
Bioactives of Momordica charantia as Potential Anti-Diabetic/Hypoglycemic Agents

 and
and
Abstract
1. Introduction
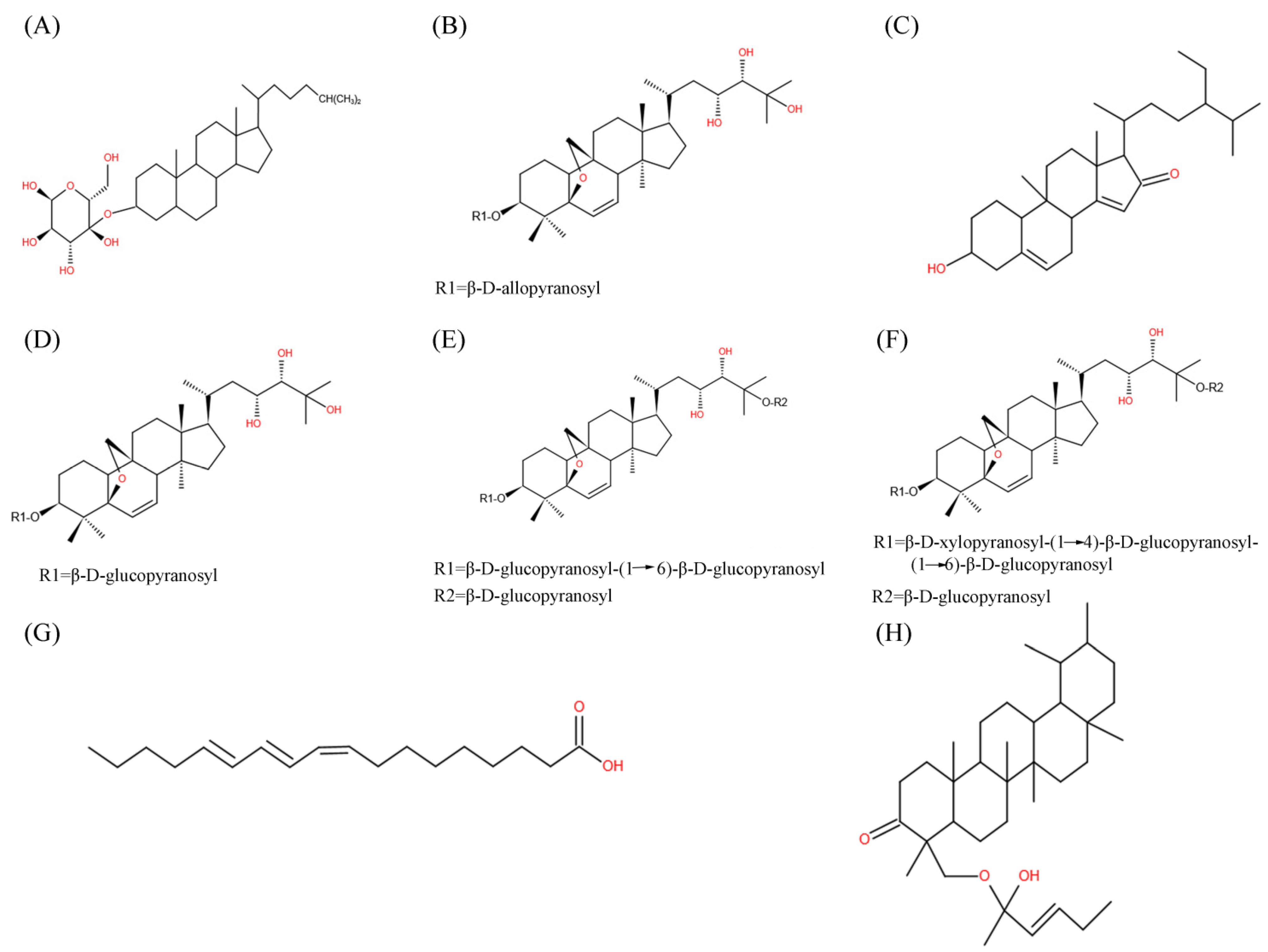
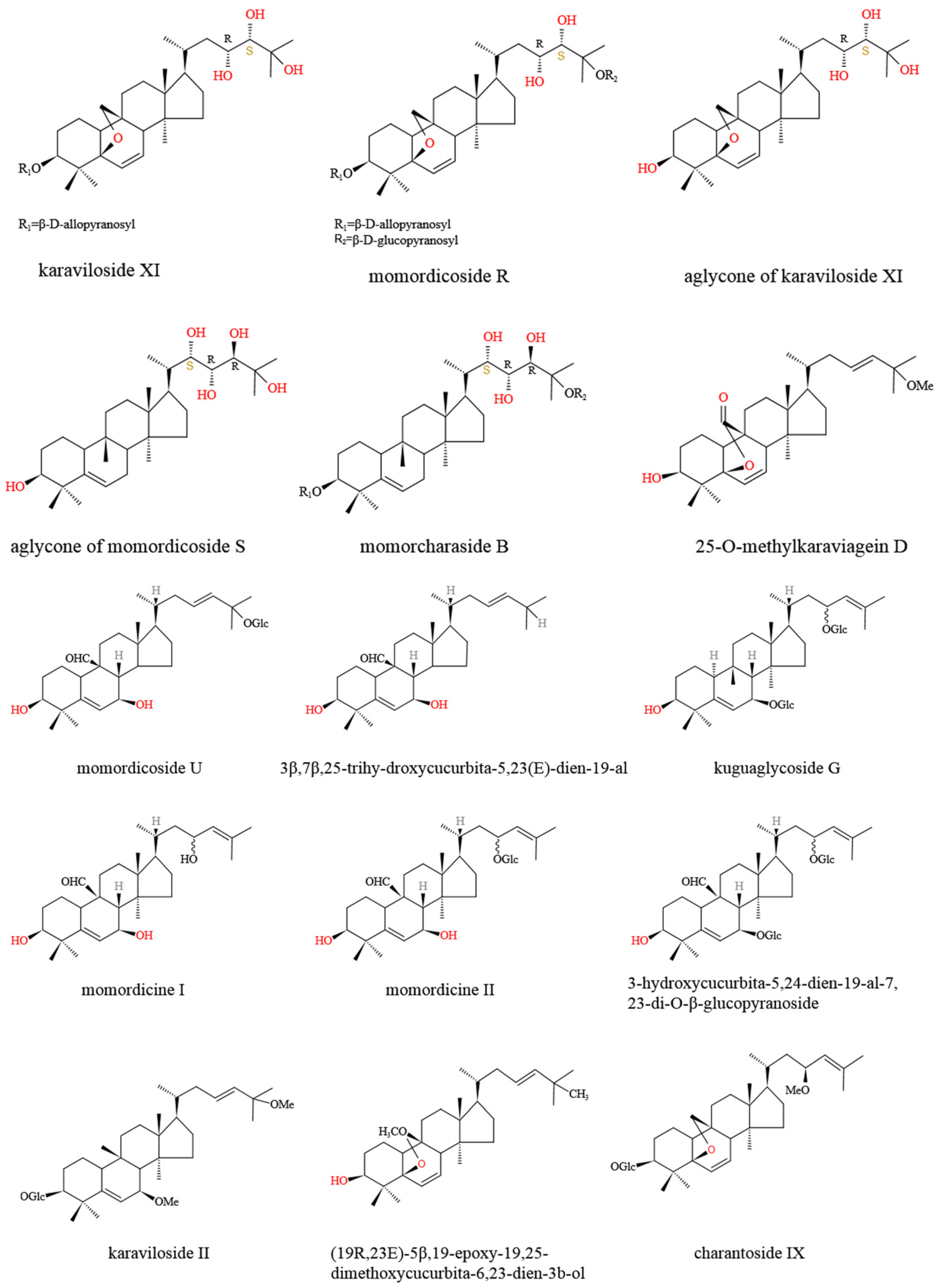
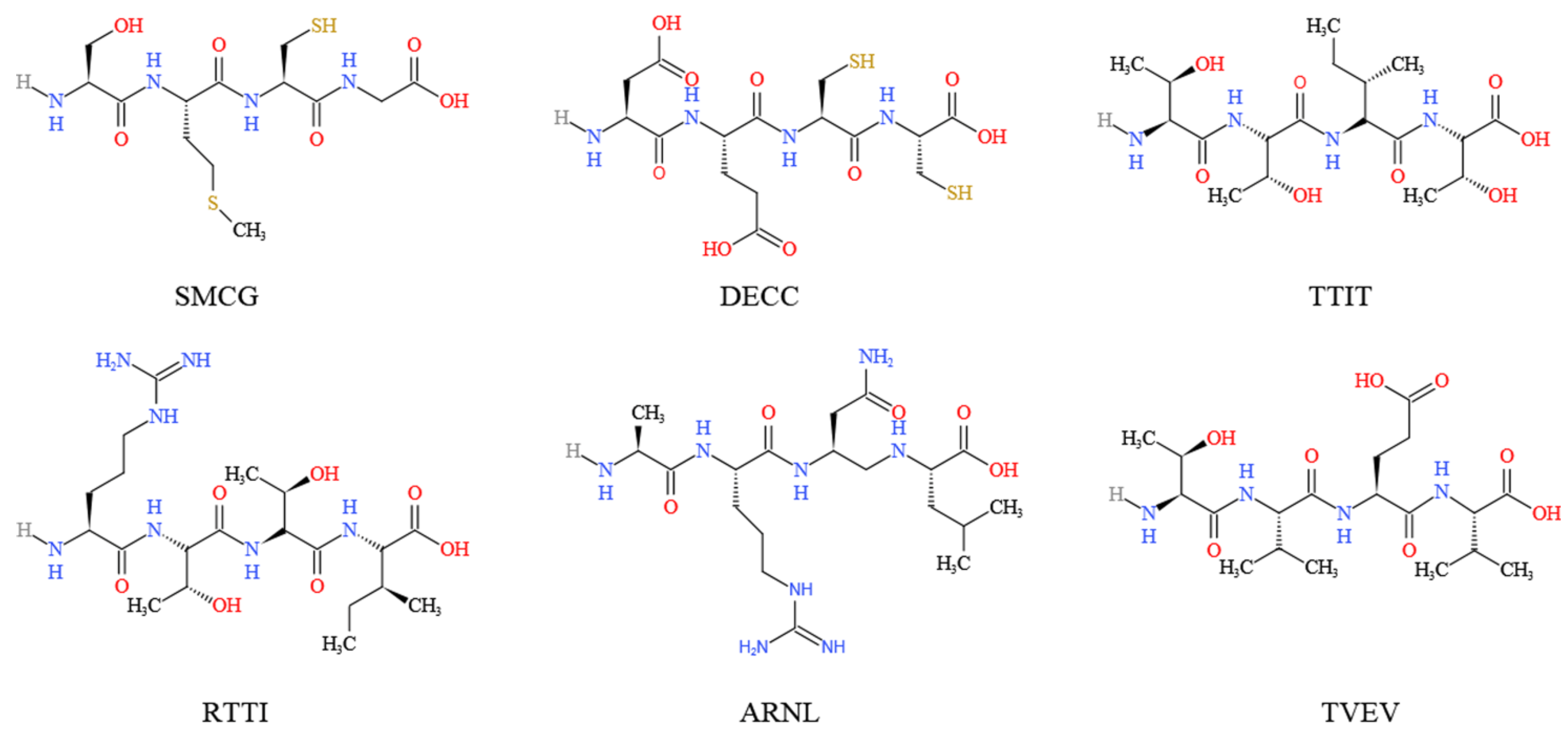
2. Bioactive Compounds of M. charantia with Hypoglycemic Potentials
3. Antidiabetic Activity of M. charantia
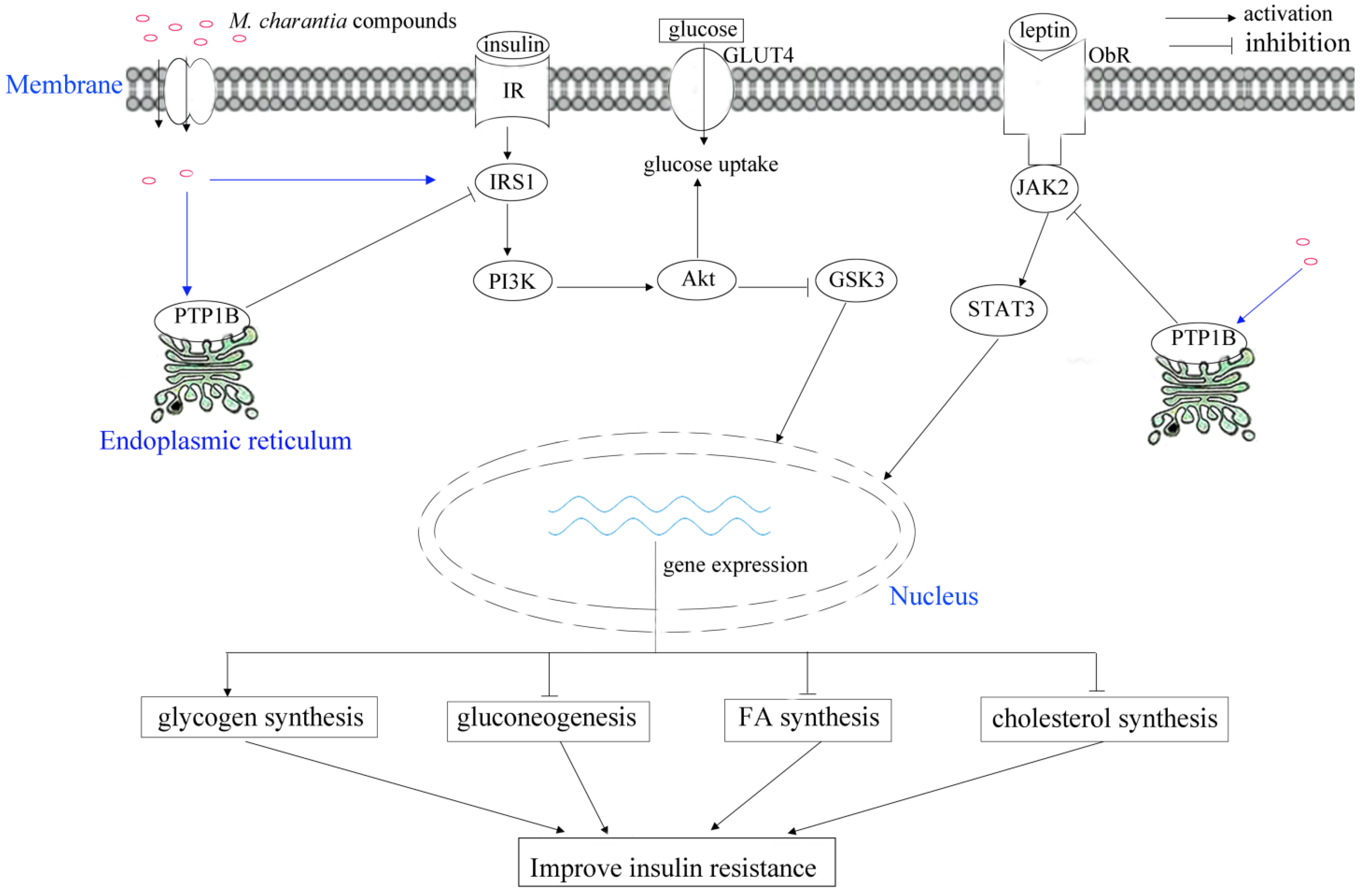
3.1. Improving Insulin Secretory and Resistance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
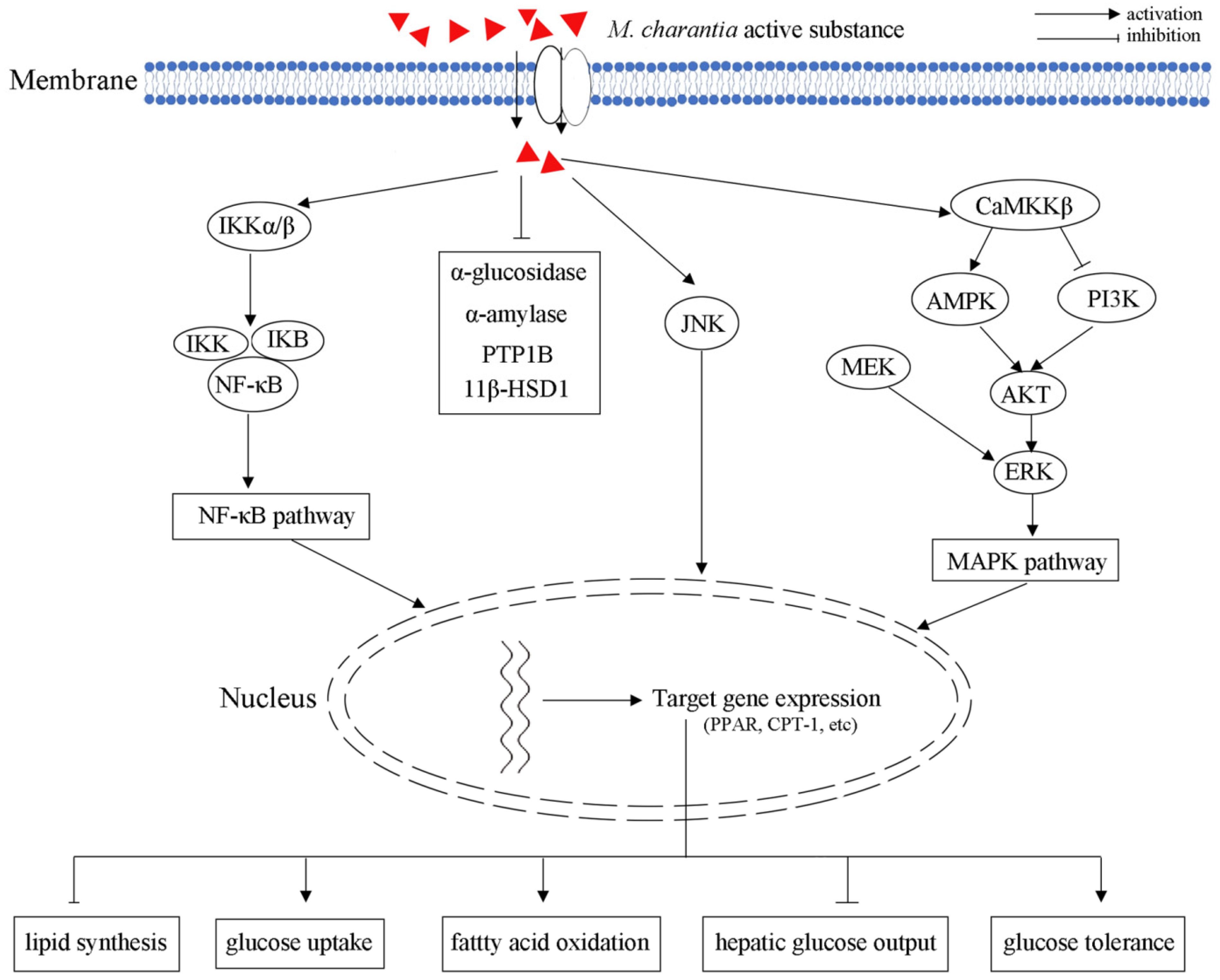{kind=link}
| Active Components | Dose | Model | Effect | References |
|---|---|---|---|---|
| water extract: platycodin-D (1:4) | _ | obese diabetic db/db mice | decrease obesity-related changes | [35] |
| acetone extract of whole fruit | 25–75 mg/100 g body weight | alloxan diabetic albino rats, | lower blood glucose, stimulate the recovery of pancreatic islet β cells | [36] |
| methanol extract of fruit | 200–600 mg/kg | normal and diabetic animals | lower blood glucose level, | [37] |
| alcoholic extract of fruit | 500 mg/kg | normal glucose primed rat | depress plasma glucose levels, enhance glycogen synthesis in liver | [38] |
| protein extract of fruit pulp | 5–10 mg/kg | normal and STZ-induced diabetic rats | exert insulin secretagogue and insulinomimetic activities, decrease plasma glucose concentrations, raise plasma insulin concentrations | [39] |
| fruit pulps | 2000 mg/day | patients with T2DM | hypoglycemic effect | [45] |
| powder | 2–4 g/day | patients with T2DM | improve blood lipids, atherogenic index, body weight, and systolic blood pressure, | [46] |
| fruit | _ | STZ-induced diabetic rats, male high-fat-fed Wistar rats, rat L6 myotubes, male Sprague-Dawley rats with diabetes | increase number of pancreatic β cells, improve insulin sensitivity, reduce fasting insulin, increase glucose uptakes, improve wound healing, increase diversity and shift overall structure of gut microbiota, stimulate amino acid uptake, normalise structural abnormalities of peripheral nerves, reduce glucose absorptions, improve body mass gain and LDL cholesterol values | [48,51,54,55,56,57,58] |
| Saponin-free methanol extract of juice | 150 mg/2 mL water | NIDDM model rats | improve insulin secretory capacity of B cells, enhance insulin action | [40] |
| saponin-rich fraction of fruit | 125 μg/mL | MIN6 β-cells | stimulate insulin secretion | [29] |
| dried powder of fruit pulp | 2000 mg/day | patients with T2DM | ameliorate diabetes associated CV risk, decrease level of glycosylated hemoglobin, increase insulin secretion | [46,49,59] |
| green fruit methanol extract and ethyl acetate fraction | INS-1 cells and rat pancreatic islets | increase ATP content, augment insulin secretion, increase serum insulin levels, decrease blood glucose levels | [50] | |
| momordicoside U | 15.8–197.2 μM | MIN6 β-cells | enhance insulin secretion | [30] |
| Momordicilin | block the active site of GSK-3, | [7] | ||
| mcIRBP | induce expression of GLUT4, stimulate phosphorylation of PDK1 and Akt, stimulate the uptake of glucose and clearance of glucose, | [27] | ||
| freeze-dried unripe fruit juice | 0.75% | rats fed high fat diets | improve insulin resistance, lower serum insulin and leptin, improve oral glucose tolerance, lower body weight and visceral fat mass, raise serum-free fatty acid concentration, reduce adiposity, | [53] |
| momordicosides (Q, R, S, and T) and karaviloside XI | _ | L6 myotubes, 3T3-L1 adipocytes, mice | enhance AMPK activity, stimulate GLUT4 translocation to the cell membrane, enhance fatty acid oxidation and glucose disposal | [32] |
| nanoparticles synthesized with filtrate of methanolic extract and silver nitrate | 50 mg/kg | STZ-induced diabetic rats | regulate signaling pathways, up-regulate expression level of glucokinase | [60] |
3.2. Regulating Glucose Uptake
3.3. Improving Glucose Metabolism
3.4. Modulating Lipid and Amino Acid Metabolism
3.5. Protective Effects of M. charantia
3.6. Inhibitory Effects of Related Enzymes
3.7. Regulation of Signal Pathways
4. Challenges and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AGE | advanced glycation end products |
| AMPK | Adenosine 5′-monophosphate (AMP)-activated protein kinase |
| adMc1 | anti-hyperglycemic protein of M. charantia |
| AUC | areas under the curve |
| 9c,11t,13t-CLN | 9cis,11trans,13trans-conjugated linolenic acid |
| CPT-1 | carnitine palmitoyltransferase I |
| Cr | chromium |
| DPP-IV | dipeptidyl peptidase-IV |
| GLUT | Glucose transporters |
| GLP-1 | glucagon-like peptide 1 |
| GIP | glucose-dependent insulinotropic peptide |
| HFD | high-fat-fed |
| IDDM | insulin-dependent diabetes mellitus |
| IR | insulin receptor |
| IRS-1 | insulin receptor subtrate-1 |
| NF-κB | nuclear factor-κB |
| NIDDM | non-insulin-dependent diabetes mellitus |
| PDK1 | protein kinase-1 |
| STZ | streptozocin |
| T1DM | Type 1 diabetes mellitus |
| T2DM | Type 2 diabetes mellitus |
| mcIRBP | M. charantia insulin receptor (IR)-binding protein |
| PPARα | Peroxisome proliferator activated receptor α |
| PTP1B | protein tyrosine phosphatase 1B |
| PPRE | peroxisome proliferator responsive element |
| SCFAs | short-chain fatty acid |
| SGLUTs | Sodium-coupled glucose transporters |
| SGLT1 | sodium dependent glucose transporter 1 |
| TAG | triacylglycerol |
References
- Xiao, J.B.; Högger, P. Dietary polyphenols and type 2 diabetes: Current insights and future perspectives. Curr. Med. Chem. 2015, 22, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Roglic, G.; Unwin, N.; Bennett, H.; Mathers, C.; Tuomilehto, J.; Nag, S.; Connolly, V.; King, H. The burden of mortality attributable to diabetes: Realistic estimates for the year 2000. Diabetes Care 2005, 28, 2130–2135. [Google Scholar] [CrossRef] [PubMed]
- ADA. Classification and diagnosis of diabetes. Diabetes Care 2015, 40, S11–S24. [Google Scholar]
- Zhao, Y.; Liu, Y.J.; Yi, F.Z.; Zhang, J.; Xu, Z.H.; Liu, Y.H.; Tao, Y. Type 2 diabetes mellitus impaired nasal immunity and increased the risk of hyposmia in COVID-19 mild pneumonia patients. Int. Immunopharmacol. 2021, 93, 107406. [Google Scholar] [CrossRef]
- Salamone, D.; Rivellese, A.A.; Vetrani, C. The relationship between gut microbiota, short-chain fatty acids and type 2 diabetes mellitus: The possible role of dietary fibre. Acta Diabetol. 2021, 58, 1131–1138. [Google Scholar] [CrossRef]
- Karagz, I.K.; Karagz, A.; Zkalayc, F.; Dogan, C.; Kocabay, G.; Elbay, A. Relation between platelet reactivity levels and diabetic retinopathy stage in patient with type 2 diabetes mellitus by using multiplate whole blood aggregometry. Semin. Ophthalmol. 2021, 36, 392–399. [Google Scholar] [CrossRef]
- Hazarika, R.; Parida, P.; Neog, B.; Yadav, R.N. Binding energy calculation of GSK-3 protein of human against some anti-diabetic compounds of Momordica charantia Linn (bitter melon). Bioinformation 2012, 8, 251–254. [Google Scholar] [CrossRef]
- Bunyatyan, N.D.; Bukhtiyarova, I.P.; Drogovoz, S.Z.; Kononenko, A.V.; Olefir, Y.V.; Prokof’ev, A.B.; Proskurina, I.A.; Goryachev, D.V. Influence of human biorhythms on the blood glucose level and the efficacy of hypoglycemic drugs (review). Pharm. Chem. J. 2017, 51, 399–401. [Google Scholar] [CrossRef]
- Lee, S.H.; Min, K.H.; Han, J.S.; Lee, D.H.; Park, D.B.; Jung, W.K.; Park, P.; Jeon, B.T.; Kim, S.K.; Jeon, Y.J. Effects of brown alga, Ecklonia cava on glucose and lipid metabolism in C57BL/KsJ-db/db mice, a model of type 2 diabetes mellitus. Food Chem. Toxicol. 2012, 50, 575–582. [Google Scholar] [CrossRef]
- Zhao, C.; Yang, C.F.; Liu, B.; Lin, L.; Sarker, S.D.; Nahar, L.; Yu, H.; Cao, H.; Xiao, J.B. Bioactive compounds from marine macroalgae and their hypoglycemic benefits. Trends Food Sci. Technol. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Jung, M.; Park, M.; Lee, H.C.; Kang, Y.H.; Kang, E.S.; Kim, S.K. Antidiabetic agents from medicinal plants. Curr. Med. Chem. 2006, 13, 1203–1218. [Google Scholar] [CrossRef] [PubMed]
- Khan, V.; Najmi, A.K.; Akhtar, M.; Aqil, M.; Mujeed, M.; Pillai, K.K. A pharmacological appraisal of medicinal plants with antidiabetic potential. J. Pharm. Bioallied Sci. 2012, 4, 27–42. [Google Scholar] [PubMed]
- Lee, S.H.; Kang, N.; Kim, E.A.; Heo, S.J.; Moon, S.H.; Jeon, B.T.; Jeon, Y.J. Antidiabetogenic and antioxidative effects of octaphlorethol a isolated from the brown algae Ishige foliacea in streptozotocin-induced diabetic mice. Food. Sci. Biotechnol. 2014, 23, 1261–1266. [Google Scholar] [CrossRef]
- Vinayagam, R.; Xiao, J.B.; Xu, B.J. An insight into anti-diabetic properties of dietary phytochemicals. Phytochem. Rev. 2017, 16, 535–553. [Google Scholar] [CrossRef]
- Yue, J.; Xu, J.; Cao, J.; Zhang, X.S.; Zhao, Y.Q. Cucurbitane triterpenoids from Momordica charantia L. and their inhibitory activity against α-glucosidase, α-amylase and protein tyrosine phosphatase 1B (PTP1B). J. Funct. Foods 2017, 37, 624–631. [Google Scholar] [CrossRef]
- Mar, K.; Fassnacht, M.; Führer-Sakel, D.; Honegger, J.B.; Weber, M.M.; Kroiss, M. The diagnosis and management of endocrine side effects of immune checkpoint inhibitors. Dtsch. Arztebl. Int. 2021, 118, 33724917. [Google Scholar]
- Preiato, V.L.; Salvagni, S.; Ricci, C.; Ardizzoni, A.; Pelusi, C. Diabetes mellitus induced by immune checkpoint inhibitors: Type 1 diabetes variant or new clinical entity? review of the literature. Rev. Endocr. Metab. Dis. 2021, 22, 337–349. [Google Scholar] [CrossRef]
- Fauchier, L.; Fauchier, G.; Bisson, A.; Bodin, A.; Herbert, J.; Angoulvant, D.; Ducluzeau, P.H.; Lip, G.Y.H. Antidiabetic drugs use and new-onset atrial fibrillation in patients with diabetes mellitus. Eur. Heart J. 2021, 42, ehab724.0457. [Google Scholar] [CrossRef]
- Wu, C.H.; Hsieh, H.T.; Lin, J.A.; Yen, G.C. Alternanthera paronychioides protects pancreatic β-cells from glucotoxicity by its antioxidant, antiapoptotic and insulin secretagogue actions. Food Chem. 2013, 139, 362–370. [Google Scholar] [CrossRef]
- Leung, L.; Birtwhistle, R.; Kotecha, J.; Hannah, S.; Cuthbertson, S. Anti-diabetic and hypoglycaemic effects of Momordica charantia (bitter melon): A mini review. Br. J. Nutr. 2009, 102, 1703–1708. [Google Scholar] [CrossRef]
- Saeed, F.; Afzaal, M.; Niaz, B.; Arshad, M.U.; Tufail, T.; Hussain, M.B.; Javed, A. Bitter melon (Momordica charantia): A natural healthy vegetable. Int. J. Food Prop. 2018, 21, 1270–1290. [Google Scholar] [CrossRef]
- Tan, H.F.; Gan, C.Y. Polysaccharide with antioxidant, α-amylase inhibitory and ace inhibitory activities from Momordica charantia. Int. J. Biol. Macromol. 2016, 85, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Shen, M.; Zhang, F.; Xie, J.H. Recent advances in Momordica charantia: Functional components and biological activities. Int. J. Mol. Sci. 2017, 18, 2555. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Assefa, A.D.; Keum, Y.S. Fatty acid and carotenoid composition of bitter melon (Momordica charantia L.) seed arils: A potentially valuable source of lycopene. J. Food Meas. Charact. 2017, 11, 1266–1273. [Google Scholar] [CrossRef]
- Wang, S.; Li, Z.; Yang, G.; Ho, C.; Li, S. Momordica charantia: A popular health-promoting vegetable with multifunctionality. Food Funct. 2017, 8, 1749–1762. [Google Scholar] [CrossRef]
- Khanna, P.; Jain, S.C.; Panagariya, A.; Dixit, V.P. Hypoglycemic activity of polypeptide-p from a plant source. J. Nat. Prod. 1981, 44, 648–655. [Google Scholar] [CrossRef]
- Lo, H.Y.; Ho, T.Y.; Li, C.C.; Chen, J.C.; Liu, J.J.; Hsiang, C.Y. A novel insulin receptor-binding protein from Momordica charantia enhances glucose uptake and glucose clearance in vitro and in vivo through triggering insulin receptor signaling pathway. J. Agric. Food Chem. 2014, 62, 8952–8961. [Google Scholar] [CrossRef]
- Chuang, C.Y.; Hsu, C.; Chao, C.Y.; Wein, Y.S.; Kuo, Y.H.; Huang, C.J. Fractionation and identification of 9c, 11t, 13t-conjugated linolenic acid as an activator of PPARα in bitter gourd (Momordica charantia L.). J. Biomed. Sci. 2006, 13, 763–772. [Google Scholar] [CrossRef]
- Keller, A.C.; Ma, J.; Kavalier, A.; He, K.; Brillantes, A.M.B.; Kennelly, E.J. Saponins from the traditional medicinal plant Momordica charantia stimulate insulin secretion in vitro. Phytomedicine 2011, 19, 32–37. [Google Scholar] [CrossRef]
- Ma, J.; Whittaker, P.; Keller, A.; Mazzola, E.P.; Pawar, R.S.; White, K.D.; Callahan, J.H.; Kennelly, E.J.; Krynitsky, A.J.; Rader, J.I. Cucurbitane-type triterpenoids from Momordica charantia. Plant. Med. 2010, 76, 1758–1761. [Google Scholar] [CrossRef]
- Iseli, T.J.; Nigel, T.; Zeng, X.Y.; Cooney, G.J.; Kraegen, E.W.; Yao, S.; Ye, Y.; James, D.E.; Ye, J.M. Activation of AMPK by bitter melon triterpenoids involves CaMKKβ. PLoS ONE 2013, 8, e62309. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.J.; Ye, J.M.; Turner, N.; Hohnen-Behrens, C.; Ke, C.Q.; Tang, C.P.; Chen, T.; Weiss, H.C.; Gesing, E.R.; Rowland, A.; et al. Antidiabetic activities of triterpenoids isolated from bitter melon associated with activation of the AMPK pathway. Chem. Biol. 2008, 15, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Harinantenaina, L.; Tanaka, M.; Takaoka, S.; Oda, M.; Mogami, O.; Uchid, M.; Askawa, Y. Momordica charantia constituents and antidiabetic screening of the isolated major compounds. Chem. Pharm. Bull. 2006, 54, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Jirousek, M.R.; Johnson, T.O.; Jacques, E. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat. Rev. Drug Discov. 2002, 1, 696–709. [Google Scholar]
- Sohn, J.H.; Kim, J.W.; Jung, G.W.; Park, D.C.; Moon, S.B.; Cho, H.R.; Ku, S.K.; Choi, J.S. Synergic antiobesity effects of bitter melon water extract and platycodin-D in genetically obese mice. J. Environ. Biol. 2018, 39, 603–611. [Google Scholar] [CrossRef]
- Singh, N.; Gupta, M. Regeneration of beta cells in islets of langerhans of pancreas of alloxan diabetic rats by acetone extract of Momordica charantia (Linn.) (bitter gourd) fruits. Indian J. Exp. Biol. 2007, 45, 1055–1062. [Google Scholar]
- Kolawole, O.T.; Abiona, F.E.; Kolawole, S.O.; Ayankunle, A.A.; OIaniran, O.I. Effect of Momordica charantia fruit extract on normal and alloxan-diabetic rats. Int. J. Pharmacol. 2011, 7, 532–535. [Google Scholar] [CrossRef]
- Sarkar, S.; Pranava, M.; Marita, R.A. Demonstration of the hypoglycemic action of Momordica charantia in a validated animal model of diabetes. Pharmacol. Res. 1996, 33, 1–4. [Google Scholar] [CrossRef]
- Yibchok-Anun, S.; Adisakwattana, S.; Yao, C.Y. Slow acting protein extract from fruit pulp of Momordica charantia with insulin secretagogue and insulinomimetic activities. Biol. Pharm. Bull. 2006, 29, 1126–1131. [Google Scholar] [CrossRef]
- Ali, L.; AzadKan, K.; Rouf Mamun, M.I.; Mosihuzzaman, M.; Nahar, N.; Nur-e-Alam, M.; Rokeya, B. Studies on hypoglycemic effects of fruit pulp, seed, and whole plant of Momordica charantia on normal and diabetic model rats. Plant. Med. 1993, 59, 408–412. [Google Scholar] [CrossRef]
- Pina, F.; Brown, A.; Luebcke, E.; Perkins-Veazie, P.; Clarke, S.; Kuvibidila, S.; Hill, M.; Linghtfoot, S.; Smith, B.; Lucas, E. Momordica charantia improves body weight and glucose tolerance in mice fed high fat diet. FASEB J. 2009, 23, 563.37. [Google Scholar] [CrossRef]
- Burnett, A.A.J.; Singh, P.D.; Simon, O.; Mckoy, M.L.G. Effect of acute administration of Momordica charantia fruit extracts on blood glucose levels of normoglycemic rats. FASEB J. 2011, 25, 889–890. [Google Scholar]
- Kim, S.K.; Jung, J.; Jung, J.H.; Yoonb, N.A.; Kangb, S.S.; Rohb, G.S.; Hahma, J.R. Hypoglycemic efficacy and safety of Momordica charantia (bitter melon) in patients with type 2 diabetes mellitus. Complement. Ther. Med. 2020, 52, 102524. [Google Scholar] [CrossRef] [PubMed]
- Krawinkel, M.B.; Ludwig, C.; Swai, M.E.; Yang, R.Y.; Chun, K.P.; Habicht, S.D. Bitter gourd reduces elevated fasting plasma glucose levels in an intervention study among prediabetics in Tanzania. J. Ethnopharmacol. 2018, 216, 1–7. [Google Scholar] [CrossRef]
- Fuangchan, A.; Sonthisombat, P.; Seubnukarn, T.; Chanouan, P.; Chotchaisuwat, P.; Sirigulsatien, V.; Lngkaninan, K.; Plianbangchang, P.; Haines, S.T. Hypoglycemic effect of bitter melon compared with metformin in newly diagnosed type 2 diabetes patients. J. Ethnopharmacol. 2011, 134, 422–428. [Google Scholar] [CrossRef]
- Rahman, I.U.; Khan, R.; Rahman, K.; Bashir, M. Lower hypoglycemic but higher antiatherogenic effects of bitter melon than glibenclamide in type 2 diabetic patients. Nutr. J. 2015, 14, 13. [Google Scholar] [CrossRef]
- Cortez-Navarrete, M.; Méndez-Del Villar, M.; Ramos-González, E.J.; Pérez-Rubio, K.G. Momordica Charantia: A Review of Its effects on metabolic diseases and mechanisms of action. J. Med. Food 2021, 24, 1017–1027. [Google Scholar] [CrossRef]
- Ahmed, I.; Adeghate, E.; Sharma, A.K.; Pallot, D.J.; Singh, B. Effects of Momordica charantia fruit juice on islet morphology in the pancreas of the streptozotocin- diabetic rat. Diabetes. Res. Clin. Pr. 1998, 40, 145–151. [Google Scholar] [CrossRef]
- Cortez-Navarrete, M.; Martínez-Abundis, E.; Pérez-Rubio, K.G.; González-Ortiz, M.; Villar, M.M.D. Momordica charantia Administration Improves Insulin Secretion in Type 2 Diabetes Mellitus. J. Med. Food. 2018, 21, 672–677. [Google Scholar] [CrossRef]
- Shimada, T.; Kato, F.; Dwijayanti, D.R.; Nagata, T.; Kinoshita, A.; Okuyama, T.; Nishizawa, M.; Mukai, E. Bitter melon fruit extract enhances intracellular ATP production and insulin secretion from rat pancreatic β-cells. Br. J. Nutr. 2022, 127, 377–383. [Google Scholar] [CrossRef]
- Sridhar, M.G.; Vinayagamoorthi, R.; Arul Suyambunathan, V.; Bobby, Z.; Selvaraj, N. Bitter gourd (Momordica charantia) improves insulin sensitivity by increasing skeletal muscle insulin-stimulated IRS-1 tyrosine phosphorylation in high-fat-fed rats. Br. J. Nutr. 2008, 99, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.Y.; Li, C.C.; Ho, T.Y.; Hsiang, C.Y. Identification of the bioactive and consensus peptide motif from Momordica charantia insulin receptor-binding protein. Food Chem. 2016, 204, 298–305. [Google Scholar] [CrossRef]
- Chen, Q.; Chan, L.L.; Li, E.T.S. Bitter melon (Momordica charantia) reduces adiposity, lowers serum insulin and normalizes glucose tolerance in rats fed a high fat diet. J. Nutr. 2003, 133, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Adeghate, E.; Cummings, E.; Singh, J. Beneficial effects and mechanism of action of Momordica charantia juice in the treatment of streptozotocin-induced diabetes mellitus in rat. Mol. Cell. Biochem. 2004, 261, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Cummings, E.; Hundal, H.S.; Wackerhage, H.; Belle, M.; Adeghate, E.; Singh, J. Momordica charantia fruit juice stimulates glucose and amino acid uptakes in l6 myotubes. Mol. Cell. Biochem. 2004, 261, 99–104. [Google Scholar] [CrossRef]
- White, P.E.; Król, E.; Szwengiel, A.; Tubacka, M.; Szczepankiewicz, D.; Staniek, H.; Vincent, J.B.; Krejpcio, Z. Effects of bitter melon and a chromium propionate complex on symptoms of insulin resistance and type 2 diabetes in rat models. Biol. Trace Elem. Res. 2021, 199, 1013–1026. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, X.; Yu, J.; Tan, Y.; Guo, P.; Wu, C.M. The gut microbiota confers the lipid-lowering effect of bitter melon (Momordica charantia L.) in high-fat diet (HFD)-induced hyperlipidemic mice. Biomed. Pharmacother. 2020, 131, 110667. [Google Scholar] [CrossRef]
- Singh, R.; Garcia-Gomez, I.; Gudehithlu, K.P.; Singh, A.K. Bitter melon extract promotes granulation tissue growth and angiogenesis in the diabetic wound. Adv. Skin Wound Care 2017, 30, 16–26. [Google Scholar] [CrossRef]
- Dans, A.M.L.; Villarruz, M.V.C.; Jimeno, C.A.; Javelosa, M.A.U.; Chua, J.; Bautista, R.; Velez, G.G.B. The effect of Momordica charantia capsule preparation on glycemic control in Type 2 Diabetes Mellitus needs further studies. J. Clin. Epidemiol. 2007, 60, 554–559. [Google Scholar] [CrossRef]
- Elekofehinti, O.O.; Oyedokun, V.O.; Iwaloye, O.; Lawal, A.O.; Ejelonu, O.C. Momordica charantia silver nanoparticles modulate SOCS/JAK/STAT and P13K/AKT/PTEN signalling pathways in the kidney of streptozotocin-induced diabetic rats. J. Diabetes Metab. Dis. 2021, 20, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Duddela, S.; Nataraj Sekhar, P.; Padmavati, G.V.; Banerjee, A.K.; Murty, U.S.N. Probing the structure of human glucose transporter 2 and analysis of protein ligand interactions. Med. Chem. Res. 2010, 19, 836–853. [Google Scholar] [CrossRef]
- Guillam, M.T.; Hümmler, E.; Schaerer, E.; Wu, J.Y.; Birnbaum, M.J.; Beermann, F.; Schmidt, A.; Deriaz, N.; Thorens, B. Early diabetes and abnormal postnatal pancreatic islet development in mice lacking glut-2. Nat. Genet. 1997, 17, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Barranco, C. SGLT2 blockers in T2DM. Nat. Rev. Cardiol. 2018, 15, 255. [Google Scholar] [CrossRef] [PubMed]
- Dobbins, R.L.; Greenway, F.L.; Chen, L.; Liu, Y.; Breed, S.L.; Andrews, S.M.; Wald, J.A.; Walker, A.; Smith, C.D. Selective sodium-dependent glucose transporter 1 inhibitors block glucose absorption and impair glucose-dependent insulinotropic peptide release. Am. J. Physiol. -Gastrointest. Liver Physiol. 2015, 308, G946–G954. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, R.S.; Snyder, A.; Rajendran, V.M.; Coon, S. GLUT2 proteins are regulated by the AKT pathway under diabetic conditions in intestinal epithelial cells. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Mahrosh, H.S.; Mehmood, R.; Bukhari, S.A.; Afzal, G.; Arif, R. Investigation of hypoglycemic peptides derived from conserved regions of adMc1 to reveal their antidiabetic activities. BioMed Res. Int. 2021, 2021, 5550180. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, K.; Ahmed, B.; Al-Abbasi, F.A.; Anwar, F.; Verma, A. Deconvoluting the dual hypoglycemic effect of wedelolactone isolated from Wedelia calendulacea: Investigation via experimental validation and molecular docking. RSC Adv. 2018, 8, 18180–18196. [Google Scholar] [CrossRef]
- Musa, E.; Matjila, M.; Levitt., N.S. Kisspeptins and glucose homeostasis in pregnancy: Implications for gestational diabetes mellitus—A review article. Reprod. Sci. 2021, 29, 321–327. [Google Scholar] [CrossRef]
- Cani, P.D.; Everard, A. Talking microbes: When gut bacteria interact with diet and host organs. Mol. Nutr. Food Res. 2016, 60, 58–66. [Google Scholar] [CrossRef]
- Kenny, D.J.; Plichta, D.R.; Shungin, D.; Koppel, N.; Hall, B.; Fu, B.; Vasan, R.S.; Shaw, S.Y.; Vlamakis, H.; Balskus, E.P.; et al. Cholesterol metabolism by uncultured human gut bacteria influences host cholesterol level. Cell Host Microbe 2020, 28, 245–257. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Interactions between gut microbiota and host metabolism predisposing to obesity and diabetes. Annu. Rev. Med. 2012, 62, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.Y.; Lo, H.Y.; Liu, I.C.; Lo, L.C.; Hsiang, C.Y.; Ho, T.Y. A gastro-resistant peptide from Momordica charantia improves diabetic nephropathy in db/db mice via its novel reno-protective and anti-inflammatory activities. Food Funct. 2022, 13, 1822–1833. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Choi, J.M.; Park, S.E.; Rheec, E.J.; Lee, W.Y.; Oh, K.W.; Park, S.W.; Park, C.Y. Preventive effects of bitter melon (Momordica charantia) against insulin resistance and diabetes are associated with the inhibition of NF-κB and JNK pathways in high-fat-fed OLETF rats. J. Nutr. Biochem. 2015, 26, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, M.; Burzynska-Pedziwiatr, I.; Wozniak, L.A.; Bukowiecka-Matusiak, M. Evidence from a Systematic Review and Meta-Analysis Pointing to the Antidiabetic Effect of Polyphenol-Rich Plant Extracts from Gymnema montanum, Momordica charantia and Moringa oleifera. Curr. Issues Mol. Biol. 2022, 44, 699–717. [Google Scholar] [CrossRef]
- Moulik, P.K.; Mtonga, R.; Gill, G. Amputation and mortality in new-onset diabetic foot ulcers stratified by etiology. Diabetes Care 2003, 26, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Ghanassia, E.; Villon, L.; Tdd, J.F.; Boegner, C.; Avignon, A.; Sultan, A. Long-term outcome and disability of diabetic patients hospitalized for diabetic foot ulcers: A 6.5-year follow-up study. Diabetes Care 2008, 31, 1288–1292. [Google Scholar] [CrossRef]
- Larsson, J.; Eneroth, M.; Apelqvist, J.; Stenstrom, A. Sustained reduction in major amputations in diabetic patients: 628 amputations in 461 patients in a defined population over a 20-year period. Acta. Orthop. 2008, 79, 665–673. [Google Scholar] [CrossRef]
- Goyal, R.; Singhai, M. Tuberculosis and non-diabetic hyperglycemia: A challenge to public health management. Med. Hypotheses 2013, 81, 1170–1171. [Google Scholar] [CrossRef]
- Wang, W.T.; Lee, P.; Yeh, H.W.; Smirnova, I.V.; Choi, I.Y. Effects of acute and chronic hyperglycemia on the neurochemical profiles in the rat brain with streptozotocin-induced diabetes detected using in vivo 1 h MR spectroscopy at 9.4 t. J. Neurochem. 2012, 121, 407–417. [Google Scholar] [CrossRef]
- Berlanga-Acosta, J.; Schultz, G.S.; Lopez-Mola, E.; Nieto, G.G.; Siverio, M.G.; Martinez, L.H. Glucose toxic effects on granulation tissue productive cells: The diabetics’ impaired healing. BioMed Res. Int. 2013, 2013, 256043. [Google Scholar] [CrossRef]
- Jia, Y.N.; Xue, Z.H.; Wang, Y.J.; Lu, Y.P.; Li, R.L.; Li, N.N.; Wang, Q.R. Chemical structure and inhibition on α-glucosidase of polysaccharides from corn silk by fractional precipitation. Carbohyd. Polym. 2021, 252, 117185. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, L.; Zhu, J.; Ai, Y.F. Utilization of maltogenic α-amylase treatment to enhance the functional properties and reduce the digestibility of pulse starches. Food Hydrocoll. 2021, 120, 106932. [Google Scholar] [CrossRef]
- Belhadj, S.; Hentati, O.; Elfeki, A.; Hamden, K. Inhibitory activities of Ulva lactuca polysaccharides on digestive enzymes related to diabetes and obesity. Arch. Physiol. Biochem. 2013, 119, 81–87. [Google Scholar]
- Li, Y.X.; Kim, S.K. Utilization of seaweed derived ingredients as potential antioxidants and functional ingredients in the food industry: An overview. Food Sci. Biotechnol. 2011, 20, 1461–1466. [Google Scholar] [CrossRef]
- Blum, A.; Loerz, C.; Martin, H.J.; Staab-Weijnitz, C.A.; Maser, E. Momordica charantia extract, a herbal remedy for type 2 diabetes, contains a specific 11β-hydroxysteroid dehydrogenase type 1 inhibitor. J. Steroid Biochem. Mol. Biol. 2012, 128, 51–55. [Google Scholar] [CrossRef]
- Iseli, T.J.; Oakhill, J.S.; Bailey, M.F.; Wee, S.; Walter, M.; Denderen, B.J.V.; Castelli, L.A.; Katsis, F.; Witters, L.A.; Stapleton, D.; et al. AMP-activated protein kinase subunit interactions. J. Biol. Chem. 2008, 283, 4799–4807. [Google Scholar] [CrossRef]
- Bijland, S.; Mancini, S.J.; Salt, I.P. Role of amp-activated protein kinase in adipose tissue metabolism and inflammation. Clin. Sci. 2013, 124, 491–507. [Google Scholar] [CrossRef]
- Zeng, W.F.; Yin, X.Z.; Jiang, Y.H.; Jin, L.T.; Liang, W. PPARα at the crossroad of metabolic–immune regulation in cancer. FESB J. 2021, 2021, 1–14. [Google Scholar] [CrossRef]
- Liu, Z.; Gong, J.; Huang, W.; Lu, F.; Dong, H. The effect of Momordica charantia in the treatment of diabetes mellitus: A review. Evid-Based Compl. Alt. 2021, 2021, 3796265. [Google Scholar] [CrossRef]
- Pahlavani, N.; Roudi, F.; Zakerian, M.; Ferns, G.A.; Navashenaq, J.G.; Mashkouri, A.; Ghayour-Mobarhan, M.; Rahimi, H. Possible molecular mechanisms of glucose- lowering activities of Momordica charantia (karela) in diabetes. J. Cell. Biochem. 2019, 120, 10921–10929. [Google Scholar] [CrossRef]
- Sur, S.; Steele, R.; Aurora, R.; Varvares, M.; Schwetye, K.E.; Ray, R.B. Bitter Melon prevents the development of 4-NQO-induced oral squamous cell carcinoma in an immunocompetent mouse model by modulating immune signaling. Cancer Prev. Res. 2018, 11, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Peter, E.L.; Kasali, F.M.; Deyno, S.; Mtewa, A.; Nagendrappa, P.B.; Tolo, C.U.; Ogwang, P.E.; Sesaazi, D. Momordica charantia L. lowers elevated glycaemia in type 2 diabetes mellitus patients: Systematic review and meta-analysis. J. Ethnopharmacol. 2019, 231, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Jensen, J.S.; Jensen, T.; Deckert, T. Serum sialic acid concentration is elevated in IDDM especially in early diabetic nephropathy. J. Intern. Med. 2010, 237, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Crook, M.A.; Pickup, J.C.; Lumb, P.J.; Georgino, F.; Webb, D.J.; Fuller, J.H. Relationship between plasma sialic acid concentration and microvascular and macrovascular complications in type 1 diabetes: The eurodiab complications study. Diabetes Care 2001, 24, 316–322. [Google Scholar] [CrossRef]
- Rahman, I.; Malik, S.A.; Bashir, M.; Khan, R.; Iqbal, M. Serum sialic acid changes in non-insulin-dependant diabetes mellitus (NIDDM) patients following bitter melon (Momordica charantia) and rosiglitazone (avandia) treatment. Phytomedicine 2009, 16, 401–405. [Google Scholar] [CrossRef]
- Sukmawati, D.; Fujimura, S.; Jitsukawa, S.; Hirano, R.I.; Ishii, T.; Sato, T.; Hayashi, A.; Itoh, S.; Mizuno, H.; Daida, H.; et al. Oxidative stress tolerance of early stage diabetic endothelial progenitor cell. Regen. Ther. 2015, 1, 38–44. [Google Scholar] [CrossRef][Green Version]
- Kizub, I.V.; Klymenko, K.I.; Soloviev, A.I. Protein kinase C in enhanced vascular tone in diabetes mellitus. Int. J. Cardiol. 2014, 174, 230–242. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, B.; Li, Z.; Zeng, T.; Zhan, J.; Wang, S.; Ho, C.-T.; Li, S. Bioactives of Momordica charantia as Potential Anti-Diabetic/Hypoglycemic Agents. Molecules 2022, 27, 2175. https://doi.org/10.3390/molecules27072175
Xu B, Li Z, Zeng T, Zhan J, Wang S, Ho C-T, Li S. Bioactives of Momordica charantia as Potential Anti-Diabetic/Hypoglycemic Agents. Molecules. 2022; 27(7):2175. https://doi.org/10.3390/molecules27072175
Chicago/Turabian StyleXu, Bilin, Zhiliang Li, Ting Zeng, Jianfeng Zhan, Shuzhen Wang, Chi-Tang Ho, and Shiming Li. 2022. "Bioactives of Momordica charantia as Potential Anti-Diabetic/Hypoglycemic Agents" Molecules 27, no. 7: 2175. https://doi.org/10.3390/molecules27072175
APA StyleXu, B., Li, Z., Zeng, T., Zhan, J., Wang, S., Ho, C.-T., & Li, S. (2022). Bioactives of Momordica charantia as Potential Anti-Diabetic/Hypoglycemic Agents. Molecules, 27(7), 2175. https://doi.org/10.3390/molecules27072175