
High-Resolution Tandem Mass Spectrometry Identifies a Particular Ganglioside Pattern in Early Diabetic Kidney Disease of Type 2 Diabetes Mellitus Patients

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Results and Discussions
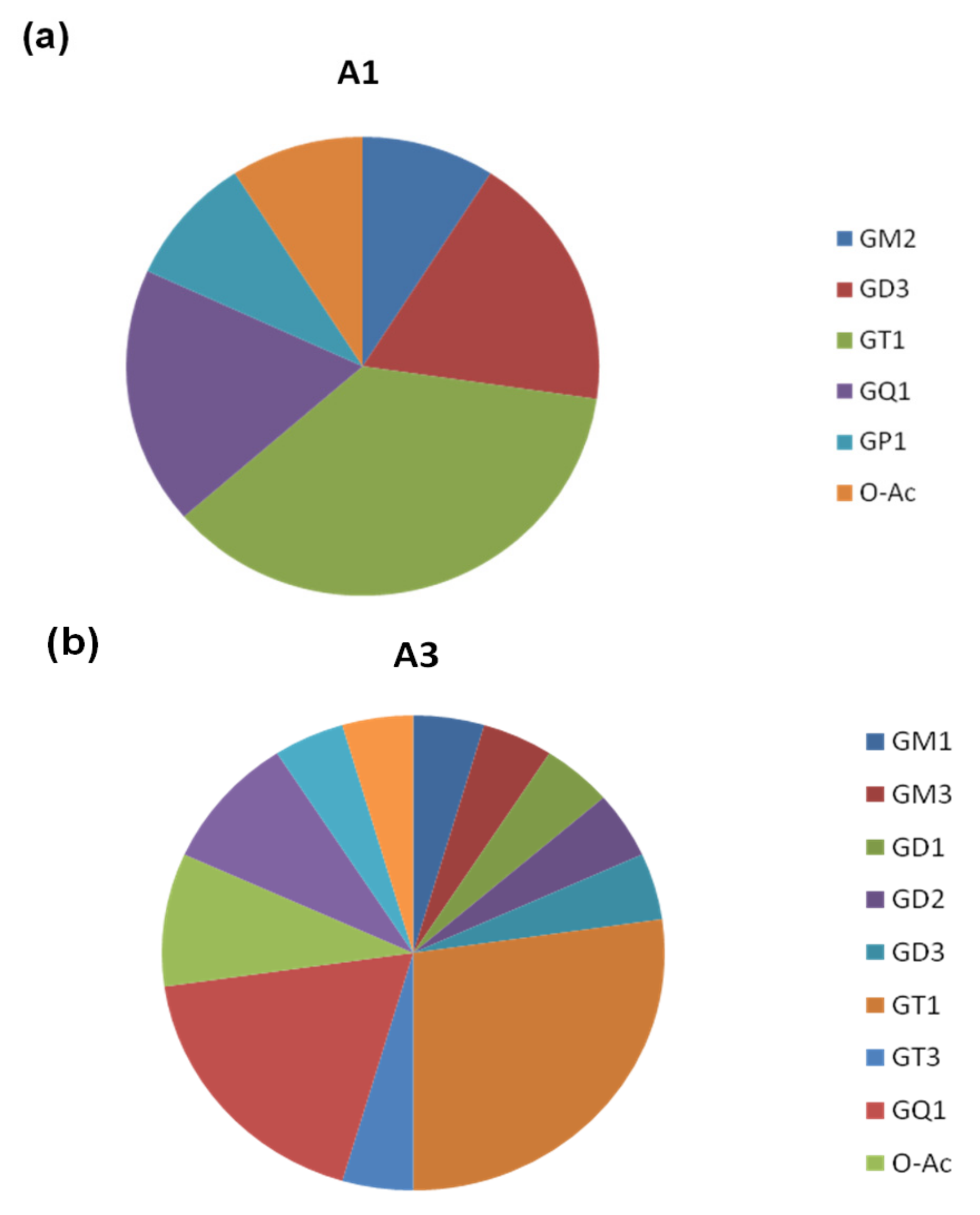
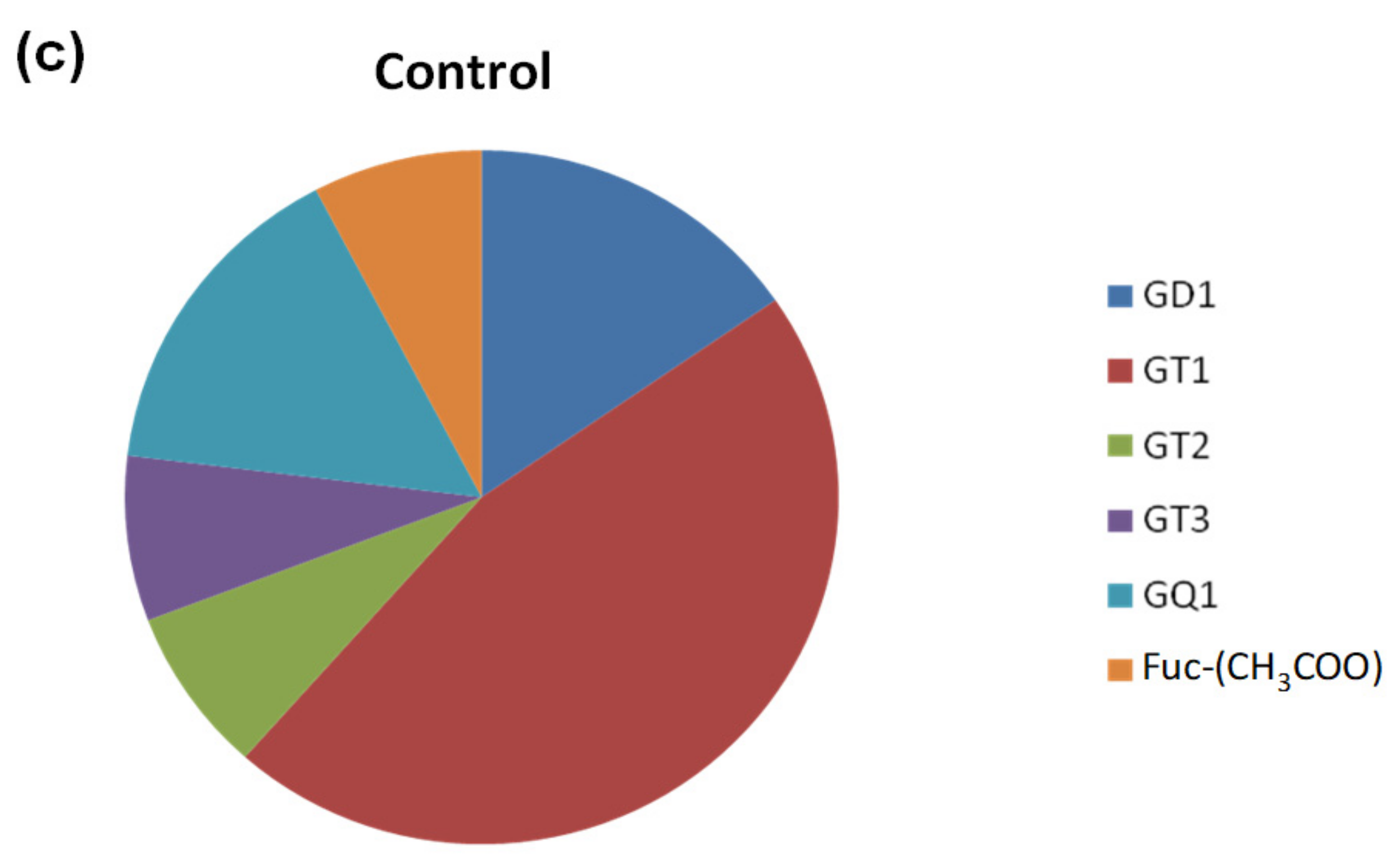
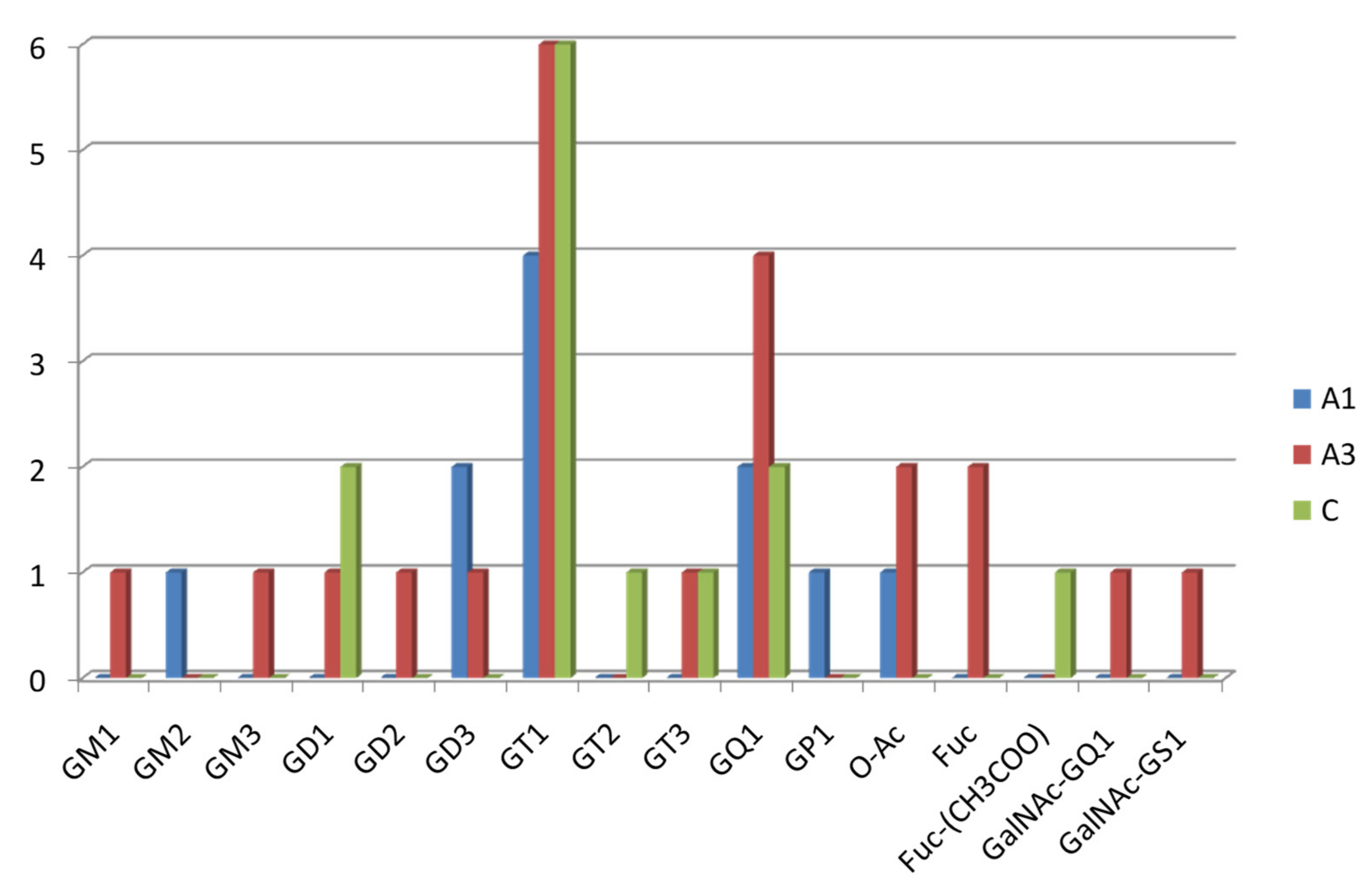
2.1. Comparative Screening of the Samples by nanoESI HR MS
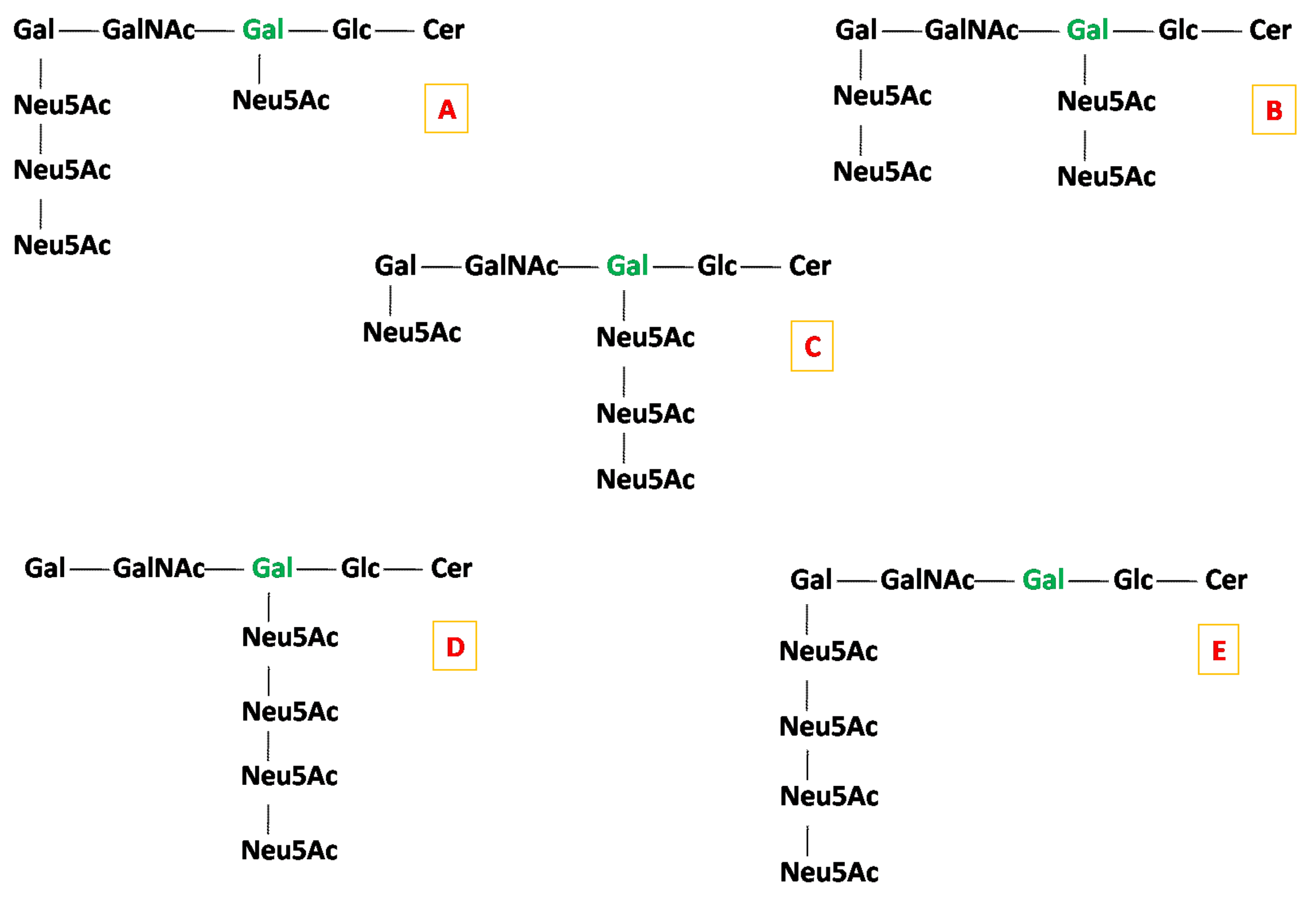
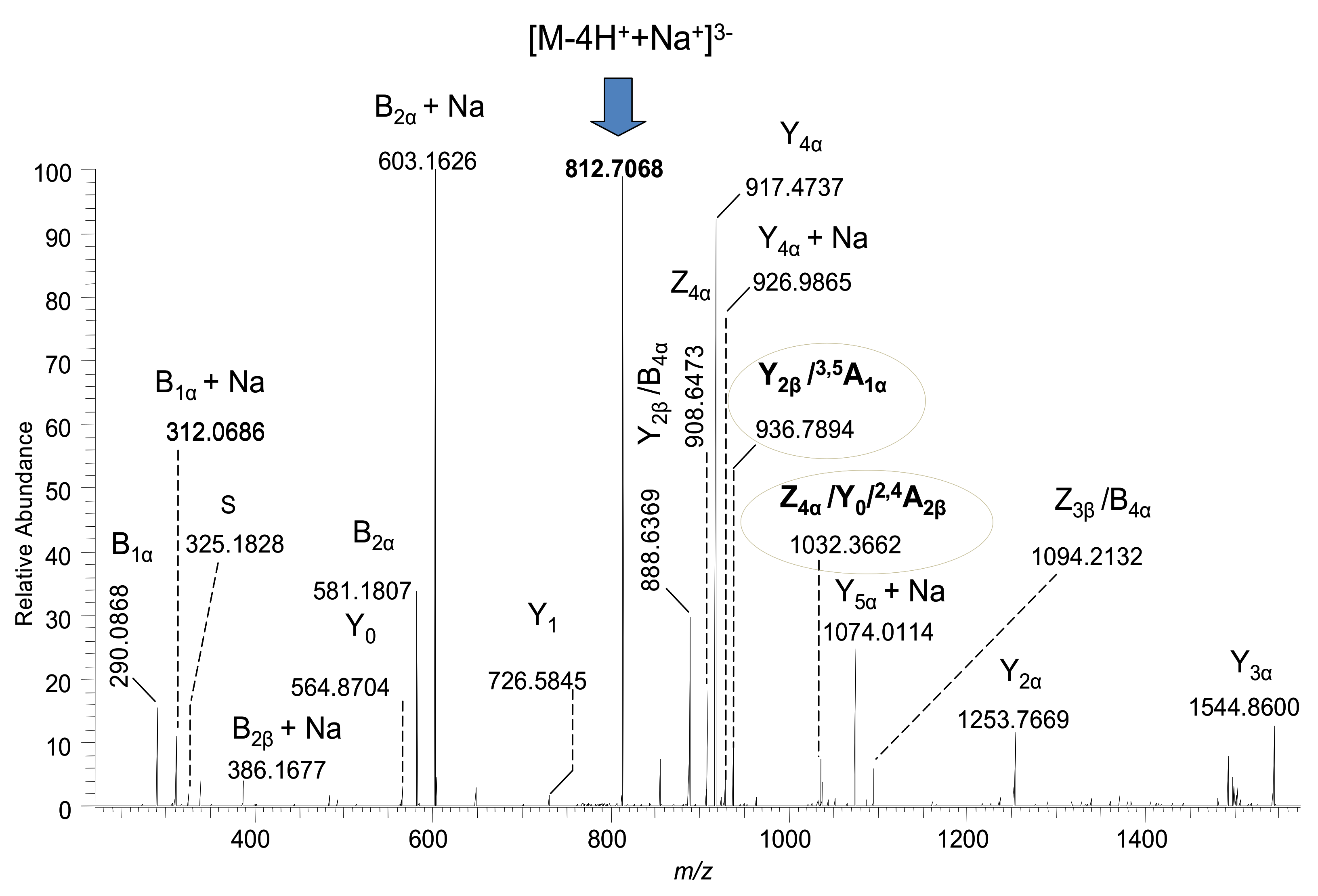
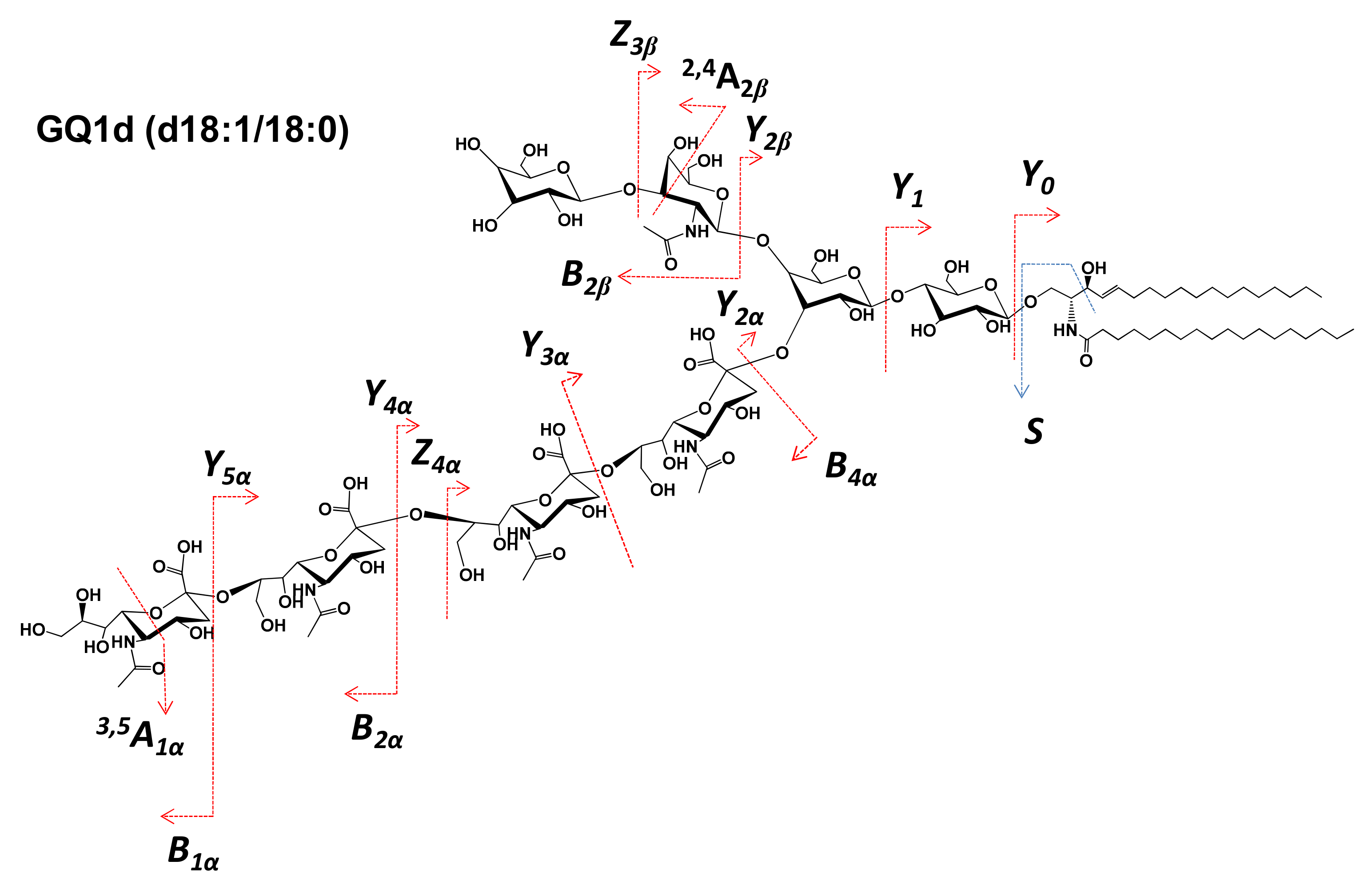
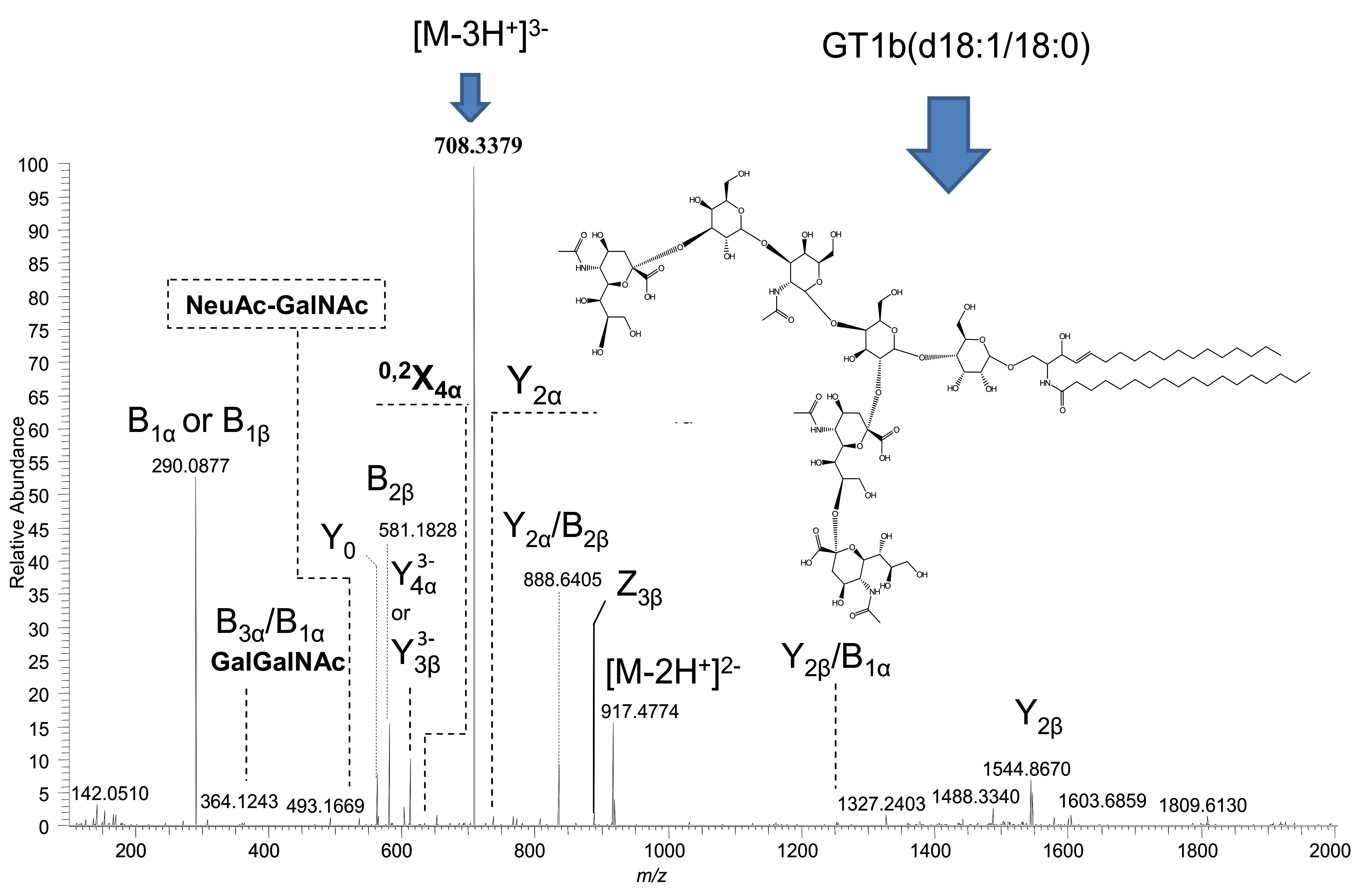
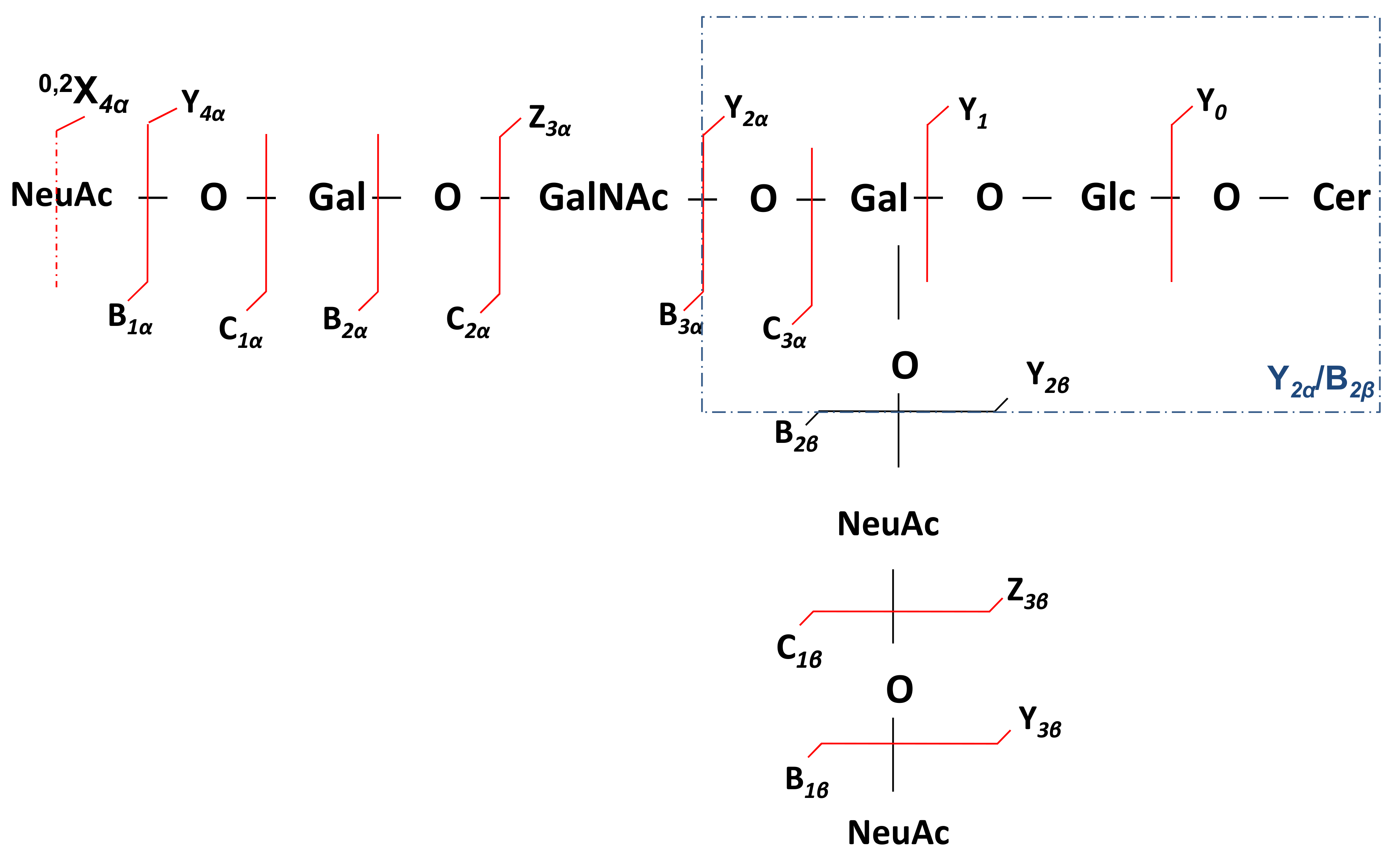
2.2. Detailed Structural Analysis of Polysialylated Species Associated to Macroalbuminuria by HCD MS/MS
3. Materials and Methods
3.1. Subject Enrollment Criteria
3.2. Ethics Statement
3.3. Laboratory Assessments
3.4. Ganglioside Extraction and Purification
3.5. Sample Preparation for Mass Spectrometry
3.6. Orbitrap Mass Spectrometry with nanoESI
3.7. Abbreviation of Gangliosides
3.8. MS Data Interpretation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghaderian, S.B.; Beladi-Mousavi, S.S. The role of diabetes mellitus and hypertension in chronic kidney disease. J. Ren. Inj. Prev. 2014, 3, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Johansen, K.L.; Chertow, G.M.; Foley, R.N.; Gilbertson, D.T.; Herzog, C.A.; Ishani, A.; Israni, A.K.; Ku, E.; Tamura, M.K.; Li, S. US Renal Data System 2020 Annual Data Report: Epidemiology of Kidney Disease in the United States. Am. J. Kidney Dis. 2021, 77, A7–A8. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chang, Y.H.; Yang, S.Y.; Wu, K.D.; Chu, T.S. Update of pathophysiology and management of diabetic kidney disease. J. Med. Assoc. 2018, 117, 662–675. [Google Scholar] [CrossRef] [PubMed]
- Vallon, V.; Thomson, S.C. Renal function in diabetic disease models: The tubular system in the pathophysiology of the diabetic kidney. Annu. Rev. Physiol. 2012, 74, 351–375. [Google Scholar] [CrossRef]
- Milas, O.; Gadalean, F.; Vlad, A.; Dumitrascu, V.; Velciov, S.; Gluhovschi, C.; Bob, F.; Popescu, R.; Ursoniu, S.; Jianu, D.C.; et al. Pro-inflammatory cytokines are associated with podocyte damage and proximal tubular dysfunction in the early stage of diabetic kidney disease in type 2 diabetes mellitus patients. J. Diab. Complicat. 2020, 34, 107479. [Google Scholar] [CrossRef]
- Petrica, L.; Ursoniu, S.; Gadalean, F.; Vlad, A.; Gluhovschi, G.; Dumitrascu, V.; Vlad, D.; Gluhovschi, C.; Velciov, S.; Bob, F.; et al. Urinary podocyte-associated mRNA levels correlate with proximal tubule dysfunction in early diabetic nephropathy of type 2 diabetes mellitus. Diabetol. Metab. Syndr. 2017, 9, 31–43. [Google Scholar] [CrossRef]
- Kwak, D.H.; Rho, Y.I.; Kwon, O.D.; Ahan, S.H.; Song, J.H.; Choo, Y.K.; Kim, S.J.; Choi, B.K.; Jung, K.Y. Decreases of ganglioside GM3 in streptozocin-induced diabetic glomeruli of rats. Life Sci. 2003, 72, 1997–2006. [Google Scholar] [CrossRef]
- Masson, E.; Troncy, L.; Ruggiero, D.; Wiernsperger, N.; Lagarde, M.; El Bawab, S. a-Series Gangliosides Mediate the Effects of Advanced Glycation End Products on Pericyte and Mesangial Cell Proliferation: A Common Mediator for Retinal and Renal Microangiopathy? Diabetes 2005, 54, 220–227. [Google Scholar] [CrossRef][Green Version]
- Zador, Z.; Deshmukh, G.D.; Kunkel, R.; Radin, N.S.; Shayman, J.A. A role for glycosphingolipids accumulation in the renal hypertrophy of streptozocin-induced diabetes mellitus. J. Clin. Investig. 1993, 91, 797–803. [Google Scholar] [CrossRef]
- Ene, C.D.; Penescu, M.; Anghel, A.; Neagu, M.; Budu, V.; Nicolae, I. Monitoring Diabetic Nephropathy by Circulating Gangliosides. J. Immunoass. Immunochem. 2016, 37, 68–79. [Google Scholar] [CrossRef]
- Nilavan, E.; Sundar, S.; Shenbagamoorthy, M.; Narayanan, H.; Nandagopal, B.; Srinivasan, R. Identification of biomarkers for early diagnosis of diabetic nephropathy disease using direct flow through mass spectrometry. Diabetes Metab. Syndr. 2020, 14, 2073–2078. [Google Scholar] [CrossRef]
- Ica, R.; Simulescu, A.; Sarbu, M.; Munteanu, C.V.A.; Vukelić, Ž.; Zamfir, A.D. High resolution mass spectrometry provides novel insights into the ganglioside pattern of brain cavernous hemangioma. Anal. Biochem. 2020, 609, 113976. [Google Scholar] [CrossRef]
- Meyer-Schwesinger, C. The ins-and-outs of podocyte lipid metabolism. Kidney Int. 2020, 98, 1087–1090. [Google Scholar] [CrossRef]
- Grove, K.J.; Voziyan, P.A.; Spraggins, J.M.; Wang, S.; Paueksakon, P.; Harris, R.C.; Hudson, B.G.; Caprioli, R.M. Diabetic nephropathy induces alterations in the glomerular and tubule lipid profiles. J. Lipid. Res. 2014, 55, 1375–1385. [Google Scholar] [CrossRef]
- Stadler, K.; Goldberg, I.J.; Susztak, K. The evolving understanding of the contribution of lipid metabolism to diabetic kidney disease. Curr. Diab. Rep. 2015, 15, 40. [Google Scholar] [CrossRef]
- Kidney Disease: Improving Global Outcomes (KDIGO) CKD Work Group. KDIGO 2012 Clinical Practice Guideline for the Evaluation and Management of Chronic Kidney Disease. Kidney Int. 2013, 3, 1–150. [Google Scholar] [CrossRef]
- Svennerholm, L.; Fredman, P. A procedure for the quantitative isolation of brain gangliosides. Biochim. Biophys. Acta 1980, 617, 97–109. [Google Scholar] [CrossRef]
- Vukelić, Ž.; Metelmann, W.; Müthing, J.; Kos, M.; Peter-Katalinić, K. Anencephaly: Structural characterization of gangliosides in defined brain regions. Biol. Chem. 2001, 382, 259–274. [Google Scholar] [CrossRef]
- Sarbu, M.; Raab, S.; Henderson, L.; Fabris, D.; Vukelić, Ž.; Clemmer, D.E.; Zamfir, A.D. Cerebrospinal fluid: Profiling and fragmentation of gangliosides by ion mobility mass spectrometry. Biochimie 2020, 170, 36–48. [Google Scholar] [CrossRef]
- Svennerholm, L. Ganglioside designation. Adv. Exp. Med. Biol. 1980, 125, 11. [Google Scholar] [CrossRef]
- Chester, M.A. IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature of glycolipids--recommendations 1997. Eur. J. Biochem. 1998, 257, 293–298. [Google Scholar] [CrossRef]
- Sarbu, M.; Petrica, L.; Clemmer, D.E.; Vukelić, Ž.; Zamfir, A.D. Gangliosides of human glioblastoma multiforme: A comprehensive mapping and structural analysis by ion mobility tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2021, 32, 1249–1257. [Google Scholar] [CrossRef]
- Zamfir, A.D. Neurological analyses: Focus on gangliosides and mass spectrometry. Adv. Exp. Med. Biol. 2014, 806, 153–204. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. A systematic nomenclature of carbohydrate fragmentation in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 1988, 5, 397–409. [Google Scholar] [CrossRef]
- Costello, C.E.; Juhasz, P.; Perreault, H. New mass spectral approaches to ganglioside structure determination. Prog. Brain Res. 1994, 101, 45–61. [Google Scholar] [CrossRef]
- Flangea, C.; Serb, A.; Sisu, E.; Zamfir, A.D. Chip-based mass spectrometry of brain gangliosides. Biochim. Biophys. Acta 2011, 1811, 513–535. [Google Scholar] [CrossRef]
- Sarbu, M.; Robu, A.; Ghiulai, R.; Vukelić, Ž.; Clemmer, D.E.; Zamfir, A.D. Electrospray ionization ion mobility mass spectrometry of human brain gangliosides. Anal. Chem. 2016, 88, 5166–5178. [Google Scholar] [CrossRef]
- Robu, A.; Schiopu, C.; Capitan, F.; Zamfir, A.D. Mass spectrometry of gangliosides in extracranial tumors: Application to adrenal neuroblastoma. Anal. Biochem. 2016, 509, 1–11. [Google Scholar] [CrossRef]
- Sarbu, M.; Ica, R.; Petrut, A.; Vukelić, Ž.; Munteanu, C.V.; Petrescu, A.; Zamfir, A.D. Gangliosidome of human anencephaly: A high resolution multistage mass spectrometry study. Biochimie 2019, 163, 142–151. [Google Scholar] [CrossRef]
- Sarbu, M.; Clemmer, D.E.; Zamfir, A.D. Ion mobility mass spectrometry of human melanoma gangliosides. Biochimie 2020, 177, 226–237. [Google Scholar] [CrossRef]
- Ica, R.; Munteanu, C.V.; Vukelić, Ž.; Zamfir, A.D. High-resolution mass spectrometry reveals a complex ganglioside pattern and novel polysialylated structures associated with the human motor cortex. Eur. J. Mass Spectrom. 2021, 27, 205–214. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Proposed Structure | m/ztheor | Molecular Ion | m/zexp A1 | m/zexp A3 | m/zexp C |
|---|---|---|---|---|---|---|
| 1 | GM2(d18:1/22:2) | 477.6110 | [M-3H+]3− | 477.6100 | - | - |
| 2 | GQ1(d18:1/16:0) | 596.7751 | [M-4H+]4− | - | - | 596.7748 |
| 3 | GQ1(d18:1/22:0) | 617.7992 | [M-4H+]4− | - | - | 617.7979 |
| 4 | GM3(d18:0/24:0) | 645.3958 | [M-H2O-4H++2Na+]2− | - | 645.3956 | - |
| 5 | GP1(d18:1/18:0) | 676.5571 | [M-4H+]4− | 676.5569 | - | - |
| 6 | GT1(d18:0/16:0) | 699.0040 | [M-3H+]3− | - | - | 699.0037 |
| 7 | GT1(d18:1/18:0) | 708.3382 | [M-3H+]3− | - | 708.3379 | - |
| 8 | GT1(d18:1/18:1) | 707.6681 | [M-3H+]3− | 707.6684 | - | 707.6684 |
| 9 | GT1(t18:0/18:0) | 714.3514 | [M-3H+]3− | 714.3513 | - | 714.3513 |
| 10 | GT1(d18:1/20:0) | 717.0221 | [M-3H+]3− | - | - | 717.0220 |
| 11 | GT1(d18:0/20:0) | 718.3636 | [M-3H+]3− | - | 718.3637 | - |
| 12 | GT1(t18:0/20:0) | 723.6952 | [M-3H+]3− | 723.6949 | - | - |
| 13 | GD3(d18:1/18:1) | 733.9043 | [M-2H+]2− | 733.9038 | - | - |
| 14 | GD3(d18:1/18:0) | 735.9199 | [M-2H+]2− | 735.9195 | - | - |
| 15 | GT1(t18:1/24:3) | 739.6953 | [M-3H+]3− | - | 739.6950 | - |
| 16 | GD3(d18:1/20:1) | 747.9199 | [M-2H+]2− | - | 747.9195 | - |
| 17 | GM1(d18:1/16:1) | 756.9070 | [M-2H+]2− | - | 756.9078 | - |
| 18 | GQ1(d18:1/18:0) | 812.7072 | [M-4H++Na+]3− | - | 812.7068 | - |
| 19 | GQ1(d18:1/20:0) | 814.7253 | [M-3H+]3− | 814.7248 | - | - |
| 20 | O-Ac GQ1(d18:1/18:0) | 819.3803 | [M-3H+]3− | - | 819.3809 | - |
| 21 | GQ1(d18:1/22:2) | 830.0509 | [M-4H++Na+]3− | - | 830.0504 | - |
| 22 | GD2(d18:1/18:0) | 847.4427 | [M-3H++Na+]2− | - | 847.4421 | - |
| 23 | GT3(d18:1/18:0) | 891.4508 | [M-3H++Na+]2− | - | - | 891.4503 |
| 24 | GD1(d18:1/16:0) | 903.4623 | [M-2H+]2− | 903.4616 | 903.4616 | |
| 25 | Fuc-GT3(t18:1/18:3) | 969.4538 | [M-3H++Na+]2− | - | 969.4534 | - |
| 26 | GT2(d18:0/16:0) | 979.9865 | [M-3H++Na+]2− | - | - | 979.9861 |
| 27 | Fuc-(CH3COO)GD1(d18:0/16:0) | 1007.5125 | [M-2H+]2− | - | - | 1007.5118 |
| 28 | GT1(d18:1/16:0) | 1059.9997 | [M-3H++Na+]2− | - | 1059.9988 | - |
| 29 | GT1(d18:1/18:0) | 1063.0263 | [M-2H+]2− | - | 1063.0260 | - |
| 30 | GT1(d18:1/18:0) | 1085.0084 | [M-4H++2Na+]2− | - | - | 1085.0077 |
| 31 | GT1(d18:1/22:0) | 1091.0573 | [M-2H+]2- | - | - | 1091.0560 |
| 32 | O-Ac GT1(d18:0/24:0) | 1129.0721 | [M-H2O-3H++Na+]2− | 1129.0652 | 1129.0649 | - |
| 33 | GalNAc-GS1(t18:1/18:0) | 1176.8277 | [M-4H++Na+]3− | - | 1176.8266 | - |
| 34 | GQ1(d18:1/18:0) | 1219.5657 | [M-3H++Na+]2− | 1219.5639 | - | - |
| 35 | GQ1(d20:1/18:0) | 1244.5711 | [M-4H++2Na+]2− | - | 1244.5669 | - |
| 36 | Fuc-GT1(d18:0/24:0) | 1179.1103 | [M-2H+]2− | - | 1179.1094 | - |
| 37 | GalNAc-GQ1(d18:1/18:0) | 1310.1289 | [M-2H+]2− | - | 1310.1271 | - |
| Ganglioside Class | A1 | A3 | Control |
|---|---|---|---|
| GM1 | - | x | - |
| GM2 | x | - | - |
| GM3 | - | x | - |
| GD1 | - | x | x |
| GD2 | - | x | - |
| GD3 | x | x | - |
| GT1 | x | x | x |
| GT2 | - | - | x |
| GT3 | - | - | x |
| GQ1 | x | x | x |
| GS1 | - | x | - |
| GP1 | x | - | - |
| O-Ac-modified species | x | x | - |
| Fuc-modified species | - | x | - |
| Fuc-CH3COO-modified species | - | - | x |
| GalNAc-modified species | - | x | - |
| No. | Type of Fragment Ions | m/z |
|---|---|---|
| 1. | Y0 | 564.5353 |
| 2. | Y1 | 726.5879 |
| 3. | C3α | 673.2298 |
| 4. | C2α | 470.1509 |
| 5. | Y2α | 734.5701 |
| 6. | B3α | 657.7908 |
| 7. | B1β or B1α | 290.0877 |
| 8. | Z3β(doubly deprotonated) | 908.4721 |
| 9. | C1β or C1α | 308.2954 |
| 10. | B2β | 581.1828 |
| 11. | Y2α/B2β | 888.6405 |
| 12. | Y2β | 1544.8670 |
| 13. | Z3α | 1674.6733 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suteanu-Simulescu, A.; Zamfir, A.D.; Ica, R.; Sarbu, M.; Munteanu, C.V.A.; Gadalean, F.; Vlad, A.; Bob, F.; Jianu, D.C.; Petrica, L. High-Resolution Tandem Mass Spectrometry Identifies a Particular Ganglioside Pattern in Early Diabetic Kidney Disease of Type 2 Diabetes Mellitus Patients. Molecules 2022, 27, 2679. https://doi.org/10.3390/molecules27092679
Suteanu-Simulescu A, Zamfir AD, Ica R, Sarbu M, Munteanu CVA, Gadalean F, Vlad A, Bob F, Jianu DC, Petrica L. High-Resolution Tandem Mass Spectrometry Identifies a Particular Ganglioside Pattern in Early Diabetic Kidney Disease of Type 2 Diabetes Mellitus Patients. Molecules. 2022; 27(9):2679. https://doi.org/10.3390/molecules27092679
Chicago/Turabian StyleSuteanu-Simulescu, Anca, Alina Diana Zamfir, Raluca Ica, Mirela Sarbu, Cristian V. A. Munteanu, Florica Gadalean, Adrian Vlad, Flaviu Bob, Dragos Catalin Jianu, and Ligia Petrica. 2022. "High-Resolution Tandem Mass Spectrometry Identifies a Particular Ganglioside Pattern in Early Diabetic Kidney Disease of Type 2 Diabetes Mellitus Patients" Molecules 27, no. 9: 2679. https://doi.org/10.3390/molecules27092679
APA StyleSuteanu-Simulescu, A., Zamfir, A. D., Ica, R., Sarbu, M., Munteanu, C. V. A., Gadalean, F., Vlad, A., Bob, F., Jianu, D. C., & Petrica, L. (2022). High-Resolution Tandem Mass Spectrometry Identifies a Particular Ganglioside Pattern in Early Diabetic Kidney Disease of Type 2 Diabetes Mellitus Patients. Molecules, 27(9), 2679. https://doi.org/10.3390/molecules27092679