
Essential Oils from Colombian Plants: Antiviral Potential against Dengue Virus Based on Chemical Composition, In Vitro and In Silico Analyses

 , , , and
, , , and
Abstract

1. Introduction
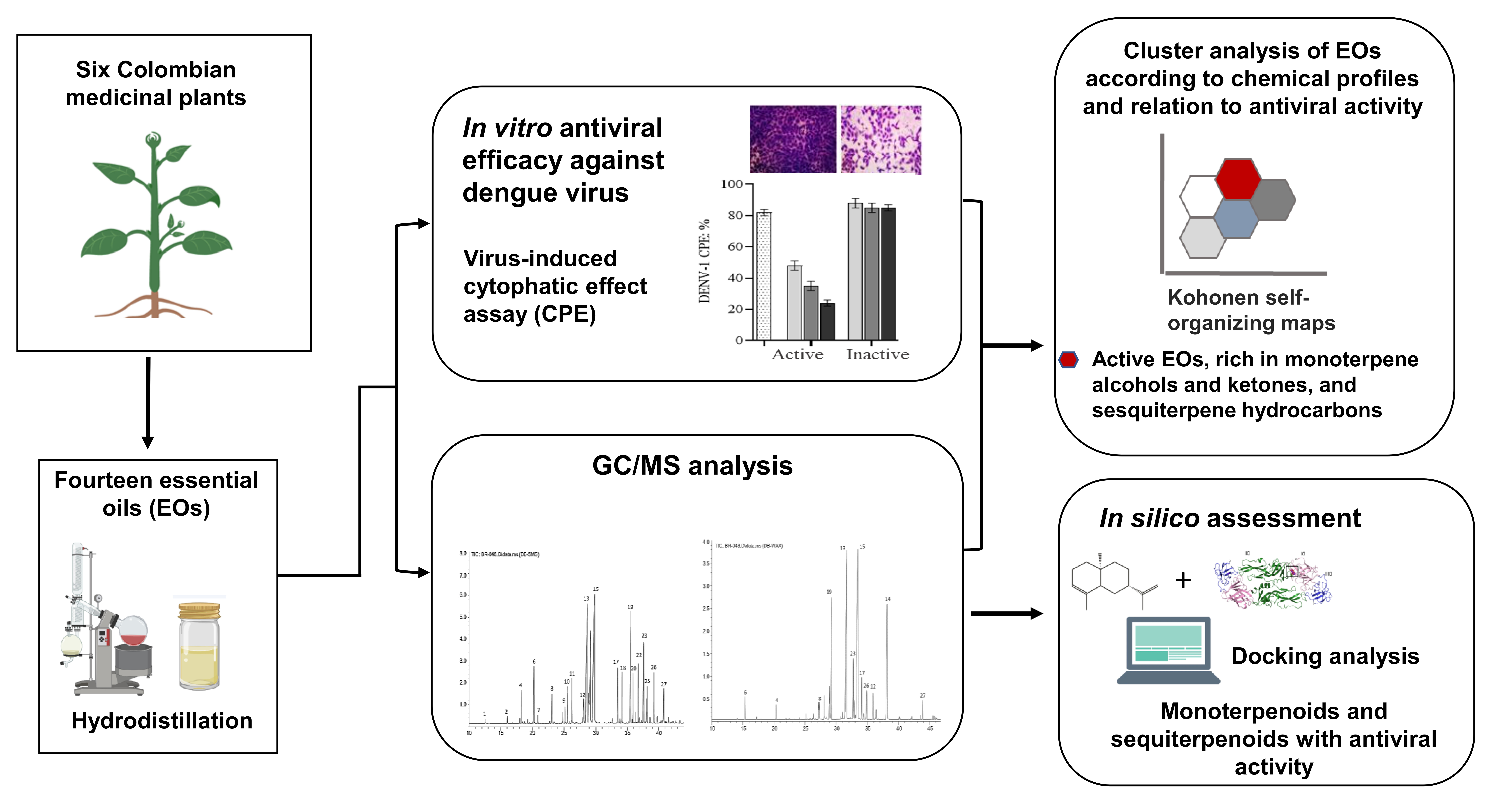
2. Results
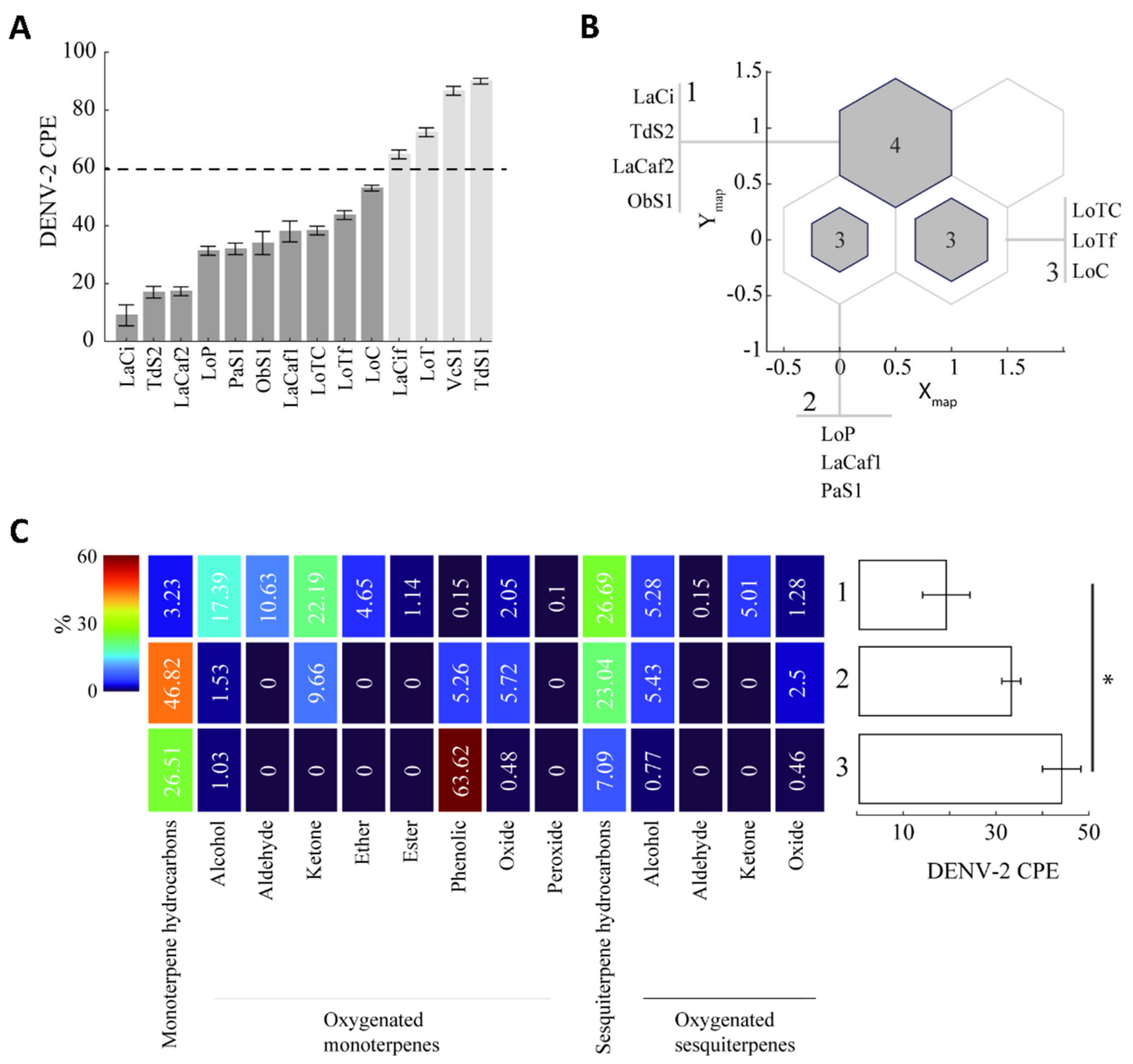
2.1. EO Chemical Composition
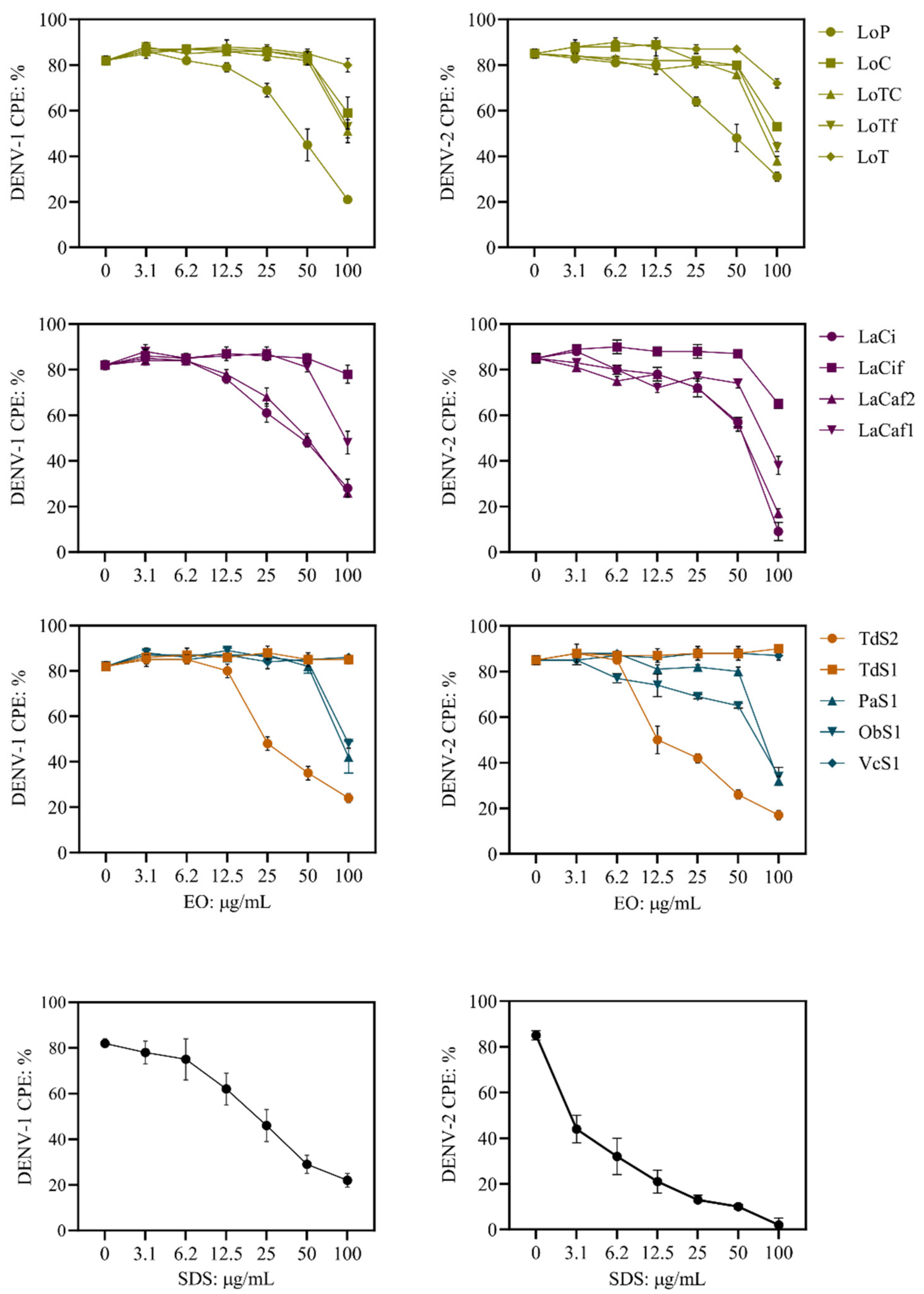
2.2. Antiviral Activity
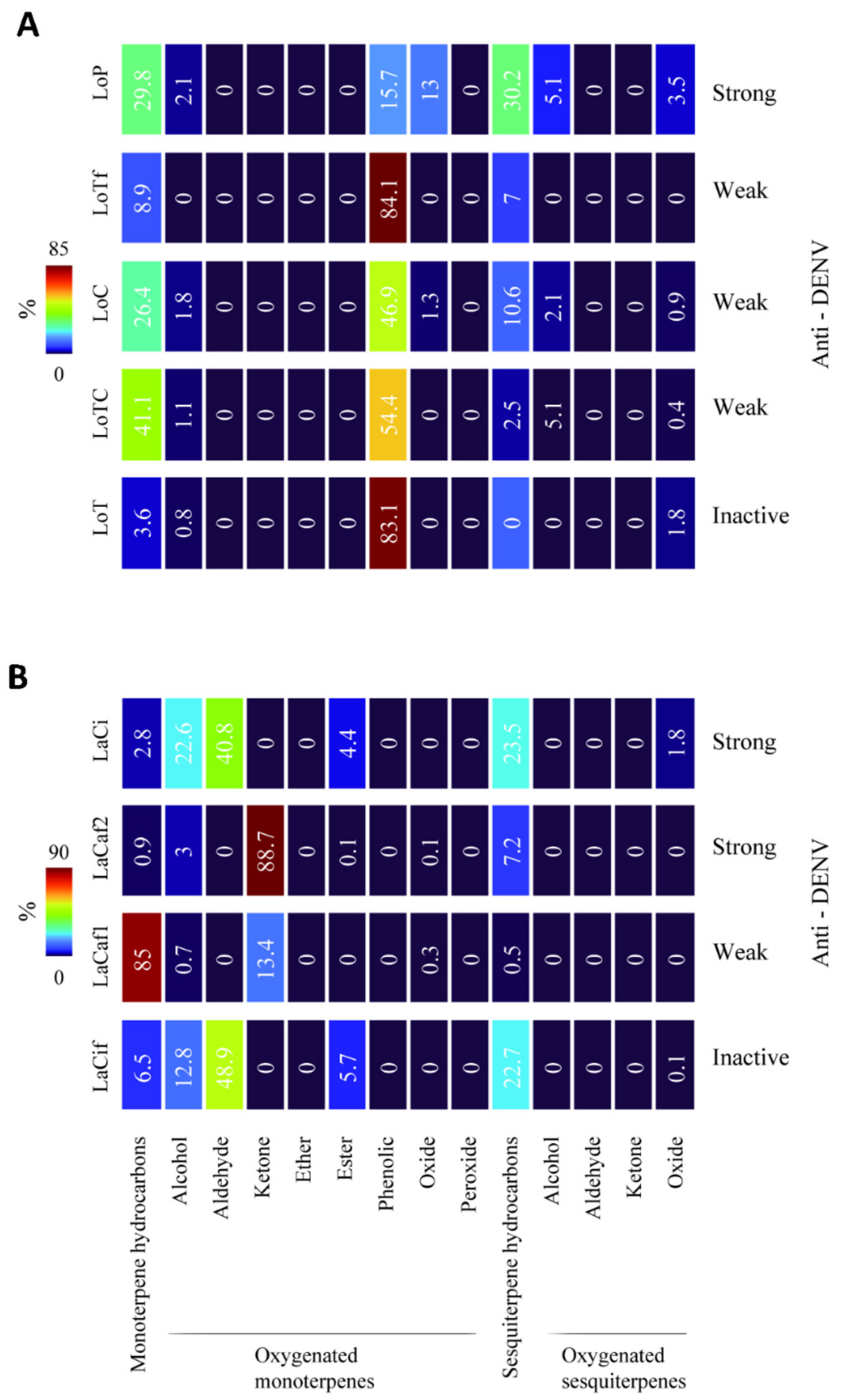
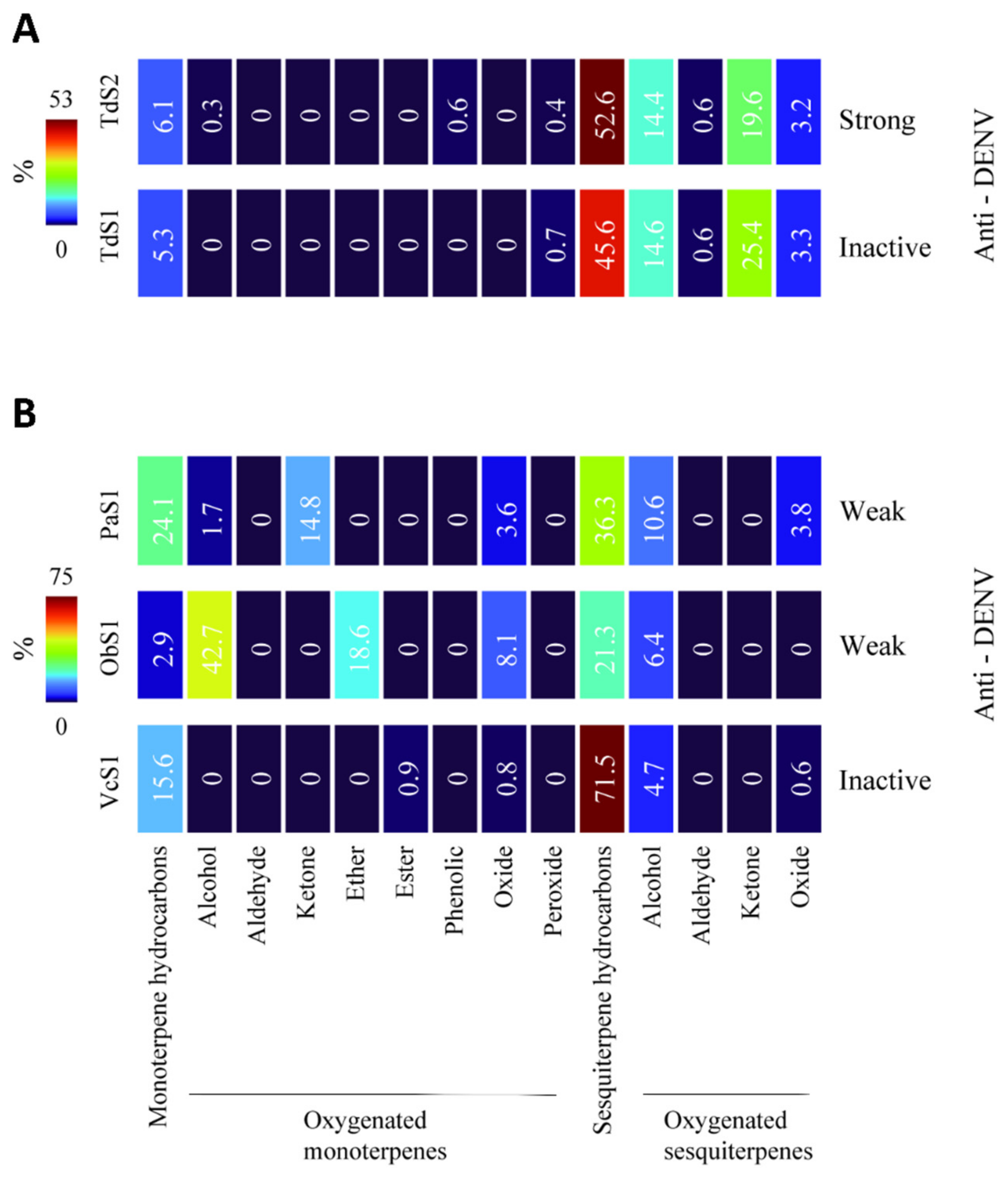
2.3. Chemical Cluster of EOs and Antiviral Activity
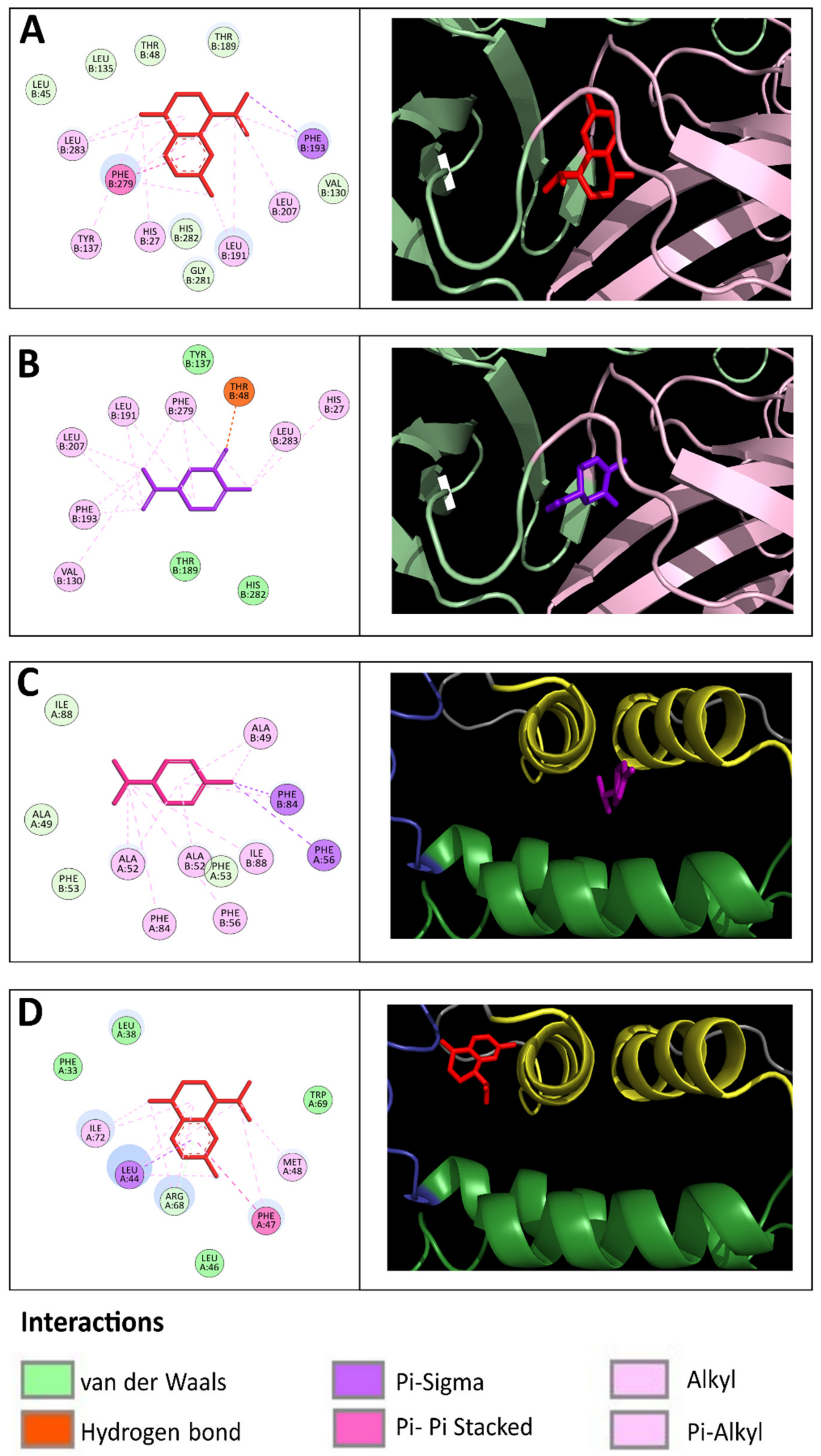
2.4. Molecular Interactions between EO Compounds and DENV-2 Proteins
3. Discussion
4. Materials and Methods
4.1. Plant Material and EO Distillation
4.2. Chromatographic Analysis
4.3. Virus and Cells
4.4. Cytotoxicity Assay
4.5. Cytopathic Effect (CPE)-Based Antiviral Assay
4.6. In Silico Analysis
4.7. Correlation and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Zeng, Z.; Zhan, J.; Chen, L.; Chen, H.; Cheng, S. Global, regional, and national dengue burden from 1990 to 2017: A systematic analysis based on the global burden of disease study 2017. EClinicalMedicine 2021, 32, 100712. [Google Scholar] [CrossRef] [PubMed]
- Kalayanarooj, S. Clinical manifestations and management of dengue/DHF/DSS. Trop. Med. Health 2011, 39 (Suppl. S4), 83–87. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Barbosa, H.; Medina-Moreno, S.; Zapata, J.; Chua, J. Dengue infections in Colombia: Epidemiological trends of a hyperendemic country. Trop. Med. Infect. Dis. 2020, 5, 156. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.P. Dengue drug discovery: Progress, challenges and outlook. Antivir. Res. 2019, 163, 156–178. [Google Scholar] [CrossRef] [PubMed]
- Obi, J.O.; Gutiérrez-Barbosa, H.; Chua, J.V.; Deredge, D.J. Current trends and limitations in dengue antiviral research. Trop. Med. Infect. Dis. 2021, 6, 180. [Google Scholar] [CrossRef]
- Lim, S.Y.M.; Chieng, J.Y.; Pan, Y. Recent insights on anti-dengue virus (DENV) medicinal plants: Review on in vitro, in vivo and in silico discoveries. All Life 2021, 14, 1–33. [Google Scholar] [CrossRef]
- Dhiman, M.; Sharma, L.; Dadhich, A.; Dhawan, P.; Sharma, M.M. Traditional knowledge to contemporary medication in the treatment of infectious disease dengue: A review. Front. Pharmacol. 2022, 13, 750494. [Google Scholar] [CrossRef]
- Chavda, V.P.; Kumar, A.; Banerjee, R.; Das, N. Ayurvedic and other herbal remedies for dengue: An update. Clin. Complement. Med. Pharmacol. 2022, 2, 100024. [Google Scholar] [CrossRef]
- Manion, C.R.; Widder, R.M. Essentials of essential oils. Am. J. Health-Syst. Pharm. 2017, 74, e153–e162. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.; Ademiluyi, A.; et al. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Wani, A.R.; Yadav, K.; Khursheed, A.; Rather, M.A. An updated and comprehensive review of the antiviral potential of essential oils and their chemical constituents with special focus on their mechanism of action against various influenza and coronaviruses. Microb. Pathog. 2021, 152, 104620. [Google Scholar] [CrossRef]
- Reichling, J. Antiviral and virucidal properties of essential oils and isolated compounds—A scientific approach. Planta Med. 2021, 88, 587–603. [Google Scholar] [CrossRef]
- Ma, L.; Yao, L. Antiviral effects of plant-derived essential oils and their components: An updated review. Molecules 2020, 25, 2627. [Google Scholar] [CrossRef]
- Anastasiou, C.; Buchbauer, G.; Musterman, M.; Placeholder, P. Essential oils as immunomodulators: Some examples. Open Chem. 2017, 15, 352–370. [Google Scholar] [CrossRef]
- Sandner, G.; Heckmann, M.; Weghuber, J. Immunomodulatory activities of selected essential oils. Biomolecules 2020, 10, 1139. [Google Scholar] [CrossRef]
- Valussi, M.; Antonelli, M.; Donelli, D.; Firenzuoli, F. Appropriate use of essential oils and their components in the management of upper respiratory tract symptoms in patients with COVID-19. J. Herb. Med. 2021, 28, 100451. [Google Scholar] [CrossRef]
- Ćavar Zeljković, S.; Schadich, E.; Džubák, P.; Hajdúch, M.; Tarkowski, P. Antiviral activity of selected Lamiaceae essential oils and their monoterpenes against SARS-Cov-2. Front. Pharmacol. 2022, 13, 893634. [Google Scholar] [CrossRef]
- Bernal, R.; Gradstein, S.R.; Celis, M. (Eds.) Catálogo de Plantas y Líquenes de Colombia; Instituto de Ciencias Naturales, Universidad Nacional de Colombia: Bogotá, Colombia, 2019. [Google Scholar]
- Mamun-Or-Rashid, A.N.M.; Sen, M.K.; Jamal, M.A.H.; Nasrin, S. A Comprehensive Ethnopharmacological Review on Lippia alba M. Int. J. Biomed. Mater. Res. 2013, 1, 14–20. [Google Scholar]
- Oliveira, D.R.; Leitão, G.G.; Fernandes, P.D.; Leitão, S.G. Ethnopharmacological studies of Lippia origanoides. Rev. Bras. Farmacogn. 2014, 24, 206–214. [Google Scholar] [CrossRef]
- Szewczyk, K.; Zidorn, C. Ethnobotany, phytochemistry, and bioactivity of the genus Turnera (Passifloraceae) with a focus on damiana—Turnera diffusa. J. Ethnopharmacol. 2014, 152, 424–443. [Google Scholar] [CrossRef]
- Durant-Archibold, A.A.; Santana, A.I.; Gupta, M.P. Ethnomedical uses and pharmacological activities of most prevalent species of genus Piper in Panama: A review. J. Ethnopharmacol. 2018, 217, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Oza, M.J.; Kulkarni, Y.A. Traditional uses, phytochemistry and pharmacology of the medicinal species of the genus Cordia (Boraginaceae). J. Pharm. Pharmacol. 2017, 69, 755–789. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Ismail, T.; Calcabrini, C.; Guescini, M.; Catanzaro, E.; Turrini, E.; Layla, A.; Akhtar, S.; Fimognari, C. The potential effects of Ocimum basilicum on health: A review of pharmacological and toxicological studies. Expert Opin. Drug Metab. Toxicol. 2018, 14, 679–692. [Google Scholar] [CrossRef]
- Yuan, Y.; Huang, M.; Pang, Y.X.; Yu, F.L.; Chen, C.; Liu, L.W.; Chen, Z.X.; Zhang, Y.B.; Chen, X.L.; Hu, X. Variations in essential oil yield, composition, and antioxidant activity of different plant organs from Blumea balsamifera (L.) DC. at different growth times. Molecules 2016, 21, 1024. [Google Scholar] [CrossRef] [PubMed]
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 1932633219. [Google Scholar]
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention Indices for frequently reported compounds of plant essential oils. J. Phys. Chem. Ref. Data 2011, 40, 1–47. [Google Scholar] [CrossRef]
- NIST Database, Version 2.3.; National Institute of Standards and Technology: Gaithersburg, MD, USA,, 2017.
- Smee, D.F.; Hurst, B.L.; Evans, W.J.; Clyde, N.; Wright, S.; Peterson, C.; Jung, K.H.; Day, C.W. Evaluation of cell viability dyes in antiviral assays with RNA viruses that exhibit different cytopathogenic properties. J. Virol. Methods 2017, 246, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Nasar, S.; Rashid, N.; Iftikhar, S. Dengue proteins with their role in pathogenesis, and strategies for developing an effective anti-dengue treatment: A review. J. Med. Virol. 2020, 92, 941–955. [Google Scholar] [CrossRef]
- Shityakov, S.; Förster, C. In silico predictive model to determine vector-mediated transport properties for the blood-brain barrier choline transporter. Adv. Appl. Bioinforma. Chem. 2014, 7, 23–36. [Google Scholar] [CrossRef]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. Structure of the dengue virus envelope protein after membrane fusion. Nature 2004, 427, 313–319. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Martínez, J.R.; Cala, M.P.; Durán, D.C.; Caballero, D. Chromatographic and mass spectrometric characterization of essential oils and extracts from Lippia (Verbenaceae) aromatic plants. J. Sep. Sci. 2012, 36, 192–202. [Google Scholar] [CrossRef]
- Vicuña, G.C.; Stashenko, E.E.; Fuentes, J.L. Chemical composition of the Lippia origanoides essential oils and their antigenotoxicity against bleomycin-induced DNA damage. Fitoterapia 2010, 81, 343–349. [Google Scholar] [CrossRef]
- Bueno, J.; Escobar, P.; Martínez, J.R.; Leal, S.M.; Stashenko, E.E. Composition of three essential oils, and their mammalian cell toxicity and antimycobacterial activity against drug resistant-tuberculosis and nontuberculous mycobacteria strains. Nat. Prod. Commun. 2011, 6, 1743–1748. [Google Scholar] [CrossRef]
- Monzote, L.; Scull, R.; Cos, P.; Setzer, W. Essential oil from Piper aduncum: Chemical analysis, antimicrobial assessment, and literature review. Medicines 2017, 4, 49. [Google Scholar] [CrossRef]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Khan, T.; Sharifi-Rad, J.; Ozleyen, A.; Turkdonmez, E.; et al. Piper species: A comprehensive review on their phytochemistry, biological activities and applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef]
- Gurav, T.P.; Dholakia, B.B.; Giri, A.P. A glance at the chemodiversity of Ocimum species: Trends, implications, and strategies for the quality and yield improvement of essential oil. Phytochem. Rev. 2021, 21, 879–913. [Google Scholar] [CrossRef]
- Marques, A.P.S.; Bonfim, F.P.G.; Dantas, W.F.C.; Puppi, R.J.; Marques, M.O.M. Chemical composition of essential oil from Varronia curassavica Jacq. accessions in different seasons of the year. Ind. Crops Prod. 2019, 140, 111656. [Google Scholar] [CrossRef]
- Grienke, U.; Mair, C.E.; Kirchmair, J.; Schmidtke, M.; Rollinger, J.M. Discovery of bioactive natural products for the treatment of acute respiratory infections—An integrated approach. Planta Med. 2018, 84, 684–695. [Google Scholar] [CrossRef]
- Ocazionez, R.E.; Meneses, R.; Torres, F.Á.; Stashenko, E. Virucidal activity of Colombian Lippia essential oils on dengue virus replication in vitro. Mem. Inst. Oswaldo Cruz 2010, 105, 304–309. [Google Scholar] [CrossRef]
- Meneses, R.; Ocazionez, R.E.; Martínez, J.R.; Stashenko, E.E. Inhibitory effect of essential oils obtained from plants grown in Colombia on yellow fever virus replication in vitro. Ann. Clin. Microbiol. Antimicrob. 2009, 8, 8. [Google Scholar]
- Tshilanda, D.D.; Ngoyi, E.M.; Kabengele, C.N.; Matondo, A.; Bongo, G.N.; Inkoto, C.L.; Mbadiko, C.M.; Gbolo, B.Z.; Lengbiye, E.M.; Kilembe, J.T.; et al. Ocimum species as potential bioresources against COVID-19: A review of their phytochemistry and antiviral activity. Int. J. Pathog. Res. 2020, 5, 42–54. [Google Scholar] [CrossRef]
- Kubiça, T.F.; Alves, S.H.; Weiblen, R.; Lovato, L.T. In vitro inhibition of the bovine viral diarrhoea virus by the essential oil of Ocimum basilicum (basil) and monoterpenes. Braz. J. Microbiol. 2014, 45, 209–214. [Google Scholar] [CrossRef][Green Version]
- Singh, P.; Chakraborty, P.; He, D.H.; Mergia, A. Extract prepared from the leaves of Ocimum basilicum inhibits the entry of Zika Virus. Acta Virol. 2019, 63, 316–321. [Google Scholar] [CrossRef]
- Lin, Q.; Lim, J.Y.; Xue, K.; Yew, P.Y.; Owh, C.; Chee, P.L.; Loh, X.J. Sanitizing agents for virus inactivation and disinfection. View 2020, 1, e16. [Google Scholar] [CrossRef]
- Cagno, V.; Sgorbini, B.; Sanna, C.; Cagliero, C.; Ballero, M.; Civra, A.; Donalisio, M.; Bicchi, C.; Lembo, D.; Rubiolo, P. In vitro anti-herpes simplex virus-2 activity of Salvia desoleana Atzei & V. Picci essential oil. PLoS ONE 2017, 12, e0172322. [Google Scholar]
- Pliego Zamora, A.; Edmonds, J.H.; Reynolds, M.J.; Khromykh, A.A.; Ralph, S.J. The in vitro and in vivo antiviral properties of combined monoterpene alcohols against West Nile virus infection. Virology 2016, 495, 18–32. [Google Scholar] [CrossRef]
- Nogueira Sobrinho, A.C.; de Morais, S.M.; Marinho, M.M.; de Souza, N.V.; Lima, D.M. Antiviral activity on the Zika virus and larvicidal activity on the Aedes spp. of Lippia alba essential oil and β-caryophyllene. Ind. Crops Prod. 2021, 162, 11381. [Google Scholar] [CrossRef]
- Yarovaya, O.I.; Salakhutdinov, N.F. Mono- and sesquiterpenes as a starting platform for the development of antiviral drugs. Russ. Chem. Rev. 2021, 90, 488–510. [Google Scholar] [CrossRef]
- Naresh, P.; Selvaraj, A.; Shyam Sundar, P.; Murugesan, S.; Sathianarayananan, S.; Namboori, P.K.K.; Jubie, S. Targeting a conserved pocket (n-octyl-β-D–glucoside) on the dengue virus envelope protein by small bioactive molecule inhibitors. J. Biomol. Struct. Dyn. 2022, 40, 4866–4878. [Google Scholar] [CrossRef]
- Flechas, M.C.; Ocazionez, R.E.; Stashenko, E.E. Evaluation of in vitro antiviral activity of essential oil compounds against Dengue Virus. Pharmacogn. J. 2018, 10, 55–59. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Kaihatsu, K.; Yamabe, M.; Ebara, Y. Antiviral mechanism of action of epigallocatechin-3-O-gallate and its fatty acid esters. Molecules 2018, 23, 2475. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A. Autodock Vina: Improving the speed and accuracy of docking. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed]
- Kohonen, T.; Somervuo, P. How to make large self-organizing maps for nonvectorial data. Neural Netw. 2002, 15, 945–952. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Material | Voucher Number | EO Identifier | EO Characteristics * |
|---|---|---|---|
| Lippia origanoides Kunth | 22035 | LoP | Phellandrene chemotype, neat EO |
| 22034 | LoC | Carvacrol chemotype, neat EO | |
| 22039 | LoTC | Thymol-carvacrol chemotype, neat EO | |
| 22036 | LoT | Thymol chemotype, neat EO | |
| LoTf | Thymol chemotype, thymol-rich fraction | ||
| Lippia alba (Mill.) N.E.Br. ex Britton & P. Wilson | 22002 | LaCi | Citral chemotype, neat EO |
| LaCif | Citral chemotype, light fraction | ||
| 22031 | LaCaf1 | Carvone chemotype, limonene-rich fraction | |
| LaCaf2 | Carvone chemotype, carvone-rich fraction | ||
| Turnera diffusa Willdenow | 22032 | TdS1 | 2019, neat EO |
| 22037 | TdS2 | 2016, neat EO | |
| Piper aduncum L. | 22033 | PaS1 | Linalool chemotype, neat EO |
| Ocimum basilicum L. | 22227 | ObS1 | Piperitone chemotype, neat EO |
| Varronia curassavica Jacq. | 20892 | VcS1 | Neat EO |
| Compound | LRIs (DB-5MS Column) | LaCi | LaCif | LaCaf1 | LaCaf2 | LoP | LoC | LoT | LoTf | LoTC | TdS1 | TdS2 | PaS1 | ObS1 | VcS1 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exp. | Lit. | |||||||||||||||
| α-Pinene * | 935 | 932 a | 0.3 | 0.2 | 0.2 | - | 1.7 | 0.4 | - | - | 0.4 | - | - | 4.6 | 0.5 | 9.4 |
| α-Phellandrene | 1005 | 1002 a | 0.1 | - | - | - | 7.1 | - | - | - | 0.5 | - | - | 4.4 | - | - |
| p-Cymene * | 1027 | 1024 a | - | - | - | - | 12.6 | 14.4 | 2.3 | 2.0 | 19.1 | 3.0 | 3.6 | 3.0 | - | - |
| Limonene * | 1034 | 1029 a | 2.4 | 5.9 | 82.2 | 0.9 | 2.1 | 0.3 | - | - | 0.9 | - | - | 6.0 | - | 0.8 |
| 1,8-Cineol * | 1036 | 1031 a | - | - | - | - | 13.0 | 1.3 | - | - | - | - | - | 3.6 | 8.1 | 0.8 |
| γ-Terpinene | 1061 | 1059 a | - | - | - | - | 2.4 | 5.3 | 0.9 | 6.9 | 9.2 | 0.6 | 0.7 | 0.8 | - | - |
| Linalool * | 1099 | 1096 a | 1.1 | 2.5 | 0.4 | 0.5 | 0.7 | 1.0 | - | - | 0.3 | - | - | 0.4 | 42.7 | - |
| Estragole * | 1203 | 1196 a | - | - | - | - | - | - | - | - | - | - | - | - | 18.6 | - |
| Neral * | 1246 | 1252 b | 11.9 | 18.1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Carvone * | 1259 | 1258 c | - | - | 12.2 | 78.2 | - | - | - | - | - | - | - | - | - | - |
| Geraniol * | 1260 | 1240 b | 19 | 8.1 | - | - | - | - | - | - | - | - | - | - | - | - |
| Piperitone | 1265 | 1264 c | - | - | - | 4.8 | - | - | - | - | - | - | - | 14.8 | - | - |
| Geranial * | 1272 | 1270 b | 24.5 | 24.8 | - | - | - | - | - | - | - | - | - | - | - | - |
| Thymol * | 1290 | 1290 a | - | - | - | - | 14.0 | 8.0 | 75.3 | 82.9 | 49.4 | - | 0.2 | - | - | - |
| Carvacrol * | 1300 | 1298 a | - | - | - | - | 0.9 | 35 | 4.9 | 1.2 | 2.7 | - | 0.4 | - | - | - |
| Piperitenone | 1347 | 1343 a | - | - | 0.3 | 0.3 | - | - | - | - | - | - | - | 14.8 | - | - |
| α-Copaene | 1385 | 1376 a | - | - | - | 0.3 | 0.6 | 0.7 | - | - | - | - | - | 2.9 | - | 7.0 |
| trans-β-Caryophyllene * | 1433 | 1427 c | 9.1 | 13.3 | - | 0.1 | 15.1 | 4.4 | 5.4 | 7.0 | 1.6 | 4.0 | 4.9 | 7.4 | 0.9 | 19.2 |
| α-Humulene * | 1467 | 1468 c | 2.8 | 2.8 | - | - | 8.1 | 1.1 | 3.2 | - | 0.9 | - | 0.4 | 1.5 | 2.5 | 2.7 |
| Aristolochene | 1483 | 1488 a | - | - | - | - | - | - | - | - | - | 17.9 | 20.9 | - | - | - |
| Germacrene D | 1492 | 1481 a | 4.3 | 1.5 | 0.1 | - | 0.9 | - | - | - | - | - | - | 1.7 | 4.9 | 12.3 |
| β-Selinene | 1502 | 1490 a | - | - | - | - | 0.5 | 0.3 | - | - | - | 5.2 | 5.8 | - | - | - |
| Valencene | 1503 | 1496 a | - | - | - | - | - | - | - | - | - | 7.4 | 6.5 | 1.2 | - | - |
| trans-β-Guaiene | 1517 | 1502 a | 2.2 | - | - | - | - | - | - | - | - | - | - | - | - | 11.8 |
| Dehydrofukinone | 1827 | 1820 c | - | - | - | - | - | - | - | - | - | 25.4 | 19.3 | - | - | - |
| Plant Material | Essential Oil Identifier | DENV-1 | DENV-2 |
|---|---|---|---|
| Lippia origanoides | LoP | Strong: IC50 (SI): 77 ± 1.1 (6.6) | Strong: IC50 (SI): 75 ± 1.0 (6.8) |
| LoC | Weak | Weak | |
| LoTC | Weak | Weak | |
| LoTf | Weak | Weak | |
| LoT | Inactive | Inactive | |
| Lippia alba | LaCi | Strong: IC50 (SI): 78 ± 1.1 (5.5) | Strong: IC50 (SI): 67 ± 1.2 (6.4) |
| LaCaf2 | Strong: IC50 (SI): 82 ± 1.1 (5.8) | Strong: IC50 (SI): 72 ± 1.1 (6.6) | |
| LaCaf1 | Weak | Weak | |
| LaCif | Inactive | Inactive | |
| Turnera diffusa | TdS2 | Strong: IC50 (SI): 54 ± 1.1 (7.7) | Strong: IC50 (SI): 29 ± 1.1 (14.3) |
| TdS1 | Inactive | Inactive | |
| Piper aduncum | PaS1 | Weak | Weak |
| Ocimum basilucum | ObS1 | Weak | Weak |
| Verronia curassavica | VcS1 | Inactive | Inactive |
| No | Compound | Kcal/mol | Structural Formula | Amino Acid Residues. H-bond in Bold Font |
|---|---|---|---|---|
| 1 | cis-Calamenene | −8.73 |  | Thr189, Leu19, Phe193, Leu167, Phe279. |
| 2 | δ-Cadinene | −8.41 |  | Leu45, Thr48, Leu135, Tyr137, Thr189, Leu191, Phe193, Phe279, Leu283. |
| 3 | α-Cadinene | −8.28 |  | Leu45, Thr48, Leu135, Tyr137, Leu191, Phe193, Phe279, Leu283. |
| 4 | α-Guaiene | −8.26 |  | Glu26, Leu45, Thr48, Tyr137, Leu191, Phe193, Phe279, Leu283. |
| 5 | γ-Cadinene | −8.19 |  | Leu45, Thr48, Leu135, Tyr137, Leu191, Phe193, Ohe279, Leu283. |
| 6 | Viridiflorene | −8.13 |  | Thr48, Leu135, Tyr137, Thr189, Leu191, Phe193, Leu207, Phe279, Leu283. |
| 7 | α-Selinene | −7.98 |  | Thr48, Leu135, Thr189, Leu191, Phe193, Phe193, Phe279, Leu283. |
| 8 | δ-Amorphene | −7.96 |  | Leu45. Thr48, Leu135, Tyr137, Leu191, Phe193, Phe279, Leu283. |
| 9 | β-Bourbonene | −7.95 |  | Thr48, Val130, Leu135, Tyr137, Leu191, Phe193, Leu207, Phe279, Leu283. |
| 10 | α-Gurjunene | −7.83 |  | Thr48, Val130, Leu135, Tyr137, Thr189, Leu191, Phe193, Leu207, Phe279. |
| 11 | α-Phellandrene | −7.60 |  | Thr48, Leu135, Tyr137, Thr189, Phe193, Phe279, Leu283 |
| 12 | Carvacrol | −7.31 |  | Thr189, Leu191, Leu207, Phe279, Leu283, His282 |
| 13 | Carvone | −7.29 |  | Thr48, Thr189, Leu191, Phe193, Leu207, Phe279. |
| 14 | γ-Terpinene | −7.28 |  | Thr48, Val130, Leu191, Leu207, Phe279, Leu283. |
| 15 | p-Cymene | −7.27 |  | Thr48, Val130, Leu191, Leu207, Phe279, Leu283. |
| 16 | Limonene | −7.24 |  | Thr48, Leu135, Phe193, Phe279, Leu283. |
| 17 | trans-Dihydrocarvone | −7.23 |  | His27, Thr48, Leu191, Phe193, Leu207, Phe279. |
| 18 | Thymol-methyl-ether | −7.11 |  | Leu191, Phe193, Leu207, Phe279. |
| 19 | α-Terpinene | −7.10 |  | Thr48, Val130, Leu191, Phe279, Leu283. |
| 20 | Terpinolene | −7.03 |  | Thr48, Leu135, Thr189, Phe193, Leu207, Leu277, Phe279. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Trujillo, L.; Quintero-Rueda, E.; Stashenko, E.E.; Conde-Ocazionez, S.; Rondón-Villarreal, P.; Ocazionez, R.E. Essential Oils from Colombian Plants: Antiviral Potential against Dengue Virus Based on Chemical Composition, In Vitro and In Silico Analyses. Molecules 2022, 27, 6844. https://doi.org/10.3390/molecules27206844
Silva-Trujillo L, Quintero-Rueda E, Stashenko EE, Conde-Ocazionez S, Rondón-Villarreal P, Ocazionez RE. Essential Oils from Colombian Plants: Antiviral Potential against Dengue Virus Based on Chemical Composition, In Vitro and In Silico Analyses. Molecules. 2022; 27(20):6844. https://doi.org/10.3390/molecules27206844
Chicago/Turabian StyleSilva-Trujillo, Lina, Elizabeth Quintero-Rueda, Elena E. Stashenko, Sergio Conde-Ocazionez, Paola Rondón-Villarreal, and Raquel E. Ocazionez. 2022. "Essential Oils from Colombian Plants: Antiviral Potential against Dengue Virus Based on Chemical Composition, In Vitro and In Silico Analyses" Molecules 27, no. 20: 6844. https://doi.org/10.3390/molecules27206844
APA StyleSilva-Trujillo, L., Quintero-Rueda, E., Stashenko, E. E., Conde-Ocazionez, S., Rondón-Villarreal, P., & Ocazionez, R. E. (2022). Essential Oils from Colombian Plants: Antiviral Potential against Dengue Virus Based on Chemical Composition, In Vitro and In Silico Analyses. Molecules, 27(20), 6844. https://doi.org/10.3390/molecules27206844