
A Comparison of Bioactive Metabolites, Antinutrients, and Bioactivities of African Pumpkin Leaves (Momordica balsamina L.) Cooked by Different Culinary Techniques

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Changes in Bioactive Metabolites during Different Household Cooking
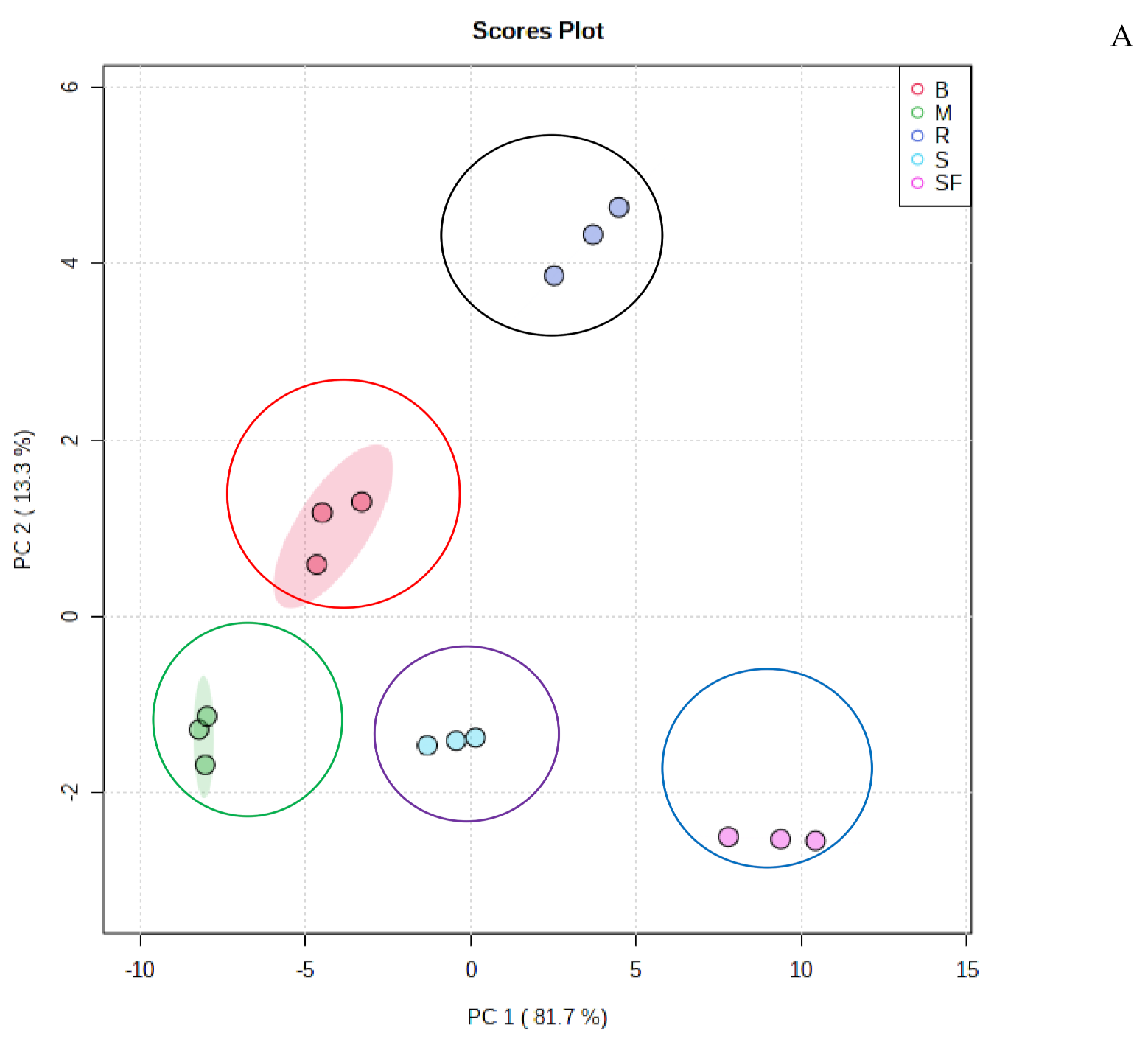
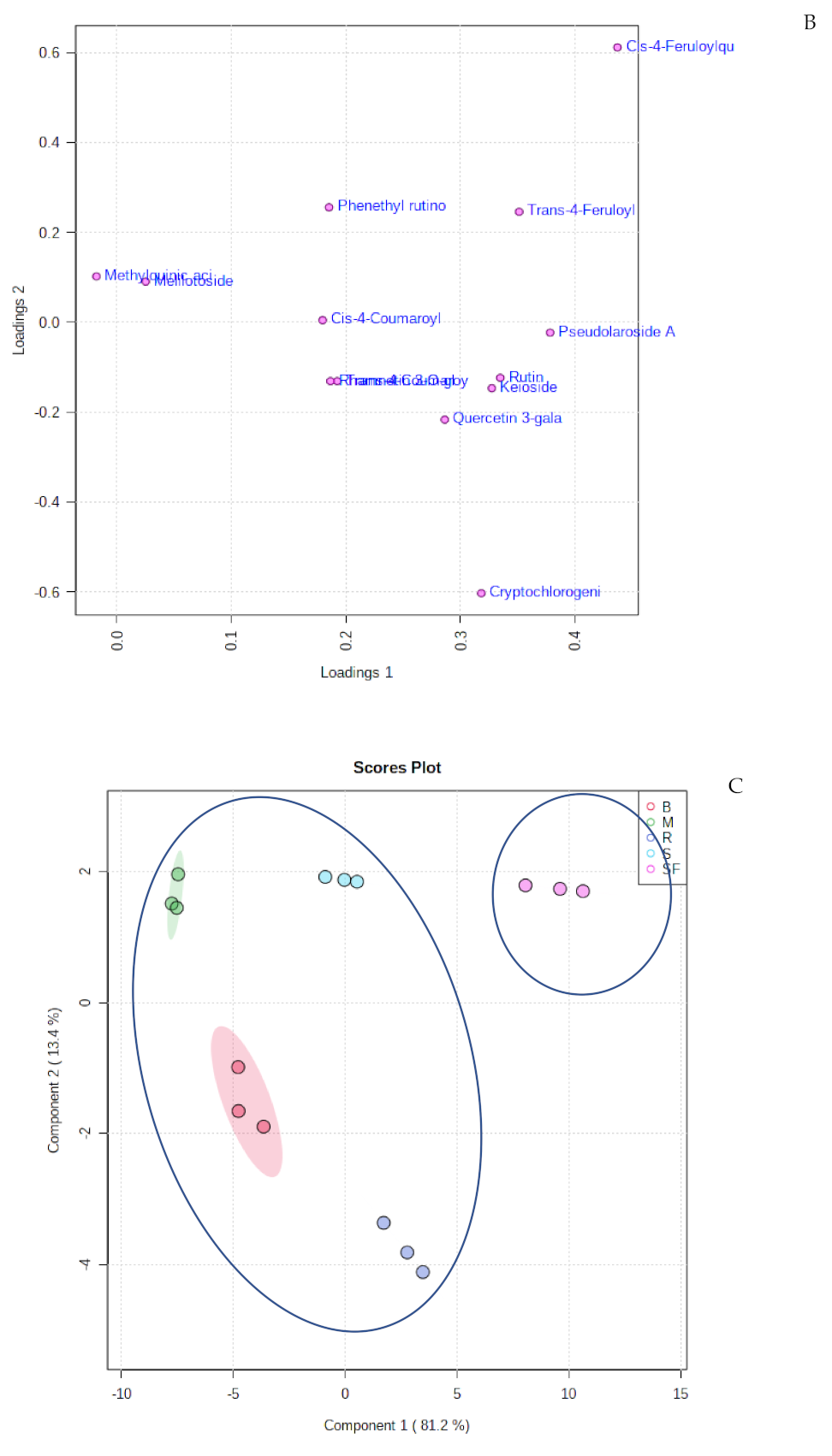
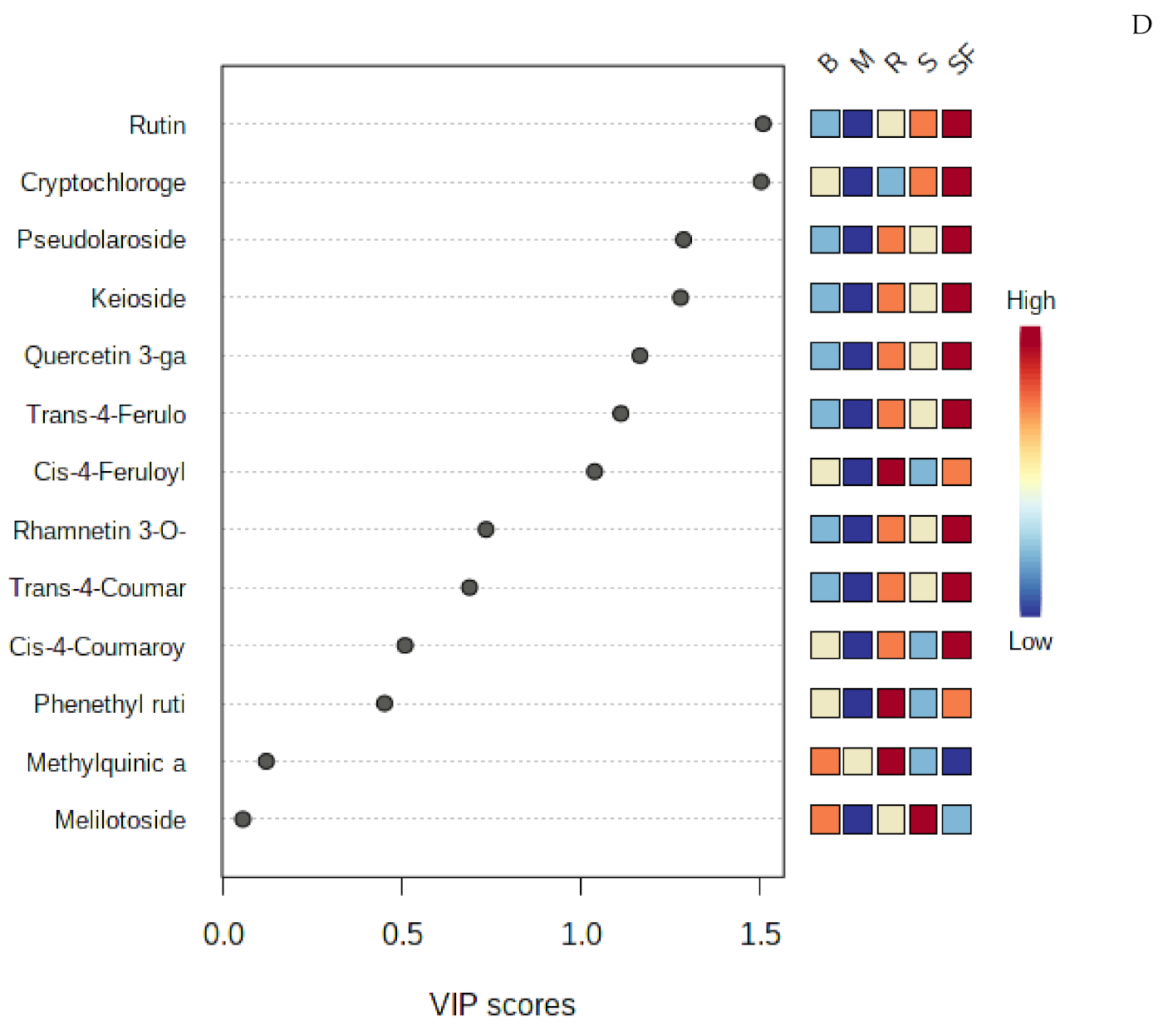
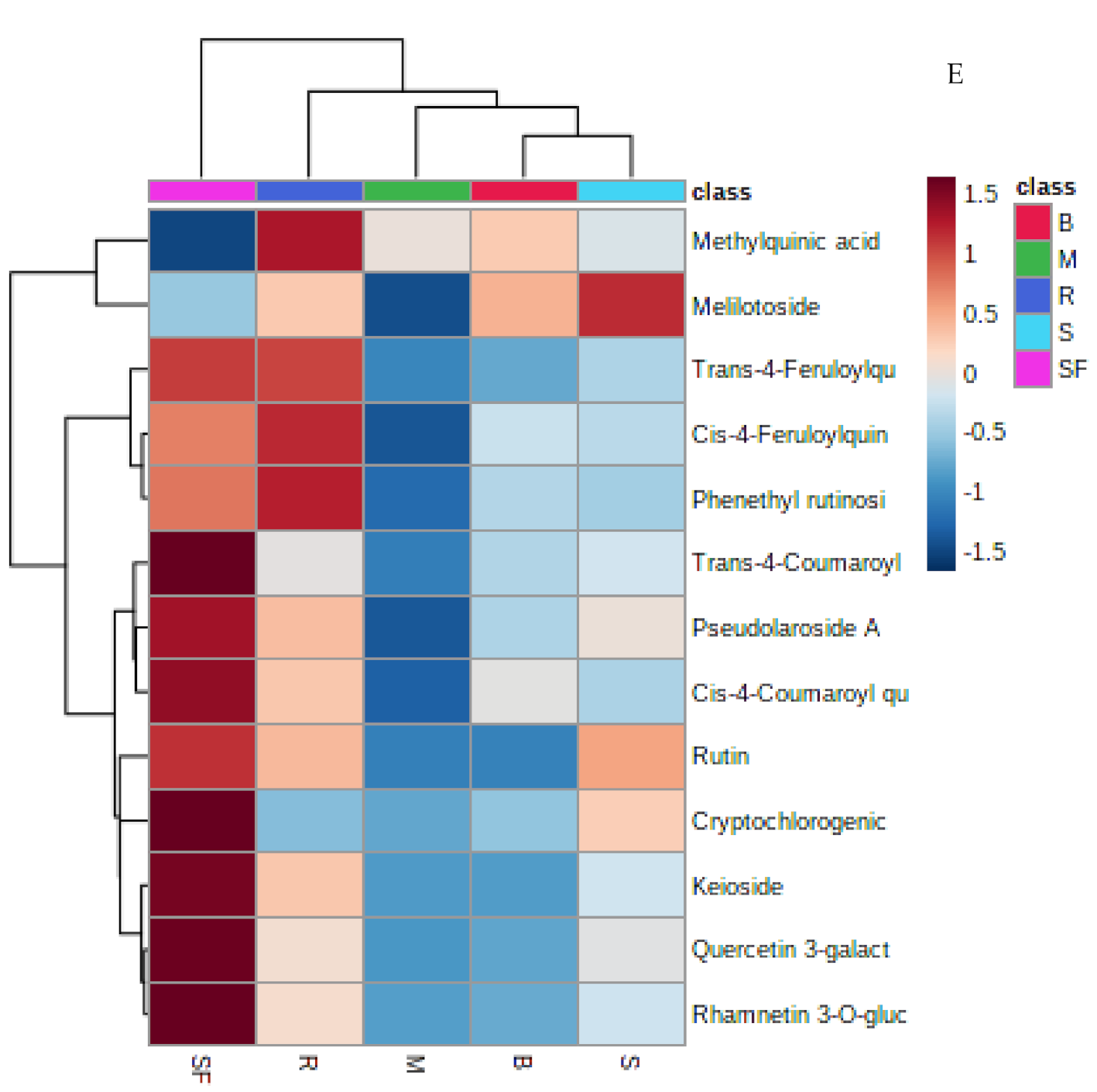
2.2. Multivariate Analysis
2.3. Carotenoid Profile in African Pumpkin Leaves (Momordica balsamina L.)
2.4. Antioxidant Capacity and Antidiabetic Activity
2.5. Antinutritive Compounds
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Household Cooking Techniques
3.4. Quantification of Phenolic Metabolites
3.5. Determination of Carotenoids
3.6. Antioxidant Capacity
3.7. Antidiabetic Activity
3.7.1. α-Glucosidase Inhibitory Activity
3.7.2. α-Amylase Inhibitory Activity
3.8. Antinutritive Compounds
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moyo, M.; Amoo, S.O.; Ncube, B.; Ndhlala, A.R.; Finnie, J.F.; Van Staden, J. Phytochemical and antioxidant properties of conventional leafy vegetables consumed in Southern Africa. S. Afr. J. Bot. 2013, 84, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Uusiku, N.P.; Oelotse, A.; Doudu, K.G.; Bester, M.J.; Faber, M. Nutritional value of leafy vegetables of Sub-Saharan Africa and their potential contribution to human health: A review. J. Food Comp. Anal. 2010, 23, 499–509. [Google Scholar] [CrossRef]
- Mshelia, H.S.; Karumi, Y.; Dibal, N.I. Therapeutic effect of Momordica balsamina L. leafy extract on ethanol-induced gastric ulcer in Wistar rats. Ann. Res. Hosp. 2017, 1, 1–5. [Google Scholar] [CrossRef]
- Siboto, A.; Sibiya, N.; Khathi, A.; Ngubane, P. The effect of Momordica balsamina L. methanolic extract on kidney function in STZ-induced diabetic rats: Effects on selected metabolic markers. J. Diabetes Res. 2018, 2018, 7341242. [Google Scholar] [CrossRef] [Green Version]
- Akdaş, Z.Z.; Bakkalbaşi, E. Influence of different cooking methods on color, bioactive compounds, and antioxidant activity of kale. Int. J. Food Prop. 2017, 20, 877–887. [Google Scholar] [CrossRef]
- Thakur, G.S.; Bag, M.; Sanodiya, B.S.; Bhadauriya, P.; Debnath, M.; Prasad, G.B.K.S.; Bisen, P.S. Momordica balsamina: A medicinal and nutraceutical plant for health care management. Curr. Pharm. Biotechnol. 2009, 10, 667–682. [Google Scholar] [CrossRef]
- USDA. Item Clusters, Percent of Consumption, and Representative Foods for Typical Choices Food Patterns. 2020. Available online: https://www.fns.usda.gov/usda-food-patterns (accessed on 15 June 2021).
- Bvenura, C.; Afolayon, A.J. The role of wild vegetables in the household food security in South Africa: A review. Food Res. Int. 2015, 76, 1001–1011. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Report on Crop Diversification for Sustainable Diets and Nutrition. In Crop Diversification for Sustainable Diets and Nutrition—The Role of FAO’s Plant Production and Protection Division; FAO: Rome, Italy, 2012; Available online: https://www.fao.org/ag/agp/greenercities/pdf/CDSDN.pdf (accessed on 15 June 2021).
- Mncwango, N.C.; Mavengahama, S.; Ntuli, N.R.; Van Jaarsveld, C.M. Diversity, consumption dynamics and ethnomedicinal claims of traditional leafy vegetables consumed by a rural community in the KwaMbonambi area, Northern KwaZulu-Natal, South Africa. Biodiversitas 2020, 21, 1201–1207. [Google Scholar] [CrossRef]
- Oboh, G. Effect of blanching on the antioxidant properties of some tropical green leafy vegetables. LWT Food Sci. Technol. 2005, 38, 513. [Google Scholar] [CrossRef]
- Managa, M.G.; Shai, J.; Phan, A.D.T.; Sultanbawa, Y.; Sivakumar, D. Impact of household cooking techniques on African nightshade and Chinese cabbage on phenolic compounds, antinutrients, in vitro antioxidants and β-glucosidase activity. Front. Nutr. 2020, 7, 550. [Google Scholar] [CrossRef]
- Moloto, R.M.; Phan, A.D.T.; Shai, J.L.; Sultanbawa, Y.; Sivakumar, D. Comparison of phenolic compounds, carotenoids, amino acid composition, in vitro antioxidants and anti-diabetic activities in the leaves of seven Cowpea (Vigna unguiculata) cultivars. Foods 2020, 9, 1285. [Google Scholar] [CrossRef]
- Mashiane, P.; Mashitoa, F.M.; Slabbert, R.M.; Sivakumar, D. Impact of household cooking techniques on colour, antioxidant and sensory properties of African pumpkin and pumpkin leaves. Int. J. Gastron. Food Sci. 2021, 23, 100307. [Google Scholar] [CrossRef]
- Subramaniam, S.; Rosdi, M.H.B.; Kuppusamy, U.R. Customized cooking methods enhance antioxidant, antiglycemic, and insulin-like properties of Momordica charantia and Moringa oleifera. J. Food Qual. 2017, 2017, 9561325. [Google Scholar] [CrossRef] [Green Version]
- Mashiane, P.; Manhivi, V.; Shoko, T.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. Cooking African pumpkin leaves (Momordica balsamina L.) by stir-frying improved bioactivity and bioaccessibility of metabolites—Metabolomic and chemometric approaches. Foods 2021, 10, 2890. [Google Scholar] [CrossRef]
- Essack, H.; Odhav, B.; Mellem, J.J. Screening of traditional South African leafy vegetables for specific anti-nutritional factors before and after processing. Food Sci. Technol. 2017, 37, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Agbaire, P.O. Levels of anti-nutritional factors in some common leafy edible vegetables of southern Nigeria. Afr. J. Food Sci. Technol. 2012, 3, 99–101. [Google Scholar]
- Mashitoa, F.M.; Shoko, T.; Shai, J.L.; Slabbert, R.M.; Sivakumar, D. Changes in phenolic metabolites and biological activities of Pumpkin leaves (Cucurbita moschata Duchesne ex Poir.) during blanching. Front. Nutr. 2021, 8, 1939. [Google Scholar] [CrossRef]
- Park, M.H.; Kim, D.Y.; Jang, H.J.; Jo, Y.H.; Jeong, J.T.; Lee, D.Y.; Baek, N.I.; Ryu, H.W.; Oh, S.R. Analysis of polyphenolic metabolites from Artemisia melinii Weber ex Stechm and regional comparison in Korea. J. Appl. Biol. Chem. 2019, 62, 433–439. [Google Scholar] [CrossRef] [Green Version]
- Fang, N.; Yu, S.; Prior, R.L. LC/MS/MS characterization of phenolic constituents in dried plums. J. Agric. Food Chem. 2002, 50, 3579–3585. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MS identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Ramabulana, A.T.; Steenkamp, P.; Madala, N.; Dubery, I.A. Profiling of chlorogenic acids from Bidens pilosa and differentiation of closely related positional isomers with the aid of UHPLC-QTOF-MS/MS-based in-source collision-induced dissociation. Metabolites 2020, 10, 178. [Google Scholar] [CrossRef]
- Shoko, T.; Maharaj, V.; Naidoo, D.; Tselanyane, M.; Nthambeleni, R.; Khorombi, E. Anti-aging potential of extracts from Sclrocarya birrea (A. rich) Hochst and its chemical profiling by UPLC-Q-TOF-MS. BMC Complement. Altern. Med. 2018, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Olennikov, D.N.; Chirikova, N.K.; Kashchenko, N.I.; Nikolaev, V.M.; Kim, S.W.; Vennos, C. Bioactive phenolics of the genus Artemisia (Asteraceae): HPLC-DAD-ESI-TQ-MS/MS profile of the Siberian species and their inhibitory potential against α-amylase and α-glucosidase. Front. Pharmacol. 2018, 9, 756. [Google Scholar] [CrossRef]
- Silva, L.M.P.; Alves, J.S.F.; da Silva Siqueira, E.M.; de Souza Neto, M.A.; Abreu, L.S.; Tavares, J.F.; Porto, D.L.; de Santis Ferreira, L.; Demarque, D.P.; Lopes, N.P. Isolation and identification of the five novel flavonoids from Genipa americana leaves. Molecules 2018, 23, 2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berardini, N.; Fezer, R.; Conrad, J.; Beifuss, U.; Carle, R.; Schieber, A. Screening of mango (Mangifera indica L.) cultivars for their contents of flavonol O-and xanthone C-glycosides, anthocyanins, and pectin. J. Agric. Food Chem. 2005, 53, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Sergio, L.; Boari, F.; Pieralice, M.; Linsalata, V.; Cantore, V.; Di Venere, D. Bioactive phenolics and antioxidant capacity of some wild edible greens as affected by different cooking treatments. Foods 2020, 9, 1320. [Google Scholar] [CrossRef] [PubMed]
- Ferracane, R.; Pellegrini, N.; Visconti, A.; Graziani, G.; Chiavaro, E.; Miglio, C.; Fogliano, V. Effects of different cooking methods on antioxidant profile, antioxidant capacity, and physical characteristics of artichoke. J. Agric. Food Chem. 2008, 56, 8601–8608. [Google Scholar] [CrossRef] [PubMed]
- Managa, M.G.; Mpai, S.; Remize, F.; Garcia, C.; Sivakumar, D. Impact of moist cooking methods on colour, anti-nutritive compounds and phenolic metabolites in African nightshade (Solanum retroflexum Dun.). Food Chem. 2020, 325, 126805. [Google Scholar] [CrossRef]
- Wang, Y.; Ho, C.T. Polyphenolic chemistry of tea and coffee: A century of progress. J. Agric. Food Chem. 2009, 7, 8109–8114. [Google Scholar] [CrossRef]
- Zeb, A.; Haq, A.; Murkovic, M. Effects of microwave cooking on carotenoids, phenolic compounds and antioxidant activity of Cichorium intybus L. (chicory) leaves. Eur. Food Res. Technol. 2019, 245, 365–374. [Google Scholar] [CrossRef]
- Cheng, X.; Hanschen, F.S.; Neugart, S.; Scheiner, M.; Vargas, S.A.; Gutschmann, B.; Baldermann, S. Boiling and steaming induced changes in the secondary metabolites in three different cultivars of Pokchoi (Brassica rapa subsp. Chinensis). J. Food Comp. Anal. 2019, 82, 103232. [Google Scholar] [CrossRef]
- Fransisco, M.; Velasco, P.; Moreno, D.A.; Garcia-Viguera, C.; Cartea, M.E. Cooking methods of Brassica rapa affect the preservation of Glucosinolates, phenolics and vitamin C. Food Res. Int. 2010, 43, 1455–1463. [Google Scholar] [CrossRef] [Green Version]
- Fiol, M.; Weckmüller, A.; Neugart, S.; Schreiner, M.; Rohn, S.; Krumbein, A.; Kroh, L.W. Thermal-induced changes of kale’s antioxidant activity analyzed by HPLC–UV/Vis-online-TEAC detection. Food Chem. 2013, 138, 857–865. [Google Scholar] [CrossRef]
- Odongo, G.A.; Schlotz, N.; Herz, C.; Hanschen, F.S.; Baldermann, S.; Neugart, S.; Trierweiler, B.; Frommherz, L.; Franz, C.M.A.P.; Ngwene, B.; et al. The role of plant processing for the cancer preventive potential of Ethiopian kale (Brassica carinata). Food Nutr. Res. 2017, 61, 1271527. [Google Scholar] [CrossRef] [Green Version]
- Miglio, C.; Chiavaro, E.; Visconti, A.; Fogliano, V.; Pellegrini, N. Effects of different cookingmethods on nutritional and physicochemical characteristics of selected vegetables. J. Agric. Food Chem. 2008, 56, 139–147. [Google Scholar] [CrossRef]
- Gunathilake, K.D.; Ranaweera, K.K.D.; Rupasinghe, H.P. Effect of different cooking methods on polyphenols, carotenoids and antioxidant activities of selected edible leaves. Antioxidants 2018, 7, 117. [Google Scholar] [CrossRef] [Green Version]
- Hossain, A.; Khatun, M.A.; Islam, M.; Huque, R. Enhancement of antioxidant quality of green leafy vegetables upon different cooking method. Prev. Nutr. Food Sci. 2017, 22, 216. [Google Scholar] [CrossRef]
- Carvalho, F.V.; Santana, L.F.; Diogenes, V.; da Silva, A.; Costa, L.; Zambotti-Villelae, L.; Colepicolo, P.; Ferraz, C.P.; Ribeiro, P.R. Combination of a multiplatform metabolite profiling approach and chemometrics as a powerful strategy to identify bioactive metabolites in Lepidium meyenii (Peruvian maca). Food Chem. 2021, 364, 130453. [Google Scholar] [CrossRef]
- Gehlenborg, N.; Wong, B. Heat maps. Nat. Methods 2012, 9, 213. [Google Scholar] [CrossRef]
- Zib, A.; Nisar, A. Effects of high temperature frying of spinach leaves in sunflower oil on carotenoids, chlorophylls, and tocopherol composition. Front. Chem. 2017, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Rodriguez, A.; Fransisco, R.M.; Ocana, A.; Soler-Rivas, C. Effect of domestic processing on bioactive compounds. Phytochem. Rev. 2008, 7, 345–384. [Google Scholar] [CrossRef]
- Pellegrini, N.; Chiavaro, E.; Gardana, C.; Mazzeo, T.; Contino, D.; Gallo, M.; Riso, P.; Fagliano, V.; Porrini, M. Effect of Different Cooking methods on color, phytochemical, concentration, and antioxidant capacity of raw and frozen brassica vegetables. J. Agric. Food Chem. 2010, 58, 4310–4321. [Google Scholar] [CrossRef]
- Popova, M.; Lyoussi, B.; Aazza, S.; Antunes, D.; Bankova, V.; Miguel, G. Antioxidant and α-gluocosidase inhibitory properties and chemical profiles of Moroccan propolis. Nat. Prod. Commun. 2015, 10, 1961–1964. [Google Scholar]
- Mashitoa, F.M.; Manhivi, V.; Slabbert, R.M.; Shai, J.L.; Sivakumar, D. Changes in antinutrients, phenolics, antioxidant activities and in vitro glucosidase inhibitory activity in pumpkin leaves (Cucurbita moschata) during different domestic cooking methods. Food Sci. Biotechnol. 2021, 30, 793–800. [Google Scholar] [CrossRef]
- Amalraj, A.; Pius, A. Bioavilability of calcium and its absorption inhibitors in raw and cooked green leafy vegetables commonly consumed in India: An in vitro study. Food Chem. 2015, 170, 430–436. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, Y.; Lai, S.; Cao, H.; Guan, Y.; San Cheang, W.; Liu, B.; Zhao, K.; Miao, S.; Riviere, C.; et al. Effects of domestic cooking process on chemical and biological properties of dietary phytochemicals. Trends Food Sci. Technol. 2019, 85, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Sarian, M.N.; Ahmed, Q.U.; Mat So’ad, S.Z.; Alhassan, A.M.; Murugesu, S.; Perumal, V.; Mohamad, S.N.A.S.; Khatib, A.; Latip, J. Antioxidant and antidiabetic effects of flavonoids: A structure-activity relationship based study. Biomed. Res. Int. 2017, 2017, 8386065. [Google Scholar] [CrossRef]
- Yadav, S.K.; Sehgal, S. Effect of domestic processing and cooking methods on total, HCl extractable iron and in vitro availability of iron in spinach and amaranth leaves. Plant Foods Hum. Nutr. 2002, 58, 1–11. [Google Scholar] [CrossRef]
- Panfili, G.; Frantiani, A.; Irano, M. Improved normal-phase high performance liquid chromatography procedure for the determination of carotenoids in cereals. J. Agric. Food Chem. 2004, 52, 6373–6380. [Google Scholar] [CrossRef] [PubMed]
- Seke, F.; Manhivi, V.E.; Shoko, T.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. Effect of freeze drying and simulated gastrointestinal digestion of phenolic metabolites and antioxidant property of Natal plum (Carissa macrocarpa). Foods 2021, 10, 1420. [Google Scholar] [CrossRef] [PubMed]
- Price, M.L.; Van Scovoc, S.; Butler, G. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites (mg kg−1) | Raw Leaves | Steaming | Microwaving | Boiling | Stir-Frying |
|---|---|---|---|---|---|
| Methylquinic acid | 3.96 ± 0.71 *a | 1.44 ± 0.26 c | 2.53 ± 3.29 b | 2.08 ± 0.99 b | 0.33 ± 0.06 d |
| 4-caffeoylquinicacid (cryptochlorogenic acid) | 6.61 ± 1.18 d | 76.01 ± 13.57 b | 3.34 ± 2.13 e | 8.35 ± 1.36 c | 507.02 ± 90.54 a |
| cis-4-coumaroylquinic acid | 33.04 ± 1.90 b | 12.17 ± 2.17 d | 1.47 ± 0.44 e | 20.82 ± 5.81 c | 99.83 ± 17.83 a |
| trans-4-coumaroylquinic acid | 17.50 ± 3.12 b | 13.61 ± 2.43 c | 1.57 ± 0.33 e | 10.34 ± 5.60 d | 116.57 ± 20.82 a |
| cis-4-feruloylquinic acid | 1505.20 ± 2.7 a | 261.19 ± 46.6 d | 21.87 ± 2.98 e | 299.05 ± 74.2 c | 983.1 ± 175.56 b |
| Trans-4-feruloylquinic acid | 244.86 ± 4.72 a | 19.54 ± 3.49 b | 1.38 ± 0.71 d | 6.06 ± 4.31 c | 259.99 ± 46.43 a |
| Quercetin-3-galactoside | 22.26 ± 3.97 b | 15.59 ± 2.78 c | 0.62 ± 0.32 e | 1.33 ± 0.92 d | 206.06 ± 36.80 a |
| Phenethyl rutinoside | 95.73 ± 17.10 a | 10.04 ± 1.79 c | 1.13 ± 0.28 d | 11.96 ± 1.52 c | 60.90 ± 10.87 b |
| Rhamnetin-3-O-glucoside | 5.76 ± 1.03 b | 2.56 ± 0.46 c | 0.17 ± 0.18 d | 0.27 ± 0.10 d | 55.43 ± 9.90 a |
| Quercetin-3-rutinoside (rutin) | 175.24 ± 3.29 c | 209.68 ± 37.4 b | 7.94 ± 2.41 d | 8.15 ± 1.49 d | 405.15 ± 72.35 a |
| Pseudolaroside A | 246.97 ± 1.10 b | 155.12 ± 27.7 c | 5.45 ± 0.62 e | 81.31 ± 24.09 d | 678.89 ± 121.2 a |
| β-d-glucosyl-2-coumarate (melilotoside) | 15.12 ± 1.70 c | 31.71 ± 5.66 a | 1.34 ± 0.54 e | 17.37 ± 1.60 b | 6.10 ± 1.09 d |
| Kaempferol-O-rutinoside (nicotiflorin) | 231.27 ± 1.30 b | 50.76 ± 9.07 c | 0.94 ± 0.69 e | 1.80 ± 0.40 d | 425.81 ± 76.04 a |
| Isorhamnetin-3-O-robinoside (keioside) | 41.76 ± 1.46 b | 12.49 ± 2.23 c | 0.52 ± 0.42 d | 0.61 ± 0.33 d | 249.26 ± 44.51 a |
| Household Cooking Methods | Lutein mg 100 g−1 | % Loss or Gain | Zeaxanthin mg 100 g−1 | % Loss or Gain | β-Carotene mg 100 g−1 | % Loss or Gain | Total Carotenoids |
|---|---|---|---|---|---|---|---|
| Raw | 34.13 ± 0.45 *c | 1.30 ± 0.20 bc | 9.05 ± 0.01 d | 44.48 d | |||
| Stir-frying | 54.69 ± 1.0 a | 60.24 ± 0.53 a | 3.20 ± 0.29 a | 146.15 ± 0.62 a | 20.68 ± 0.10 a | 123.51 ± 0.70 a | 78.57 a |
| Boiling | 43.08 ± 1.8 b | 26.22 ± 0.20 c | 2.25 ± 0.25 ab | 73.08 ± 0.75 b | 12.41 ± 0.08c | 37.13±0.52 c | 57.74 c |
| Steaming | 44.51 ± 1.1 b | 30.41 ± 0.60 b | 1.85 ± 1.24 b | 42.31 ± 0.64 d | 17.33 ± 0.03 b | 91.49 ± 0.43 b | 63.69 bc |
| Microwave | 41.71 ± 1.2 b | 22.21 ± 0.33 c | 0.64 ± 0.20 c | 50.77 ± 0.54 c | 7.97 ± 0.03 e | 11.93 ± 0.54 d | 50.32 cd |
| Household Cooking Techniques | DPPH IC50 mg mL−1 | ABTS IC50 mg mL−1 | α-Glucosidase IC50 mg mL−1 | α-Amylase IC50 mg mL−1 |
|---|---|---|---|---|
| Raw | 1.78 ± 0.10 *d | 0.78 ± 0.02 d | 2.23 ± 0.20 d | 1.31 ± 0.21 d |
| Stir-frying | 0.71 ± 0.20 e | 0.61 ± 0.04 e | 1.35 ± 0.09 e | 0.80 ± 0.16 e |
| Microwave | 4.16 ± 0.08 a | 3.52 ± 0.25 a | 5.97 ± 0.79 a | 4.64 ± 0.20 a |
| Steaming | 2.62 ± 0.11 c | 1.21 ± 0.03 c | 3.28 ± 0.28 c | 2.58 ± 0.25 c |
| Boiling | 3.82 ± 0.25 b | 2.62 ± 0.11 b | 4.14 ± 0.00 b | 3.81 ± 0.11 b |
| Acarbose | 6.87 ± 0.22 | 3.14 ± 0.13 |
| Household Cooking Techniques | Tannins mg 100 g−1 | % Loss | Phytates mg 100 g−1 | % Loss | Alkaloids mg 100 g−1 | % Loss |
|---|---|---|---|---|---|---|
| Raw | 165.20±1.68 *a | 41.37 ± 0.36 a | 35.00 ± 0.05 a | |||
| Stir fry | 157.31 ± 0.45 b | 5.88 ±0.52 c | 33.00 ± 1.40 b | 20.23 ± 0.18 c | 23.60 ± 0.29 b | 32.57 ± 0.53 d |
| Boiling | 56.43 ± 1.26 e | 66.25 ± 0.17 a | 18.41 ± 0.53 d | 55.49 ± 0.28 a | 4.04 ± 0.08 e | 88.46 ± 0.26 a |
| Steaming | 131.85 ± 0.99 c | 21.14 ± 0.17 b | 28.75 ± 1.03 c | 30.51 ± 0.39 b | 17.73 ± 0.28 c | 49.34 ± 0.47 c |
| Microwave | 63.48 ± 1.18 d | 62.03 ± 0.08 a | 20.15 ± 0.38 d | 51.29 ± 0.44 a | 14.88 ± 0.29 d | 57.49 ± 0.45 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mashiane, P.; Shoko, T.; Manhivi, V.; Slabbert, R.; Sultanbawa, Y.; Sivakumar, D. A Comparison of Bioactive Metabolites, Antinutrients, and Bioactivities of African Pumpkin Leaves (Momordica balsamina L.) Cooked by Different Culinary Techniques. Molecules 2022, 27, 1901. https://doi.org/10.3390/molecules27061901
Mashiane P, Shoko T, Manhivi V, Slabbert R, Sultanbawa Y, Sivakumar D. A Comparison of Bioactive Metabolites, Antinutrients, and Bioactivities of African Pumpkin Leaves (Momordica balsamina L.) Cooked by Different Culinary Techniques. Molecules. 2022; 27(6):1901. https://doi.org/10.3390/molecules27061901
Chicago/Turabian StyleMashiane, Petunia, Tinotenda Shoko, Vimbainashe Manhivi, Retha Slabbert, Yasmina Sultanbawa, and Dharini Sivakumar. 2022. "A Comparison of Bioactive Metabolites, Antinutrients, and Bioactivities of African Pumpkin Leaves (Momordica balsamina L.) Cooked by Different Culinary Techniques" Molecules 27, no. 6: 1901. https://doi.org/10.3390/molecules27061901
APA StyleMashiane, P., Shoko, T., Manhivi, V., Slabbert, R., Sultanbawa, Y., & Sivakumar, D. (2022). A Comparison of Bioactive Metabolites, Antinutrients, and Bioactivities of African Pumpkin Leaves (Momordica balsamina L.) Cooked by Different Culinary Techniques. Molecules, 27(6), 1901. https://doi.org/10.3390/molecules27061901