
The Phenolics and Antioxidant Properties of Black and Purple versus White Eggplant Cultivars

 ,
, 
 ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. Variation in Total Phenolic Compounds (TPC), Flavonoid (TF) Contents and Antioxidant Capacity (AC) Values in Eggplants
2.2. Variation in Phenolic Acids in Eggplants
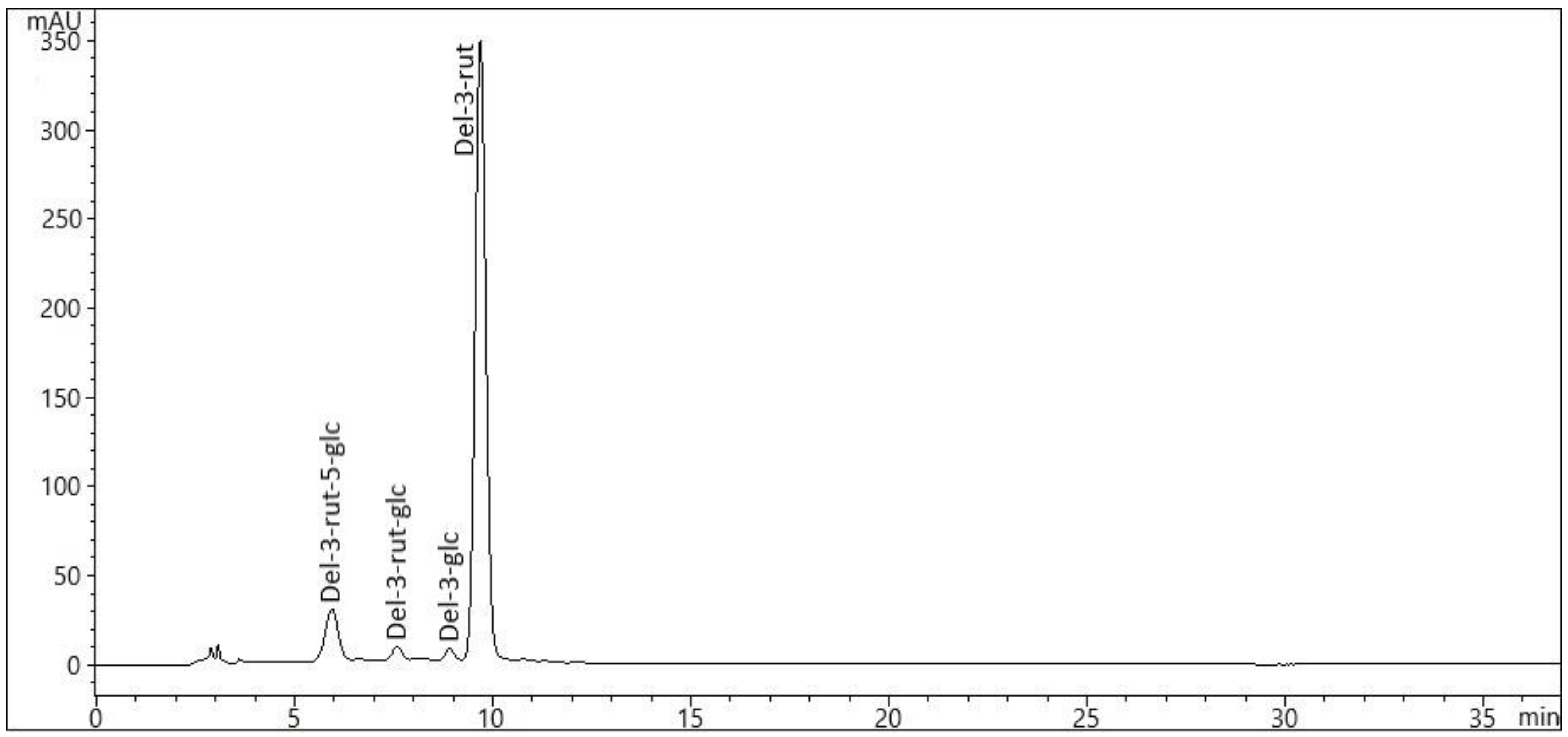
2.3. Variation in Anthocyanin Composition in Eggplants
2.4. Variation in α-Amylase Activity in Eggplants
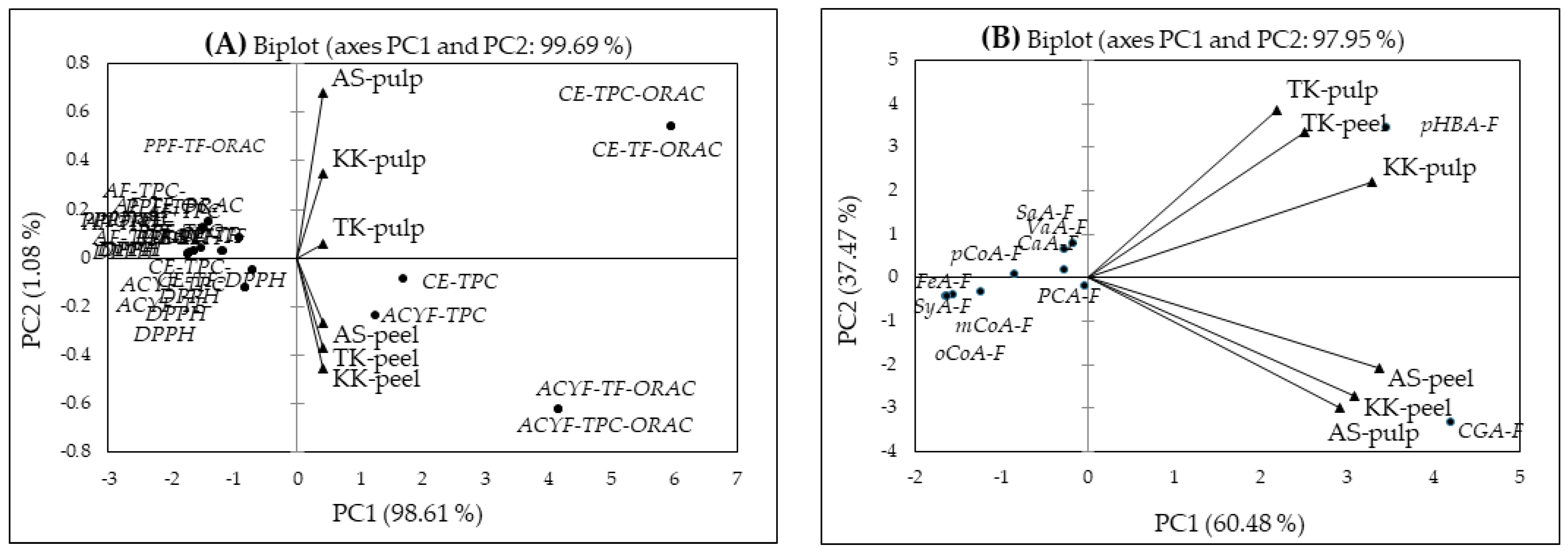
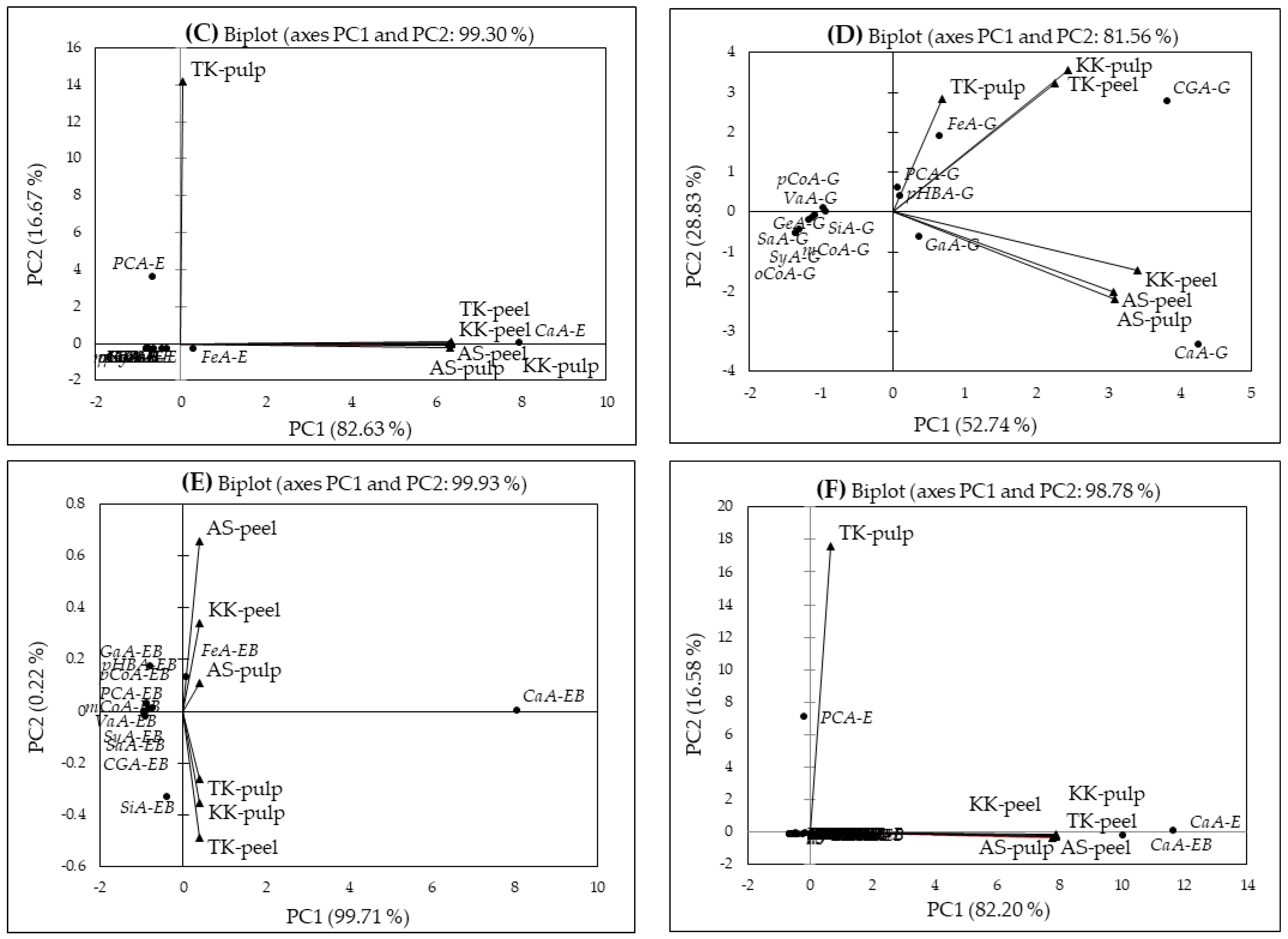
2.5. Principal Component Analysis (PCA)
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Extraction of Phenolics
3.4. Determination of Total Phenolic Compounds (TPC), Flavonoid (TF) and Anthocyanin (ACY) Contents
3.5. Determination of Antioxidant Capacity (AC)
3.6. UHPLC-MS/MS Determination of Phenolic Acids in Eggplants
3.7. Extraction and HPLC-DAD/ESI-MS Analysis of Anthocyanins in Eggplants
3.8. α-Amylase Inhibition Assay
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Salerno, L.; Modica, M.N.; Pittalà, V.; Romeo, G.; Siracusa, M.A.; Di Giacomo, C.; Sorrenti, V.; Acquaviva, R. Antioxidant activity and phenolic content of microwave-assisted Solanum melongena Extracts. Sci. World J. 2014, 11, 719486. [Google Scholar]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Niño-Medina, G.; Urías-Orona, V.; Muy-Rangel, M.D.; Heredia, J.B. Structure and content of phenolics in eggplant (Solanum melongena)—A review. S. Afr. J. Bot. 2017, 111, 161–169. [Google Scholar] [CrossRef]
- Stommel, J.R.; Whitaker, B.D. Phenolic acid content and composition of eggplant fruit in a germplasm core subset. J. Am. Soc. Hortic. Sci. 2003, 128, 704–710. [Google Scholar] [CrossRef]
- Singh, A.P.; Luthria, D.; Wilson, T.; Vorsa, N.; Singh, V.; Banuelos, G.S.; Pasakdee, S. Polyphenols content and antioxidant capacity of eggplant. Food Chem. 2009, 114, 955–961. [Google Scholar] [CrossRef]
- Zaro, M.J.; Keunchkarian, S.; Chaves, A.R.; Vicente, A.R.; Concellón, A. Changes in bioactive compounds and response to postharvest storage conditions in purple eggplants as affected by fruit developmental stage. Postharvest Biol. Technol. 2014, 96, 110–117. [Google Scholar] [CrossRef]
- Zaro, M.J.; Ortiz, L.C.; Keunchkarian, S.; Chaves, A.R.; Vicente, A.R.; Cocellon, A. Chlorogenic acid retention in white and purple eggplant after processing and cooking. LWT Food Sci. Technol. 2015, 64, 802–808. [Google Scholar] [CrossRef]
- Boyacı, H.; Topçu, V. Development of eggplant hybrid cultivar ‘BATEM FILIZI’ and determination of yield performance. Derim 2014, 31, 11–22. [Google Scholar] [CrossRef][Green Version]
- Güvenç, İ. Türkiye ve Avrupa Birliği ülkelerinin sebze üretimi ve kendine yeterlilik bakımından karşılaştırılması. Yyü. Tar. Bil. Derg. 2018, 28, 530–535. [Google Scholar] [CrossRef][Green Version]
- Sadilova, E.; Stintzing, F.C.; Carle, R. Anthocyanins, colour and antioxidant properties of eggplant (Solanum melongena L.) and violet pepper (Capsicum annuum L.) peel extracts. Z. Naturforsch. C 2006, 7–8, 527–535. [Google Scholar] [CrossRef]
- Hanson, P.M.; Yang, R.Y.; Tsou, S.C.S.; Ledesma, D.; Engle, L.; Lee, T.C. Diversity on eggplant (Solanum melongena) for superoxide scavenging activity, total phenolics, and ascorbic acid. J. Food Comp. Anal. 2006, 19, 594–600. [Google Scholar] [CrossRef]
- Raigón, M.D.; Prohens, J.; Muñoz-Falcón, J.E.; Nuez, F. Comparison of eggplant landraces and commercial varieties for fruit content of phenolics, minerals, dry matter and protein. J. Food Comp. Anal. 2008, 21, 370–376. [Google Scholar] [CrossRef]
- Colak, N.; Primetta, A.K.; Riihinen, K.R.; Jaakola, L.; Grúz, J.; Strnad, M.; Torun, H.; Ayaz, F.A. Phenolic compounds and antioxidant capacity in different-colored and non-pigmented berries of bilberry (Vaccinium myrtillus L.). Food Biosci. 2017, 20, 67–78. [Google Scholar] [CrossRef]
- Hano, C.; Tungmunnithum, D. Plant polyphenols, more than just simple natural antioxidants: Oxidative stress, aging and age-related diseases. Medicines 2020, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant flavonoids: Chemical characteristics and biological activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Plazas, M.; Andújar, I.; Vilanova, S.; Hurtado, M.; Gramazio, P.; Herráiz, F.J.; Prohens, J. Breeding for chlorogenic acid content in eggplant: Interest and prospects. Not. Bot. Horti Agrobot. Cluj Napoca 2013, 41, 26–35. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- Luthria, D.L. A simplified UV spectral scan method for the estimation of phenolic acids and antioxidant capacity in eggplant pulp extracts. J. Funct. Foods 2012, 4, 238–242. [Google Scholar] [CrossRef]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. In vitro studies of eggplant (Solanum melongena) phenolics as inhibitors of key enzymes relevant for type 2 diabetes and hypertension. Bioresour. Technol. 2008, 99, 2981–2988. [Google Scholar] [CrossRef]
- Li, W.; Yuan, G.; Pan, Y.; Wang, C.; Chen, H. Network pharmacology studies on the bioactive compounds and action mechanisms of natural products for the treatment of diabetes mellitus: A Review. Front. Pharmacol. 2017, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Chaudhary, G.; Yadav, D.K. Genetic diversity of Indian isolates of Ralstonia solanacearum causing bacterial wilt of eggplant (Solanum melongena). Ind. J. Agric. Sci. 2017, 87, 1466–1475. [Google Scholar]
- Sharma, M.; Kaushik, P. Biochemical Composition of Eggplant Fruits: A Review. Appl. Sci. 2021, 11, 7078. [Google Scholar] [CrossRef]
- Sun, L.; Warren, F.J.; Netzel, G.; Gidley, M.J. 3 or 3′-Galloyl substitution plays an important role in association of catechins and theaflavins with porcine pancreatic α-amylase: The kinetics of inhibition of α-amylase by tea polyphenols. J. Funct. Foods 2016, 26, 144–156. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.; Miao, M. Inhibition of α-amylase by polyphenolic compounds: Substrate digestion, binding interactions and nutritional intervention. Trends Food Sci. Technol. 2020, 104, 190–207. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, Y.; Han, L.; Zhang, S.; Duan, X.; Sun, L.; Wang, M. Number of galloyl moieties and molecular flexibility are both important in alpha-amylase inhibition by galloyl-based polyphenols. Food Funct. 2020, 11, 3838–3850. [Google Scholar] [CrossRef]
- Torun, H.; Kolcuoğlu, Y.; Ayaz, F.A.; Çolak, A.; Glew, R.H. Characterization of polyphenol oxidase during three ripening stages of an eggplant (Solanum melongena L.) fruit: A local type in northeast Anatolia. Turk. J. Biochem. 2015, 40, 44–50. [Google Scholar]
- Akanitapichat, P.; Phraibung, K.; Nuchklang, K.; Prompitakkul, S. Antioxidant and hepatoprotective activities of five eggplant varieties. Food Chem. Toxicol. 2010, 48, 3017–3021. [Google Scholar] [CrossRef]
- Bor, J.Y.; Chen, H.Y.; Yen, G.H. Evaluation of antioxidant activity and inhibitory effect on nitric oxide production of some common vegetables. J. Agric. Food Chem. 2006, 54, 1680–1686. [Google Scholar] [CrossRef]
- Boulekbache-Makhlouf, L.; Medouni, L.; Medouni-Adrar, S.; Arkoub, L.; Madani, K. Effect of solvents extraction on phenolic content and antioxidant activity of the by product of eggplant. Ind. Crops Prod. 2013, 49, 668–674. [Google Scholar] [CrossRef]
- Valanciene, E.; Jonuskiene, I.; Syrpas, M.; Augustiniene, E.; Matulis, P.; Simonavicius, A.; Malys, N. Advances and prospects of phenolic acids production, biorefinery and analysis. Biomolecules 2020, 10, 874. [Google Scholar] [CrossRef] [PubMed]
- Gürbüz, N.; Uluişik, S.; Frary, A.; Frary, A.; Doğanlar, S. Health benefits and bioactive compounds of eggplant. Food Chem. 2018, 268, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Tanchev, S.S.; Ruskov, P.J.; Timberlake, C.F. The anthocyanins of Bulgarian aubergine (Solanum melongena). Phytochemistry 1970, 9, 1681–1682. [Google Scholar] [CrossRef]
- Azuma, K.; Ohyama, A.; Ippoushi, K.; Ichiyanagi, T.; Tekeuchi, A.; Saito, T.; Fukuoka, H. Structures and antioxidant activity of anthocyanins in many accessions of eggplant and its related species. J. Agric. Food Chem. 2008, 56, 10154–10159. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Prior, R.L. Identification and characterization of anthocyanins by high performance liquid chromatography-electrospray ionization-tandem mass spectrometry in common foods in the United States: Vegetables, nuts, and grains. J. Agric. Food Chem. 2005, 53, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Calumpang, C.L.F.; Saigo, T.; Watanabe, M.; Tohge, T. Cross-Species Comparison of Fruit-Metabolomics to Elucidate Metabolic Regulation of Fruit Polyphenolics Among Solanaceous Crops. Metabolites 2020, 10, 209. [Google Scholar] [CrossRef] [PubMed]
- Toppino, L.; Barchi, L.; Lo Scalzo, R.; Palazzolo, E.; Francese, G.; Fibiani, M.; D’Alessandro, A.; Papa, V.; Laudicina, V.A.; Sabatino, L.; et al. Mapping quantitative trait loci affecting biochemical and morphological fruit properties in eggplant (Solanum melongena L.). Front. Plant Sci. 2016, 7, 256. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Z.; Chu, G.; Huang, C.; Tian, S.; Zhao, Z.; Chen, G. Anthocyanin accumulation and molecular analysis of anthocyanin biosynthesis-associated genes in eggplant (Solanum melongena L.). J. Agric. Food Chem. 2014, 62, 2906–2912. [Google Scholar] [CrossRef]
- Stommel, J.R.; Dumm, J.M. Coordinated regulation of biosynthetic and regulatory genes coincides with Anthocyanin Accumulation in developing eggplant fruit. J. Am. Soc. Hortic. Sci. 2015, 140, 129–135. [Google Scholar] [CrossRef]
- Gisbert, C.; Dumm, J.M.; Prohens, J.; Vilanova, S.; Stommel, J.R. A spontaneous eggplant (Solanum melongena L.) color mutant conditions anthocyanin-free fruit pigmentation. Hortscience 2016, 51, 793–798. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Ren, L.; Lian, H.L.; Chen, H.Y. Molecular cloning and characterization of anthocyanin biosynthesis genes in eggplant (Solanum melongena L.). Acta Physiol. Plant. 2016, 38, 163. [Google Scholar] [CrossRef]
- Woodward, G.; Kroon, P.; Cassidy, A.; Kay, C. Anthocyanin stability and recovery: Implications for the analysis of clinical and experimental samples. J. Agric. Food Chem. 2009, 57, 5271–5278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef]
- Lachman, J.; Hamouz, K. Red and purple coloured potatoes as a significant antioxidant source in human nutrition—A review. Plant Soil Environ. 2005, 51, 477–482. [Google Scholar] [CrossRef]
- Miao, M.; Jiang, H.; Jiang, B.; Li, Y.; Cui, S.W.; Zhang, T. Structure elucidation of catechins for modulation of starch digestion. LWT Food Sci. Technol. 2014, 57, 188–193. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, P.; Zhou, M.; Wang, T.; Fang, S.; Shang, X.; Fu, X. Geographic variation in the chemical composition and antioxidant properties of phenolic compounds from Cyclocarya paliurus (Batal) Iljinskaja Leaves. Molecules 2018, 23, 2440. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Principal component analysis. WIREs Comp. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Hossain, M.; Li, J.; Sikdar, A.; Hasanuzzaman, M.; Uzizerimana, F.; Muhammad, I.; Yuan, Y.; Zhang, C.; Feng, B. Exogenous melatonin modulates the physiological and biochemical mechanisms of drought tolerance in tartary buckwheat (Fagopyrum tataricum (L.) Gaertn). Molecules 2020, 25, 2828. [Google Scholar] [CrossRef]
- Colak, N.; Kurt-Celebi, A.; Fauzan, R.; Torun, H.; Ayaz, F.A. The protective effect of exogenous salicylic and gallic acids ameliorates the adverse effects of ionizing radiation stress in wheat seedlings by modulating the antioxidant defence system. Plant Physiol. Biochem. 2021, 168, 526–545. [Google Scholar] [CrossRef]
- Torun, H.; Novák, O.; Mikulík, J.; Strnad, M.; Ayaz, F.A. The Effects of exogenous salicylic acid on endogenous phytohormone status in Hordeum vulgare L. under salt stress. Plants 2022, 11, 618. [Google Scholar] [CrossRef]
- Rodriguez-Saona, L.E.; Wrolstad, R.E. Extraction, Isolation, and Purification of Anthocyanins. Curr. Protoc. Food Anal. Chem 2001, 1, F1-1. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Huang, D.J.; Lin, C.D.; Chen, H.J.; Lin, Y.H. Antioxidant and antiproliferative activities of sweet potato [Ipomoea batatas (L.) Lam ‘Tainong 57′] constituents. Bot. Bull. Acad. Sinica 2004, 45, 179–186. [Google Scholar]
- Giusti, M.; Rodríguez-Saona, L.E.; Wrolstad, R.E. Molar absorptivity and color characteristics of acylated and non-acylated pelargonidin-based anthocyanins. J. Agric. Food Chem. 1999, 47, 4631–4637. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Ayaz, F.A.; Hayirlioglu-Ayaz, S.; Gruz, J.; Novak, O.; Strnad, M. Separation, characterization, and quantitation of phenolic acids in a little-known blueberry (Vaccinium arctostaphylos L.) fruit by HPLC-MS. J. Agric. Food Chem. 2005, 53, 8116–8122. [Google Scholar] [CrossRef]
- Gruz, J.; Novák, O.; Strnad, M. Rapid analysis of phenolic acids in beverages by UPLC–MS/MS. Food Chem. 2008, 111, 789–794. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, H.-J.; Choung, M.-G. Anthocyanin compositions and biological activities from the red petals of Korean edible rose (Rosa hybrida cv. Noblered). Food Chem. 2011, 129, 272–278. [Google Scholar] [CrossRef]
- Phan, M.A.T.; Wang, J.; Tang, J.; Lee, Y.L.; Ng, K. Evaluation of α-glucosidase inhibition potential of some flavonoids from Epimedium brevicornum. LWT Food Sci. Technol. 2013, 53, 492–498. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Rodríguez-Werner, M.; Schlösser, A.; Winterhalter, P.; Rimbach, G. Fractionation, enzyme inhibitory and cellular antioxidant activity of bioactives from purple sweet potato (Ipomoea batatas). Food Chem. 2017, 221, 447–456. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | ‘Aydin Siyahi’ (Black) | ‘Kadife Kemer’ (Purple) | ‘Trabzon Kadife’ (White) | |||
|---|---|---|---|---|---|---|
| Peel | Pulp | Peel | Pulp | Peel | Pulp | |
| Total Phenolic Compounds (TPC) ‡ | ||||||
| Crude Phenolic (CE) | 17,193.44 ± 48.84 Fd | 6551.92 ± 26.743 Cd | 12,329.44 ± 52.24 Ed | 6027.75 ± 42.37 Bd | 8724.73 ± 27.07 Dd | 3875.04 ± 3121 Ad |
| Aqueous Fraction (AF) | 353.53 ± 0.47 Ca | 650.62 ± 1.84 Fb | 306.03 ± 0.60 Ba | 641.05 ± 3.71 Eb | 288.56 ± 7.01 Aa | 514.10 ± 8.72 Db |
| Polyphenolic Fraction (PPF) | 1341.20 ± 10.74 Fb | 609.53 ± 6.02 Da | 630.68 ± 12.46 Eb | 348.16 ± 4.25 Ba | 507.67 ± 3.47 Cb | 269.04 ± 5.16 Aa |
| Anthocyanin Fraction (ACYF) | 16,119.47 ± 49.74 Fc | 5397.43 ± 28.96 Cc | 11,586.29 ± 66.23 Ec | 5089.54 ± 9.19 Bc | 8010.09 ± 71.38 Dc | 2917.69 ± 14.83 Ac |
| Total Flavonoids (TF) § | ||||||
| Crude Phenolic (CE) | 3019.41 ± 20.65 Ed | 1159.74 ± 18.27 Bd | 1883.96 ± 56.74 Dd | 1130.19 ± 41.25 Bd | 1371.11 ± 22.32 Cd | 632.53 ±15.44 Ad |
| Aqueous Fraction (AF) | 22.50 ± 0.24 Fa | 21.60 ± 0.15 Da | 19.38 ± 0.09 Ca | 20.46 ± 0.37 Ca | 13.10 ± 0.13 Ba | 10.11 ± 0.08 Aa |
| Polyphenolic Fraction (PPF) | 180.88 ± 3.96 Fb | 85.92 ± 2.04 Db | 98.05 ± 1.30 Eb | 34.23 ± 0.97 Ab | 54.67 ± 0.64 Bb | 65.84 ± 1.38 Cb |
| Anthocyanin Fraction (ACYF) | 2883.76 ± 58.41 Fc | 1064.60 ± 25.40 Bc | 1742.59 ± 10.3 EDc | 1088.13 ± 44.09 Cc | 1322.73 ± 37.08 Dc | 614.79 ± 5.69 Ac |
| DPPH ¶ | ||||||
| Crude Phenolic (CE) | 8156.0 ± 81.76 Fd | 2334.53 ± 35.35 Cd | 3366.18 ± 103.22 Ed | 1453.16 ± 21.94 Bd | 2634.13 ± 17.81 Dc | 662.39 ± 27.35 Ad |
| Aqueous Fraction (AF) | 5.47 ± 0.28 Ba | 8.16 ± 0.34 Ca | 4.87 ± 0.22 Ba | 10.16 ± 0.13 Da | 4.16 ± 0.36 Aa | 10.61 ± 0.61 Da |
| Polyphenolic Fraction (PPF) | 579.51 ± 3.89 Fb | 291.32 ± 9.96 Eb | 265.34 ± 7.30 Db | 131.25 ± 3.85 Ab | 218.53 ± 6.47 Cb | 203.22 ± 9.45 Bb |
| Anthocyanin Fraction (ACYF) | 7646.63 ± 27.84 Fc | 1872.22 ± 14.05 Cc | 3210.73 ± 38.79 Ec | 1080.95 ± 20.22 Bc | 2608.37 ± 73.41 Dc | 541.24 ± 8.65 Ac |
| ORAC ¶ | ||||||
| Crude Phenolic (CE) | 37,886.96 ± 124.19 Fd | 17,648.20 ± 22.19 Dd | 22,670.88 ± 16.19 Ed | 14,352.79 ± 131.19 Bd | 19,554.97 ± 101.19 Cd | 8392.16 ± 42.19 Ad |
| Aqueous Fraction (AF) | 1217.64 ± 14.60 Da | 990.43 ± 37.42 Ca | 522.12 ± 28.61 Aa | 899.34 ± 6.17 Ba | 505.68 ± 38.59 Aa | 516.39 ± 15.27 Aa |
| Polyphenolic Fraction (PPF) | 5036.76 ± 14.80 Fb | 1878.19 ± 6.86 Db | 2607.07 ± 22.87 Eb | 1559.02 ± 5.54 Cb | 1314.20 ± 10.88 Bb | 915.11 ± 12.39 Ab |
| Anthocyanin Fraction (ACYF) | 31,929.87 ± 244.71 Ec | 9718.59 ± 155.84 Bc | 20,871.84 ± 330.72 Dc | 9484.73 ± 180.31 Bc | 17,676.07 ± 207.51 Cc | 6482.34 ± 21.79 Ac |
| Total Anthocyanin (ACY) ∞ | 16,835.45 ± 53.42 Fe | 6167.11 ± 33.71 Cc | 12,081.72 ± 42.51 Ee | 5715.37 ± 18.64 Bb | 8486.52 ± 63.62 Dd | 3218.81 ± 13.56 Aa |
| ‘Aydin Siyahi’, Black | ‘Kadife Kemer’, Purple | ‘Trabzon Kadife’, White | ||||
|---|---|---|---|---|---|---|
| Phenolic Acids | Peel | Pulp | Peel | Pulp | Peel | Pulp |
| Free (F) Form | ||||||
| Protocatechuic acid φ | 0.96 ± 0.05 C | 1.05 ± 0.03 C | 3.09 ± 0.08 D | 0.64 ± 0.01 B | 0.60 ± 0.03 B | 0.41 ± 0.02 A |
| p-Hydroxybenzoic acid φ | 2.15 ± 0.15 B | 0.65 ± 0.03 A | 5.57 ± 0.17 D | 2.06 ± 0.07 B | 5.00 ± 0.05 C | 2.07 ± 0.03 B |
| Vanilic acid φ | 0.51 ± 0.03 B | 0.18 ± 0.01 A | 0.99 ± 0.09 D | 0.83 ± 0.02 C | 0.70 ± 0.06 C | 0.81 ± 0.02 C |
| Salicylic acid φ | 0.63 ± 0.05 D | 0.17 ± 0.01 A | 0.50 ± 0.07 C | 0.80 ± 0.01 E | 0.34 ± 0.04 B | 0.81 ± 0.01 E |
| Syringic acid φ | 0.02 ± 0.00 A | 0.06 ± 0.00 BC | 0.25 ± 0.00 D | 0.05 ± 0.00 B | 0.07 ± 0.00 C | 0.05 ± 0.00 B |
| Caffeic acid δ | 0.25 ± 0.01 A | 0.61 ± 0.03 C | 3.77 ± 0.03 E | 0.56 ± 0.06 C | 1.05 ± 0.01 D | 0.43 ± 0.03 B |
| p-Coumaric acid δ | 0.42 ± 0.05 BC | 0.17 ± 0.00 A | 0.59 ± 0.08 D | 0.46 ± 0.03 C | 0.34 ± 0.02 B | 0.36 ± 0.01 B |
| Ferulic acid δ | 0.11 ± 0.00 A | 0.29 ± 0.05 B | 1.22 ± 0.13 C | 0.17 ± 0.00 A | 0.13 ± 0.01 A | 0.19 ± 0.01 AB |
| m-Coumaric acid δ | 0.03 ± 0.00 D | 0.02 ± 0.00 B | 0.03 ± 0.00 C | 0.02 ± 0.00 A | 0.03 ± 0.00 C | 0.02 ± 0.00 B |
| o-Coumaric acid δ | 0.06 ± 0.00 B | 0.01 ± 0.00 A | 0.11 ± 0.01 C | 0.01 ± 0.00 A | 0.06 ± 0.00 B | 0.02 ± 0.00 A |
| Ester (E) Form | ||||||
| Gallic acid φ | 458.47 ± 5.53 E | 144.81 ± 0.54 D | 72.37 ± 4.40 C | 8.19 ± 0.34 A | 14.87 ± 0.6 B | 10.40 ± 1.36 AB |
| Protocatechuic acid φ | 50.42 ± 0.10 F | 35.40 ± 0.53 E | 31.35 ± 1.27 D | 9.93 ± 0.16 A | 28.39 ± 0.25 C | 27.15 ± 0.92 B |
| Gentisic acid φ | ND | ND | ND | ND | ND | 0.02 ± 0.00 |
| p-Hydroxybenzoic acid φ | 30.37 ± 0.18 E | 8.01 ± 0.39 C | 10.38 ± 0.31 D | 3.32 ± 0.13 A | 7.04 ± 0.22 B | 3.44 ± 0.26 A |
| Vanilic acid φ | 1.17 ± 0.15 A | 1.50 ± 0.13 B | 2.15 ± 0.16 C | 1.16 ± 0.13 A | 1.15 ± 0.12 A | 1.19 ± 0.05 A |
| Syringic acid φ | 0.28 ± 0.00 A | 0.28 ± 0.01 A | 0.81 ± 0.08 C | 0.97 ± 0.12 D | 0.64 ± 0.02 B | 0.56 ± 0.02 C |
| Salicylic acid φ | 0.57 ± 0.04 D | 0.39 ± 0.03 B | 0.30 ± 0.02 A | 12.44 ± 0.03 E | 0.33 ± 0.02 A | 0.48 ± 0.00 C |
| Caffeic acid δ | 2620.35 ± 0.78 E | 2512.26 ± 18.34 D | 2830.44 ± 20.34 F | 1250.97 ± 37.80 A | 2136.59 ± 11.72 B | 2457.75 ± 0.35 C |
| p-Coumaric acid δ | 37.39 ± 0.53 B | 55.48 ± 1.61 E | 41.89 ± 2.36 C | 23.36 ± 1.17 A | 34.60 ± 1.43 B | 46.94 ± 2.17 D |
| Sinapic acid δ | 16.72 ± 0.13 A | 79.59 ± 0.71 C | 46.73 ± 1.74 B | 91.68 ± 2.63 D | 153.04 ± 2.82 F | 138.97 ± 1.91 E |
| Ferulic acid δ | 243.36 ± 3.40 C | 341.41 ± 3.02 E | 176.08 ± 2.98 A | 289.64 ± 20.71 D | 212.07 ± 0.01 B | 297.74 ± 1.73 D |
| m-Coumaric acid δ | 0.18 ± 0.00 A | 0.18 ± 0.02 A | 0.22 ± 0.01 B | 0.16 ± 0.01 A | 0.22 ± 0.00 B | 0.27 ± 0.00 C |
| o-Coumaric acid δ | ND | ND | ND | ND | ND | 0.01 ± 0.00 |
| Glycoside (G) Form | ||||||
| Gallic acid φ | 399.07 ± 19.01 B | 17.49 ± 0.27 A | 17.34 ± 0.18 A | 7.89 ± 0.59 A | 14.22 ± 0.16 A | 13.09 ± 0.23 A |
| Protocatechuic acid φ | 43.60 ± 0.04 F | 38.91 ± 0.65 E | 24.47 ± 0.05 B | 14.64 ± 0.35 A | 26.88 ± 0.79 C | 30.81 ± 0.06 D |
| Gentisic acid φ | 1.14 ± 0.09 B | 0.63 ± 0.01 A | 0.26 ± 0.01 A | 9.79 ± 0.06 D | 0.49 ± 0.00 A | 2.86 ± 0.02 C |
| p-Hydroxybenzoic acid | 85.19 ± 3.60 E | 51.75 ± 0.60 D | 18.57 ± 0.14 B | 9.82 ± 0.25 A | 16.38 ± 0.06 B | 37.74 ± 0.23 C |
| Vanilic acid φ | 7.31 ± 0.29 C | 6.22 ± 0.28 B | 3.58 ± 0.05 A | 9.12 ± 0.07 D | 3.59 ± 0.11 A | 9.26 ± 0.45 D |
| Syringic acid φ | 1.19 ± 0.03 B | 1.99 ± 0.04 D | 0.83 ± 0.02 A | 3.87 ± 0.09 E | 1.71 ± 0.09 C | 6.68 ± 0.07 F |
| Salicylic acid φ | 0.26 ± 0.02 B | 0.15 ± 0.00 AB | 0.06 ± 0.00 A | 9.17 ± 0.19 D | 0.21 ± 0.01 AB | 1.30 ± 0.01 C |
| Caffeic acid δ | 611.86 ± 8,65 E | 195.00 ± 5.75 B | 220.36 ± 2.45 C | 9.17 ± 0.19 D | 0.21 ± 0.01 AB | 1.30 ± 0.01 C |
| p-Coumaric acid δ | 7.10 ± 0.28 D | 2.95 ± 0.02 B | 2.40 ± 0.07 A | 8.40 ± 0.09 E | 3.33 ± 0.09 C | 12.97 ± 0.02 F |
| Sinapic acid δ | 1.09 ± 0.04 D | 0.72 ± 0.00 B | 0.81 ± 0.02 C | 0.70 ± 0.00 B | 0.21 ± 0.00 A | 1.84 ± 0.02 E |
| Ferulic acid δ | 10.66 ± 0.20 A | 31.42 ± 0.38 D | 48.02 ± 0.72 E | 29.83 ± 0.46 C | 21.79 ± 0.48 B | 59.86 ± 0.83 F |
| m-Ccoumaric acid δ | 0.04 ± 0.00 C | 0.03 ± 0.00 B | 0.01 ± 0.00 A | 0.03 ± 0.00 B | 0.05 ± 0.00 D | 0.04 ± 0.00 C |
| o-Coumaric acid δ | 0.04 ± 0.00 E | 0.03 ± 0.00 D | 0.02 ± 0.00 C | 0.01 ± 0.00 B | 0.00 ± 0.00 A | 0.02 ± 0.00 C |
| Ester-Bound (EB) Form | ||||||
| Gallic acid φ | 181.16 ± 1,60 E | 19.47 ± 0.42 D | 11.64 ± 0.34 B | 1.42 ± 0.02 A | 14.65 ± 0.22 C | 1.34 ± 0.03 A |
| Protocatechuic acid φ | 39.09 ± 0.72 F | 28.99 ± 0.38 E | 21.21 ± 0.42 C | 5.19 ± 0.06 A | 24.45 ± 0.57 D | 14.00 ± 0.11 B |
| p-Hydroxybenzoic acid φ | 41.51 ± 0.60 E | 8.73 ± 0.38 B | 12.17 ± 0.27 D | 3.00 ± 0.10 A | 10.12 ± 0.11 C | 2.66 ± 0.06 A |
| Vanilic acid φ | 1.58 ± 0.06 C | 1.52 ± 0.00 C | 3.79 ± 0.01 E | 1.94 ± 0.08 D | 1.31 ± 0.04 B | 0.82 ± 0.03 A |
| Syringic acid φ | 0.20 ± 0.00 A | 0.32 ± 0.00 B | 1.50 ± 0.04 E | 1.10 ± 0.04 D | 0.88 ± 0.03 C | 0.35 ± 0.00 B |
| Salicylic acid φ | 1.23 ± 0.01 D | 0.87 ± 0.02 B | 1.15 ± 0.00 C | 13.16 ± 0.08 E | 0.88 ± 0.01 B | 0.50 ± 0.07 A |
| Caffeic acid δ | 2384.72 ± 24.34 E | 1824.14 ± 24.98 C | 2593.88 ± 8.71 D | 1072.75 ± 11.66 A | 2067.31 ± 31.95 D | 1122.41 ± 5.52 B |
| p-Coumaric acid δ | 70.30 ± 1.02 F | 47.55 ± 0.13 C | 52.37 ± 0.60 D | 17.24 ± 0.05 A | 56.05 ± 1.02 E | 22.58 ± 0.22 B |
| Sinapic acid δ | 45.67 ± 1.21 B | 103.91 ± 2.97 D | 16.97 ± 0.39 A | 104.22 ± 3.24 D | 234.92 ± 9.87 E | 84.51 ± 1.58 C |
| Ferulic acid δ | 335.09 ± 6.71 F | 265.41 ± 1.85 D | 273.39 ± 1.06 E | 117.99 ± 0.51 B | 197.17 ± 8.26 C | 93.73 ± 1.58 A |
| m-Coumaric acid δ | 0.33 ± 0.00 C | 0.20 ± 0.00 A | 0.40 ± 0.00 D | 0.20 ± 0.00 A | 0.28 ± 0.00 B | 0.20 ± 0.00 A |
| ‘Aydin Siyahi’, Black | ‘Kadife Kemer’, Purple | ‘Trabzon Kadife’, White | ||||
|---|---|---|---|---|---|---|
| Chlorogenic Acid (CGA) | Peel | Pulp | Peel | Pulp | Peel | Pulp |
| Free form (F) | 6.08 ± 0.44 D ** | 4.71 ± 0.10 C | 27.55 ± 0.82 E | 1.14 ± 0.11 B | 0.69 ± 0.03 AB | 0.22 ± 0.02 A |
| Ester form (E) | 7.82 ± 0.36 E | 3.06 ± 0.23 D | 0.46 ± 0.09 BC | 0.00 ± 0.00 A | 0.54 ± 0.07 C | 0.15 ± 0.00 AB |
| Glycoside form (G) | 294.05 ± 4.81 F | 52.28 ± 0.94 B | 108.16 ± 1.26 D | 60.12 ± 0.20 C | 213.45 ± 2.67 E | 15.68 ± 0.54 A |
| Ester-bound form (EB) | 2.41 ± 0.23 E | 1.23 ± 0.04 D | 1.00 ± 0.01 C | 0.29 ± 0.02 B | 0.43 ± 0.01 B | 0.10 ± 0.00 A |
| Anthocyanin φ | Peak No | Retention Time (Rt, min) | λmax (nm) | MS [M + H]+ | ‘Aydin Siyahi’, Black | ‘Kadife Kemer’, Purple | ‘Trabzon Kadife’, White | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Peel | Pulp | Peel | Pulp | Peel | Pulp | |||||
| del-3-rut-5-glc | 1 | 5.9 | 525 | 773.1 | trace ψ | Trace | trace | trace | trace | trace |
| del-3-rut-glc | 2 | 7.6 | 522 | 773.1 | trace | Trace | trace | trace | trace | trace |
| del-3-glc | 3 | 8.9 | 528 | 465.1 | trace | Trace | trace | trace | trace | trace |
| del-3-rut | 4 | 9.7 | 526 | 611.1 | 1162.22 ± 5.56 D * | 194.62 ± 4.54 B | 336.59 ± 11.20 C | 45.45 ± 1.70 A | 215.11 ± 2.9 2 B | 72.44 ± 1.97 A |
| Total Phenolic Contents and Antioxidant Capacity φ | F1 | Phenolic Acid δ | F1 | F2 | Phenolic Acid δ | F1 | F2 | Phenolic Acid δ | F1 | F2 | F3 | Phenolic Acid δ | F1 | Phenolic Acid δ | F1 | Phenolic Acid δ | F1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CE-TPC | 1.696 | PCA-F | −0.040 | −0.189 | GaA-E | −0.312 | −0.305 | GaA-G | 0.362 | −0.610 | −0.118 | GaA-EB | −0.775 | PCA-F | −0.618 | CGA-E | −0.613 |
| AF-TPC | −1.499 | p-HBA-F | 3.454 | 3.452 | PCA-E | −0.657 | 3.594 | PCA-G | 0.076 | 0.629 | 0.854 | PCA-EB | −0.822 | p-HBA-F | −0.607 | GaA-G | −0.201 |
| PPF-TPC | −1.504 | VaA-F | −0.164 | 0.784 | GeA-E | −0.787 | −0.305 | GeA-G | −1.078 | −0.089 | −0.482 | p-HBA-EB | −0.871 | VaA-F | −0.620 | PCA-G | −0.465 |
| ACYF-TPC | 1.250 | SaA-F | −0.278 | 0.651 | p-HBA-E | −0.744 | −0.305 | p-HBA-G | 0.110 | 0.401 | 1.372 | VaA-EB | −0.917 | SaA-F | −0.621 | GeA-G | −0.603 |
| CE-TF | −1.157 | SyA-F | −1.568 | −0.407 | VaA-E | −0.781 | −0.305 | VaA-G | −0.936 | 0.020 | −0.152 | SyA-EB | −0.922 | SyA-F | −0.624 | p-HBA-G | −0.440 |
| AF-TF | −1.728 | CaA-F | −0.273 | 0.170 | SyA-E | −0.784 | −0.305 | SyA-G | −1.175 | −0.206 | −0.240 | SaA-EB | −0.904 | CaA-F | −0.618 | VaA-G | −0.586 |
| PPF-TF | −1.701 | p-CoA-F | −0.843 | 0.076 | SaA-E | −0.768 | −0.306 | SaA-G | −1.113 | −0.148 | −0.554 | CaA-EB | 8.055 | p-CoA-F | −0.622 | SyA-G | −0.611 |
| ACYF-TF | −1.186 | FeA-F | −1.231 | −0.328 | CaA-E | 7.960 | 0.017 | CaA-G | 4.258 | −3.325 | 0.387 | p-CoA-EB | −0.718 | FeA-F | −0.622 | SaA-G | −0.606 |
| CE-TPC-DPPH | −0.704 | m-CoA-F | −1.641 | −0.443 | p-CoA-E | −0.636 | −0.299 | p-CoA-G | −0.974 | 0.113 | 0.025 | SiA-EB | −0.374 | m-CoA-F | −0.624 | CaA-G | 0.338 |
| AF-TPC-DPPH | −1.732 | o-CoA-F | −1.631 | −0.444 | SiA-E | −0.438 | −0.289 | SiA-G | −1.306 | −0.451 | −0.447 | FeA-EB | 0.093 | o-CoA-F | −0.624 | p-CoA-G | −0.592 |
| PPF-TPC-DPPH | −1.622 | CGA-F | 4.215 | −3.323 | FeA-E | 0.299 | −0.274 | FeA-G | 0.655 | 1.898 | 2.210 | m-CoA-EB | −0.925 | CGA-F | −0.587 | SiA-G | −0.620 |
| ACYF-TPC-DPPH | −0.821 | m-CoA-E | −0.786 | −0.305 | m-CoA-G | −1.352 | −0.513 | −0.534 | CGA-EB | −0.922 | GaA-E | 0.035 | FeA-G | −0.457 | |||
| CE-TPC-ORAC | 5.947 | o-CoA-E | −0.787 | −0.305 | o-CoA-G | −1.352 | −0.515 | −0.534 | PCA-E | −0.200 | m-CoA-G | −0.624 | |||||
| AF-TPC-ORAC | −1.394 | CGA-E | −0.779 | −0.305 | CGA-G | 3.824 | 2.795 | −1.787 | GeA-E | −0.624 | o-CoA-G | −0.624 | |||||
| PPF-TPC-ORAC | −0.915 | p-HBA-E | −0.565 | CGA-G | 0.141 | ||||||||||||
| ACYF-TPC-ORAC | 4.155 | VaA-E | −0.616 | GaA-EB | −0.412 | ||||||||||||
| CE-TF-DPPH | −0.704 | SyA-E | −0.620 | PCA-EB | −0.503 | ||||||||||||
| AF-TF-DPPH | −1.732 | SaA-E | −0.598 | p-HBA-EB | −0.550 | ||||||||||||
| PPF-TF-DPPH | −1.622 | CaA-E | 11.632 | VaA-EB | −0.613 | ||||||||||||
| ACYF-TF-DPPH | −0.821 | p-CoA-E | −0.412 | SyA-EB | −0.619 | ||||||||||||
| CE-TF-ORAC | 5.947 | SiA-E | −0.136 | SaA-EB | −0.594 | ||||||||||||
| AF-TF-ORAC | −1.394 | FeA-E | 0.910 | CaA-EB | 10.054 | ||||||||||||
| PPF-TF-ORAC | −0.915 | m-CoA-E | −0.623 | p-CoA-EB | −0.370 | ||||||||||||
| ACYF-TF-ORAC | 4.155 | o-CoA-E | −0.624 | SiA-EB | 0.005 | ||||||||||||
| FeA-EB | 0.640 | ||||||||||||||||
| m-CoA-EB | −0.623 | ||||||||||||||||
| CGA-EB | −0.619 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colak, N.; Kurt-Celebi, A.; Gruz, J.; Strnad, M.; Hayirlioglu-Ayaz, S.; Choung, M.-G.; Esatbeyoglu, T.; Ayaz, F.A. The Phenolics and Antioxidant Properties of Black and Purple versus White Eggplant Cultivars. Molecules 2022, 27, 2410. https://doi.org/10.3390/molecules27082410
Colak N, Kurt-Celebi A, Gruz J, Strnad M, Hayirlioglu-Ayaz S, Choung M-G, Esatbeyoglu T, Ayaz FA. The Phenolics and Antioxidant Properties of Black and Purple versus White Eggplant Cultivars. Molecules. 2022; 27(8):2410. https://doi.org/10.3390/molecules27082410
Chicago/Turabian StyleColak, Nesrin, Aynur Kurt-Celebi, Jiri Gruz, Miroslav Strnad, Sema Hayirlioglu-Ayaz, Myoung-Gun Choung, Tuba Esatbeyoglu, and Faik Ahmet Ayaz. 2022. "The Phenolics and Antioxidant Properties of Black and Purple versus White Eggplant Cultivars" Molecules 27, no. 8: 2410. https://doi.org/10.3390/molecules27082410
APA StyleColak, N., Kurt-Celebi, A., Gruz, J., Strnad, M., Hayirlioglu-Ayaz, S., Choung, M.-G., Esatbeyoglu, T., & Ayaz, F. A. (2022). The Phenolics and Antioxidant Properties of Black and Purple versus White Eggplant Cultivars. Molecules, 27(8), 2410. https://doi.org/10.3390/molecules27082410