
Factors Impacting Invader-Mediated Recognition of Double-Stranded DNA

Abstract
1. Introduction
2. Results and Discussion
2.1. Invader Probe Design
2.2. Thermal Denaturation Properties of Invader Probes
2.3. Driving Force for Recognition of dsDNA Targets
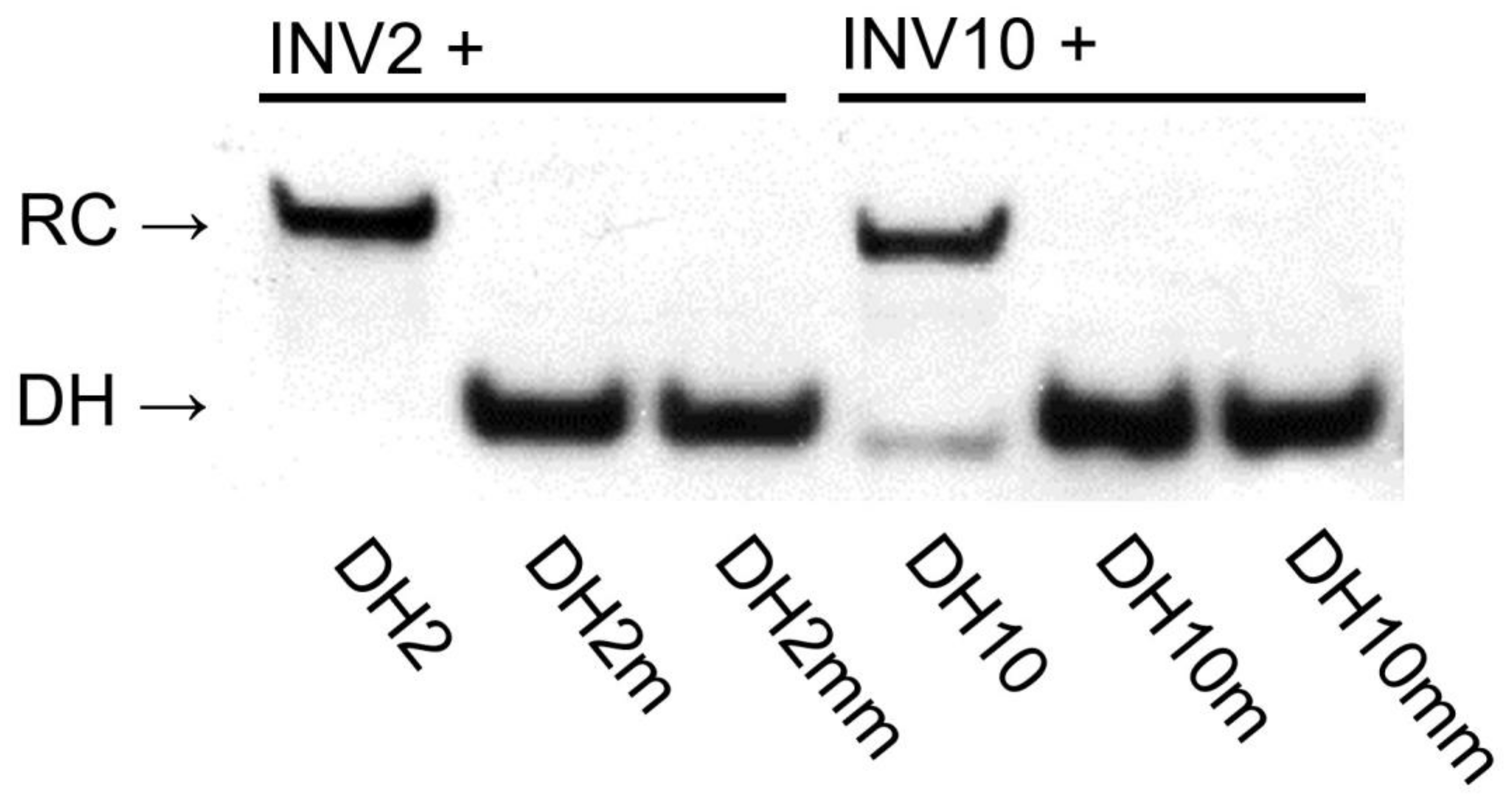
2.4. Recognition of Mixed-Sequence Model DNA Hairpin Targets
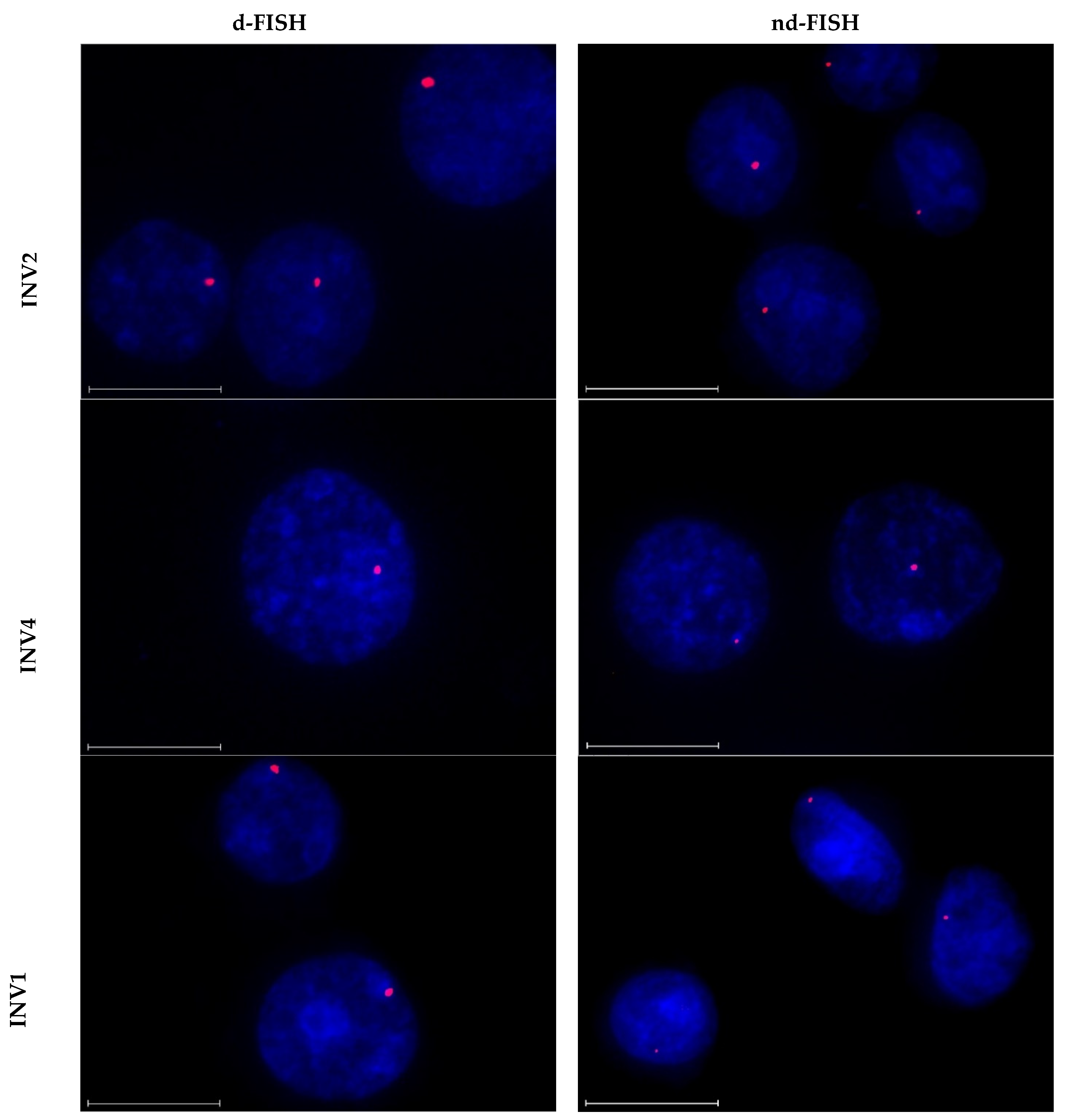
2.5. Targeting Chromosomal DNA—Fluorescence In Situ Hybridization (FISH) Assays
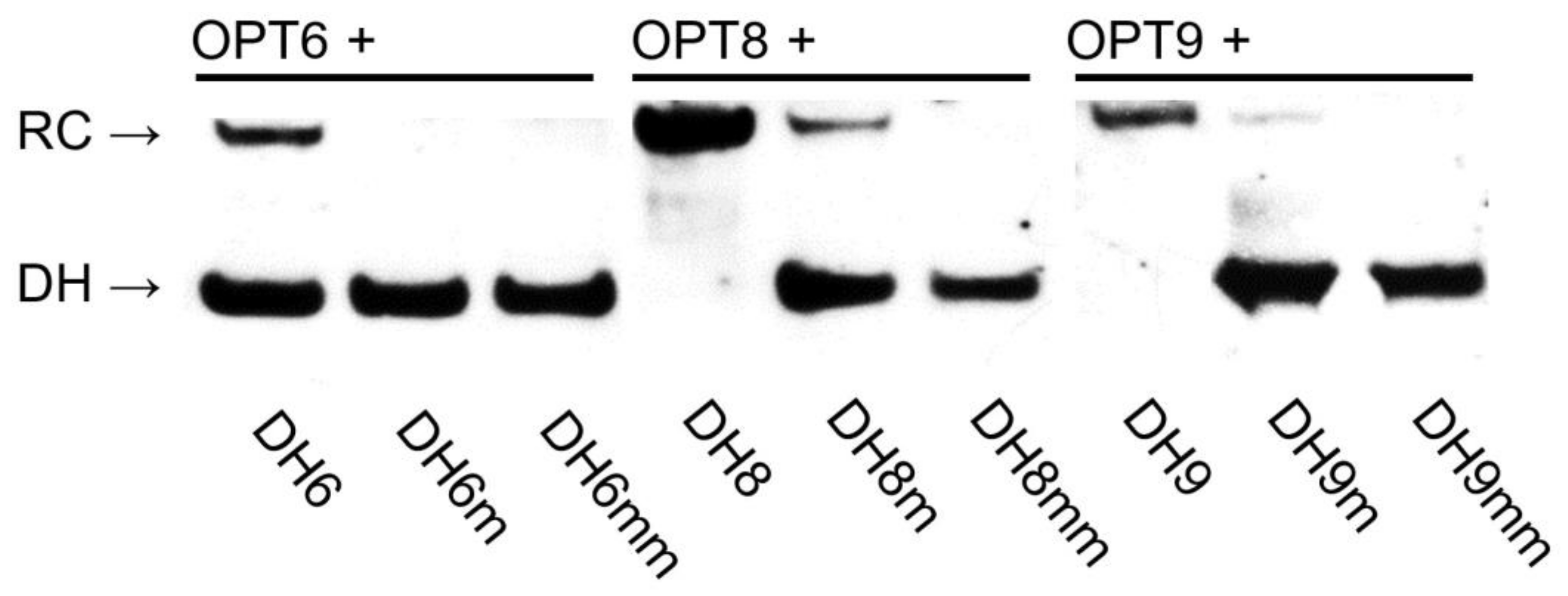
2.6. Design of Optimized Invader Probes
2.7. Thermal Denaturation and Thermodynamic Properties of Optimized Invader Probes
2.8. Recognition of Model DNA Hairpin Targets by Optimized Invader Probes
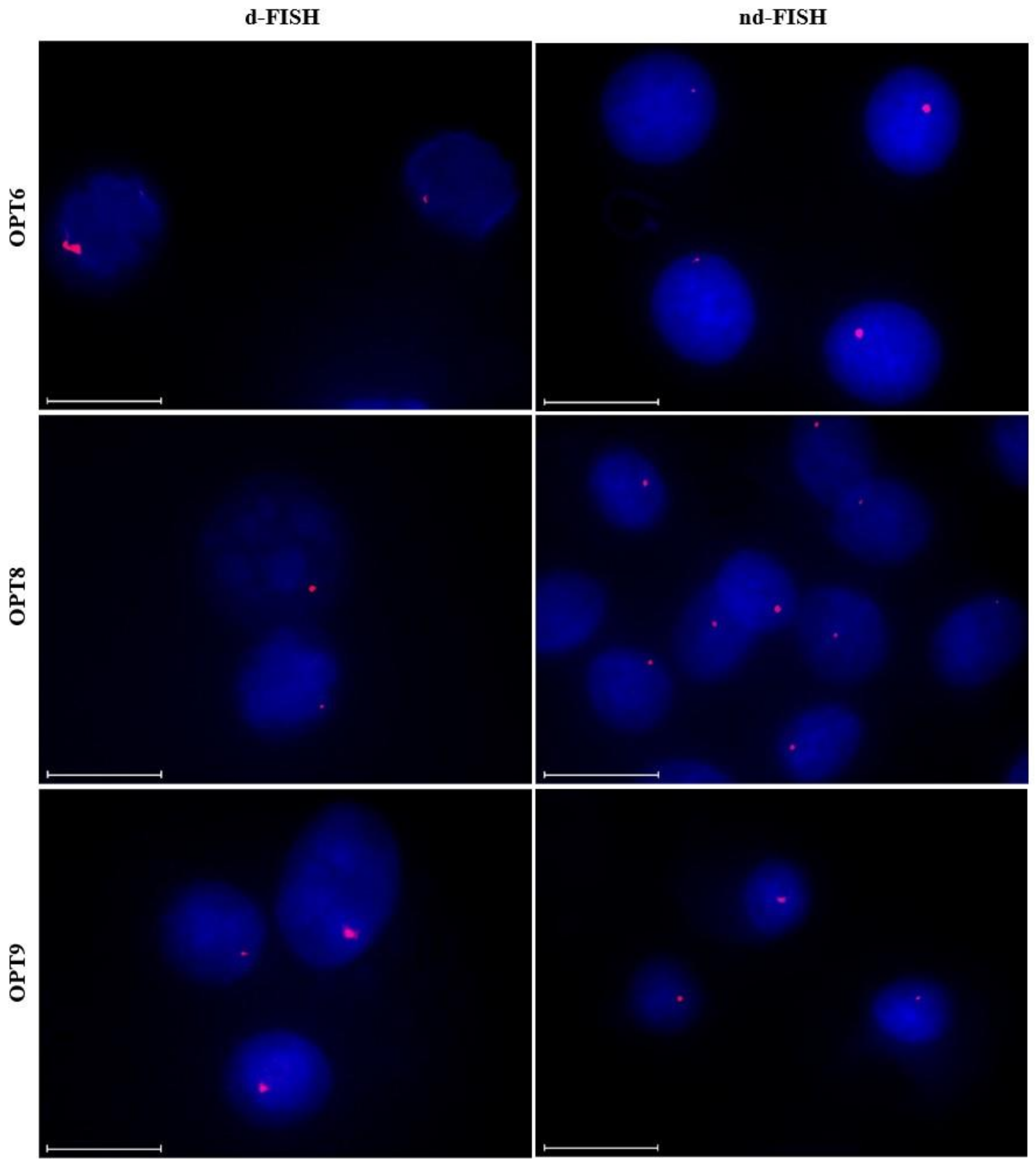
2.9. Targeting Chromosomal DNA using Optimized Invader Probes
3. Materials and Methods
3.1. Synthesis and Purification of Probe Strands
3.2. Thermal Denaturation Experiments
3.3. Electrophoretic Mobility Shift Assays
3.4. Spearman Rank-Order Correlation Analysis
3.5. Cell Culture and Nuclei Preparation
3.6. Preparation of Slides for FISH Assays
3.7. Fluorescence In Situ Hybridization Experiments and Image Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References and Notes
- Muangkaew, P.; Vilaivan, T. Modulation of DNA and RNA by PNA. Bioorg. Med. Chem. Lett. 2020, 30, 127064. [Google Scholar] [CrossRef] [PubMed]
- Brodyagin, N.; Katkevics, M.; Kotikam, V.; Ryan, C.A.; Rozners, E. Chemical approaches to discover the full potential of peptide nucleic acids in biomedical applications. Beilstein J. Org. Chem. 2021, 17, 1641–1688. [Google Scholar] [CrossRef] [PubMed]
- Duca, M.; Vekhoff, P.; Oussedik, K.; Halby, L.; Arimondo, P.B. The triple helix: 50 years later, the outcome. Nucleic Acids Res. 2008, 36, 5123–5138. [Google Scholar] [CrossRef] [PubMed]
- Hari, Y.; Obika, S.; Imanishi, T. Towards the sequence-selective recognition of double-stranded DNA containing pyrimidine-purine interruptions by triplex-forming oligonucleotides. Eur. J. Org. Chem. 2012, 2875–2887. [Google Scholar] [CrossRef]
- Dervan, P.B.; Edelson, B.S. Recognition of the DNA minor groove by pyrrole-imidazole polyamides. Curr. Opin. Struct. Biol. 2003, 13, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, Y.; Bando, T.; Sugiyama, H. Sequence-specific DNA binding pyrrole-imidazole polyamides and their applications. Bioorg. Med. Chem. 2018, 26, 1393–1411. [Google Scholar] [CrossRef]
- Dragulescu-Andrasi, A.; Rapireddy, S.; Frezza, B.M.; Gayathri, C.; Gil, R.R.; Ly, D.H. A simple γ-backbone modification preorganizes peptide nucleic acid into a helical structure. J. Am. Chem. Soc. 2006, 128, 10258–10267. [Google Scholar] [CrossRef]
- Bahal, R.; Sahu, B.; Rapireddy, S.; Lee, C.-M.; Ly, D.H. Sequence-unrestricted, Watson-Crick recognition of double helical B-DNA by (R)-MiniPEG-γPNAs. ChemBioChem 2012, 131, 56–60. [Google Scholar] [CrossRef]
- Bohländer, P.R.; Vilaivan, T.; Wagenknecht, H.-A. Strand displacement and duplex invasion into double-stranded DNA by pyrrolidinyl peptide nucleic acids. Org. Biomol. Chem. 2015, 13, 9223–9230. [Google Scholar] [CrossRef]
- Thadke, S.A.; Hridya, V.M.; Perera, J.D.R.; Gil, R.R.; Mukherjee, A.; Ly, D.H. Shape selective bifacial recognition of double helical DNA. Commun. Chem. 2018, 1, 79. [Google Scholar] [CrossRef]
- Zheng, H.; Botos, I.; Clausse, V.; Nikolayevskiy, H.; Rastede, E.E.; Fouz, M.F.; Mazur, S.J.; Appella, D.H. Conformational constraints of cyclopentane peptide nucleic acids facilitate tunable binding to DNA. Nucleic Acids Res. 2021, 49, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Suparpprom, C.; Vilaivan, T. Perspectives on conformationally constrained peptide nucleic acid (PNA): Insights into the structural design, properties and applications. RSC Chem. Biol. 2022, 3, 648–697. [Google Scholar] [CrossRef] [PubMed]
- Lohse, J.; Dahl, O.; Nielsen, P.E. Double duplex invasion by peptide nucleic acid: A general principle for sequence-specific targeting of double-stranded DNA. Proc. Natl. Acad. Sci. USA 1999, 96, 11804–11808. [Google Scholar] [CrossRef] [PubMed]
- Aiba, Y.; Honda, Y.; Komiyama, M. Promotion of double-duplex invasion of peptide nucleic acids through conjugation with nuclear localization signal peptide. Chem. Eur. J. 2015, 21, 4021–4026. [Google Scholar] [CrossRef] [PubMed]
- Aiba, Y.; Shibata, M.; Shoji, O. Sequence-specific recognition of double-stranded DNA by peptide nucleic acid forming double-duplex invasion complex. Appl. Sci. 2022, 12, 3677. [Google Scholar] [CrossRef]
- Filichev, V.V.; Christensen, U.B.; Pedersen, E.B.; Babu, B.R.; Wengel, J. Locked nucleic acids and intercalating nucleic acids in the design of easily denaturing nucleic acids: Thermal stability studies. ChemBioChem 2004, 5, 1673–1679. [Google Scholar] [CrossRef] [PubMed]
- Filichev, V.V.; Vester, B.; Hansen, L.H.; Pedersen, E.B. Easily denaturing nucleic acids derived from intercalating nucleic acids: Thermal stability studies, dual duplex invasion and inhibition of transcription start. Nucleic Acids Res. 2005, 33, 7129–7137. [Google Scholar] [CrossRef]
- Yotapan, N.; Nim-anussornkul, D.; Vilaivan, T. Pyrrolidinyl peptide nucleic acid terminally labeled with fluorophore and end-stacking quencher as a probe for highly specific DNA sequence discrimination. Tetrahedron 2016, 72, 7992–7999. [Google Scholar] [CrossRef]
- Asanuma, H.; Niwa, R.; Akahane, M.; Murayama, K.; Kashida, H.; Kamiya, Y. Strand-invading linear probe combined with unmodified PNA. Bioorg. Med. Chem. 2016, 24, 4129–4137. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Kawabata, H.; Fujimoto, K. Double duplex invasion of DNA induced by ultrafast photo-cross-linking using 3-cyanovinylcarbazole for antigene methods. Chem. Commun. 2017, 53, 7616–7619. [Google Scholar] [CrossRef]
- Hibino, M.; Aiba, Y.; Shoji, O. Cationic guanine: Positively charged nucleobase with improved DNA affinity inhibits self-duplex formation. Chem. Commun. 2020, 56, 2546–2549. [Google Scholar] [CrossRef] [PubMed]
- Emehiser, R.G.; Hrdlicka, P.J. Chimeric γPNA–Invader probes: Using intercalator-functionalized oligonucleotides to enhance the DNA-targeting properties of γPNA. Org. Biomol. Chem. 2020, 18, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Emehiser, R.G.; Dhuri, K.; Shepard, C.; Karmakar, S.; Bahal, R.; Hrdlicka, P.J. Serine-γPNA, Invader probes, and chimeras thereof: Three probe chemistries that enable sequence-unrestricted recognition of double-stranded DNA. Org. Biomol. Chem. 2022, 20, 8714–8724. [Google Scholar] [CrossRef] [PubMed]
- Guenther, D.C.; Anderson, G.H.; Karmakar, S.; Anderson, B.A.; Didion, B.A.; Guo, W.; Verstegen, J.P.; Hrdlicka, P.J. Invader probes: Harnessing the energy of intercalation to facilitate recognition of chromosomal DNA for diagnostic applications. Chem. Sci. 2015, 6, 5006–5015. [Google Scholar] [CrossRef] [PubMed]
- For a definition of the interstrand zipper nomenclature, see the Supporting Materials.
- Crothers, D.M. Calculation of binding isotherms for heterogeneous polymers. Biopolymers 1968, 6, 575–584. [Google Scholar] [CrossRef]
- Jain, S.C.; Tsai, C.; Sobell, H.M. Visualization of drug-nucleic acid interactions at atomic resolution. II. Structure of an ethidium-dinucleoside monophosphate crystalline complex, ethidium:5-iodocytidylyl (3′-5′) guanosine. J. Mol. Biol. 1977, 114, 317–331. [Google Scholar] [CrossRef]
- Williams, L.D.; Egli, M.; Gao, Q.; Rich, A. DNA intercalation: Helix unwinding and neighbor-exclusion. In Structure and Function: Nucleic Acids; Sarma, R.H., Sarma, M.H., Eds.; Adenine Press: Albany, NY, USA, 1992; Volume 1, pp. 107–125. [Google Scholar]
- Ihmels, H.; Otto, D. Intercalation of organic dye molecules into double-stranded DNA-General principles and recent developments. Top. Curr. Chem. 2005, 258, 161–204. [Google Scholar]
- Sau, S.P.; Madsen, A.S.; Podbevsek, P.; Andersen, N.K.; Kumar, T.S.; Andersen, S.; Rathje, R.L.; Anderson, B.A.; Guenther, D.C.; Karmakar, S.; et al. Identification and characterization of second-generation Invader locked nucleic acids (LNAs) for mixed-sequence recognition of double-stranded DNA. J. Org. Chem. 2013, 78, 9560–9570. [Google Scholar] [CrossRef]
- Karmakar, S.; Madsen, A.S.; Guenther, D.C.; Gibbons, B.C.; Hrdlicka, P.J. Recognition of double-stranded DNA using energetically activated duplexes with interstrand zippers of 1-, 2- or 4-pyrenyl-functionalized O2′-alkylated RNA monomers. Org. Biomol. Chem. 2014, 12, 7758–7773. [Google Scholar] [CrossRef]
- Denn, B.; Karmakar, S.; Guenther, D.C.; Hrdlicka, P.J. Sandwich assay for mixed-sequence recognition of double-stranded DNA: Invader-based detection of targets specific to foodborne pathogens. Chem. Commun. 2013, 49, 9851–9853. [Google Scholar] [CrossRef]
- Emehiser, R.G.; Hall, E.; Guenther, D.C.; Karmakar, S.; Hrdlicka, P.J. Head-to-head comparison of LNA, MPγPNA, INA and Invader probes targeting mixed-sequence double-stranded DNA. Org. Biomol. Chem. 2020, 18, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.A.; Onley, J.J.; Hrdlicka, P.J. Recognition of double-stranded DNA using energetically activated duplexes modified with N2′-pyrene-, perylene-, or coronene-functionalized 2′-N-methyl-2′-amino-DNA monomers. J. Org. Chem. 2015, 80, 5395–5406. [Google Scholar] [CrossRef] [PubMed]
- Guenther, D.C.; Karmakar, S.; Hrdlicka, P.J. Bulged Invader probes: Activated duplexes for mixed-sequence dsDNA recognition with improved thermodynamic and kinetic profiles. Chem. Commun. 2015, 51, 15051–15054. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.P.; Vukelich, P.; Guenther, D.C.; Karmakar, S.; Hrdlicka, P.J. Recognition of double-stranded DNA using LNA-modified toehold Invader probes. Org. Biomol. Chem. 2021, 19, 9276–9290. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Guenther, D.C.; Hrdlicka, P.J. Recognition of mixed-sequence DNA duplexes: Design guidelines for Invaders based on 2′-O-(pyren-1-yl)methyl-RNA monomers. J. Org. Chem. 2013, 78, 12040–12048. [Google Scholar] [CrossRef]
- Perret, J.; Shia, Y.C.; Fries, R.; Vassart, G.; Georges, M. A polymorphic satellite sequence maps to the pericentric region of the bovine Y-chromosome. Genomics 1990, 6, 482–490. [Google Scholar] [CrossRef]
- Mergny, J.L.; Lacroix, L. Analysis of thermal melting curves. Oligonucleotides 2003, 13, 515–537. [Google Scholar] [CrossRef]
- The driving force for dsDNA-recognition with INV4 and INV9 is due to favorable changes in entropy.
- Preliminary studies suggested that shorter incubation (i.e., 2.5 h) resulted in incomplete recognition for some probes. For more details, see the Supporting Materials.
- Stadlbauer, S.; Kührová, P.; Wicherek, L.; Banáš, P.; Otyepla, M.; Trantírek, L.; Šponer, J. Parallel G-triplexes and G-hairpins as potential transitory ensembles in the folding of parallel-stranded DNA G-quadruplexes. Nucleic Acids Res. 2019, 47, 7276–7293. [Google Scholar] [CrossRef]
- Similar results were previously reported for INV4 in [24].
- Karmakar, S.; Anderson, B.A.; Rathje, R.L.; Andersen, S.; Jensen, T.B.; Nielsen, P.; Hrdlicka, P.J. High-affinity DNA targeting using readily accessible mimics of N2′-functionalized 2′-Amino-α-L-LNA. J. Org. Chem. 2011, 76, 7119–7131. [Google Scholar] [CrossRef] [PubMed]
- Dioubankova, N.N.; Malakhov, A.D.; Stetsenko, D.A.; Gait, M.J.; Volynsky, P.E.; Efremov, R.G.; Korshun, V.A. Pyrenemethyl ara-uridine-2′-carbamate: A strong interstrand excimer in the major groove of a DNA duplex. ChemBioChem 2003, 4, 841–847. [Google Scholar] [CrossRef]
- Morgan, M.A.; Okamoto, K.; Kahn, J.D.; English, D.S. Single-molecule spectroscopic determination of lac repressor-DNA loop conformation. Biophys. J. 2005, 89, 2588–2596. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.M. A step-by-step guide to non-linear regression analysis of experimental data using a Microsoft Excel spreadsheet. Comput. Meth. Prog. Biomed. 2001, 65, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.real-statistics.com/free-download/real-statistics-resource-pack/ (accessed on 12 December 2022).
- For example, it would only be possible to add one more hotspot to INV4, i.e., the 5′-TG-3′ step at position 8-9 to give a hotspot content of ~29%.










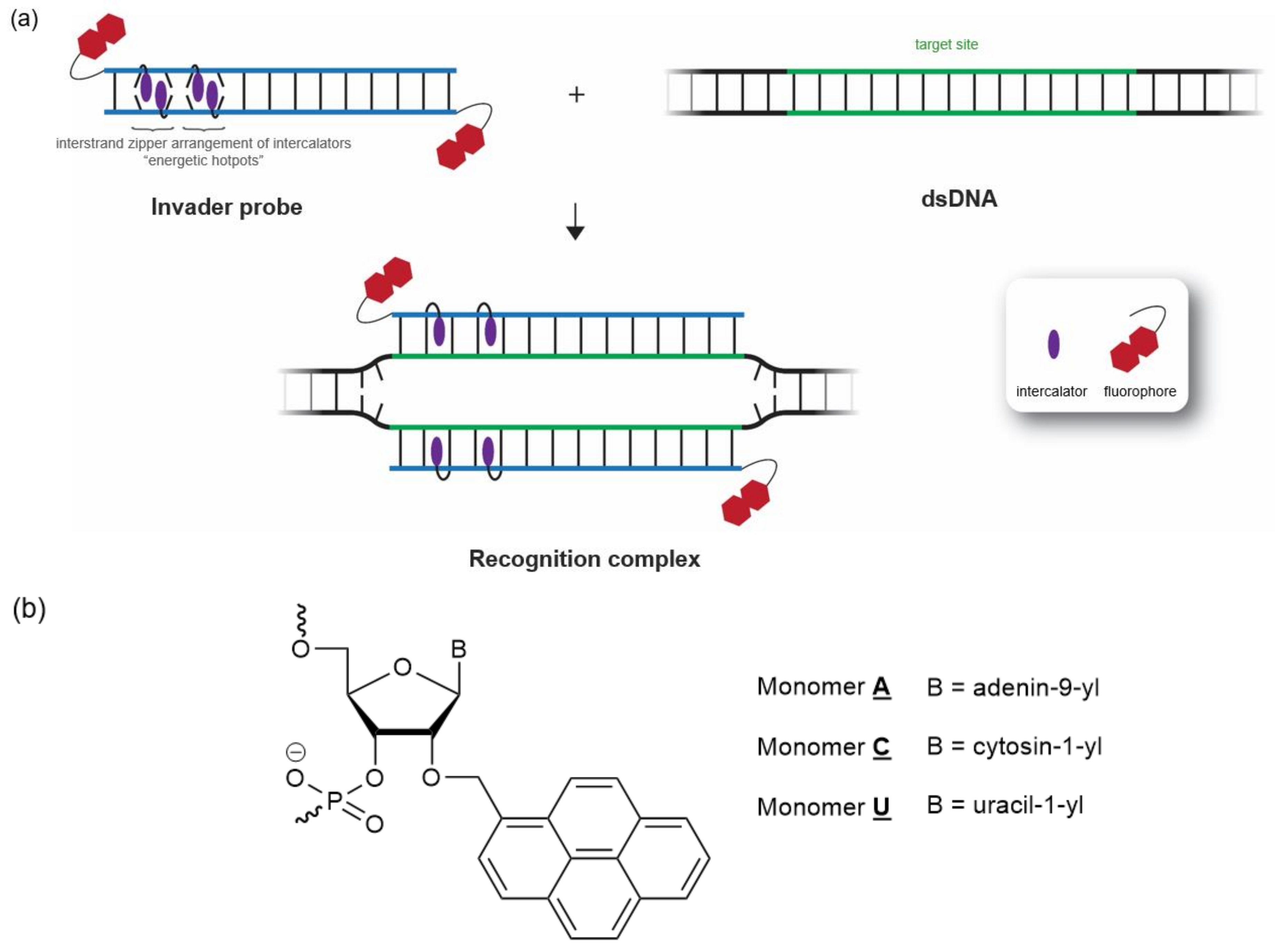{kind=link}
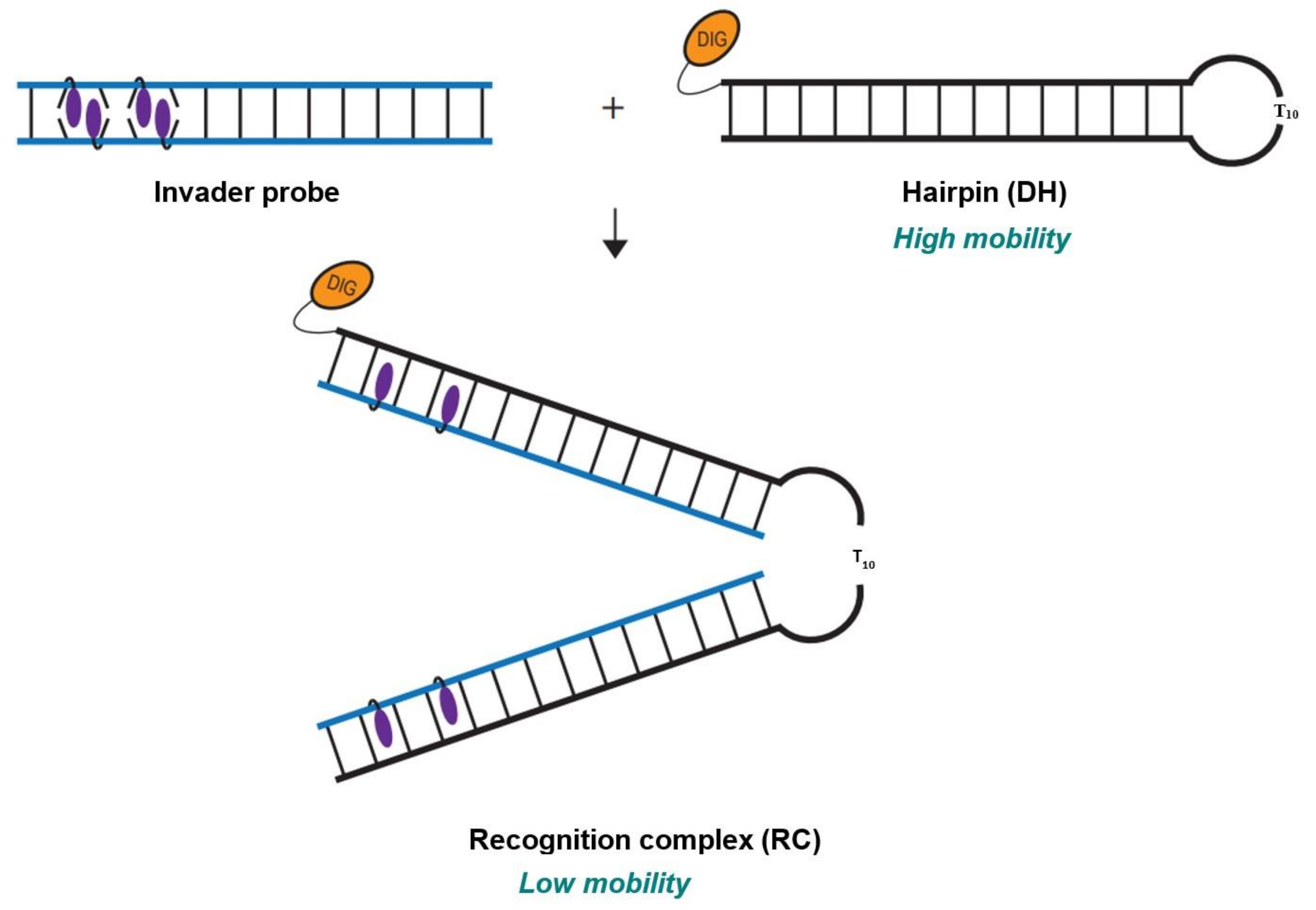{kind=link}
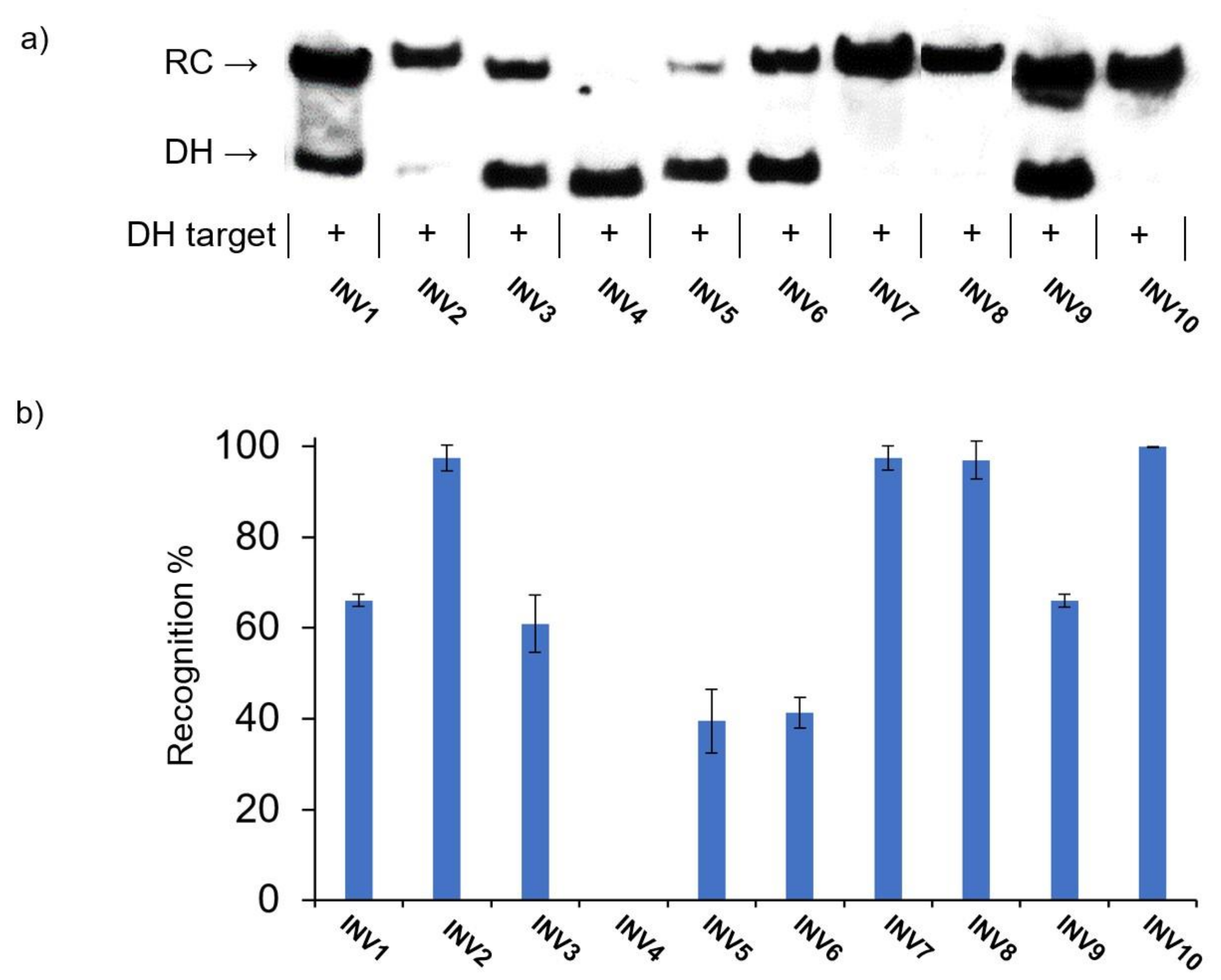{kind=link}
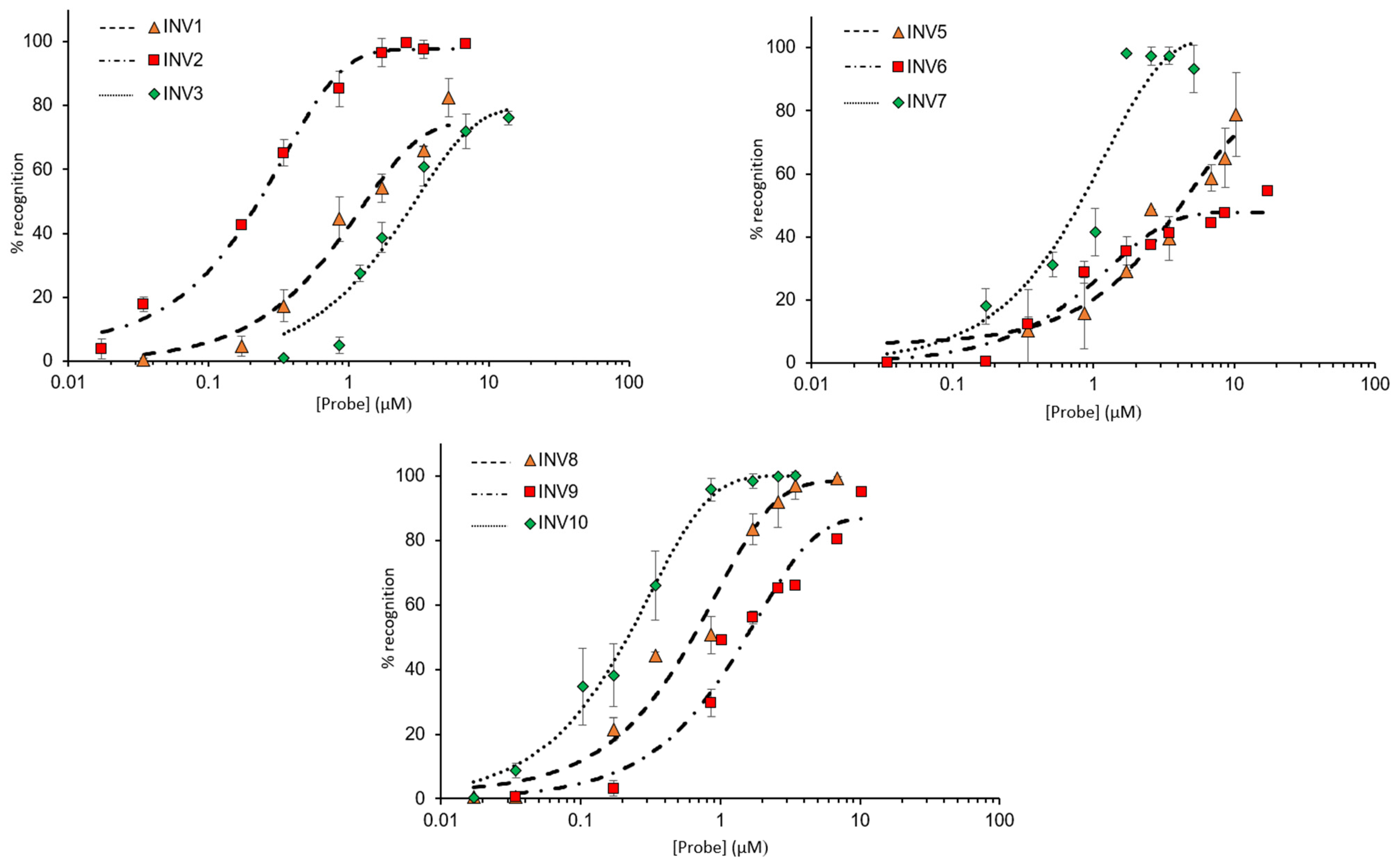{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| . | . | Tm [ΔTm] (°C) | . | |||||
|---|---|---|---|---|---|---|---|---|
| Probe (Length) | Sequence | Probe Duplex | 5′-ON: cDNA | 3′-ON: cDNA | TA (°C) | ΔG (kJ/mol) | mod% | GC% |
| INV1 (15) | 5′-Cy3-TUATCAGCACUGUGC-3′ | 52.0 b | 65.5 | 66.0 | +23.5 | ND | 20.0% | 46.7% |
| 3′-AAUAGTCGTGACACG-Cy3-5′ | [−4.0] | [+9.5] | [+10.0] | |||||
| INV2 (16) | 5′-Cy3-AUACUGGTTTGUGUTC-3′ | 34.5 b | 66.0 | 66.0 | +44.5 | −50 | 25.0% | 37.5% |
| 3′-TAUGACCAAACACAAG-Cy3-5′ | [−18.5] | [+13.0] | [+13.0] | |||||
| INV3 (15) | 5′-Cy3-TUGUGCCCTGGCAAC-3′ | NT | 64.0 | 62.0 | ND | ND | 20.0% | 60.0% |
| 3′-AACACGGGACCGTUG-Cy3-5′ | [+5.5] | [+3.5] | ||||||
| INV4 (14) | 5′-Cy3-AGCCCUGTGCCCTG-3′ | 61.5 | 69.5 | 75.5 | +23.0 | −7 | 21.4% | 71.4% |
| 3′-TCGGGACACGGGAC-Cy3-5′ | [+1.0] | [+9.0] | [+15.0] | |||||
| INV5 (16) | 5′-Cy3-GATTTCAGCCAUGUGC-3′ | 45.0 | 63.0 | 69.5 | +30.5 | −29 | 18.8% | 50.0% |
| 3′-CTAAAGTCGGTACACG-Cy3-5′ | [−12.0] | [+6.0] | [+12.5] | |||||
| INV6 (16) | 5′-Cy3-CUGUGCAACTGGTUTG-3′ | 63.0 | 65.5 | 69.0 | +13.5 | −22 | 18.8% | 50.0% |
| 3′-GACACGTTGACCAAAC-Cy3-5′ | [+5.0] | [+7.5] | [+11.0] | |||||
| INV7 (16) | 5′-Cy3-CUGUGCAAUATTTUGT-3′ | 55.0 | 73.0 | 71.0 | +38.0 | −46 | 25.0% | 31.3% |
| 3′-GACACGTTAUAAAACA-Cy3-5′ | [+4.0] | [+22.0] | [+20.0] | |||||
| INV8 (15) | 5′-Cy3-TTCACAGCCCUGUGC-3′ | 58.5 b | 70.5 | 74.5 | +26.5 | −52 | 20.0% | 60.0% |
| 3′-AAGUGTCGGGACACG-Cy3-5′ | [−1.5] | [+10.5] | [+14.5] | |||||
| INV9 (15) | 5′-Cy3-TUAUATGCTGUTCTC-3′ | 55.0 | 58.0 | 64.0 | +21.5 | −19 | 20.0% | 33.3% |
| 3′-AAUAUACGACAAGAG-Cy3-5′ | [+9.5] | [+12.5] | [+18.5] | |||||
| INV10 (14) | 5′-Cy3-GUGUAGTGUAUATG-3′ | 45.5 | 65.0 | 64.5 | +40.5 | −56 | 28.6% | 35.7% |
| 3′-CACAUCACAUAUAC-Cy3-5′ | [+2.0] | [+21.5] | [+21.0] | |||||
| OPT6 (16) | 5′-Cy3-CUGUGCAACUGGTUTG-3′ | 49.0 | 75.0 | 75.0 | +43.0 | −84 | 31.3% | 50.0% |
| 3′-GACACGUTGACCAAAC-Cy3-5′ | [−9.0] | [+17.0] | [+17.0] | |||||
| OPT8 (15) | 5′-Cy3-TTCACAGCCCUGUGC-3′ | 38.0 b | 77.0 | 76.5 | +55.5 | −93 | 26.7% | 60.0% |
| 3′-AAGUGUCGGGACACG-Cy3-5′ | [−22.0] | [+17.0] | [+16.5] | |||||
| OPT9 (15) | 5′-Cy3-TUAUAUGCUGUTCTC-3′ | 29.0 b | 65.0 | 64.0 | +54.5 | −59 | 33.3% | 33.3% |
| 3′-AAUAUACGACAAGAG-Cy3-5′ | [−16.5] | [+19.5] | [+18.5] | |||||
| Entry | Parameter Pair | Correlation Coefficient rs | p-Value |
|---|---|---|---|
| 1 | probe duplex ΔTm × length | −0.169 | 0.664 |
| 2 | probe duplex ΔTm × GC% | −0.328 | 0.388 |
| 3 | probe duplex ΔTm × mod% | −0.037 | 0.924 |
| 4 | probe duplex ΔTm × #mod | −0.091 | 0.815 |
| 5 | probe duplex ΔTm × stretch | −0.244 | 0.526 |
| 6 | 5′-ON:cDNA ΔTm × mod% | 0.774 | 0.009 |
| 7 | 3′-ON:cDNA ΔTm × mod% | 0.661 | 0.037 |
| 8 | 5′-ON:cDNA ΔTm × stretch | −0.810 | 0.005 |
| 9 | 3′-ON:cDNA ΔTm × stretch | −0.853 | 0.002 |
| 10 | 5′-ON:cDNA ΔTm × GC% | −0.762 | 0.010 |
| 11 | 3′-ON:cDNA ΔTm × GC% | −0.518 | 0.125 |
| 12 | 5′-ON:cDNA ΔTm × dsDNA Tm | −0.697 | 0.025 |
| 13 | 3′-ON:cDNA ΔTm × dsDNA Tm | −0.515 | 0.128 |
| 14 | TA × probe duplex ΔTm | −0.567 | 0.112 |
| 15 | TA × 5′-ON:cDNA ΔTm | 0.583 | 0.099 |
| 16 | TA × 3′-ON:cDNA ΔTm | 0.333 | 0.381 |
| 17 | TA × probe duplex Tm | −0.778 | 0.014 |
| 18 | TA × GC% | −0.359 | 0.343 |
| 19 | TA × mod% | 0.662 | 0.052 |
| 20 | TA × #mod | 0.822 | 0.007 |
| 21 | ΔG × TA | −0.814 | 0.014 |
| 22 | ΔG × mod% | −0.583 | 0.129 |
| 23 | ΔG × #mod | −0.620 | 0.101 |
| Probe | Rec100× (%) | C50 (µM) |
|---|---|---|
| INV1 | 66 ± 1.3 | 1.3 |
| INV2 | 97 ± 2.8 | 0.2 |
| INV3 | 60 ± 6.2 | 2.9 |
| INV4 | <5 | ND |
| INV5 | 39 ± 7.0 | 4.1 |
| INV6 | 41 ± 3.4 | >10 |
| INV7 | 97 ± 2.6 | 0.7 |
| INV8 | 96 ± 4.2 | 0.6 |
| INV9 | 66 ± 1.4 | 1.5 |
| INV10 | 99 ± 0.0 | 0.2 |
| OPT6 | 42 ± 3.4 | >10 |
| OPT8 | 99 ± 0.4 | 0.2 |
| OPT9 | 94 ± 8.9 | 0.6 |
| Entry | Parameter Pair | Correlation Coefficient rs | p-Value |
|---|---|---|---|
| 1 | C50 × mod% | −0.850 | 0.008 |
| 2 | C50 × #mod | −0.732 | 0.039 |
| 3 | C50 × stretch | 0.735 | 0.038 |
| 4 | C50 × TA | −0.598 | 0.156 |
| 5 | C50 × ΔG | 0.772 | 0.072 |
| 6 | C50 × 5′-ON:cDNA ΔTm | −0.782 | 0.022 |
| 7 | C50 × 3′-ON:cDNA ΔTm | −0.566 | 0.144 |
| 8 | C50 × 5′-ON:cDNA ΔG310 | 0.604 | 0.113 |
| 9 | C50 × 3′-ON:cDNA ΔG310 | 0.749 | 0.032 |
| 10 | C50 × probe duplex Tm | 0.032 | 0.945 |
| 11 | C50 × probe duplex ΔTm | 0.010 | 0.983 |
| 12 | C50 × probe duplex ΔG310 | −0.187 | 0.723 |
| 13 | C50 × probe duplex ΔΔG310 | 0.138 | 0.795 |
| 14 | C50 × GC% | 0.323 | 0.435 |
| 15 | d-FISH × mod% | 0.713 | 0.021 |
| 16 | d-FISH × C50 | −0.853 | 0.007 |
| 17 | nd-FISH × mod% | 0.738 | 0.015 |
| 18 | nd-FISH × C50 | −0.710 | 0.049 |
| 19 | d-FISH × #mod | 0.558 | 0.094 |
| 20 | d-FISH × stretch | −0.711 | 0.021 |
| 21 | nd-FISH × #mod | 0.547 | 0.102 |
| 22 | nd-FISH × stretch | −0.590 | 0.073 |
| 23 | nd-FISH × TA | 0.505 | 0.165 |
| 24 | nd-FISH × ΔG | −0.583 | 0.129 |
| 25 | d-FISH × TA | 0.274 | 0.476 |
| 26 | d-FISH × ΔG | −0.319 | 0.441 |
| 27 | d-FISH × GC% | −0.099 | 0.785 |
| 28 | nd-FISH × GC% | 0.191 | 0.597 |
| Probe | d-FISH | nd-FISH |
|---|---|---|
| INV1 | 0% | 0% |
| INV2 | ~90% | ~85% |
| INV3 | ~40% | ~30% |
| INV4 | ~90% | ~90% |
| INV5 | 0% | 0% |
| INV6 | ~60% | ~20% |
| INV7 | ~60% | ~25% |
| INV8 | ~60% | ~25% |
| INV9 | ~60% | 0% |
| INV10 | ~90% | ~90% |
| OPT6 | ~90% | ~85% |
| OPT8 | ~90% | ~75% |
| OPT9 | ~75% | ~25% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shepard, C.P.; Emehiser, R.G.; Karmakar, S.; Hrdlicka, P.J. Factors Impacting Invader-Mediated Recognition of Double-Stranded DNA. Molecules 2023, 28, 127. https://doi.org/10.3390/molecules28010127
Shepard CP, Emehiser RG, Karmakar S, Hrdlicka PJ. Factors Impacting Invader-Mediated Recognition of Double-Stranded DNA. Molecules. 2023; 28(1):127. https://doi.org/10.3390/molecules28010127
Chicago/Turabian StyleShepard, Caroline P., Raymond G. Emehiser, Saswata Karmakar, and Patrick J. Hrdlicka. 2023. "Factors Impacting Invader-Mediated Recognition of Double-Stranded DNA" Molecules 28, no. 1: 127. https://doi.org/10.3390/molecules28010127
APA StyleShepard, C. P., Emehiser, R. G., Karmakar, S., & Hrdlicka, P. J. (2023). Factors Impacting Invader-Mediated Recognition of Double-Stranded DNA. Molecules, 28(1), 127. https://doi.org/10.3390/molecules28010127