, J.; Różalska, B. Novel properties of Hippophae rhamnoides L. twig and leaf extracts—Anti-virulence action and synergy with antifungals studied in vitro on Candida spp. model. Microb. Pathog. 2017, 107, 372–379. [Google Scholar] [CrossRef]
- Sun, L.; Liao, K.; Wang, D. Effects of magnolol and honokiol on adhesion, yeast-hyphal transition, and formation of biofilm by Candida albicans. PLoS ONE 2015, 10, e01176952015. [Google Scholar] [CrossRef]
- Souza, L.B.F.C.; Silva-Rocha, W.P.; Ferreira, M.R.A.; Soares, L.A.L.; Svidzinski, T.I.E.; Milan, E.P.; Pires, R.H.; Almeida, A.M.F.; Mendes-Giannini, M.J.S.; Chaves, G.M. Influence of Eugenia uniflora extract on adhesion to human buccal epithelial cells, biofilm formation, and cell surface hydrophobicity of Candida spp. from the oral cavity of kidney transplant recipients. Molecules 2018, 23, 2418. [Google Scholar] [CrossRef]
- Ma, C.; Du, F.; Yan, L.; He, G.; He, J.; Wang, C.; Rao, G.; Jiang, Y.; Xu, G. Potent activities of roemerine against Candida albicans and the underlying mechanisms. Molecules 2015, 20, 17913–17928. [Google Scholar] [CrossRef]
- Onsare, J.G.; Arora, D.S. Antibiofilm potential of flavonoids extracted from Moringa oleifera Seed Coat against Staphylococcus aureus, Pseudomonas aeruginosa and Candida albicans. J. Appl. Microbiol. 2015, 118, 313–325. [Google Scholar] [CrossRef]
- Sardi, J.d.C.O.; Freires, I.A.; Lazarini, J.G.; Infante, J.; de Alencar, S.M.; Rosalen, P.L. Unexplored endemic fruit species from Brazil: Antibiofilm properties, insights into mode of action, and systemic toxicity of four Eugenia spp. Microb. Pathog. 2017, 105, 280–287. [Google Scholar] [CrossRef]
- Madeira, P.L.B.; Carvalho, L.T.; Paschoal, M.A.B.; de Sousa, E.M.; Moffa, E.B.; da Silva, M.A.; Tavarez, R.d.J.R.; Gonçalves, L.M. In vitro effects of lemongrass extract on Candida albicans biofilms, human cells viability, and denture surface. Front. Cell. Infect. Microbiol. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Abu-Darwish, M.S.; Cabral, C.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Zulfiqar, A.; Khan, I.A.; Efferth, T.; Salgueiro, L. Chemical composition and biological activities of Artemisia judaica essential oil from southern desert of jordan. J. Ethnopharmacol. 2016, 191, 161–168. [Google Scholar] [CrossRef]
- Sadowska, B.; Budzyńska, A.; Wieckowska-Szakiel, M.; Paszkiewicz, M.; Stochmal, A.; Moniuszko-Szajwaj, B.; Kowalczyk, M.; Rózalska, B. New Pharmacological properties of Medicago sativa and Saponaria officinalis saponin-rich fractions addressed to Candida albicans. J. Med. Microbiol. 2014, 63, 1076–1086. [Google Scholar] [CrossRef]
- Liu, R.H.; Shang, Z.C.; Li, T.X.; Yang, M.H.; Kong, L.Y. In vitro antibiofilm activity of eucarobustol e against Candida albicans. Antimicrob. Agents Chemother. 2017, 61, 1–15. [Google Scholar] [CrossRef]
- Messier, C.; Grenier, D. Effect of licorice Compounds licochalcone A, glabridin and glycyrrhizic acid on growth and virulence properties of Candida albicans. Mycoses 2011, 54, 801–806. [Google Scholar] [CrossRef]
- Sudjana, A.N.; Carson, C.F.; Carson, K.C.; Riley, T.V.; Hammer, K.A. Candida albicans adhesion to human epithelial cells and polystyrene and formation of biofilm is reduced by sub-inhibitory Melaleuca alternifolia (tea tree) essential oil. Med. Mycol. 2012, 50, 863–870. [Google Scholar] [CrossRef]
- Curvelo, J.A.R.; Marques, A.M.; Barreto, A.L.S.; Romanos, M.T.V.; Portela, M.B.; Kaplan, M.A.C.; Soares, R.M.A. A Novel nerolidol-rich essential oil from Piper claussenianum modulates Candida albicans biofilm. J. Med. Microbiol. 2014, 63, 697–702. [Google Scholar] [CrossRef]
- Sharma, Y.; Rastogi, S.K.; Perwez, A.; Rizvi, M.A.; Manzoor, N. β-Citronellol alters cell surface properties of Candida albicans to influence pathogenicity related traits. Med. Mycol. 2020, 58, 93–106. [Google Scholar] [CrossRef]
- Zhong, H.; Hu, D.D.; Hu, G.H.; Su, J.; Bi, S.; Zhang, Z.E.; Wang, Z.; Zhang, R.L.; Xu, Z.; Jiang, Y.Y.; et al. Activity of sanguinarine against Candida albicans biofilms. Antimicrob. Agents Chemother. 2017, 61, 1–9. [Google Scholar] [CrossRef]
- de Oliveira, J.R.; de Aguiar-Almeida, R.B.; das Graças Figueiredo-Vilela, P.; de Oliveira, F.E.; da Rocha, R.F.; Jorge, A.O.C.; de Oliveira, L.D. Control of microorganisms of oral health interest with Arctium lappa L. (Burdock) extract non-cytotoxic to cell culture of macrophages (RAW 264.7). Arch. Oral Biol. 2014, 59, 808–814. [Google Scholar] [CrossRef]
- de Oliveira, J.R.; de Jesus, D.; Figueira, L.W.; de Oliveira, F.E.; Pacheco-Soares, C.; Camargo, S.E.A.; Jorge, A.O.C.; de Oliveira, L.D. Biological activities of Rosmarinus officinalis L. (Rosemary) extract as analyzed in microorganisms and cells. Exp. Biol. Med. 2017, 242, 625–634. [Google Scholar] [CrossRef]
- Endo, E.H.; Ueda-Nakamura, T.; Nakamura, C.V.; Filho, B.P.D. Activity of spray-dried microparticles containing Pomegranate peel extract against Candida albicans. Molecules 2012, 17, 10094–10107. [Google Scholar] [CrossRef]
- Kim, D.J.; Lee, M.W.; Choi, J.S.; Lee, S.G.; Park, J.Y.; Kim, S.W. Inhibitory activity of hinokitiol against biofilm formation in fluconazole-resistant Candida species. PLoS ONE 2017, 12, 1–11. [Google Scholar] [CrossRef]
- Vale, J.P.C.D.; Ribeiro, L.H.D.F.; Vasconcelos, M.A.D.; Sá-Firmino, N.C.; Pereira, A.L.; Nascimento, M.F.D.; Rodrigues, T.H.S.; Silva, P.T.D.; Sousa, K.C.D.; Silva, R.B.D.; et al. Chemical composition, antioxidant, antimicrobial and antibiofilm activities of Vitex gardneriana schauer leaves’s essential oil. Microb. Pathog. 2019, 135, 103608. [Google Scholar] [CrossRef]
- Arora, D.S.; Mahajan, H. Major phytoconstituents of Prunus cerasoides responsible for antimicrobial and antibiofilm potential against some reference strains of pathogenic bacteria and clinical isolates of MRSA. Appl. Biochem. Biotechnol. 2019, 188, 1185–1204. [Google Scholar] [CrossRef]
- Rivas da Silva, A.C.; Lopes, P.M.; Barros de Azevedo, M.M.; Costa, D.C.; Alviano, C.S.; Alviano, D.S. Biological activities of α-pinene and β-pinene enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef]
- Mo, F.; Ma, J.; Yang, X.; Zhang, P.; Li, Q.; Zhang, J. In vitro and in vivo effects of the combination of myricetin and miconazole nitrate incorporated to thermosensitive hydrogels, on C. albicans Biofilms. Phytomedicine 2020, 71, 153223. [Google Scholar] [CrossRef]
- Alves-Silva, J.M.; Zuzarte, M.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Salgueiro, L. Unveiling the bioactive potential of the essential oil of a portuguese endemism, Santolina impressa. J. Ethnopharmacol. 2019, 244, 112120. [Google Scholar] [CrossRef]
- Nakamura, M.; Fujibayashi, T.; Tominaga, A.; Satoh, N.; Kawarai, T.; Shinozuka, O.; Watanabe, H.; Yamazaki, T.; Senpuku, H. Hinokitiol inhibits Candida albicans adherence to oral epithelial cells. J. Oral. Biosci. 2010, 52, 42–50. [Google Scholar] [CrossRef]
- Ivanov, M.; Kannan, A.; Stojković, D.S.; Glamočlija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Soković, M. Camphor and eucalyptol—Anticandidal spectrum, antivirulence effect, efflux pumps interference and cytotoxicity. Int. J. Mol. Sci. 2021, 22, 483. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, K.Y. Adenophora triphylla var. japonica inhibits Candida biofilm formation, increases susceptibility to antifungal agents and reduces infection. Int. J. Mol. Sci. 2021, 22, 12523. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira-Zoccolotti, J.; Cavalheiro, A.J.; Tasso, C.O.; Ribas, B.R.; Ferrisse, T.M.; Jorge, J.H. Antimicrobial efficacy and biocompatibility of extracts from Cryptocarya Species. PLoS ONE 2021, 16, 1–17. [Google Scholar] [CrossRef]
- Pereira, J.V.; Freires, I.A.; Castilho, A.R.; da Cunha, M.G.; Alves, H.D.S.; Rosalen, P.L. Antifungal potential of Sideroxylon obtusifolium and Syzygium Cumini and their mode of action against Candida albicans. Pharm. Biol. 2016, 54, 2312–2319. [Google Scholar] [CrossRef] [PubMed]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards advances in medicinal plant antimicrobial activity: A review study on challenges and future perspectives. Microorganism 2021, 9, 2041. [Google Scholar] [CrossRef] [PubMed]
- EUCAST: Breakpoints for Antifungals. Available online: https://www.eucast.org/astoffungi/clinicalbreakpointsforantifungals/ (accessed on 20 October 2022).
- Munn, Z.; Peters, M.D.J.; Stern, C.; Tufanaru, C.; Mcarthur, A.; Aromataris, E. Systematic review or scoping review? guidance for authors when choosing between a systematic or scoping review approach. BMC Med. Res. Methodol. 2018, 18, 143. [Google Scholar] [CrossRef] [PubMed]
- Sarkis-Onofre, R.; Skupien, J.; Cenci, M.S.; Moraes, R.R.; Pereira-Cenci, T. The role of resin cement on bond strength of glass-fiber posts luted into root canals: A systematic review and meta- analysis of in vitro studies. Oper. Dent. 2014, 39, 31–44. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).