
Components of Banisteriopsis caapi, a Plant Used in the Preparation of the Psychoactive Ayahuasca, Induce Anti-Inflammatory Effects in Microglial Cells

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Standards
2.2. Plant Extraction and Spectrometric Analysis
2.3. Cell Culture
2.4. Viability Assay
2.5. Apoptosis/Necrosis Assay
2.6. ROS Production
2.7. Cytokine Production
2.8. Statistical Analyses
3. Results and Discussion
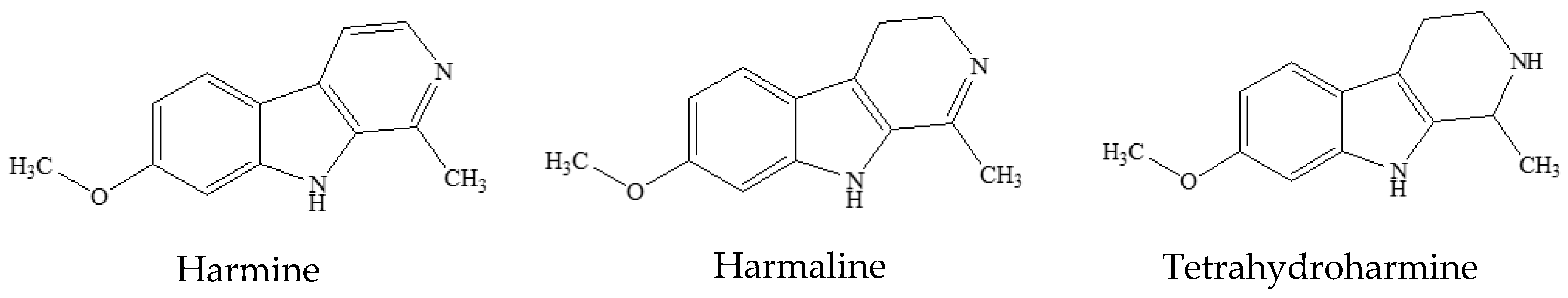
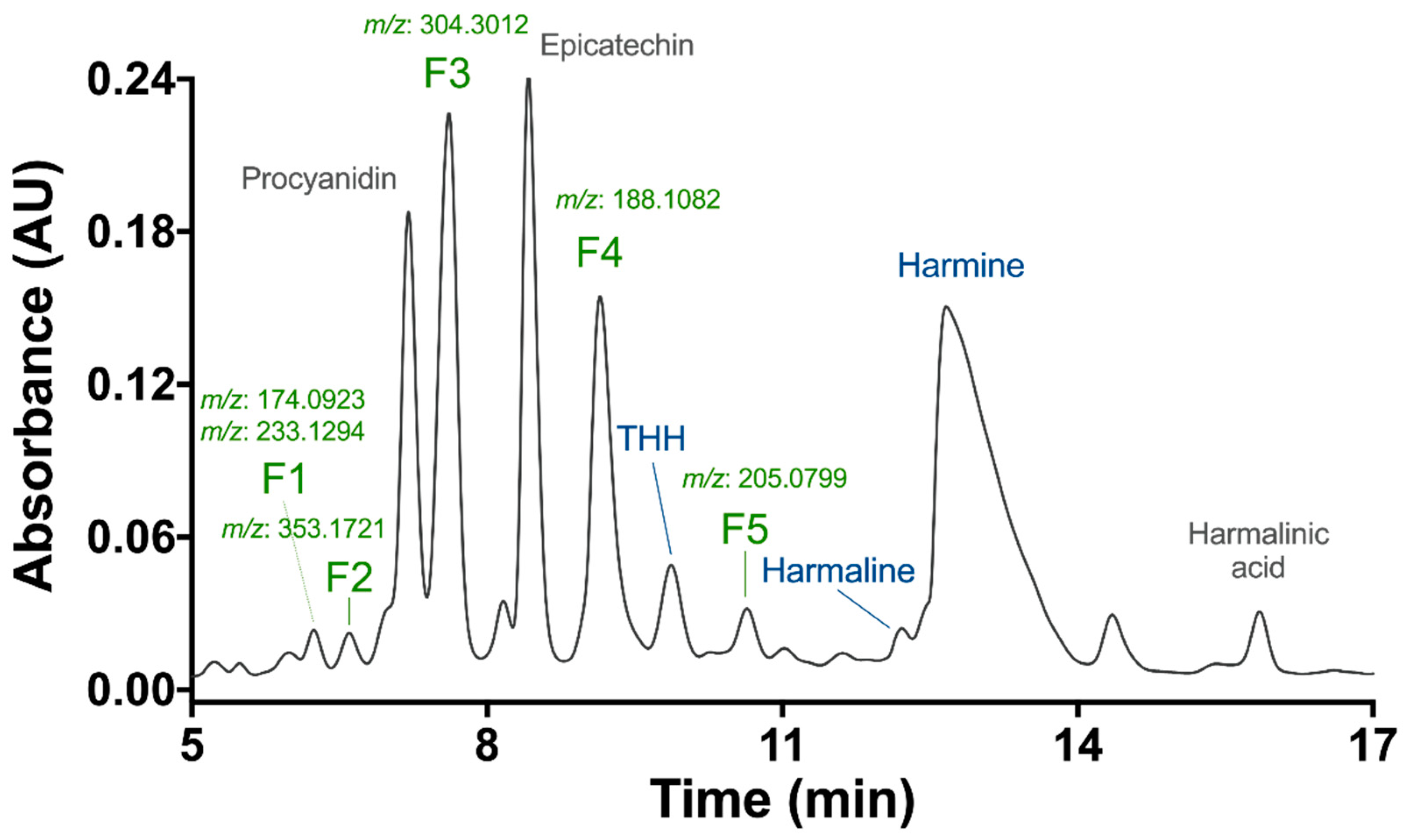
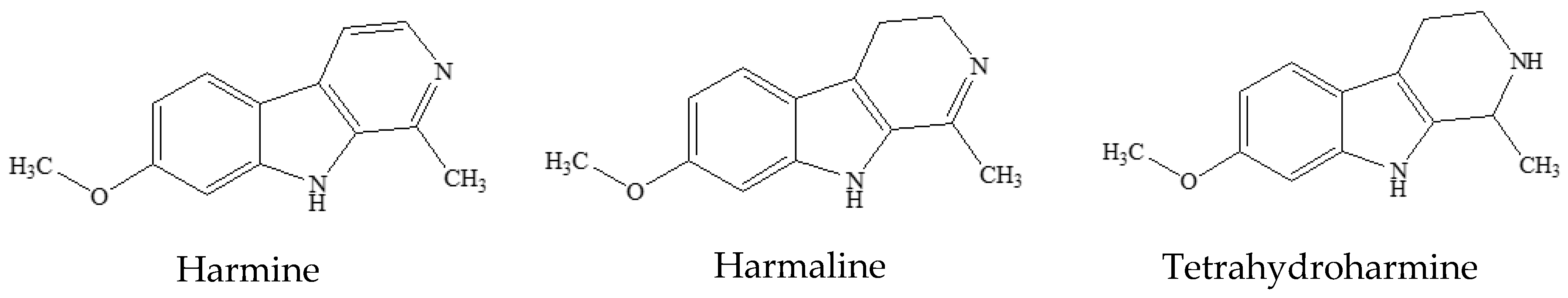
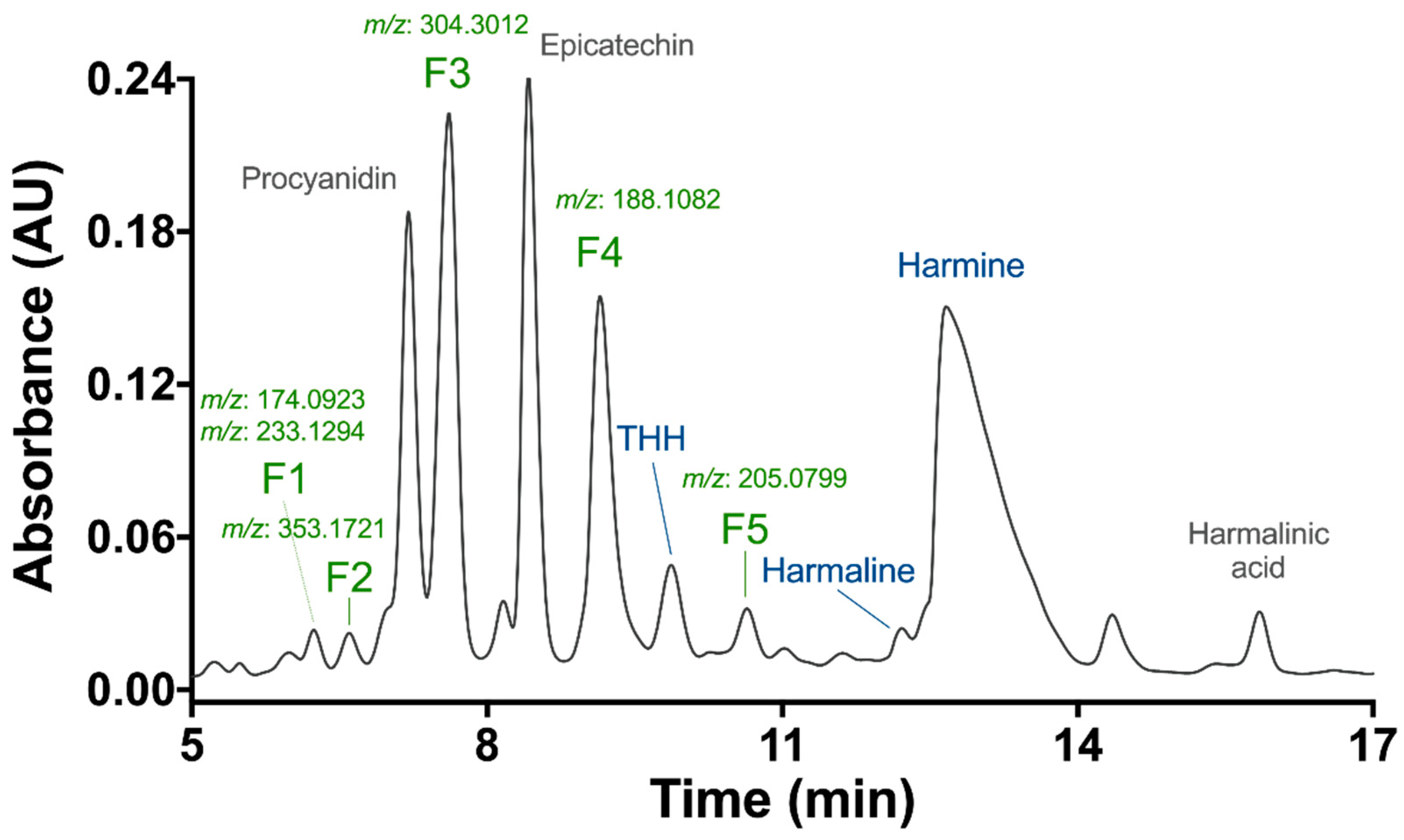
3.1. Preliminary Phytochemical Analyses
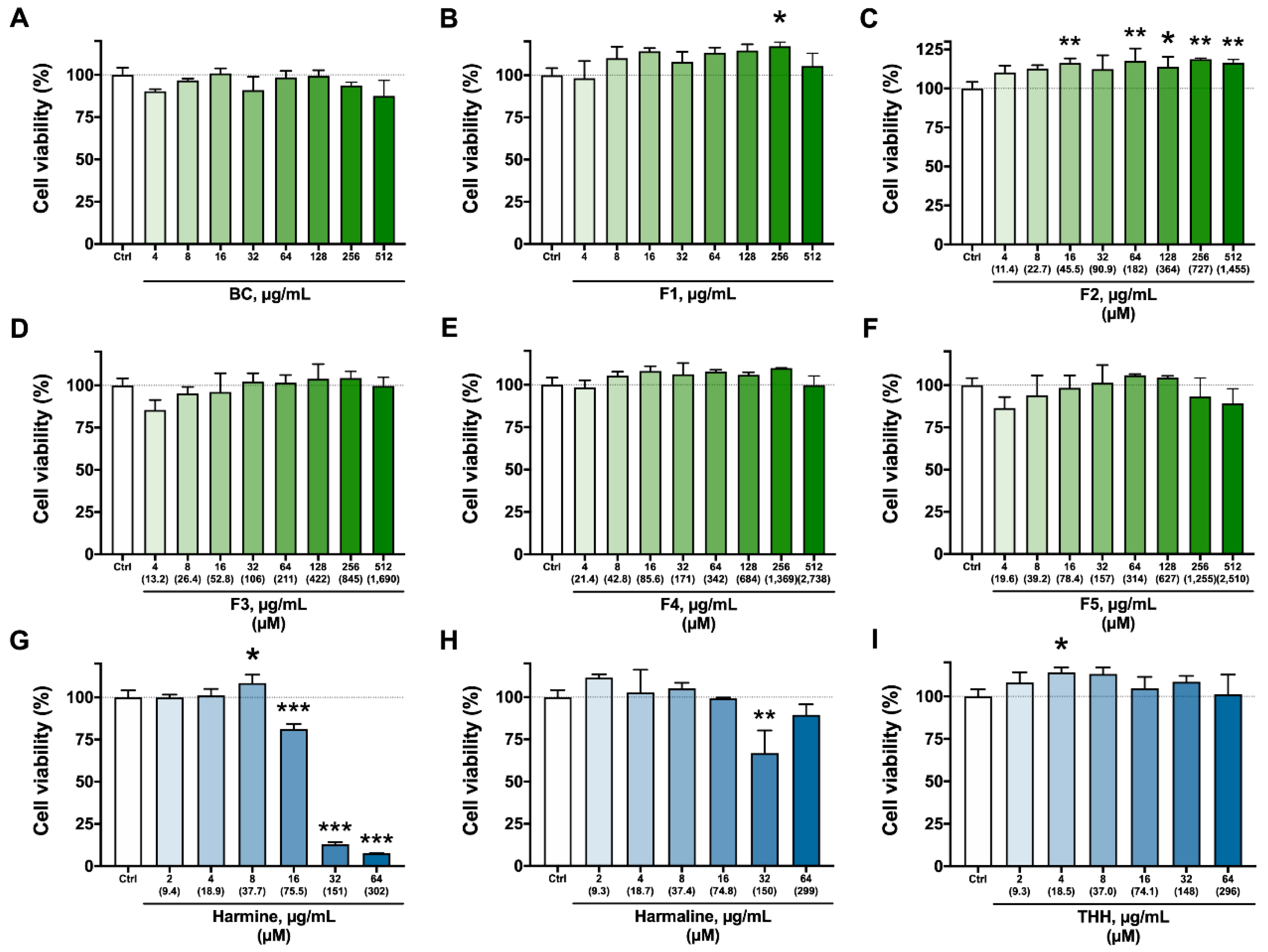
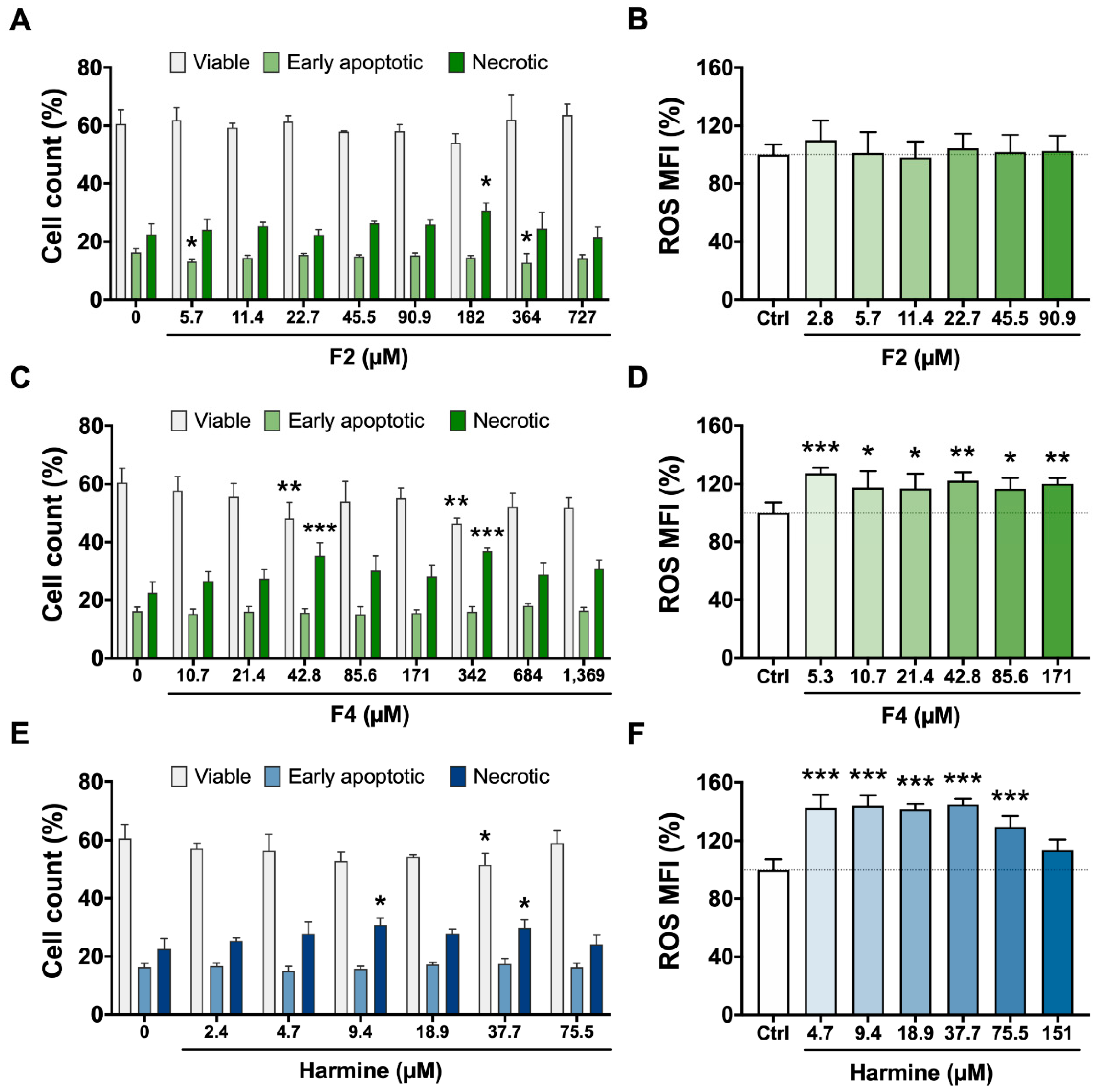
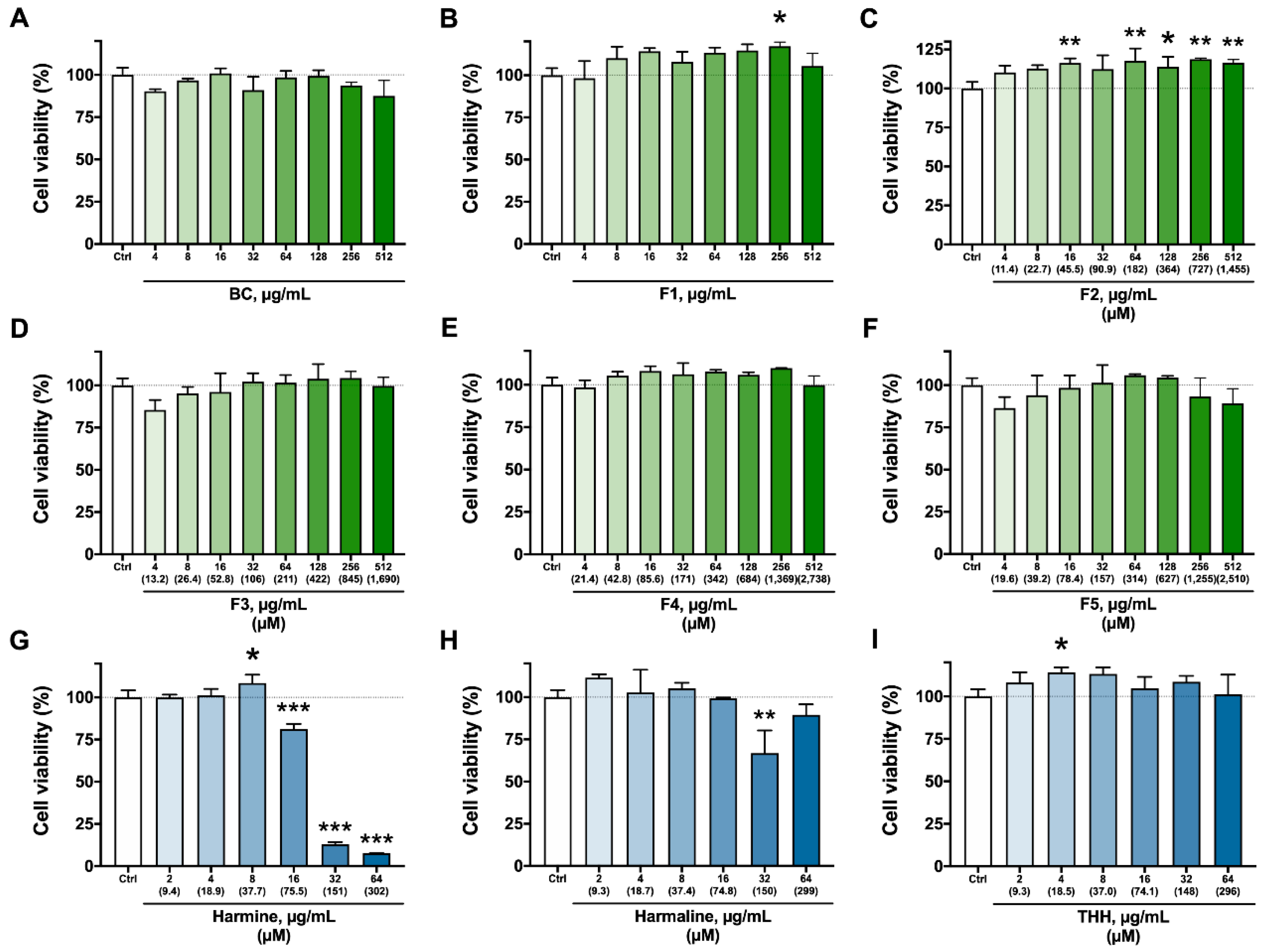
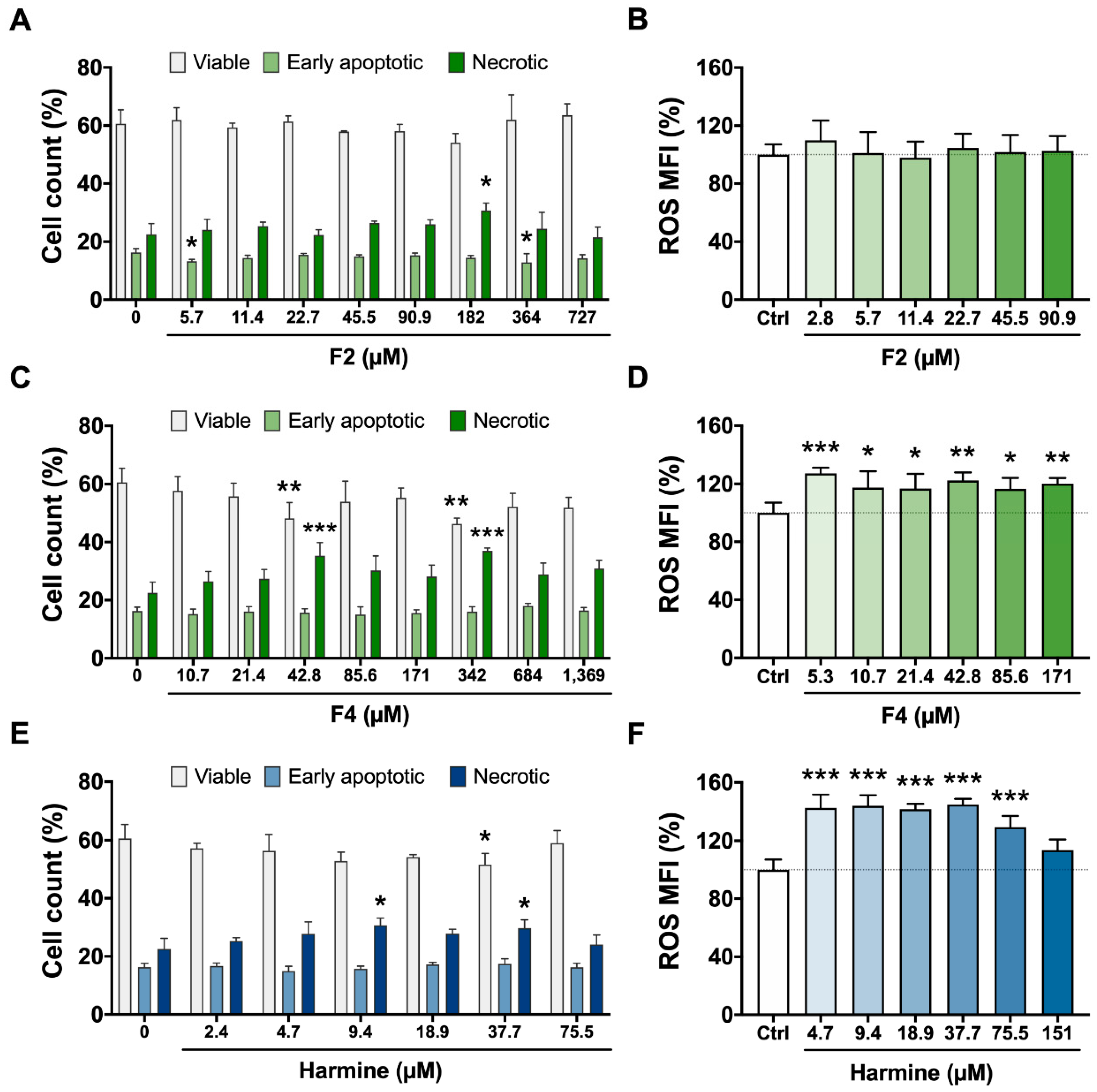
3.2. Cytotoxicity Assays—Viability, Apoptosis/Necrosis, and ROS Production
3.3. Cytokine Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Labate, B.C.; Assis, G.L. A critical review of the literature on the diaspora of Brazilian ayahuasca religions. In The Expanding World Ayahuasca Diaspora, 1st ed.; Labate, B., Cavnar, C., Eds.; Routledge: New York, NY, USA, 2018; pp. 1–21. [Google Scholar]
- Colaço, C.S.; Alves, S.S.; Nolli, L.M.; Pinheiro, W.O.; Oliveira, D.G.R.; Santos, B.W.L.; Pic-Taylor, A.; Mortari, M.R.; Caldas, E.D. Toxicity of ayahuasca after 28 days daily exposure and effects on monoamines and brain-derived neurotrophic factor (BDNF) in brain of Wistar rats. Metab. Brain Dis. 2020, 35, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Inserra, A. Hypothesis: The psychedelic ayahuasca heals traumatic memories via a Sigma 1 receptor-mediated epigenetic-mnemonic process. Front. Pharmacol. 2018, 9, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osório, F.d.L.; Sanches, R.F.; Macedo, L.R.; dos Santos, R.G.; Maia-de-Oliveira, J.P.; Wichert-Ana, L.; de Araujo, D.B.; Riba, J.; Crippa, J.A.; Hallak, J. Antidepressant effects of a single dose of ayahuasca in patients with recurrent depression: A preliminary report. Rev. Bras. Psiquiatr. 2015, 37, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Palhano-Fontes, F.; Barreto, D.; Onias, H.; Andrade, K.C.; Novaes, M.; Pessoa, J.; Mota-Rolim, S.; Osorio, F.L.; Sanches, R.; dos Santos, R. Rapid antidepressant effects of the psychedelic ayahuasca in treatment-resistant depression: A randomised placebo-controlled trial. Psychol. Med. 2017, 49, 655–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pic-Taylor, A.; da Motta, L.G.; de Morais, J.A.; Junior, W.M.; de Fátima Andrade Santos, A.; Campos, L.A.; Mortari, M.R.; von Zuben, M.V.; Caldas, E.D. Behavioural and neurotoxic effects of ayahuasca infusion (Banisteriopsis caapi and Psychotria viridis) in female Wistar rat. Behav. Process. 2015, 118, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.P.; Venable, K.; Destouni, A.; Billac, G.; Ebenezer, P.; Stadler, K.; Nichols, C.; Barker, S.; Francis, J. Pharmahuasca and DMT rescue ROS production and differentially expressed genes observed after predator and psychosocial stress: Relevance to human PTSD. ACS Chem. Neurosci. 2022, 13, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Cesar, P.; Barbosa, R.; Tófoli, L.F.; Bogenschutz, M.P.; Hoy, R.; Berro, L.F.; Marinho, E.A.V.; Areco, K.N.; Winkelman, M.J. Assessment of alcohol and tobacco use disorders among religious users of ayahuasca. Front. Psychiatry 2018, 9, 136. [Google Scholar]
- Fábregas, J.M.; González, D.; Fondevila, S.; Cutchet, M.; Fernández, X.; Barbosa, P.C.R.; Alcázar-Córcoles, M.Á.; Barbanoj, M.J.; Riba, J.; Bouso, J.C. Assessment of addiction severity among ritual users of ayahuasca. Drug Alcohol Depend. 2010, 111, 257–261. [Google Scholar] [CrossRef]
- Nolli, L.M.; de Oliveira, D.; Alves, S.S.; von Zuben, M.V.; Pic-Taylor, A.; Mortari, M.R.; Caldas, E.D. Effects of the hallucinogenic beverage ayahuasca on voluntary ethanol intake by rats and on cFos expression in brain areas relevant to drug addiction. Alcohol 2020, 84, 67–75. [Google Scholar] [CrossRef]
- Oliveira-Lima, A.J.; Santos, R.; Hollais, A.W.; Gerardi-Junior, C.A.; Baldaia, M.A.; Wuo-Silva, R.; Yokoyama, T.S.; Costa, J.L.; Malpezzi-Marinho, E.L.A.; Ribeiro-Barbosa, P.C.; et al. Effects of ayahuasca on the development of ethanol-induced behavioral sensitization and on a post-sensitization treatment in mice. Physiol. Behav. 2015, 142, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.; Lucas, P.; Capler, N.R.; Tupper, K.W.; Martin, G. Ayahuasca-assisted therapy for addiction: Results from a preliminary observational study in Canada. Curr. Drug Abus. Rev. 2013, 6, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Samoylenko, V.; Rahman, M.M.; Tekwani, B.L.; Tripathi, L.M.; Wang, Y.H.; Khan, S.I.; Khan, I.A.; Miller, L.S.; Joshi, V.C.; Muhammad, I. Banisteriopsis caapi, a unique combination of MAO inhibitory and antioxidative constituents for the activities relevant to neurodegenerative disorders and Parkinson’s disease. J. Ethnopharmacol. 2010, 127, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, M.J.; Houghton, P.J.; Rose, S.; Jenner, P.; Lees, A.D. Activities of extract and constituents of Banisteriopsis caapi relevant to parkinsonism. Pharmacol. Biochem. Behav. 2003, 75, 627–633. [Google Scholar] [CrossRef]
- Wang, Y.H.; Samoylenko, V.; Tekwani, B.L.; Khan, I.A.; Miller, L.S.; Chaurasiya, N.D.; Rahman, M.M.; Tripathi, L.M.; Khan, S.I.; Joshi, V.C.; et al. Composition, standardization and chemical profiling of Banisteriopsis caapi, a plant for the treatment of neurodegenerative disorders relevant to Parkinson’s disease. J. Ethnopharmacol. 2010, 128, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Göckler, N.; Jofre, G.; Papadopoulos, C.; Soppa, U.; Tejedor, F.J.; Becker, W. Harmine specifically inhibits protein kinase DYRK1A and interferes with neurite formation. FEBS J. 2009, 276, 6324–6337. [Google Scholar] [CrossRef] [Green Version]
- Frost, D.; Meechoovet, B.; Wang, T.; Gately, S.; Giorgetti, M.; Shcherbakova, I.; Dunckley, T. β-carboline compounds, including harmine, inhibit DYRK1A and Tau phosphorylation at multiple Alzheimer’s disease-related sites. PLoS ONE 2011, 6, e19264. [Google Scholar] [CrossRef] [Green Version]
- Fortunato, J.J.; Réus, G.Z.; Kirsch, T.R.; Stringari, R.B.; Stertz, L.; Kapczinski, F.; Pinto, J.P.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; et al. Acute harmine administration induces antidepressive-like effects and increases BDNF levels in the rat hippocampus. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 1425–1430. [Google Scholar] [CrossRef]
- Fortunato, J.J.; Réus, G.Z.; Kirsch, T.R.; Stringari, R.B.; Fries, G.R.; Kapczinski, F.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Quevedo, J. Chronic administration of harmine elicits antidepressant-like effects and increases BDNF levels in rat hippocampus. J. Neural Transm. 2010, 117, 1131–1137. [Google Scholar] [CrossRef]
- Morales-García, J.A.; De La Fuente Revenga, M.; Alonso-Gil, S.; Rodríguez-Franco, M.I.; Feilding, A.; Perez-Castillo, A.; Riba, J. The alkaloids of Banisteriopsis caapi, the plant source of the Amazonian hallucinogen Ayahuasca, stimulate adult neurogenesis in vitro. Sci. Rep. 2017, 7, 5309. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, Y.; Kawanis, K. New organic bases from Amazonian Banisteriopsis caapi. Phytochemistry 1975, 14, 1633–1635. [Google Scholar] [CrossRef]
- McKenna, D.J.; Towers, G.H.N.; Abbott, F. Monoamine oxidase inhibitors in South American hallucinogenic plants: Tryptamine and ß-carboline constituents of ayahuasca. J. Ethnopharmacol. 1984, 10, 195–223. [Google Scholar] [CrossRef]
- Galloway, D.A.; Phillips, A.E.M.; Owen, D.R.J.; Moore, C.S. Phagocytosis in the brain: Homeostasis and disease. Front. Immunol. 2019, 10, 790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolivel, V.; Brun, S.; Binamé, F.; Benyounes, J.; Taleb, O.; Bagnard, D.; De Sèze, J.; Patte-Mensah, C.; Mensah-Nyagan, A.G. Microglial cell morphology and phagocytic activity are critically regulated by the neurosteroid allopregnanolone: A possible role in neuroprotection. Cells 2021, 10, 698. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 polarization and metabolic states. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef]
- Block, M.L.; Hong, J.S. Microglia and inflammation-mediated neurodegeneration: Multiple triggers with a common mechanism. Prog. Neurobiol. 2005, 76, 77–98. [Google Scholar] [CrossRef]
- Carson, M.J.; Doose, J.M.; Melchior, B.; Schmid, C.D.; Ploix, C.C. CNS immune privilege: Hiding in plain sight. Immunol. Rev. 2006, 213, 48–65. [Google Scholar] [CrossRef]
- Rahimian, R.; Wakid, M.; O’Leary, L.A.; Mechawar, N. The emerging tale of microglia in psychiatric disorders. Neurosci. Biobehav. Rev. 2021, 131, 1–29. [Google Scholar] [CrossRef]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 2012, 87, 10–20. [Google Scholar] [CrossRef]
- Van Eldik, L.J.; Thompson, W.L.; Ranaivo, H.R.; Behanna, H.A.; Martin Watterson, D. Glia proinflammatory cytokine upregulation as a therapeutic target for neurodegenerative diseases: Function-based and target-based discovery approaches. Int. Rev. Neurobiol. 2007, 82, 277–296. [Google Scholar]
- He, Y.; Gao, Y.; Zhang, Q.; Zhou, G.; Cao, F.; Yao, S. IL-4 Switches Microglia/macrophage M1/M2 Polarization and Alleviates Neurological Damage by Modulating the JAK1/STAT6 Pathway Following ICH. Neuroscience 2020, 437, 161–171. [Google Scholar] [CrossRef]
- Santos, B.W.L.; Oliveira, R.C.; Sonsin-Oliveira, J.; Fagg, C.W.; Barbosa, J.B.F.; Caldas, E.D. Biodiversity of β-Carboline Profile of Banisteriopsis caapi and ayahuasca, a plant and a brew with neuropharmacological potential. Plants 2020, 9, 870. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Suzuki, T.; Ishii, H.; Ogata, A.; Nakae, D. Mitochondrial dysfunction and biotransformation of β-carboline alkaloids, harmine and harmaline, on isolated rat hepatocytes. Chem. Biol. Interact. 2010, 188, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Simão, A.Y.; Gonçalves, J.; Gradillas, A.; García, A.; Restolho, J.; Fernández, N.; Rodilla, J.M.; Barroso, M.; Duarte, A.P.; Cristóvão, A.C. Evaluation of the cytotoxicity of ayahuasca beverages. Molecules 2020, 25, 5594. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Wang, Z.; Ma, K.; Chen, Q. Harmine Induces Gastric Cancer Cell Apoptosis through the ROS mediated PI3K/AKT Signaling Pathway. Curr. Signal Transduct. Ther. 2015, 10, 112–118. [Google Scholar] [CrossRef]
- Zhang, X.F.; Sun, R.Q.; Jia, Y.F.; Chen, Q.; Tu, R.F.; Li, K.K.; Zhang, X.D.; Du, R.L.; Cao, R.H. Synthesis and mechanisms of action of novel harmine derivatives as potential antitumor agents. Sci. Rep. 2016, 6, 33204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, C.; Jiang, C.; Zeng, H.; He, X. Novel mechanism of harmaline on inducing G2/M cell cycle arrest and apoptosis by up-regulating Fas/FasL in SGC-7901 cells. Sci. Rep. 2015, 5, 18613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Li, M.; Yu, Y.; He, J.; Hu, S.; Pan, M.; Lu, S.; Liao, K.; Pan, Z.; Zhou, Y. Effects of harmaline on cell growth of human liver cancer through the p53/p21 and Fas/FasL signaling pathways. Oncol. Lett. 2018, 15, 1931–1936. [Google Scholar] [CrossRef] [Green Version]
- Hancock, J.T.; Desikan, R.; Neill, S.J. Role of reactive oxygen species in cell signalling pathways. Biochem. Soc. Trans. 2001, 29, 345–350. [Google Scholar] [CrossRef]
- Figueroa, D.; Asaduzzaman, M.; Young, F. Real time monitoring and quantification of reactive oxygen species in breast cancer cell line MCF-7 by 2′,7′–dichlorofluorescin diacetate (DCFDA) assay. J. Pharmacol. Toxicol. Methods 2018, 94, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, M.F.; Oxenius, A. Interleukin 2: From immunostimulation to immunoregulation and back again. EMBO Rep. 2007, 8, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Pol, J.G.; Caudana, P.; Paillet, J.; Piaggio, E.; Kroemer, G. Effects of interleukin-2 in immunostimulation and immunosuppression. J. Exp. Med. 2020, 217, e20191247. [Google Scholar] [CrossRef] [Green Version]
- Cobourne-Duval, M.K.; Taka, E.; Mendonca, P.; Soliman, K. Thymoquinone increases the expression of neuroprotective proteins while decreasing the expression of pro-inflammatory cytokines and the gene expression NFκB pathway signaling targets in LPS/IFNγ-activated BV-2 microglia cells. J. Neuroimmunol. 2018, 320, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zhu, Y.; He, G.; Ni, H.; Liu, C.; Ma, L.; Zhang, L.; Shi, D. Dexmedetomidine attenuates neuroinflammation in LPS-stimulated BV2 microglia cells through upregulation of miR-340. Drug Des. Devel. Ther. 2019, 13, 3465–3475. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, V.; Cornelius, C.; Dinkova-Kostova, A.T.; Calabrese, E.J.; Mattson, M. P Cellular stress responses, the hormesis paradigm, and vitagenes: Novel targets for therapeutic intervention in neurodegenerative disorders. Antioxid. Redox Signal. 2010, 13, 1763–1811. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, M.; Tan, S.; Wang, C.; Fan, S.; Huang, C. Harmine is an inflammatory inhibitor through the suppression of NF-κB signaling. Biochem. Biophys. Res. Commun. 2017, 489, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, e17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, X.; Yao, Q.; Li, W.; Zang, L.; Li, W.; Zhao, J.; Liu, F.; Zhi, W. Harmine mitigates LPS-induced acute kidney injury through inhibition of the TLR4-NF-κB/NLRP3 inflammasome signalling pathway in mice. Eur. J. Pharmacol. 2019, 849, 160–169. [Google Scholar] [CrossRef]
- Galvão-Coelho, N.L.; de Menezes Galvão, A.C.; de Almeida, R.N.; Palhano-Fontes, F.; Campos Braga, I.; Lobão Soares, B.; Maia-de-Oliveira, J.P.; Perkins, D.; Sarris, J.; de Araujo, D.B. Changes in inflammatory biomarkers are related to the antidepressant effects of Ayahuasca. J. Psychopharmacol. 2020, 34, 1125–1133. [Google Scholar] [CrossRef]
- Gonçalves, J.; Luís, Â.; Gradillas, A.; García, A.; Restolho, J.; Fernández, N.; Domingues, F.; Gallardo, E.; Duarte, A.P. Ayahuasca beverages: Phytochemical analysis and biological properties. Antibiotics 2020, 9, 731. [Google Scholar] [CrossRef]
- da Silva, M.G.; Daros, G.C.; de Bitencourt, R.M. Anti-inflammatory activity of ayahuasca: Therapeutical implications in neurological and psychiatric diseases. Behav. Brain Res. 2021, 400, 113003. [Google Scholar] [CrossRef]
- Brundin, L.; Bryleva, E.Y.; Thirtamara Rajamani, K. Role of Inflammation in Suicide: From Mechanisms to Treatment. Neuropsychopharmacology 2017, 42, 271–283. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction µM | IL-2 (pg/mL) | IL-4 (pg/mL) | IL-6 (pg/mL) | IL-10 (pg/mL) | IL-17A (pg/mL) | IFN-γ (pg/mL) | TNF (pg/mL) |
|---|---|---|---|---|---|---|---|
| Control (n = 6) | 3.84 ± 0.96 | 1.49 ± 0.51 | 1.43 ± 0.22 | 1.77 ± 0.44 | 0.54 ± 0.1 | 0.88 ± 0.08 | 7.59 ± 0.81 |
| F3 (n = 3, except for 1.7 and 211 µM, n = 6) | |||||||
| 1.7 | 1.16 ± 0.65 | 2.57 ± 1.19 | 0.86 ± 0.86 | 2.08 ± 1.08 | 0.19 ± 0.19 | 0.71 ± 0.11 | 5.57 ± 0.82 |
| 6.6 | 0.00 ± 0.00 | 3.86 ± 1.45 | 0.93 ± 0.43 | 1.24 ± 0.71 | 0.42 ± 0.42 | 0.75 ± 0.15 | 5.43 ± 1.4 |
| 13.2 | 1.46 ± 1.46 | 4.61 ± 0.39 | 1.72 ± 0.23 | 2.09 ± 1.53 | 0.49 ± 0.49 | 0.58 ± 0.15 | 5.22 ± 0.21 |
| 26.4 | 5.01 ± 1.67 | 3.81 ± 0.53 | 0.81 ± 0.17 | 3.06 ± 1.06 | 0.00 ± 0.00 | 0.27 ± 0.14 | 6.56 ± 0.82 |
| 52.8 | 1.37 ± 1.37 | 2.93 ± 1.54 | 1.06 ± 0.15 | 1.70 ± 1.27 | 0.29 ± 0.29 | 0.54 ± 0.13 | 6.85 ± 0.44 |
| 106 | 0.00 ± 0.00 | 1.95 ± 1.26 | 0.58 ± 0.29 | 0.39 ± 0.39 | 0.18 ± 0.18 | 0.41 ± 0.17 | 5.53 ± 2.16 |
| 211 | 0.00 ± 0.00 * | 1.32 ± 0.87 | 0.39 ± 0.12 * | 0.19 ± 0.19 | 0.01 ± 0.01 | 0.43 ± 0.13 | 5.34 ± 0.84 |
| F4 (n = 3, except for 1.7 µM, n = 6) | |||||||
| 2.7 | 2.93 ± 1.11 | 2.15 ± 1.1 | 0.88 ± 0.08 | 1.61 ± 0.57 | 0.54 ± 0.16 | 0.55 ± 0.09 | 6.83 ± 0.73 |
| 10.7 | 0.00 ± 0.00 | 2.66 ± 1.5 | 0.60 ± 0.3 * | 1.44 ± 0.81 | 0.00 ± 0.00 | 0.28 ± 0.25 * | 4.24 ± |
| 21.4 | 0.00 ± 0.00 | 0.60 ± 0.60 | 0.00 ± 0.00 *** | 0.43 ± 0.43 | 0.00 ± 0.00 | 0.08 ± 0.08 ** | 4.42 ± 0.84 |
| 42.8 | 0.00 ± 0.00 | 1.04 ± 1.04 | 0.00 ± 0.00 *** | 0.00 ± 0.00 | 0.16 ± 0.16 | 0.03 ± 0.03 *** | 2.01 ± 1.08 ** |
| 85.6 | 0.00 ± 0.00 | 2.70 ± 1.64 | 0.25 ± 0.25 ** | 0.00 ± 0.00 | 0.32 ± 0.32 | 0.15 ± 0.15 ** | 2.68 ± 1.36 * |
| 171 | 0.00 ± 0.00 | 3.96 ± 0.91 | 0.89 ± 0.11 | 1.16 ± 0.35 | 0.12 ± 0.12 | 0.65 ± 0.07 | 6.80 ± 1.01 |
| 342 | 0.30 ± 0.30 | 3.03 ± 0.11 | 0.23 ± 0.11 ** | 0.14 ± 0.14 * | 0.00 ± 0.00 ± | 0.61 ± 0.12 | 4.04 ± 0.22 |
| F5 (n = 3, except for 2.5 µM, n = 5) | |||||||
| 2.5 | 2.43 ± 1.02 | 1.93 ± 0.97 | 0.54 ± 0.15 | 1.27 ± 1.21 | 0.09 ± 0.09 ** | 0.57 ± 0.09 | 5.67 ± 0.48 |
| 9.8 | 0.95 ± 0.95 | 3.44 ± 1.08 | 0.88 ± 0.61 | 1.32 ± 0.42 | 0.00 ± 0.00 ** | 0.67 ± 0.14 | 8.47 ± 2.51 |
| 19.6 | 0.00 ± 0.00 * | 2.28 ± 1.32 | 0.49 ± 0.12 | 0.46 ± 0.46 | 0.17 ± 0.09 | 0.52 ± 0.19 | 3.99 ± 0.21 |
| 39.2 | 0.00 ± 0.00 * | 1.95 ± 0.2 | 0.09 ± 0.09 ** | 0.00 ± 0.00 | 0.00 ± 0.00** | 0.16 ± 0.10 ** | 1.81 ± 0.14 ** |
| 78.4 | 0.00 ± 0.00 * | 0.68 ± 0.68 | 0.00 ± 0.00 ** | 0.00 ± 0.00 | 0.00 ± 0.00 ** | 0.16 ± 0.16 ** | 1.80 ± 1.13 ** |
| 157 | 0.00 ± 0.00 * | 0.00 ± 0.00 | 0.00 ± 0.00 ** | 0.00 ± 0.00 | 0.00 ± 0.00 ** | 0.00 ± 0.00 *** | 0.35 ± 0.35 *** |
| 314 | 0.00 ± 0.00 * | 0.00 ± 0.00 | 0.00 ± 0.00 ** | 0.00 ± 0.00 | 0.00 ± 0.00 ** | 0.00 ± 0.00 *** | 0.01 ± 0.01 *** |
| Harmine (n = 6, except for 18.9 µM, n = 5) | |||||||
| 2.4 | 1.23 ± 0.85 | 2.69 ± 0.64 | 0.81 ± 0.16 | 0.84 ± 0.34 | 0.33 ± 0.13 | 0.56 ± 0.04 | 5.77 ± 0.61 |
| 9.4 | 1.84 ± 0.87 | 1.91 ± 0.90 | 0.86 ± 0.31 | 1.18 ± 0.62 | 0.17 ± 0.09 | 0.70 ± 0.17 | 5.15 ± 0.95 |
| 18.9 | 0.29 ± 0.29 * | 2.35 ± 0.97 | 0.72 ± 0.21 | 0.44 ± 0.23 | 0.11 ± 0.11 | 0.55 ± 0.10 | 4.25 ± 0.51 * |
| 37.7 | 2.33 ± 0.76 | 2.51 ± 1.35 | 1.39 ± 0.27 | 1.66 ± 0.51 | 0.51 ± 0.30 | 0.87 ± 0.14 | 4.04 ± 0.61 * |
| 75.5 | 0.20 ± 0.2 * | 1.38 ± 0.76 | 0.63 ± 0.17 | 1.25 ± 0.75 | 0.30 ± 0.13 | 0.78 ± 0.09 | 3.77 ± 0.61 ** |
| Harmaline (n = 3, except for 18.7 µM, n = 6) | |||||||
| 2.4 | 0.00 ± 0.00 ** | 1.32 ± 1.32 | 0.18 ± 0.18 | 0.59 ± 0.59 | 0.26 ± 0.26 | 0.31 ± 0.31 | 3.12 ± 3.12 |
| 9.3 | 0.00 ± 0.00 ** | 1.57 ± 1.23 | 0.51 ± 0.43 | 0.73 ± 0.73 | 0.14 ± 0.14 | 0.33 ± 0.33 | 2.91 ± 1.89 |
| 18.7 | 0.00 ± 0.00 ** | 0.00 ± 0.00 | 0.48 ± 0.31 | 0.12 ± 0.12 * | 0.02 ± 0.02 * | 0.20 ± 0.12 * | 1.29 ± 0.74 ** |
| 37.4 | 0.00 ± 0.00** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 * | 0.01 ± 0.01 * | 0.15 ± 0.15 ** |
| 74.8 | 0.00 ± 0.00** | 0.00± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 * | 0.00 ± 0.00 ** | 0.00 ± 0.00 ** |
| THH (n = 3) | |||||||
| 2.3 | 4.25 ± 2.25 | 6.14 ± 0.47 ** | 1.52 ± 0.37 | 3.16 ± 1.10 | 0.80 ± 0.41 | 0.77 ± 0.06 | 6.68 ± 0.38 |
| 9.3 | 0.00 ± 0.00 | 4.32 ± 1.43 | 0.00 ± 0.00 ** | 1.01 ± 1.01 | 0.39 ± 0.39 | 0.31 ± 0.17 ** | 2.58 ± 1.12 ** |
| 18.5 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 ** | 0.17 ± 0.17 | 0.00 ± 0.00 | 0.08 ± 0.08 *** | 0.85 ± 0.85 *** |
| 37.0 | 0.00 ± 0.00 | 1.45 ± 1.45 | 0.00 ± 0.00 ** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.01 *** | 0.72 ± 0.68 *** |
| 74.1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 ** | 0.00± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 *** | 0.78 ± 0.78 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, B.W.L.; Moreira, D.C.; Borges, T.K.d.S.; Caldas, E.D. Components of Banisteriopsis caapi, a Plant Used in the Preparation of the Psychoactive Ayahuasca, Induce Anti-Inflammatory Effects in Microglial Cells. Molecules 2022, 27, 2500. https://doi.org/10.3390/molecules27082500
Santos BWL, Moreira DC, Borges TKdS, Caldas ED. Components of Banisteriopsis caapi, a Plant Used in the Preparation of the Psychoactive Ayahuasca, Induce Anti-Inflammatory Effects in Microglial Cells. Molecules. 2022; 27(8):2500. https://doi.org/10.3390/molecules27082500
Chicago/Turabian StyleSantos, Beatriz Werneck Lopes, Daniel Carneiro Moreira, Tatiana Karla dos Santos Borges, and Eloisa Dutra Caldas. 2022. "Components of Banisteriopsis caapi, a Plant Used in the Preparation of the Psychoactive Ayahuasca, Induce Anti-Inflammatory Effects in Microglial Cells" Molecules 27, no. 8: 2500. https://doi.org/10.3390/molecules27082500
APA StyleSantos, B. W. L., Moreira, D. C., Borges, T. K. d. S., & Caldas, E. D. (2022). Components of Banisteriopsis caapi, a Plant Used in the Preparation of the Psychoactive Ayahuasca, Induce Anti-Inflammatory Effects in Microglial Cells. Molecules, 27(8), 2500. https://doi.org/10.3390/molecules27082500