
Potential of the Compounds from Bixa orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols

 , ,
, ,  ,
, 
Abstract
:
1. Introduction
2. Results and Discussion
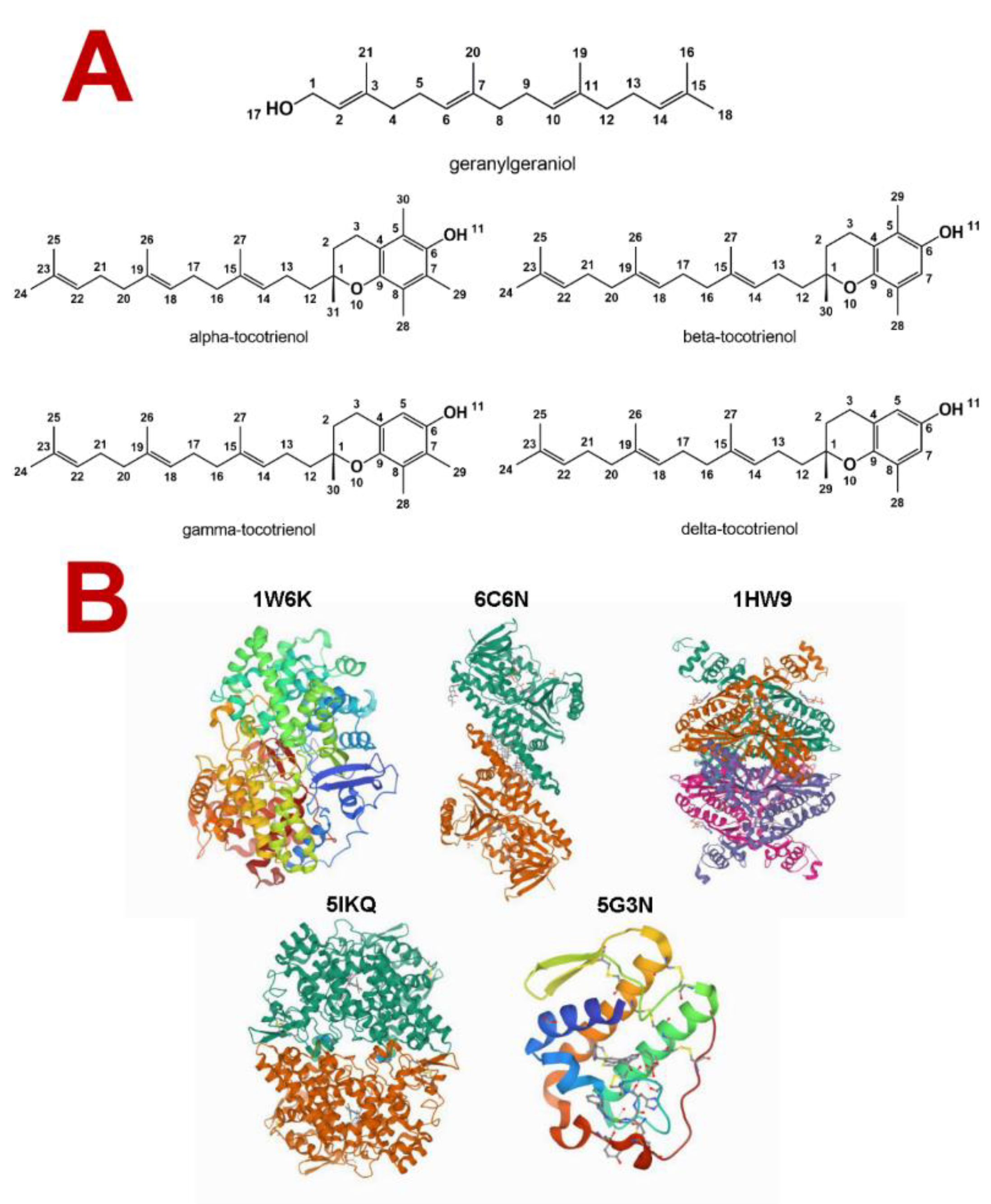
2.1. Molecules’ Structure Obtention and Biological Activity Prediction
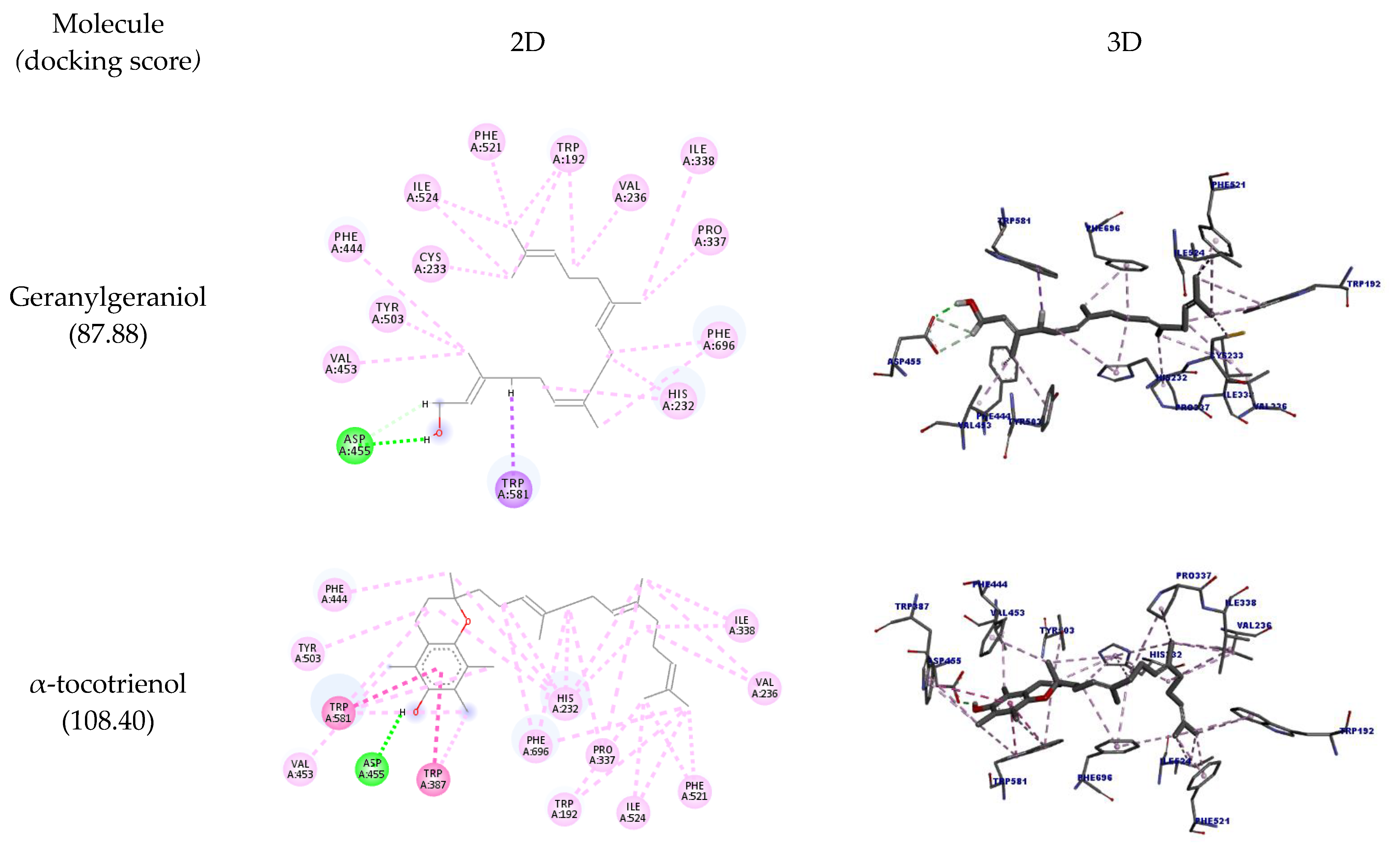
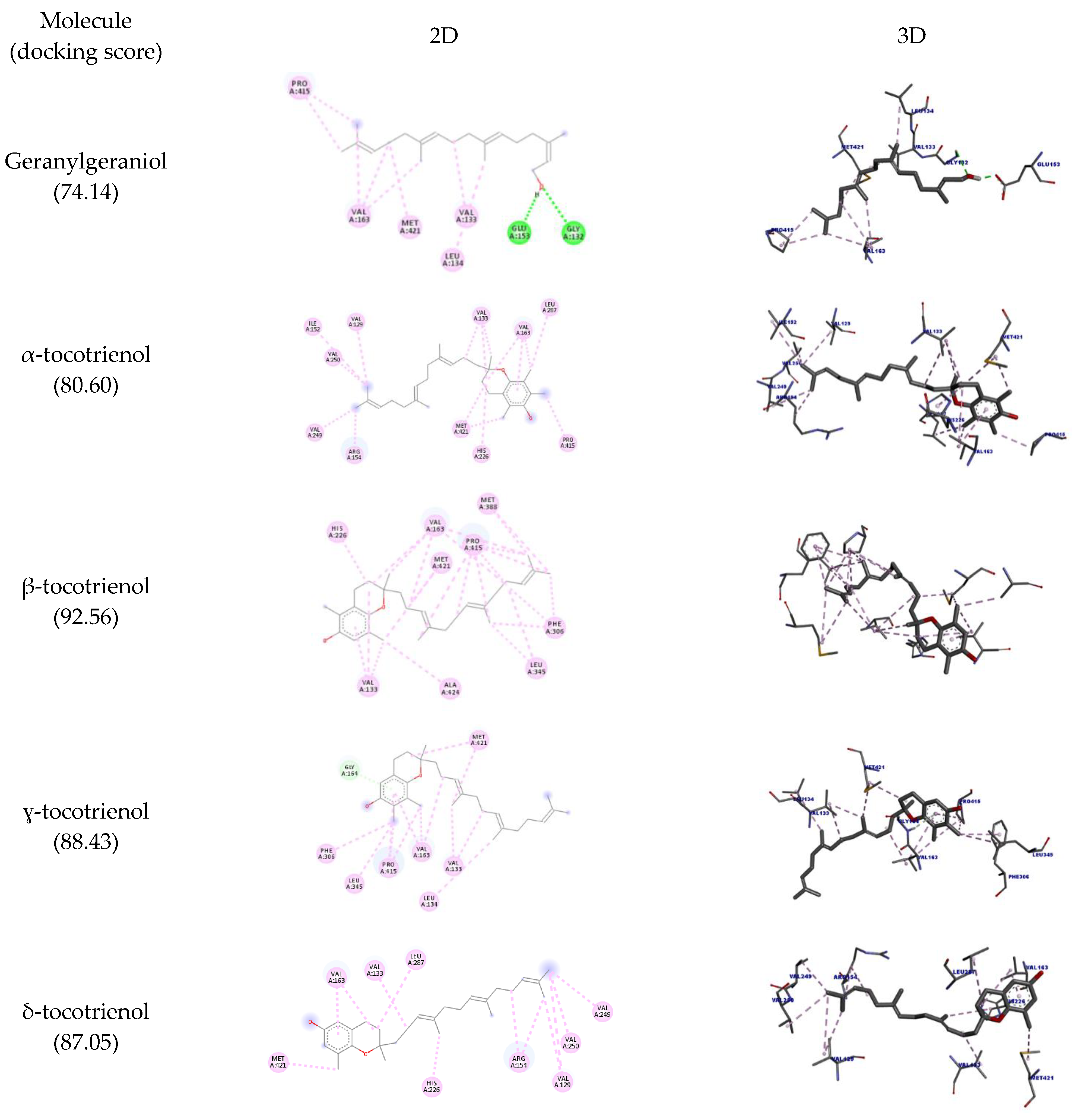
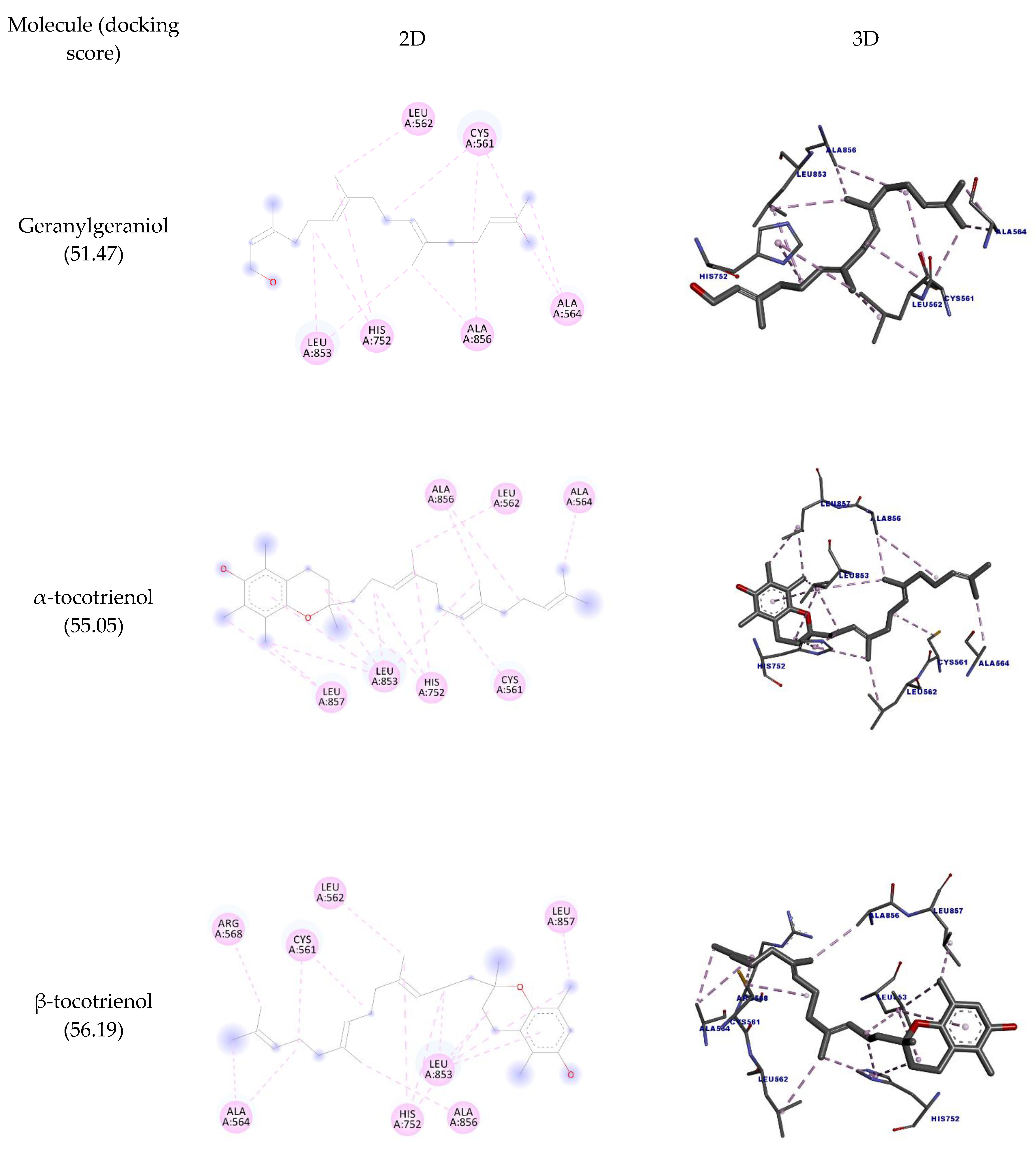
2.2. Molecular Docking
2.3. Pharmacokinetic Property Prediction
2.4. Toxicological Property Prediction
3. Materials and Methods
3.1. Molecules Studied
3.2. Biological Activities Prediction
3.3. Molecular Docking
3.4. Pharmacokinetic Prediction
3.5. Toxicological Prediction
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.S.; Li, J.; Wang, Z.; Mi, C.; Ma, J.; Piao, L.X.; Xu, G.H.; Li, X.; Jin, X. Artemisinin inhibits inflammatory response via regulating NF-κB and MAPK signaling pathways. Immunopharmacol. Immunotoxicol. 2017, 39, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Jing, L.; Tian, Y.; Yan, H.; Zhang, B.; Sun, Q.; Dai, D.; Shi, L.; Liu, D.; Yang, Z.; et al. Epidemiology of dyslipidemia and associated cardiovascular risk factors in northeast China: A cross-sectional study. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 2262–2270. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.; Zhu, X.; Zhang, Y.; Shen, Y. Metabolomic characterization of hypertension and dyslipidemia. Metabolomics 2018, 14, 117. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. Exploring the potential of tocotrienol from Bixa orellana as a single agent targeting metabolic syndrome and bone loss. Bone 2018, 116, 8–21. [Google Scholar] [CrossRef]
- Pacheco, S.D.G.; Gasparin, A.T.; Jesus, C.H.A.; Sotomaior, B.B.; Ventura, A.C.S.S.B.; Redivo, D.D.B.; Cabrini, D.D.A.; Gaspari Dias, J.D.F.; Miguel, M.D.; Miguel, O.G.; et al. Antinociceptive and Anti-Inflammatory Effects of Bixin, a Carotenoid Extracted from the Seeds of Bixa orellana. Planta Med. 2019, 85, 1216–1224. [Google Scholar] [CrossRef]
- Rivera-Madrid, R.; Aguilar-Espinosa, M.; Cárdenas-Conejo, Y.; Garza-Caligaris, L.E. Carotenoid derivates in achiote (Bixa orellana) seeds: Synthesis and health promoting properties. Front. Plant Sci. 2016, 7, 1406. [Google Scholar] [CrossRef] [Green Version]
- Matias Pereira, A.C.; de Oliveira Carvalho, H.; Gonçalves, D.E.S.; Picanço, K.R.T.; de Lima Teixeira dos Santos, A.V.T.; da Silva, H.R.; Braga, F.S.; Bezerra, R.M.; de Sousa Nunes, A.; Nazima, M.T.S.T.; et al. Co-treatment of purified annatto oil (Bixa orellana l.) and its granules (chronic®) improves the blood lipid profile and bone protective effects of testosterone in the orchiectomy-induced osteoporosis in wistar rats. Molecules 2021, 26, 4720. [Google Scholar] [CrossRef]
- Kamal-Eldin, A.; Appelqvist, L.Å. The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids 1996, 31, 671–701. [Google Scholar] [CrossRef]
- Medvedev, O.; Ivanova, A.; Medvedeva, N. Biological properties of tocotrienols. Vopr. Pitan. 2018, 87, 5–16. [Google Scholar] [CrossRef]
- Irwin, J.C.; Fenning, A.S.; Vella, R.K. Geranylgeraniol prevents statin-induced skeletal muscle fatigue without causing adverse effects in cardiac or vascular smooth muscle performance. Transl. Res. 2020, 215, 17–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCully, K.S. Chemical Pathology of Homocysteine VIII. Effects of Tocotrienol, Geranylgeraniol, and Squalene on Thioretinaco Ozonide, Mitochondrial Permeability, and Oxidative Phosphorylation in Arteriosclerosis, Cancer, Neurodegeneration and Aging. Ann. Clin. Lab. Sci. 2020, 50, 567–577. [Google Scholar]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef]
- Bolchi, C.; Bavo, F.; Appiani, R.; Roda, G.; Pallavicini, M. 1,4-Benzodioxane, an evergreen, versatile scaffold in medicinal chemistry: A review of its recent applications in drug design. Eur. J. Med. Chem. 2020, 200, 112419. [Google Scholar] [CrossRef]
- Song, T.-Q.; Ding, M.-Z.; Zhai, F.; Liu, D.; Liu, H.; Xiao, W.-H.; Yuan, Y.-J. Engineering Saccharomyces cerevisiae for geranylgeraniol overproduction by combinatorial design. Sci. Rep. 2017, 7, 14991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, H.; Jeter, R.; Bachmann, A.; Yount, S.T.; Shen, C.L.; Yeganehjoo, H. The potential of isoprenoids in adjuvant cancer therapy to reduce adverse effects of statins. Front. Pharmacol. 2019, 9, 1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagunin, A.; Zakharov, A.; Filimonov, D.; Poroikov, V. QSAR modelling of rat acute toxicity on the basis of PASS prediction. Mol. Inform. 2011, 30, 241–250. [Google Scholar] [CrossRef]
- Filimonov, D.A.; Lagunin, A.A.; Gloriozova, T.A.; Rudik, A.V.; Druzhilovskii, D.S.; Pogodin, P.V.; Poroikov, V.V. Prediction of the biological activity spectra of organic compounds using the pass online web resource. Chem. Heterocycl. Compd. 2014, 50, 444–457. [Google Scholar] [CrossRef]
- Druzhilovskiy, D.S.; Rudik, A.V.; Filimonov, D.A.; Gloriozova, T.A.; Lagunin, A.A.; Dmitriev, A.V.; Pogodin, P.V.; Dubovskaya, V.I.; Ivanov, S.M.; Tarasova, O.A.; et al. Computational platform Way2Drug: From the prediction of biological activity to drug repurposing. Russ. Chem. Bull. 2017, 66, 1832–1841. [Google Scholar] [CrossRef]
- Rudik, A.V.; Dmitriev, A.V.; Lagunin, A.A.; Filimonov, D.A.; Poroikov, V.V. PASS-based prediction of metabolites detection in biological systems. SAR QSAR Environ. Res. 2019, 30, 751–758. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sundaram, C.; Prasad, S.; Kannappan, R. Tocotrienols, the vitamin E of the 21st century: Its potential against cancer and other chronic diseases. Biochem. Pharmacol. 2010, 80, 1613–1631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keiser, M.J.; Roth, B.L.; Armbruster, B.N.; Ernsberger, P.; Irwin, J.J.; Shoichet, B.K. Relating protein pharmacology by ligand chemistry. Nat. Biotechnol. 2007, 25, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Liang, L.; Yin, Z.; Lin, J. Improving chemical similarity ensemble approach in target prediction. J. Cheminform. 2016, 8, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, K.-Y.; Pang, K.-L.; Soelaiman, I.-N. Tocotrienol and Its Role in Chronic Diseases. In Advances in Experimental Medicine and Biology; Gupta, S.C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 928, pp. 97–130. ISBN 9783319413341. [Google Scholar]
- Sever, N.; Song, B.L.; Yabe, D.; Goldstein, J.L.; Brown, M.S.; DeBose-Boydb, R.A. Insig-dependent Ubiquitination and Degradation of Mammalian 3-Hydroxy-3-methylglutaryl-CoA Reductase Stimulated by Sterols and Geranylgeraniol. J. Biol. Chem. 2003, 278, 52479–52490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, M.M.; Elsabrouty, R.; Seemann, J.; Jo, Y.; DeBose-Boyd, R.A. The prenyltransferase UBIAD1 is the target of geranylgeraniol in degradation of HMG CoA reductase. eLife 2015, 4, e05560. [Google Scholar] [CrossRef]
- Scarpino, A.; Ferenczy, G.G.; Keserű, G.M. Comparative Evaluation of Covalent Docking Tools. J. Chem. Inf. Model. 2018, 58, 1441–1458. [Google Scholar] [CrossRef]
- Morand, O.H.; Aebi, J.D.; Dehmlow, H.; Ji, Y.H.; Gains, N.; Lengsfeld, H.; Himber, J. Ro 48-8071, a new 2,3-oxidosqualene:lanosterol cyclase inhibitor lowering plasma cholesterol in hamsters, squirrel monkeys, and minipigs: Comparison to simvastatin. J. Lipid Res. 1997, 38, 373–390. [Google Scholar] [CrossRef]
- Thoma, R.; Schulz-Gasch, T.; D’Arcy, B.; Benz, J.; Aebi, J.; Dehmlow, H.; Hennig, M.; Stihle, M.; Ruf, A. Insight into steroid scaffold formation from the structure of human oxidosqualene cyclase. Nature 2004, 432, 118–122. [Google Scholar] [CrossRef]
- Padyana, A.K.; Gross, S.; Jin, L.; Cianchetta, G.; Narayanaswamy, R.; Wang, F.; Wang, R.; Fang, C.; Lv, X.; Biller, S.A.; et al. Structure and inhibition mechanism of the catalytic domain of human squalene epoxidase. Nat. Commun. 2019, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Gunasekaran, B.; Shukor, M.Y. HMG-CoA Reductase as Target for Drug Development. Methods Mol. Biol. 2020, 2089, 245–250. [Google Scholar] [CrossRef]
- Nepomuceno, R.; de, F. Vallerini, B.; da Silva, R.L.; Corbi, S.C.T.; Bastos, A.D.S.; dos Santos, R.A.; Takahashi, C.S.; Regina, P.; Orrico, S.; Scarel-Caminaga, R.M. Systemic expression of genes related to inflammation and lipid metabolism in patients with dyslipidemia, type 2 diabetes mellitus and chronic periodontitis. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 2715–2722. [Google Scholar] [CrossRef]
- Jiang, Y.; Du, H.; Liu, X.; Fu, X.; Li, X.; Cao, Q. Artemisinin alleviates atherosclerotic lesion by reducing macrophage inflammation via regulation of AMPK/NF-κB/NLRP3 inflammasomes pathway. J. Drug Target. 2020, 28, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Lontchi-Yimagou, E.; Sobngwi, E.; Matsha, T.E.; Kengne, A.P. Diabetes mellitus and inflammation. Curr. Diabetes Rep. 2013, 13, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, A.; Chopra, K. Tocotrienol attenuates oxidative-nitrosative stress and inflammatory cascade in experimental model of diabetic neuropathy. Neuropharmacology 2009, 57, 456–462. [Google Scholar] [CrossRef]
- Chung, E.; Elmassry, M.M.; Kottapalli, P.; Kottapalli, K.R.; Kaur, G.; Dufour, J.M.; Wright, K.; Ramalingam, L.; Moustaid-Moussa, N.; Wang, R.; et al. Metabolic benefits of annatto-extracted tocotrienol on glucose homeostasis, inflammation, and gut microbiome. Nutr. Res. 2020, 77, 97–107. [Google Scholar] [CrossRef]
- Kuhad, A.; Bishnoi, M.; Tiwari, V.; Chopra, K. Suppression of NF-κβ signaling pathway by tocotrienol can prevent diabetes associated cognitive deficits. Pharmacol. Biochem. Behav. 2009, 92, 251–259. [Google Scholar] [CrossRef]
- Wong, S.K.; Chin, K.Y.; Suhaimi, F.H.; Ahmad, F.; Ima-Nirwana, S. The effects of palm tocotrienol on metabolic syndrome and bone loss in male rats induced by high-carbohydrate high-fat diet. J. Funct. Foods 2018, 44, 246–254. [Google Scholar] [CrossRef]
- Kim, Y.; Wang, W.; Okla, M.; Kang, I.; Moreau, R.; Chung, S. Suppression of NLRP3 inflammasome by γ -tocotrienol ameliorates type 2 diabetes. J. Lipid Res. 2016, 57, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Kuhad, A.; Chopra, K. Attenuation of diabetic nephropathy by tocotrienol: Involvement of NFkB signaling pathway. Life Sci. 2009, 84, 296–301. [Google Scholar] [CrossRef]
- Li, S.; Jiang, M.; Wang, L.; Yu, S. Combined chemotherapy with cyclooxygenase-2 (COX-2) inhibitors in treating human cancers: Recent advancement. Biomed. Pharmacother. 2020, 129, 110389. [Google Scholar] [CrossRef]
- Orlando, B.J.; Malkowski, M.G. Substrate-selective Inhibition of Cyclooxygeanse-2 by Fenamic Acid Derivatives Is Dependent on Peroxide Tone. J. Biol. Chem. 2016, 291, 15069–15081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shridas, P.; Webb, N.R. Diverse Functions of Secretory Phospholipases A2. Adv. Vasc. Med. 2014, 2014, 689815. [Google Scholar] [CrossRef] [Green Version]
- Hay, M.; Thomas, D.W.; Craighead, J.L.; Economides, C.; Rosenthal, J. Clinical development success rates for investigational drugs. Nat. Biotechnol. 2014, 32, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.H.; Le, J.; Abraham, M.H.; Hersey, A.; Eddershaw, P.J.; Luscombe, C.N.; Boutina, D.; Beck, G.; Sherborne, B.; Cooper, I.; et al. Evaluation of human intestinal absorption data and subsequent derivation of a quantitative structure—Activity relationship (QSAR) with the Abraham descriptors. J. Pharm. Sci. 2001, 90, 749–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Zoete, V. A Boiled-Egg to Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.M.V.; de Andrade, F.d.C.P.; Filgueiras, L.A.; de Carvalho Maia, O.A.; Cunha, R.L.; Rodezno, S.V.; Maia Filho, A.L.M.; de Amorim Carvalho, F.A.; Braz, D.C.; Mendes, A.N. preADMET analysis and clinical aspects of dogs treated with the Organotellurium compound RF07: A possible control for canine visceral leishmaniasis? Environ. Toxicol. Pharmacol. 2020, 80, 103470. [Google Scholar] [CrossRef]
- Yamashita, S.; Konishi, K.; Yamazaki, Y.; Taki, Y.; Sakane, T.; Sezaki, H.; Furuyama, Y. New and better protocols for a short-term Caco-2 cell culture system. J. Pharm. Sci. 2002, 91, 669–679. [Google Scholar] [CrossRef]
- Roman, D.L.; Roman, M.; Som, C.; Schmutz, M.; Hernandez, E.; Wick, P.; Casalini, T.; Perale, G.; Ostafe, V.; Isvoran, A. Computational Assessment of the Pharmacological Profiles of Degradation Products of Chitosan. Front. Bioeng. Biotechnol. 2019, 7, 214. [Google Scholar] [CrossRef]
- Ambrogini, P.; Torquato, P.; Bartolini, D.; Albertini, M.C.; Lattanzi, D.; Di Palma, M.; Marinelli, R.; Betti, M.; Minelli, A.; Cuppini, R.; et al. Excitotoxicity, neuroinflammation and oxidant stress as molecular bases of epileptogenesis and epilepsy-derived neurodegeneration: The role of vitamin E. Biochim. Biophys. Acta-Mol. Basis Dis. 2019, 1865, 1098–1112. [Google Scholar] [CrossRef]
- Gumpricht, E.; Rockway, S. Can ω-3 fatty acids and tocotrienol-rich vitamin E reduce symptoms of neurodevelopmental disorders? Nutrition 2014, 30, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, A.A.; Khan, D.A. Pharmacokinetics and Bioavailability of Annatto δ-tocotrienol in Healthy Fed Subjects. J. Clin. Exp. Cardiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.A.; Khan, D.A. Evaluation of Pharmacokinetics, and Bioavailability of Higher Doses of Tocotrienols in Healthy Fed Humans. J. Clin. Exp. Cardiol. 2016, 7, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Cardoza, C.G.; Vique-Sánchez, J.L. Potential inhibitors of the interaction between ACE2 and SARS-CoV-2 (RBD), to develop a drug. Life Sci. 2020, 256, 117970. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.A. Die Geschichte des ChemDraw-Projekts. Angew. Chem. 2014, 126, 11320–11325. [Google Scholar] [CrossRef]
- Nagamani, S.; Kesavan, C.; Muthusamy, K. Atom-based and Pharmacophore-based 3D–QSAR Studies on Vitamin D Receptor (VDR). Comb. Chem. High Throughput Screen. 2018, 21, 329–343. [Google Scholar] [CrossRef]
- Schyman, P.; Liu, R.; Wallqvist, A. General Purpose 2D and 3D Similarity Approach to Identify hERG Blockers. J. Chem. Inf. Model. 2016, 56, 213–222. [Google Scholar] [CrossRef]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, D.; Davis, A.M.; Kleywegt, G.J.; Schmitt, S. An Alternative Method for the Evaluation of Docking Performance: RSR vs RMSD. J. Chem. Inf. Model. 2008, 48, 1411–1422. [Google Scholar] [CrossRef]
- Matias Pereira, A.C.; Sánchez-Ortíz, B.L.; de Melo, E.L.; da Silva Hage-Melim, L.I.; Borges, R.S.; Hu, X.; Carvalho, J.C.T. Perillyl alcohol decreases the frequency and severity of convulsive-like behavior in the adult zebrafish model of acute seizures. Naunyn. Schmiedebergs Arch. Pharmacol. 2021, 394, 1177–1190. [Google Scholar] [CrossRef] [PubMed]
- Ruswanto; Siswandono; Richa, M.; Tita, N.; Tresna, L. Molecular docking of 1-benzoyl-3-methylthiourea as anti cancer candidate and its absorption, distribution, and toxicity prediction. J. Pharm. Sci. Res. 2017, 9, 680–684. [Google Scholar]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Pa | Pi | Activity Prediction |
|---|---|---|---|
| Geranylgeraniol | 0.953 | 0.003 | Mucous membrane protection |
| 0.885 | 0.004 | Lipid metabolism regulation | |
| 0.840 | 0.003 | TNF inhibitor | |
| 0.770 | 0.004 | Antiulcerative | |
| 0.743 | 0.049 | Antineoplastic | |
| 0.686 | 0.015 | Hypolipidemic | |
| 0.636 | 0.007 | NF kappa B regulator | |
| 0.643 | 0.024 | Anti-inflammatory | |
| 0.570 | 0.015 | Antihypercholesterolemic | |
| 0.549 | 0.005 | Antioxidant | |
| 0.538 | 0.03 | Cholesterol antagonist | |
| 0.498 | 0.019 | Antineoplastic | |
| 0.437 | 0.007 | Cholesterol synthesis inhibitor | |
| α-tocotrienol | 0.989 | 0.001 | Lipid peroxidase inhibitor |
| 0.973 | 0.002 | Antioxidant | |
| 0.962 | 0.002 | Antihypercholesterolemic | |
| 0.900 | 0.005 | Treatment of acute neural disorders | |
| 0.892 | 0.005 | Cerebral anti-ischemic | |
| 0.866 | 0.005 | Anti-inflammatory | |
| 0.863 | 0.003 | Peroxidase inhibitor | |
| 0.763 | 0.005 | Hepatoprotector | |
| 0.753 | 0.034 | Mucous membrane protection | |
| 0.713 | 0.008 | Cholesterol antagonist | |
| 0.702 | 0.001 | Cholesterol synthesis inhibition | |
| 0.685 | 0.003 | NOS2 expression inhibition | |
| 0.621 | 0.009 | Antineoplastic (breast cancer) | |
| 0.456 | 0.033 | NF kappa B inhibitor | |
| 0.426 | 0.031 | Atherosclerosis treatment | |
| 0.435 | 0.046 | TNF inhibitor | |
| 0.397 | 0.044 | Antipsoriasis | |
| 0.255 | 0.017 | Phospholipase A2 inhibition | |
β-tocotrienol | 0.957 | 0.002 | Lipid peroxidase inhibition |
| 0.951 | 0.002 | Antioxidant | |
| 0.951 | 0.002 | Antihypercholesterolemic | |
| 0.881 | 0.004 | Hypolipidemic | |
| 0.835 | 0.005 | Anti-inflammatory | |
| 0.812 | 0.005 | Anticarcinogenic | |
| 0.787 | 0.004 | Antiulcerative | |
| 0.744 | 0.002 | NOS2 expression inhibition | |
| 0.738 | 0.040 | Mucous membrane protection | |
| 0.692 | 0.001 | Cholesterol synthesis inhibition | |
| 0.714 | 0.026 | Cerebral anti-ischemic | |
| 0.685 | 0.008 | Hepatoprotector | |
| 0.648 | 0.035 | Antineoplastic | |
| 0.602 | 0.019 | Cholesterol antagonist | |
| 0.475 | 0.027 | Antipsoriasis | |
| 0.481 | 0.034 | TNF inhibitor | |
| 0.355 | 0.010 | NF kappa B inhibitor | |
| 0.271 | 0.026 | Lipoprotein disorder treatment | |
| 0.198 | 0.025 | Phospholipase A2 inhibition | |
| ɣ-tocotrienol | 0.977 | 0.002 | Lipid peroxidase inhibition |
| 0.953 | 0.002 | Antioxidant | |
| 0.944 | 0.002 | Antihypercholesterolemic | |
| 0.882 | 0.004 | Hypolipidemic | |
| 0.846 | 0.005 | Anti-inflammatory | |
| 0.811 | 0.005 | Anticarcinogenic | |
| 0.776 | 0.017 | Cerebral anti-ischemic | |
| 0.762 | 0.004 | Antiulcerative | |
| 0.686 | 0.001 | Cholesterol synthesis inhibitor | |
| 0.682 | 0.008 | Hepatoprotector | |
| 0.719 | 0.008 | Mucous membrane protection | |
| 0.683 | 0.003 | NOS2 expression inhibition | |
| 0.593 | 0.011 | Antineoplastic (breast cancer) | |
| 0.452 | 0.041 | TNF inhibitor | |
| 0.464 | 0.061 | Lipid metabolism inhibitor | |
| 0.402 | 0.043 | Antipsoriasis | |
| 0.271 | 0.014 | NF kappa B inhibitor | |
| 0.230 | 0.016 | Phospholipase A2 inhibition | |
| 0.280 | 0.091 | Atherosclerosis treatment | |
| δ-tocotrienol | 0.941 | 0.002 | Lipid peroxidase inhibition |
| 0.913 | 0.003 | Antioxidant | |
| 0.813 | 0.006 | Anti-inflammatory | |
| 0.803 | 0.005 | Antihypercholesterolemic | |
| 0.791 | 0.008 | Hypolipidemic | |
| 0.789 | 0.022 | Mucous membrane protection | |
| 0.745 | 0.002 | NOS2 expression inhibition | |
| 0.683 | 0.005 | Antiulcerative | |
| 0.663 | 0.001 | Cholesterol synthesis inhibition | |
| 0.650 | 0.011 | Anticarcinogenic | |
| 0.642 | 0.036 | Antineoplastic | |
| 0.589 | 0.013 | Hepatoprotector | |
| 0.522 | 0.025 | TNF inhibition | |
| 0.512 | 0.027 | Antithrombotic | |
| 0.515 | 0.041 | Lipid metabolism regulation | |
| 0.458 | 0.03 | Antipsoriasis | |
| 0.444 | 0.147 | Cerebral anti-ischemic | |
| 0.385 | 0.007 | NF kappa B inhibitor | |
| 0.224 | 0.038 | Lipoprotein disorder regulator | |
| 0.201 | 0.024 | Phospholipase A2 inhibitor |
| Molecule | Target | p-Value | Max TC |
|---|---|---|---|
| Geranylgeraniol | Squalene monooxygenase | 2.641 × 10−27 | 0.65 |
| Lanosterol synthase | 4.01 × 10−19 | 0.40 | |
| Phospholipase A2 | 7.305 × 10−18 | 0.31 | |
| Protein-S-isoprenylcysteine O-methyltransferase | 1.703 × 10−65 | 0.53 | |
| Geranylgeranyl pyrophosphate synthase | 1.409 × 10−61 | 0.50 | |
| Transient receptor potential cation channel subfamily V member 2 | 1.407 × 10−49 | 0.38 | |
| Transient receptor potential cation channel subfamily A member 1 | 6.621 × 10−40 | 0.40 | |
| Protein farnesyltransferase subunit beta | 2.46 × 10−10 | 0.53 | |
| Protein farnesyltransferase/geranylgeranyltransferase type 1 subunit alpha | 9.389 × 10−10 | 0.53 | |
| α-tocotrienol | Alpha-tocopherol transfer protein | 4.81 × 10−63 | 0.52 |
| PH domain leucine-rich repeat-containing protein phosphatase 1 | 4.19 × 10−15 | 0.34 | |
| Phospholipase A2 | 6.66 × 10−09 | 0.30 | |
| Squalene monooxygenase | 8.61 × 10−09 | 0.31 | |
| DNA polymerase lambda | 1.39 × 10−08 | 0.29 | |
| Lanosterol synthase | 2.04 × 10−08 | 0.31 | |
| Geranylgeranyl pyrophosphate synthase | 9.35 × 10−04 | 0.29 | |
| β-tocotrienol | PH domain leucine-rich repeat-containing protein phosphatase 1 | 6.38 × 10−51 | 0.39 |
| Alpha-tocopherol transfer protein | 1.62 × 10−21 | 0.36 | |
| DNA Polymerase lambda | 1.69 × 10−20 | 0.33 | |
| Phospholipase A2 | 3 × 10−09 | 0.31 | |
| Squalene monooxygenase | 2.23 × 10−08 | 0.3 | |
| Lanosterol synthase | 1.2 × 10−06 | 0.3 | |
| γ-tocotrienol | PH domain leucine-rich repeat-containing protein phosphatase 1 | 3.13 × 10−65 | 0.57 |
| DNA polymerase lambda | 1.69 × 10−20 | 0.33 | |
| Alpha-tocopherol transfer protein | 1.61 × 10−09 | 0.32 | |
| Phospholipase A2 | 3 × 10−09 | 0.31 | |
| Squalene monooxygenase | 2.23 × 10−08 | 0.3 | |
| Lanosterol synthase | 9.42 × 10−07 | 0.31 | |
| δ-tocotrienol | PH domain leucine-rich repeat-containing protein phosphatase 1 | 7.59 × 10−50 | 0.39 |
| DNA polymerase lambda | 8.97 × 10−20 | 0.32 | |
| Phospholipase A2 | 3 × 10−09 | 0.31 | |
| Squalene monooxygenase | 2.23 × 10−08 | 0.3 | |
| Lanosterol synthase | 9.42 × 10−07 | 0.31 | |
| Hypoxia-inducible factor 1-alpha | 5.31 × 10−06 | 0.36 |
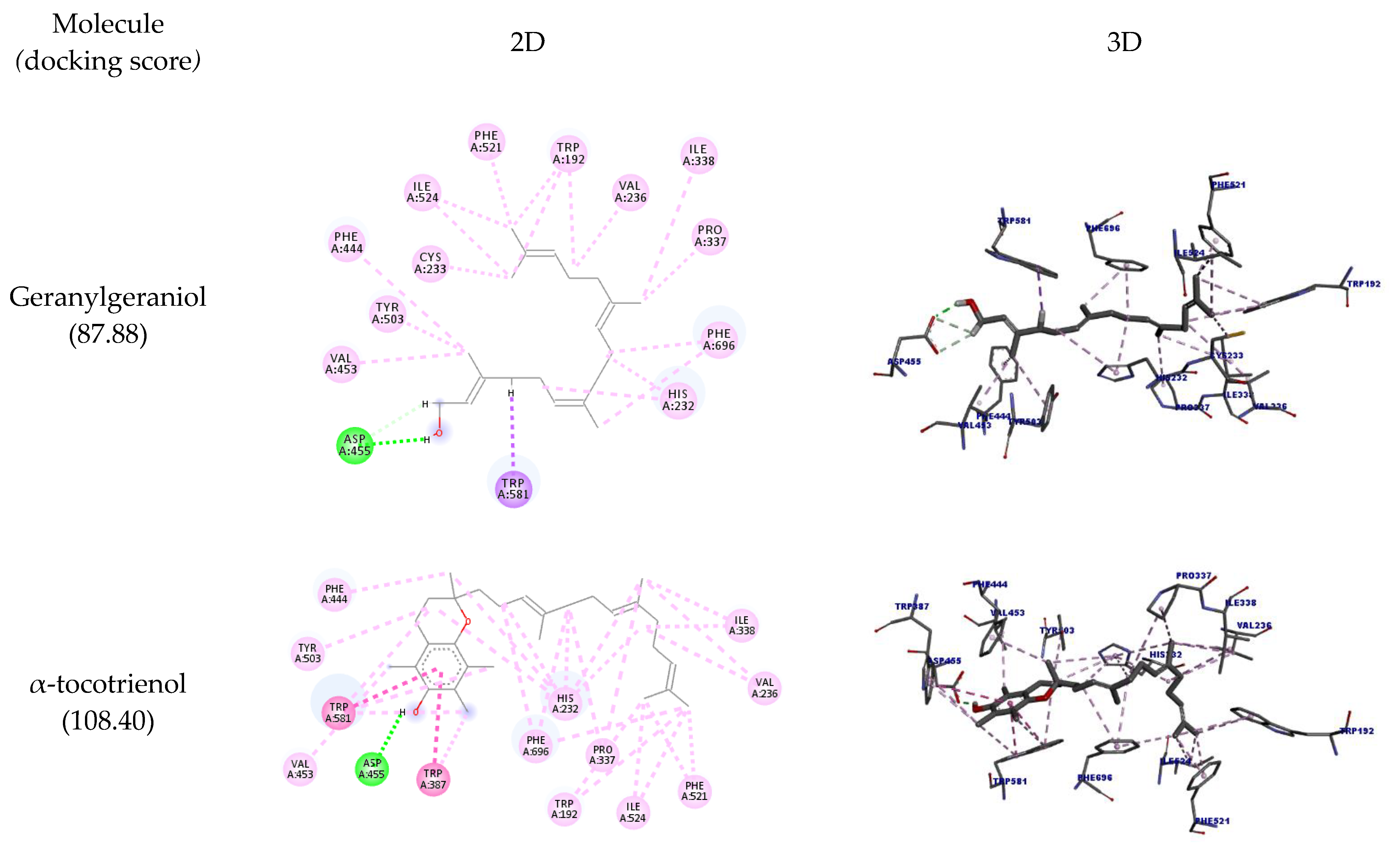
| Molecule | Amino Acid | Ligand Atom | Category | Types | Distance (Å) | Score |
|---|---|---|---|---|---|---|
| Geranylgeraniol | A:ASP455 | H25 | Hydrogen bond | Conventional hydrogen bond | 2.08 | 87.88 |
| 2.65 | ||||||
| 2.81 | ||||||
| A:TRP581 | H28 | Hydrophobic | Pi-sigma | 2.87 | ||
| A:VAL236 | Ligand | Alkyl | 5.36 | |||
| A:VAL453 | C14 | 3.94 | ||||
| A:PRO337 | C16 | 4.90 | ||||
| A:ILE338 | 5.25 | |||||
| A:ILE524 | C20 | 3.70 | ||||
| A:CYS233 | C21 | 4.53 | ||||
| A:ILE524 | 4.57 | |||||
| A:TRP192 | Ligand | Pi-alkyl | 5.12 | |||
| C20 | 5.40 | |||||
| C21 | 4.63 | |||||
| A:HIS232 | Ligand | 4.69 | ||||
| 4.24 | ||||||
| A:PHE444 | C14 | 4.92 | ||||
| A:TYR503 | 4.76 | |||||
| A:PHE521 | C20 | 3.93 | ||||
| A:PHE696 | Ligand | 3.91 | ||||
| C15 | 4.10 | |||||
| α-tocotrienol | A:ASP455 | H39 | Hydrogen bond | Conventional hydrogen bond | 1.86 | 108.40 |
| A:TRP581 | Ligand | Hydrophobic | Pi-pi stackedPi-pi T-shaped | 4.38 | ||
| 4.26 | ||||||
| A:TRP387 | 5.60 | |||||
| A:VAL236 | Alkyl | 5.28 | ||||
| A:PRO337 | 5.33 | |||||
| A:VAL453 | C11 | 4.26 | ||||
| A:ILE338 | Ligand | 5.18 | ||||
| A:VAL236 | C26 | 5.03 | ||||
| A:PRO337 | 4.99 | |||||
| A:ILE338 | 4.29 | |||||
| A:ILE524 | C30 | 3.55 | ||||
| C31 | 5.29 | |||||
| A:TRP192 | C30 | Pi-alkyl | 4.71 | |||
| C31 | 5.27 | |||||
| A:HIS232 | ligand | 4.84 | ||||
| 5.07 | ||||||
| C16 | 5.24 | |||||
| ligand | 5.31 | |||||
| 4.69 | ||||||
| C26 | 4.93 | |||||
| A:TRP387 | C13 | 4.96 | ||||
| 4.30 | ||||||
| A:PHE444 | C16 | 4.46 | ||||
| A:TYR503 | Ligand | 5.11 | ||||
| A:PHE521 | C30 | 4.58 | ||||
| C31 | 3.62 | |||||
| A:TRP581 | C13 | 5.00 | ||||
| Ligand | 4.64 | |||||
| C14 | 5.26 | |||||
| A:PHE696 | ligand | 4.76 | ||||
| 4.55 | ||||||
| C31 | 5.39 | |||||
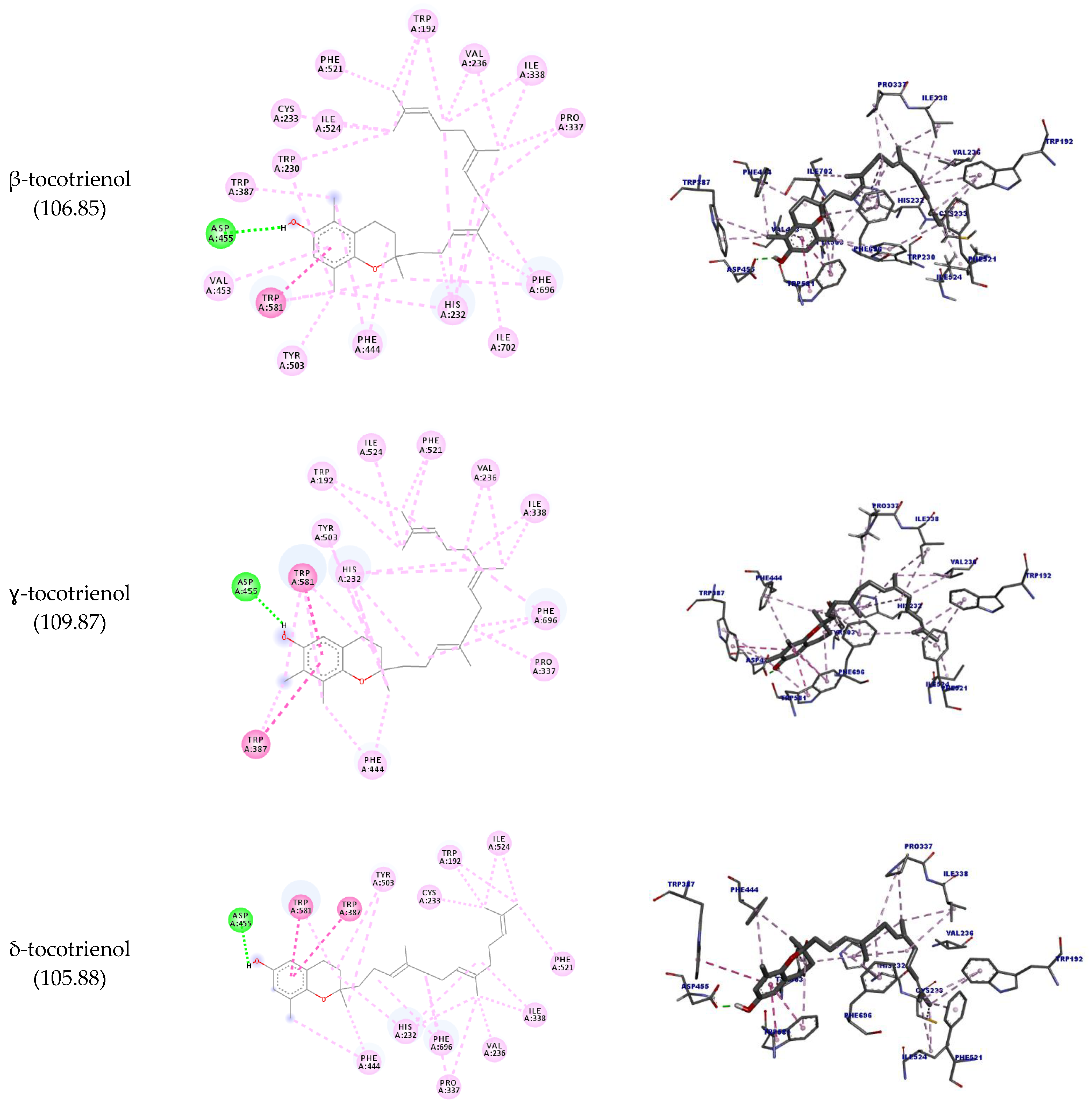
| β-tocotrienol | A:ASP455 | H39 | Hydrogen bond | Conventional hydrogen bond | 2.17 | 106.85 |
| A:TRP581 | Ligand | Hydrophobic | Pi-pi stacked | 4.80 | ||
| 4.27 | ||||||
| A:VAL236 | Alkyl | 4.48 | ||||
| A:PRO337 | 4.72 | |||||
| A:ILE702 | C20 | 4.41 | ||||
| A:ILE338 | Ligand | 5.15 | ||||
| C25 | 5.33 | |||||
| A:PRO337 | 4.40 | |||||
| A:ILE338 | 4.09 | |||||
| A:CYS233 | C29 | 4.57 | ||||
| A:ILE524 | 3.70 | |||||
| A:TRP192 | Ligand | Pi-alkyl | 5.17 | |||
| C29 | 5.46 | |||||
| C30 | 5.08 | |||||
| A:TRP230 | C13 | 5.26 | ||||
| 4.85 | ||||||
| C29 | 5.01 | |||||
| A:HIS232 | C13 | 4.70 | ||||
| Ligand | 4.15 | |||||
| 5.49 | ||||||
| 5.23 | ||||||
| C25 | 5.33 | |||||
| A:TRP387 | C11 | 4.72 | ||||
| 3.73 | ||||||
| A:PHE444 | Ligand | 4.13 | ||||
| C11 | 4.51 | |||||
| A:TYR503 | C13 | 4.59 | ||||
| A:PHE521 | C30 | 3.69 | ||||
| A:TRP581 | C13 | 4.02 | ||||
| C15 | 4.97 | |||||
| A:PHE696 | 5.20 | |||||
| Ligand | 5.19 | |||||
| C20 | 3.83 | |||||
| A:VAL453 | Ligand | 5.32 | ||||
| ɣ-tocotrienol | A:ASP455 | H36 | Hydrogen bond | Conventional hydrogen bond | 1.83 | 109.87 |
| A:TRP581 | Ligand | Hydrophobic | Pi-pi stacked | 4.31 | ||
| A:TRP581 | 4.18 | |||||
| A:TRP387 | 5.69 | |||||
| A:VAL236 | Alkyl | 5.38 | ||||
| A:PRO337 | 5.46 | |||||
| A:ILE338 | 5.15 | |||||
| A:VAL236 | 5.07 | |||||
| A:ILE338 | C24 | 4.37 | ||||
| A:ILE524 | 3.63 | |||||
| A:TRP192 | C28 | Pi-alkyl | 4.59 | |||
| C29 | 5.20 | |||||
| A:HIS232 | Ligand | 4.75 | ||||
| 5.09 | ||||||
| C14 | 5.07 | |||||
| Ligand | 4.95 | |||||
| C24 | 4.77 | |||||
| A:TRP387 | C30 | 4.90 | ||||
| 4.17 | ||||||
| A:PHE444 | C12 | 5.14 | ||||
| C14 | 4.59 | |||||
| A:TYR503 | Ligand | 5.01 | ||||
| C14 | 5.30 | |||||
| A:PHE521 | C28 | 4.18 | ||||
| C29 | 3.59 | |||||
| A:TRP581 | C30 | 4.96 | ||||
| Ligand | 4.61 | |||||
| C12 | 5.42 | |||||
| A:PHE696 | Ligand | 4.80 | ||||
| 4.34 | ||||||
| C29 | 5.23 | |||||
| δ-tocotrienol | A:ASP455 | H36 | Hydrogen bond | Conventional hydrogen bond | 1.66 | 105.88 |
| A:TRP581 | Ligand | Hydrophobic | Pi-pi stacked | 4.28 | ||
| 4.15 | ||||||
| A:TRP387 | Pi-pi T-shaped | 5.80 | ||||
| A:PRO337 | Alkyl | 5.47 | ||||
| A:ILE338 | 5.00 | |||||
| A:VAL236 | C24 | 5.46 | ||||
| A:PRO337 | 4.58 | |||||
| A:ILE338 | C28 | 4.28 | ||||
| A:CYS233 | 4.29 | |||||
| A:ILE524 | C29 | 3.72 | ||||
| C28 | 4.78 | |||||
| A:TRP192 | C29 | Pi-alkyl | 5.01 | |||
| Ligand | 5.31 | |||||
| A:HIS232 | 4.85 | |||||
| C24 | 4.95 | |||||
| C12 | 5.03 | |||||
| A:PHE444 | C14 | 4.48 | ||||
| Ligand | 3.76 | |||||
| A:TYR503 | C14 | 5.22 | ||||
| C29 | 5.42 | |||||
| A:PHE521 | Ligand | 3.92 | ||||
| A:TRP581 | 5.18 | |||||
| A:PHE696 | 5.18 | |||||
| H55 | 4.32 |
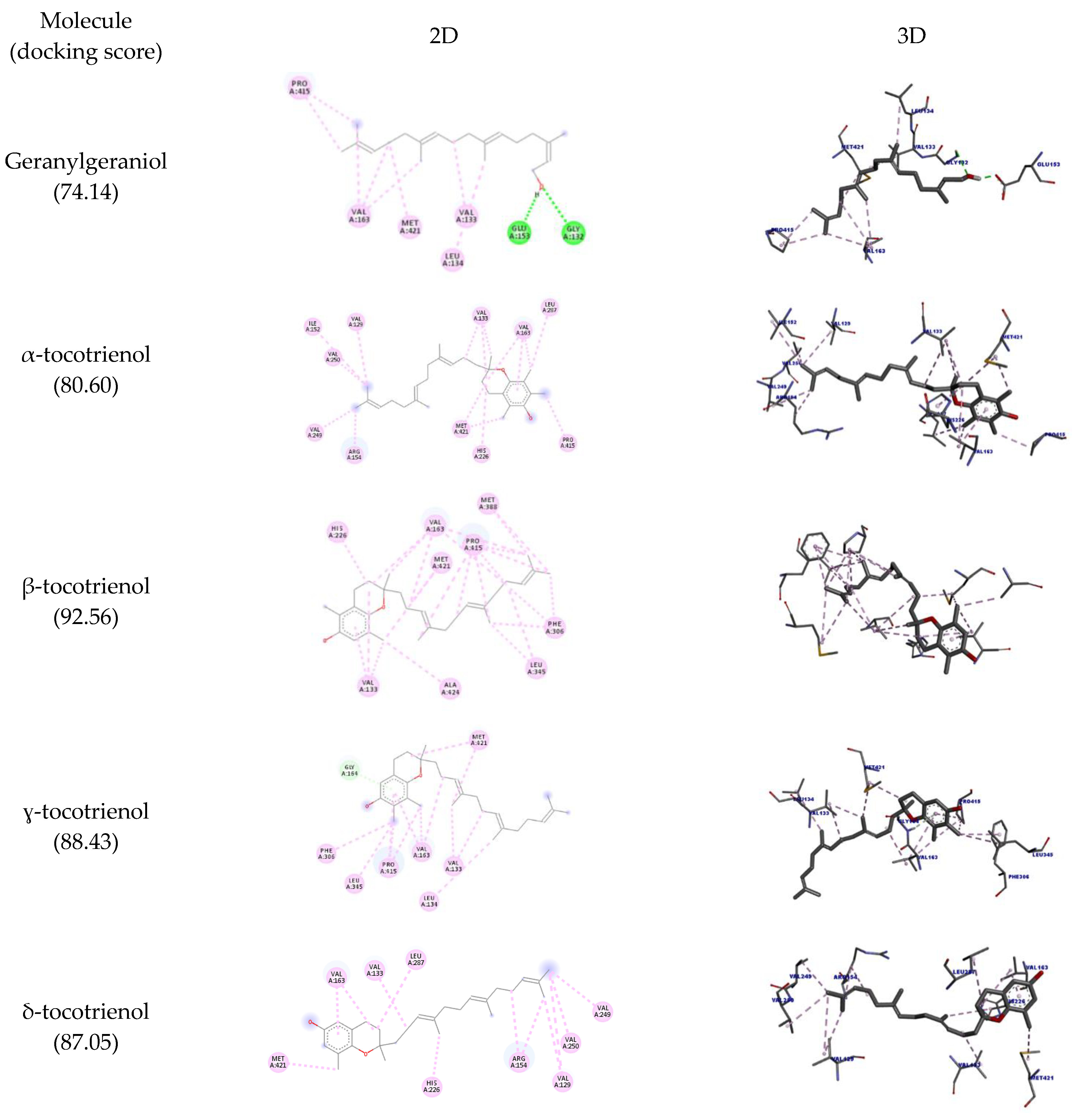
| Molecule | Amino Acid | Ligand Atom | Category | Types | Distance (Å) | Score |
|---|---|---|---|---|---|---|
| Geranylgeraniol | A:GLY132 | O24 | Hydrogen bond | Conventional hydrogen bond | 2.83 | 74.14 |
| A:GLU153 | H55 | 1.70 | ||||
| A:VAL133 | Ligand | Hydrophobic | Alkyl | 4.06 | ||
| A:VAL163 | 5.13 | |||||
| A:MET421 | 5.04 | |||||
| A:LEU134 | C15 | 4.41 | ||||
| A:VAL163 | C16 | 4.56 | ||||
| C20 | 4.57 | |||||
| A:PRO415 | 4.05 | |||||
| C21 | 4.27 | |||||
| α-tocotrienol | A:VAL133 | Ligand | Hydrophobic | Alkyl | 4.33 | 80.60 |
| 5.27 | ||||||
| A:VAL163 | 5.21 | |||||
| A:MET421 | 4.42 | |||||
| C11 | 4.04 | |||||
| A:PRO415 | C13 | 3.96 | ||||
| A:VAL163 | C14 | 3.71 | ||||
| A:LEU287 | 4.19 | |||||
| A:VAL133 | C16 | 4.70 | ||||
| A:VAL129 | C30 | 4.93 | ||||
| A:ILE152 | 5.02 | |||||
| A:VAL250 | 4.05 | |||||
| A:ARG154 | C31 | 4.14 | ||||
| A:VAL249 | 4.20 | |||||
| A:HIS226 | Ligand | Pi-alkyl | 4.96 | |||
| A:VAL163 | 4.36 | |||||
| β-tocotrienol | A:VAL133 | Ligand | Hydrophobic | Alkyl | 5.10 | 92.56 |
| A:VAL163 | 4.65 | |||||
| 5.00 | ||||||
| 4.55 | ||||||
| A:PRO415 | 4.92 | |||||
| 4.72 | ||||||
| A:ALA424 | C13 | 3.84 | ||||
| A:VAL133 | 4.39 | |||||
| A:MET421 | 5.43 | |||||
| Ligand | 4.71 | |||||
| A:VAL163 | C15 | 4.73 | ||||
| A:PRO415 | C20 | 4.76 | ||||
| A:LEU345 | Ligand | 5.16 | ||||
| C25 | 4.75 | |||||
| A:PRO415 | 4.32 | |||||
| A:VAL163 | C29 | 4.76 | ||||
| A:MET388 | 4.47 | |||||
| A:PRO415 | 4.60 | |||||
| A:MET388 | C30 | 4.90 | ||||
| A:PRO415 | 4.11 | |||||
| A:HIS226 | Ligand | Pi-alkyl | 4.95 | |||
| A:PHE306 | 5.25 | |||||
| C25 | 4.84 | |||||
| C30 | 4.56 | |||||
| A:VAL133 | Ligand | 3.93 | ||||
| ɣ-tocotrienol | GLY164 | ligand | Hydrogen bond | Pi-donor hydrogen bond | 2.67 | 88.43 |
| VAL133 | Hydrophobic | Alkyl | 4.71 | |||
| VAL163 | 4.67 | |||||
| MET421 | 4.68 | |||||
| VAL163 | C12 | 3.82 | ||||
| PRO415 | 4.40 | |||||
| VAL133 | C19 | 3.89 | ||||
| MET421 | 4.79 | |||||
| LEU134 | C24 | 5.06 | ||||
| LEU345 | C30 | 4.32 | ||||
| PRO415 | 4.60 | |||||
| PHE306 | Pi-alkyl | 4.73 | ||||
| VAL163 | Ligand | 5.45 | ||||
| PRO415 | 4.94 | |||||
| δ-tocotrienol | VAL133 | Ligand | Hydrophobic | Alkyl | 4.37 | 87.05 |
| ARG154 | 4.86 | |||||
| VAL163 | 5.06 | |||||
| LEU287 | 4.66 | |||||
| MET421 | C12 | 4.62 | ||||
| VAL129 | C28 | 5.47 | ||||
| VAL249 | 4.49 | |||||
| VAL250 | 4.51 | |||||
| VAL129 | C29 | 5.18 | ||||
| ARG154 | 5.13 | |||||
| HIS226 | C19 | Pi-alkyl | 5.47 | |||
| VAL163 | Ligand | 3.93 |
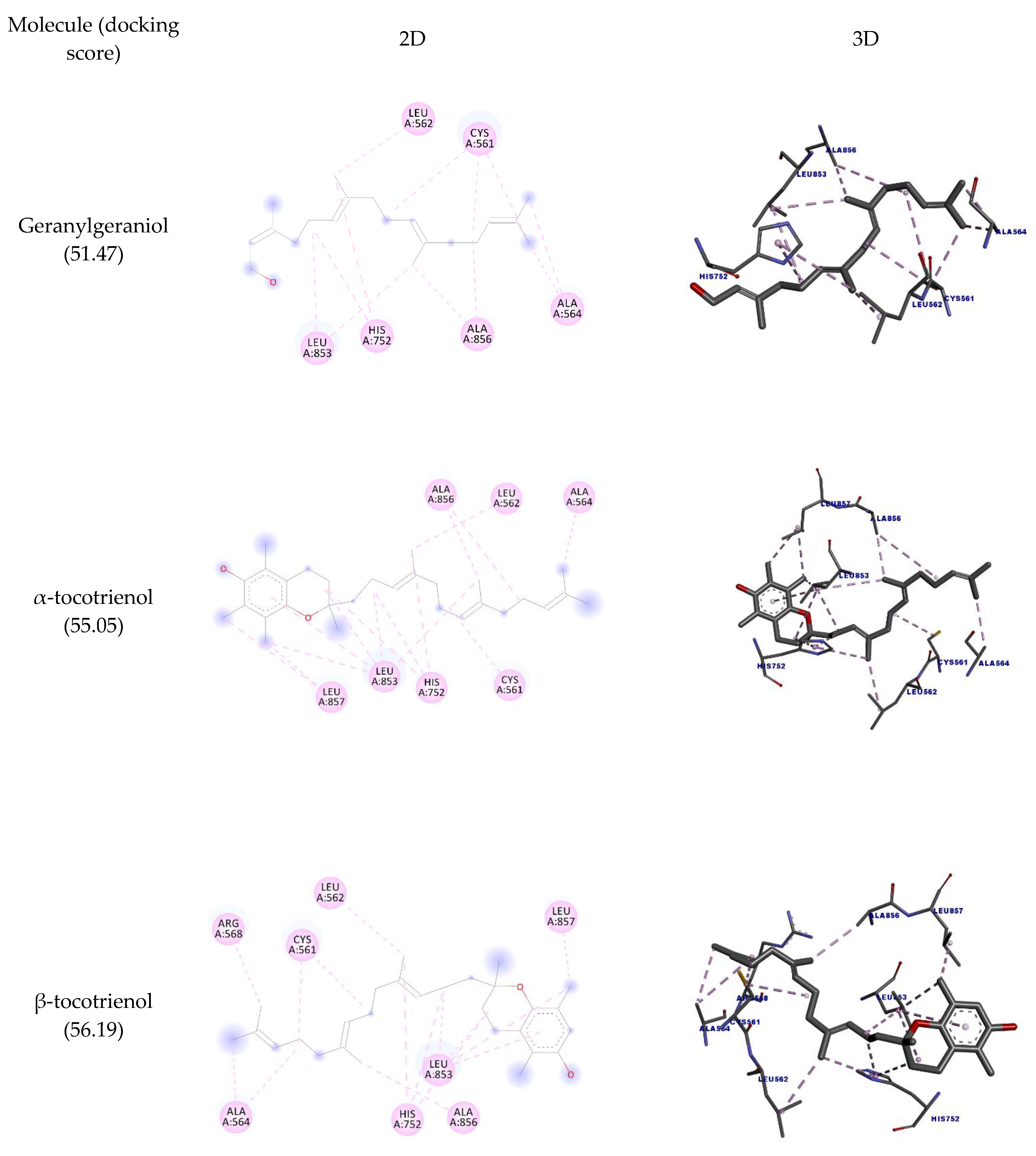
| Molecule | Amino Acid | Ligand Atom | Category | Types | Distance (Å) | Score |
|---|---|---|---|---|---|---|
| Geranylgeraniol | CYS561 | C9 | Hydrophobic | Alkyl | 4.49 | 51.47 |
| C17 | 5.46 | |||||
| ALA564 | C20 | 3.30 | ||||
| C21 | 3.71 | |||||
| ALA856 | C17 | 4.86 | ||||
| C16 | 3.49 | |||||
| LEU853 | C5 | 4.21 | ||||
| LEU562 | C15 | 3.83 | ||||
| LEU853 | C16 | 4.26 | ||||
| CYS561 | C20 | 3.86 | ||||
| HIS752 | C5 | Pi-alkyl | 4.36 | |||
| C15 | 4.97 | |||||
| α-tocotrienol | CYS561 | C22 | Hydrophobic | Alkyl | 4.18 | 55.05 |
| ALA564 | C31 | 3.51 | ||||
| ALA856 | C27 | 5.19 | ||||
| C26 | 3.30 | |||||
| LEU853 | C9 | 5.05 | ||||
| LEU857 | C13 | 4.29 | ||||
| LEU853 | C14 | 4.63 | ||||
| LEU857 | 4.30 | |||||
| LEU853 | C17 | 4.16 | ||||
| LEU562 | C21 | 3.90 | ||||
| LEU853 | C26 | 4.73 | ||||
| HIS752 | C9 | Pi-alkyl | 5.03 | |||
| C17 | 4.37 | |||||
| C21 | 4.95 | |||||
| LEU853 | Anel Ar. | 5.15 | ||||
| β-tocotrienol | CYS561 | C21 | Hydrophobic | Alkyl | 4.31 | 56.19 |
| C26 | 4.40 | |||||
| ALA564 | 4.37 | |||||
| C29 | 4.15 | |||||
| ALA856 | C25 | 3.63 | ||||
| LEU853 | C9 | 5.33 | ||||
| C13 | 4.56 | |||||
| LEU857 | 4.20 | |||||
| LEU853 | C16 | 4.19 | ||||
| LEU562 | C20 | 3.92 | ||||
| ARG568 | C30 | 3.97 | ||||
| HIS752 | C9 | 5.32 | ||||
| C16 | Pi-alkyl | 4.53 | ||||
| C20 | 4.71 | |||||
| LEU853 | Anel Ar. | 5.35 | ||||
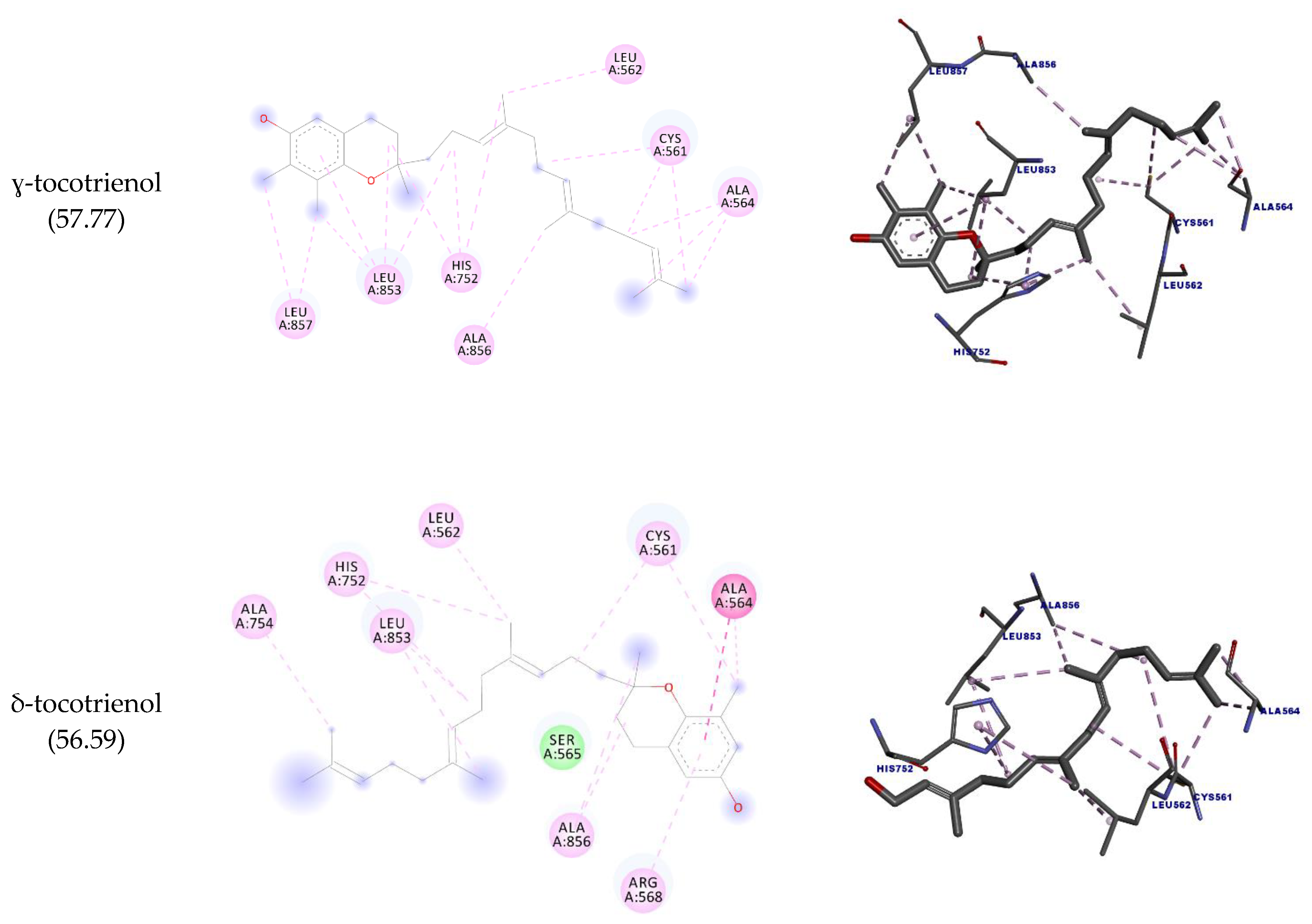
| ɣ-tocotrienol | CYS561 | C20 | Hydrophobic | Alkyl | 4.15 | 57.77 |
| C25 | 4.97 | |||||
| ALA564 | 4.87 | |||||
| C28 | 3.34 | |||||
| C29 | 3.70 | |||||
| ALA856 | C24 | 3.32 | ||||
| LEU853 | C9 | 4.93 | ||||
| C12 | 4.68 | |||||
| LEU857 | C12 | 4.41 | ||||
| LEU853 | C15 | 4.11 | ||||
| LEU562 | C19 | 4.04 | ||||
| CYS561 | C28 | 3.75 | ||||
| LEU857 | C30 | 4.24 | ||||
| HIS752 | C9 | Pi-alkyl | 4.88 | |||
| C15 | 4.40 | |||||
| C19 | 5.06 | |||||
| LEU853 | Anel Ar. | 5.01 | ||||
| δ-tocotrienol | ALA564 | Anel Ar. | Hydrophobic | Amide-pi stacked | 4.02 | 56.59 |
| CYS561 | C15 | Alkyl | 4.18 | |||
| ALA564 | C12 | 3.33 | ||||
| ALA754 | C29 | 4.03 | ||||
| ALA856 | C9 | 4.26 | ||||
| C14 | 4.43 | |||||
| CYS561 | C12 | 3.32 | ||||
| LEU853 | C20 | 4.24 | ||||
| LEU562 | C19 | 4.24 | ||||
| LEU853 | C24 | 4.74 | ||||
| HIS752 | C20 | Pi-alkyl | 4.27 | |||
| C19 | 4.56 | |||||
| ALA564 | Anel Ar. | 4.21 | ||||
| ARG568 | 5.40 |
| Molecule | Amino Acid | Ligand Atom | Category | Types | Distance (Å) | Score |
|---|---|---|---|---|---|---|
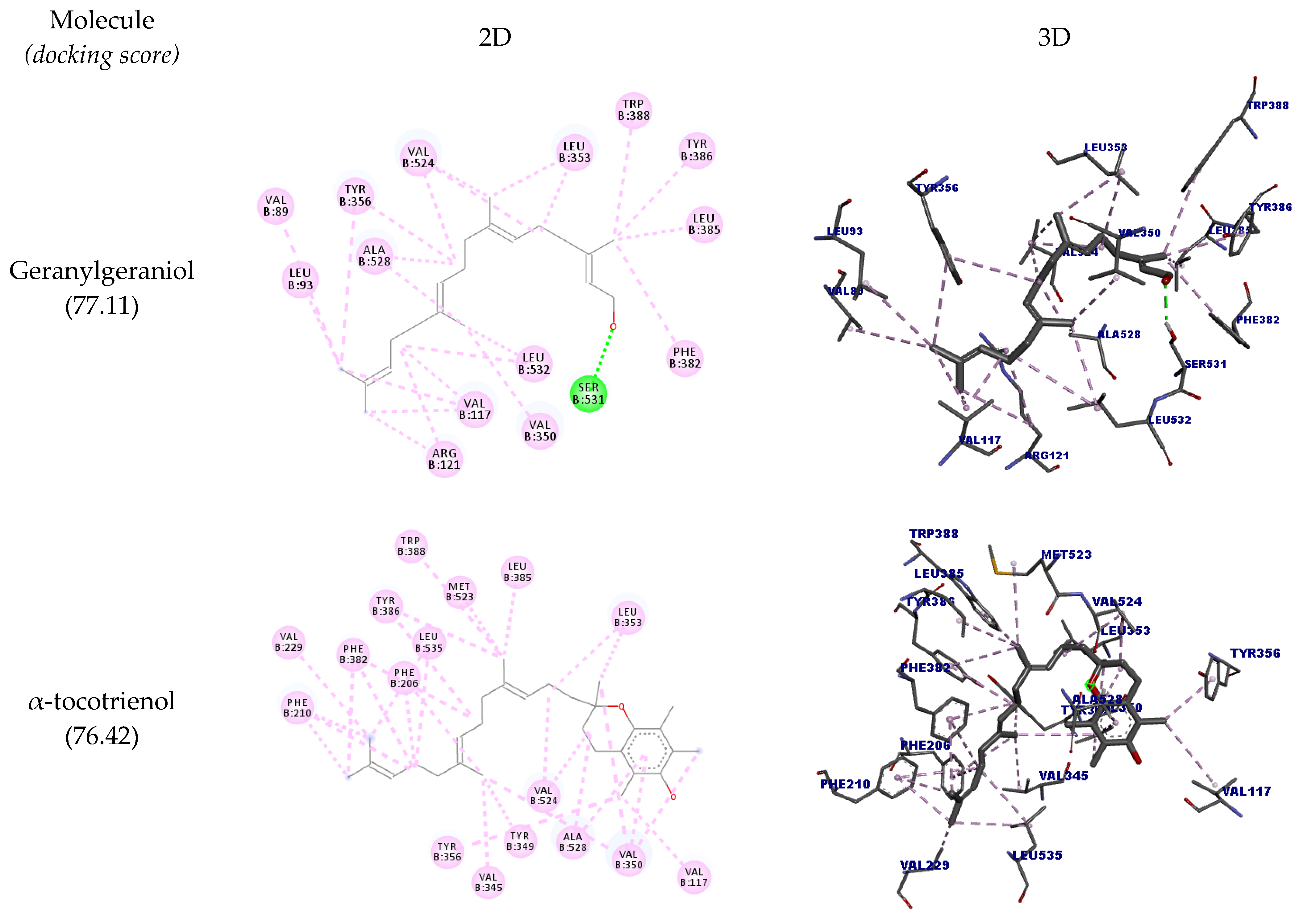
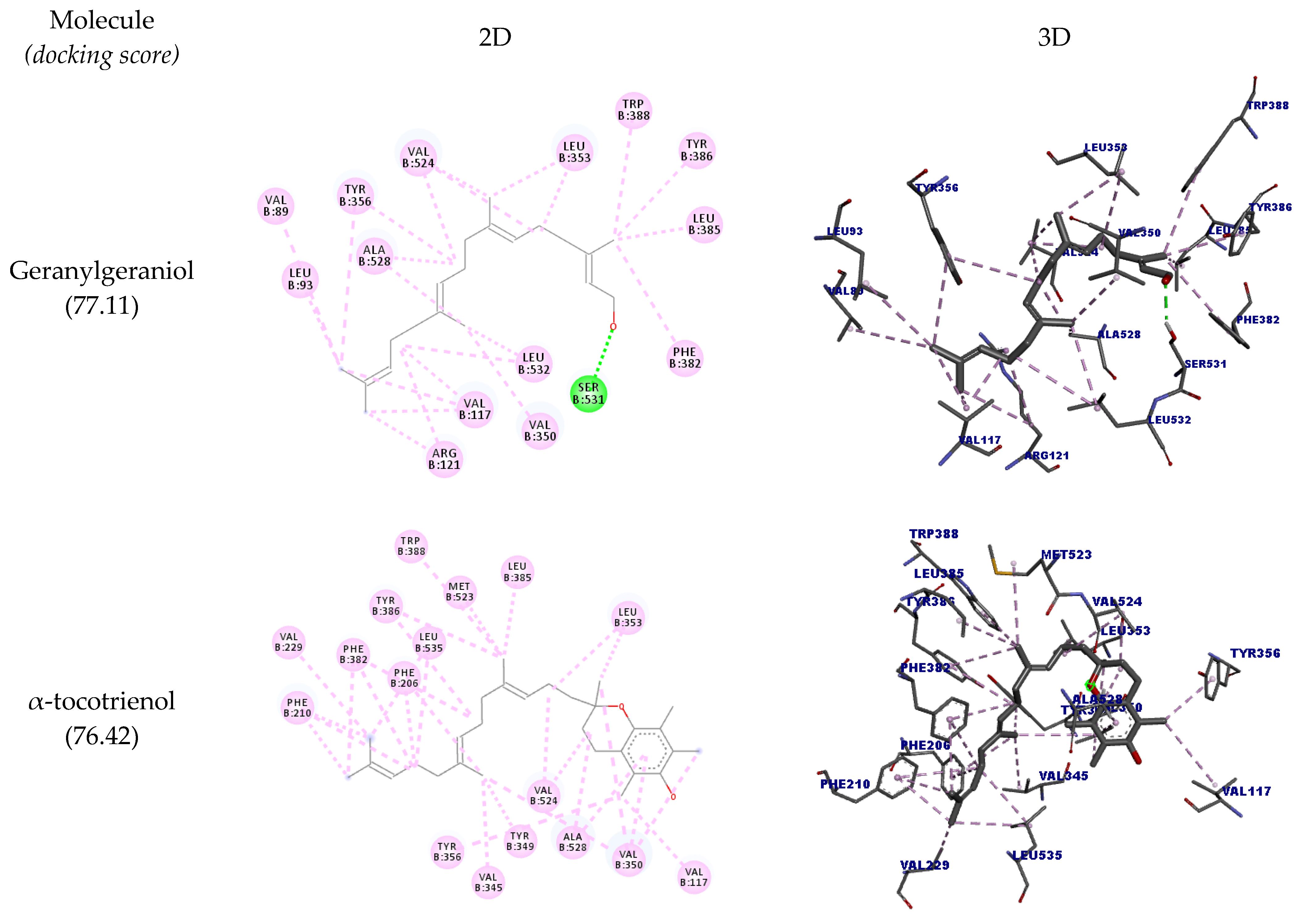
| Geranylgeraniol | B:SER531 | O24 | Hydrogen bond | Conventional hydrogen bond | 2.16 | 77.11 |
| B:VAL117 | Ligand | Hydrophobic | Alkyl | 3.86 | ||
| B:ARG121 | 5.14 | |||||
| B:VAL524 | 4.13 | |||||
| 4.59 | ||||||
| B:ALA528 | 3.99 | |||||
| C16 | 3.39 | |||||
| B:LEU353 | Ligand | 5.04 | ||||
| B:LEU532 | 5.28 | |||||
| B:LEU385 | C14 | 5.05 | ||||
| B:LEU353 | C15 | 4.18 | ||||
| B:VAL524 | 3.88 | |||||
| B:VAL350 | C16 | 4.57 | ||||
| B:LEU532 | 4.40 | |||||
| B:VAL89 | C20 | 5.46 | ||||
| B:LEU93 | 4.92 | |||||
| B:VAL117 | 4.43 | |||||
| C21 | 3.45 | |||||
| B:ARG121 | 4.48 | |||||
| B:TYR356 | Ligand | Pi-alkyl | 5.30 | |||
| C20 | 5.09 | |||||
| B:PHE382 | C14 | 5.48 | ||||
| B:TYR386 | 4.31 | |||||
| B:TRP388 | 4.91 | |||||
| α-tocotrienol | B:VAL524 | Ligand | Hydrophobic | Alkyl | 3.65 | 76.42 |
| 4.60 | ||||||
| B:ALA528 | 3.85 | |||||
| B:VAL117 | C11 | 5.03 | ||||
| B:VAL350 | C13 | 3.68 | ||||
| B:LEU353 | Ligand | 4.79 | ||||
| B:VAL350 | C16 | 5.10 | ||||
| B:LEU353 | 4.14 | |||||
| B:LEU385 | C21 | 4.79 | ||||
| B:MET523 | 4.91 | |||||
| B:LEU535 | Ligand | 4.79 | ||||
| B:VAL345 | C26 | 4.76 | ||||
| B:VAL350 | 5.09 | |||||
| B:VAL229 | C30 | 4.50 | ||||
| B:LEU535 | 4.93 | |||||
| B:PHE206 | Ligand | Pi-alkyl | 4.82 | |||
| C26 | 5.43 | |||||
| B:PHE210 | Ligand | 4.89 | ||||
| C30 | 4.27 | |||||
| C31 | 4.24 | |||||
| B:TYR349 | C26 | 4.50 | ||||
| B:TYR356 | C11 | 4.00 | ||||
| B:PHE382 | Ligand | 4.43 | ||||
| 5.15 | ||||||
| C31 | 4.60 | |||||
| B:TYR386 | Ligand | 4.49 | ||||
| C21 | 4.91 | |||||
| B:TRP388 | 4.99 | |||||
| B:VAL350 | Ligand | 4.04 | ||||
| B:ALA528 | 3.86 | |||||
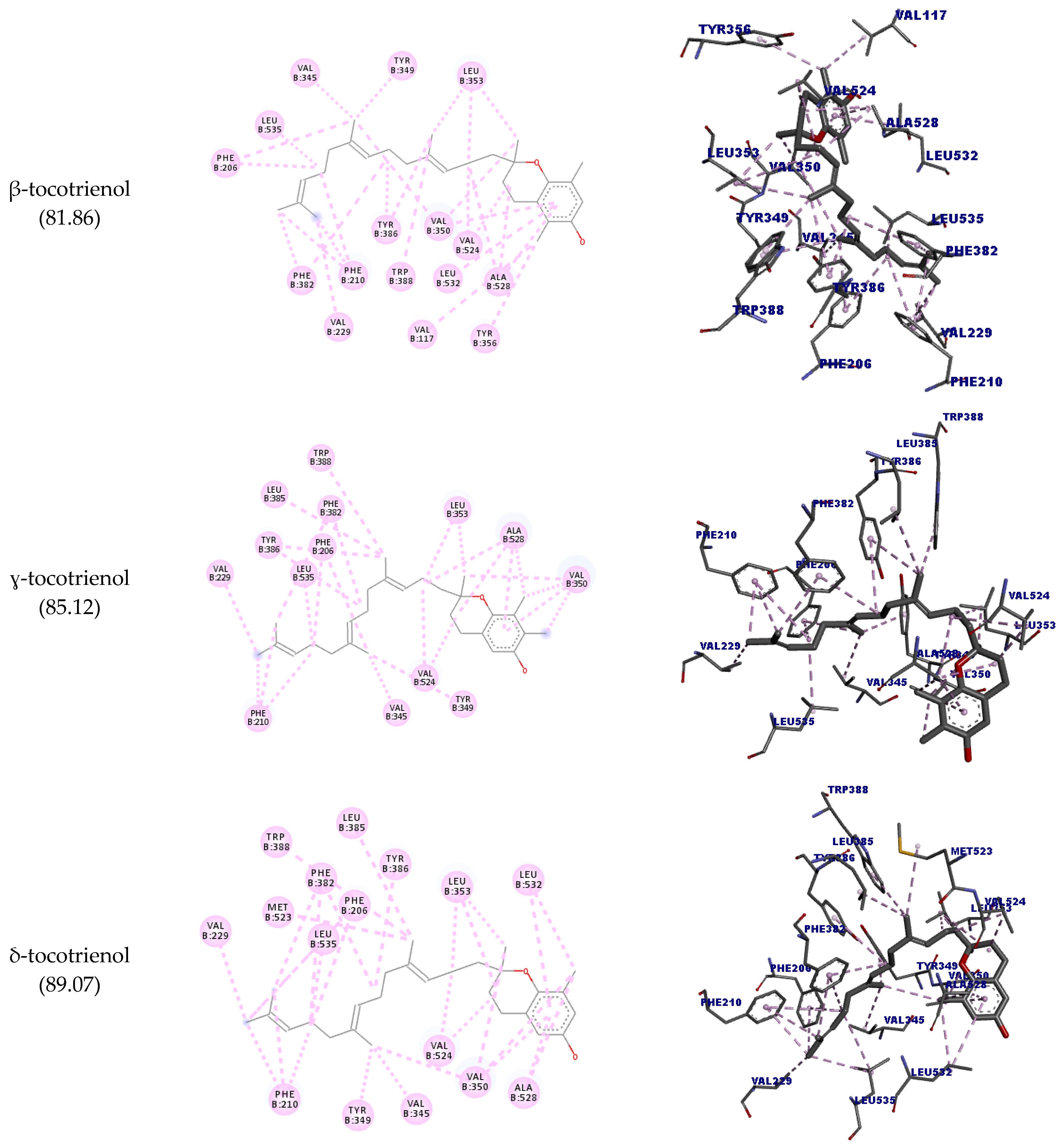
| β-tocotrienol | B:VAL524 | Ligand | Hydrophobic | Alkyl | 3.84 | 81.86 |
| 4.58 | ||||||
| B:ALA528 | 4.17 | |||||
| 4.48 | ||||||
| B:VAL117 | C11 | 5.10 | ||||
| B:LEU353 | Ligand | 5.34 | ||||
| B:VAL350 | C15 | 4.49 | ||||
| B:LEU353 | 3.92 | |||||
| C20 | 4.72 | |||||
| B:LEU535 | Ligand | 4.26 | ||||
| B:VAL345 | C25 | 4.39 | ||||
| B:VAL350 | 5.49 | |||||
| B:VAL229 | C29 | 4.89 | ||||
| B:PHE206 | Ligand | Pi-alkyl | 4.50 | |||
| C25 | 4.87 | |||||
| B:PHE210 | Ligand | 4.84 | ||||
| C29 | 3.85 | |||||
| C30 | 4.64 | |||||
| B:TYR349 | C25 | 4.77 | ||||
| B:TYR356 | C11 | 4.36 | ||||
| B:PHE382 | Ligand | 4.85 | ||||
| C30 | 4.53 | |||||
| B:TYR386 | Ligand | 4.57 | ||||
| C20 | 4.74 | |||||
| B:TRP388 | 4.55 | |||||
| B:VAL350 | Ligand | 4.32 | ||||
| B:ALA528 | 3.74 | |||||
| B:LEU532 | 4.74 | |||||
| ɣ-tocotrienol | B:VAL524 | Ligand | Hydrophobic | Alkyl | 3.62 | 85.12 |
| 4.85 | ||||||
| B:ALA528 | 4.50 | |||||
| C12 | 3.69 | |||||
| Ligand | 4.81 | |||||
| B:VAL350 | C12 | 4.19 | ||||
| B:LEU353 | Ligand | 4.95 | ||||
| B:VAL350 | C14 | 4.83 | ||||
| B:LEU353 | 3.85 | |||||
| B:LEU385 | C19 | 4.93 | ||||
| B:LEU535 | Ligand | 4.36 | ||||
| B:VAL345 | C24 | 4.59 | ||||
| B:VAL229 | C28 | 4.90 | ||||
| B:VAL350 | C30 | 4.34 | ||||
| B:PHE206 | Ligand | Pi-alkyl | 4.67 | |||
| C24 | 4.89 | |||||
| B:PHE210 | Ligand | 4.82 | ||||
| C28 | 3.84 | |||||
| C29 | 4.55 | |||||
| B:TYR349 | C24 | 4.63 | ||||
| B:PHE382 | Ligand | 4.60 | ||||
| C29 | 4.45 | |||||
| B:TYR386 | Ligand | 4.54 | ||||
| C19 | 4.58 | |||||
| B:TRP388 | C19 | 4.90 | ||||
| B:VAL350 | Ligand | 4.42 | ||||
| B:ALA528 | 4.37 | |||||
| δ-tocotrienol | B:VAL524 | Ligand | Hydrophobic | Alkyl | 3.81 | 89.07 |
| 4.52 | ||||||
| B:ALA528 | C12 | 3.63 | ||||
| B:LEU353 | Ligand | 5.45 | ||||
| B:VAL350 | C12 | 4.34 | ||||
| B:LEU532 | 4.66 | |||||
| B:LEU353 | Ligand | 4.84 | ||||
| B:VAL350 | C14 | 4.32 | ||||
| B:LEU353 | 3.75 | |||||
| B:LEU385 | C19 | 4.74 | ||||
| B:MET523 | 4.65 | |||||
| B:LEU535 | Ligand | 4.89 | ||||
| B:VAL345 | C24 | 5.08 | ||||
| B:VAL350 | 4.84 | |||||
| B:VAL229 | C28 | 4.45 | ||||
| B:LEU535 | 4.71 | |||||
| B:PHE206 | Ligand | Pi-alkyl | 4.84 | |||
| C28 | 5.37 | |||||
| B:PHE210 | Ligand | 5.04 | ||||
| C28 | 4.26 | |||||
| C29 | 4.39 | |||||
| B:TYR349 | C24 | 4.51 | ||||
| B:PHE382 | Ligand | 4.81 | ||||
| 5.13 | ||||||
| C29 | 4.65 | |||||
| B:TYR386 | Ligand | 4.52 | ||||
| B:TRP388 | C19 | 5.15 | ||||
| B:VAL350 | Ligand | 4.84 | ||||
| B:ALA528 | 3.55 | |||||
| B:LEU532 | 5.25 |
| Molecule | Amino Acid | Ligand Atom | Category | Types | Distance (Å) | Score |
|---|---|---|---|---|---|---|
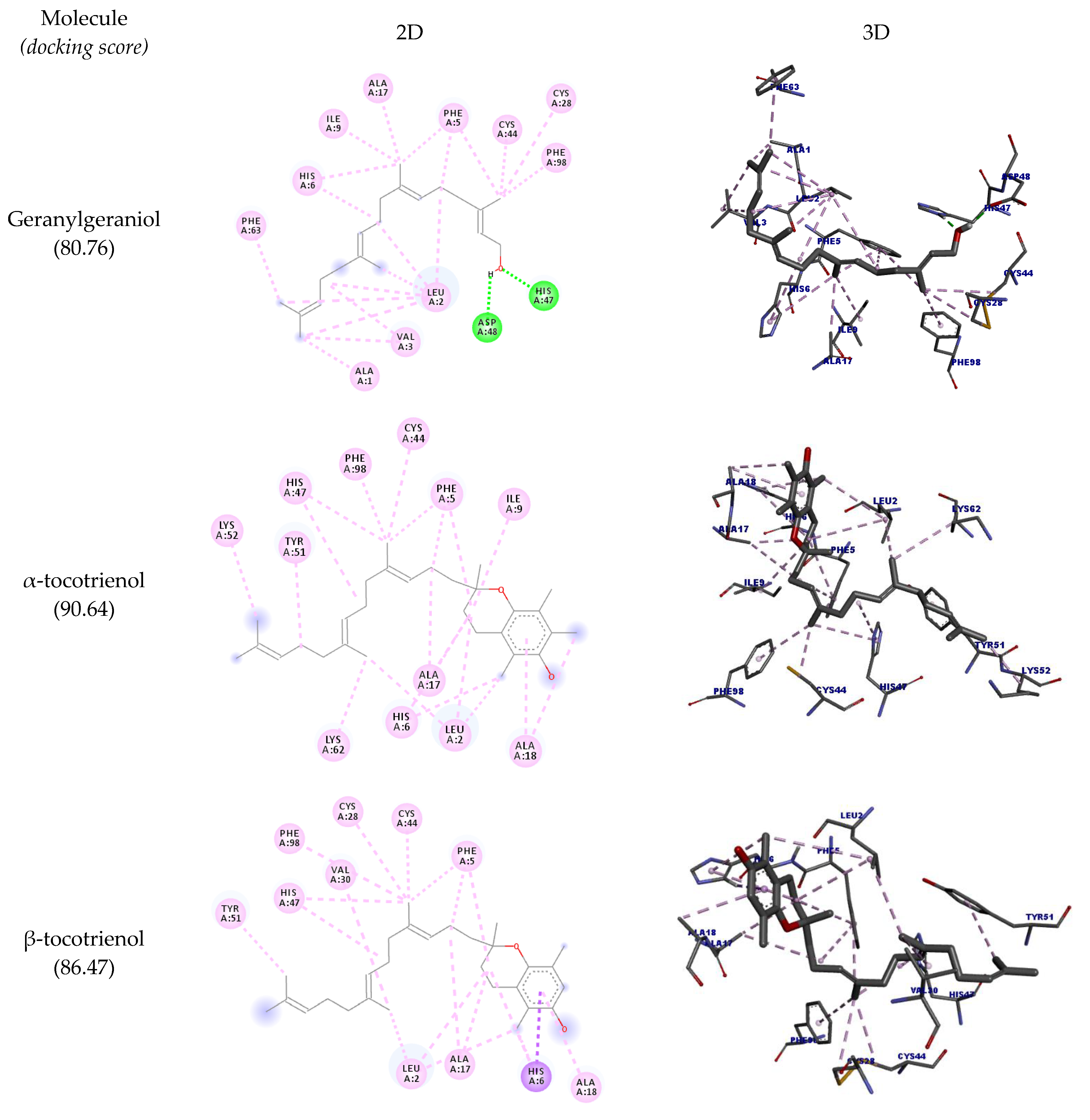
| Geranylgeraniol | HIS47 | O24 | Hydrogen bond | Conventional hydrogen bond | 1.61 | 80.76 |
| ASP48 | H55 | 1.97 | ||||
| ALA1 | C21 | Hydrophobic | Alkyl | 3.79 | ||
| VAL3 | Ligand | 4.87 | ||||
| ALA17 | C15 | 3.70 | ||||
| LEU2 | Ligand | 5.21 | ||||
| 4.34 | ||||||
| 4.89 | ||||||
| CYS28 | C14 | 4.24 | ||||
| CYS44 | 4.33 | |||||
| ILE9 | C15 | 4.98 | ||||
| LEU2 | C16 | 3.94 | ||||
| C20 | 4.71 | |||||
| C21 | 5.30 | |||||
| VAL3 | 4.51 | |||||
| PHE5 | Ligand | Pi-alkyl | 4.84 | |||
| C14 | 5.20 | |||||
| C15 | 4.31 | |||||
| HIS6 | Ligand | 4.98 | ||||
| C15 | 5.02 | |||||
| PHE63 | C20 | 4.26 | ||||
| PHE98 | C14 | 4.85 | ||||
| α-tocotrienol | ALA17 | Ligand | Hydrophobic | Alkyl | 3.99 | 90.64 |
| 5.28 | ||||||
| C13 | 4.21 | |||||
| LEU2 | Ligand | 4.98 | ||||
| ILE9 | 5.49 | |||||
| LEU2 | C11 | 4.89 | ||||
| CYS44 | C21 | 3.95 | ||||
| LEU2 | C26 | 4.39 | ||||
| LYS62 | 4.68 | |||||
| LYS52 | C30 | 3.99 | ||||
| PHE5 | Ligand | Pi-alkyl | 4.58 | |||
| PHE5 | 4.92 | |||||
| C21 | 4.59 | |||||
| HIS6 | Ligand | 5.40 | ||||
| C21 | 4.71 | |||||
| HIS47 | Ligand | 4.84 | ||||
| C21 | 4.78 | |||||
| TYR51 | Ligand | 4.82 | ||||
| PHE98 | C21 | 4.51 | ||||
| ALA18 | Ligand | 4.67 | ||||
| β-tocotrienol | HIS6 | Ligand | Hydrophobic | Pi-sigma | 2.89 | 86.47 |
| ALA17 | Alkyl | 4.15 | ||||
| 5.17 | ||||||
| LEU2 | 4.76 | |||||
| C11 | 5.48 | |||||
| CYS28 | C20 | 4.48 | ||||
| CYS44 | 3.99 | |||||
| LEU2 | C25 | 4.67 | ||||
| VAL30 | 4.90 | |||||
| PHE5 | Ligand | Pi-alkyl | 5.02 | |||
| 4.80 | ||||||
| C20 | 4.96 | |||||
| HIS6 | Ligand | 5.03 | ||||
| C11 | 4.63 | |||||
| HIS47 | Ligand | 4.58 | ||||
| C20 | 5.06 | |||||
| TYR51 | C30 | 4.52 | ||||
| PHE98 | C20 | 5.11 | ||||
| ALA18 | Ligand | 4.50 | ||||
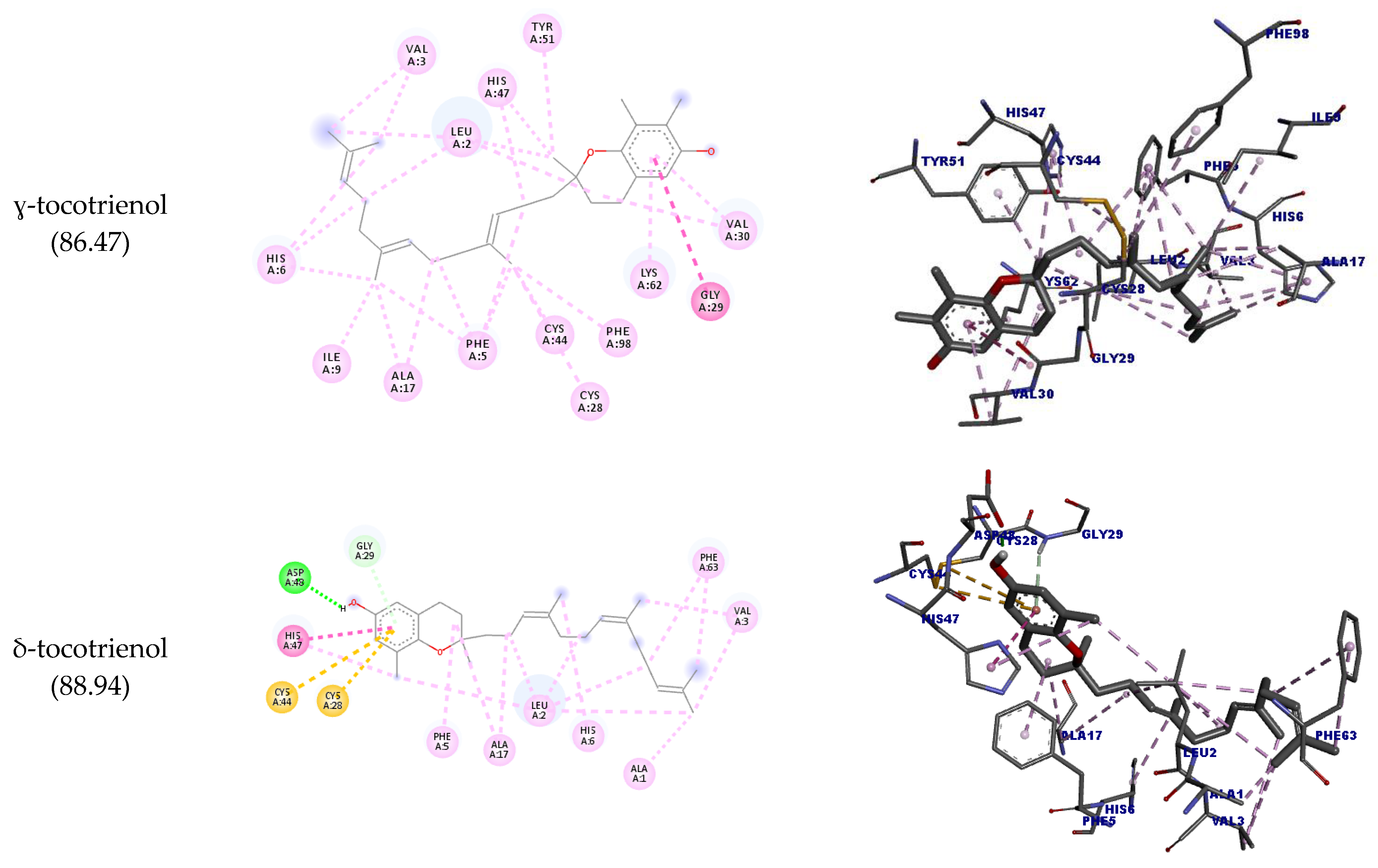
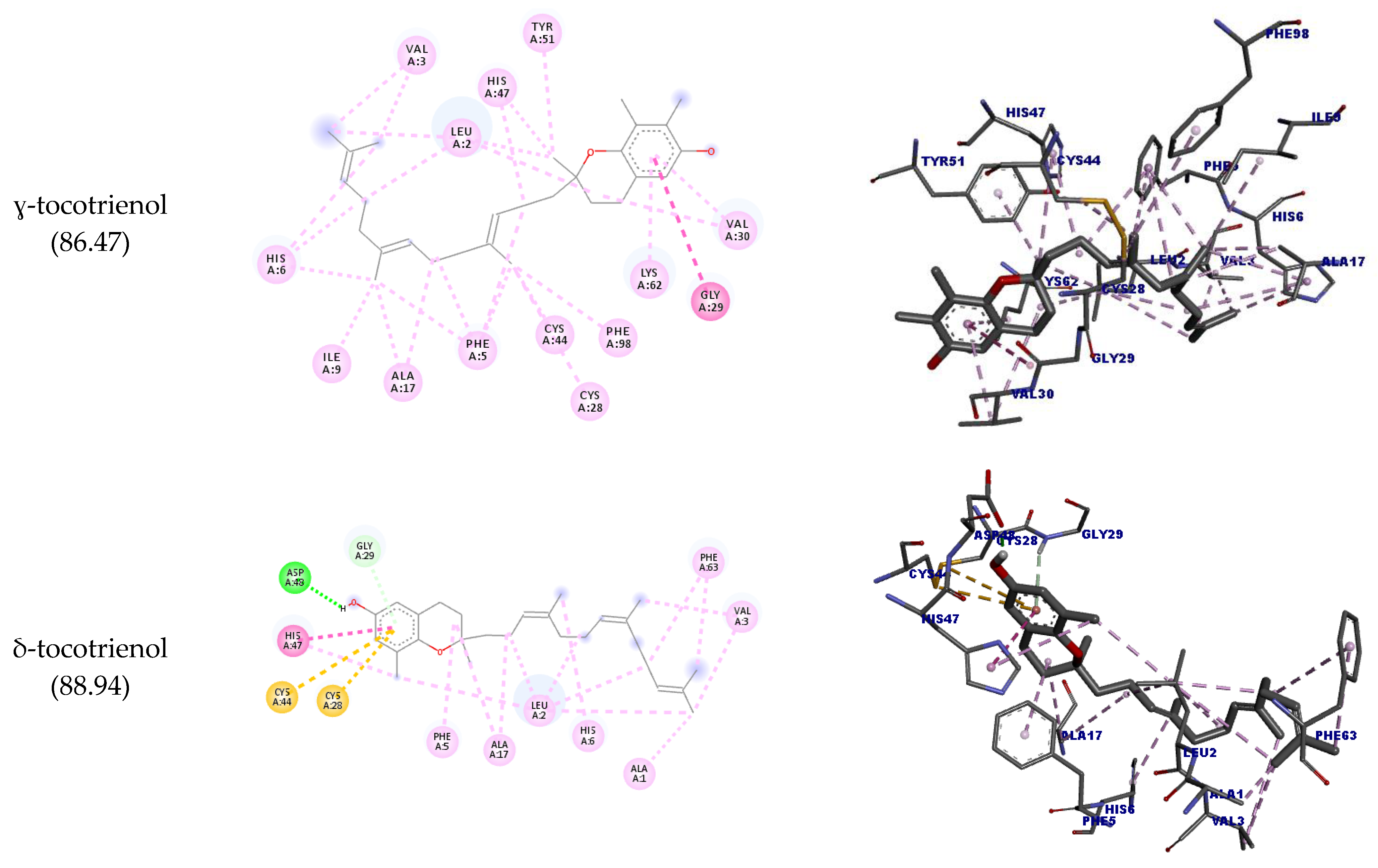
| ɣ-tocotrienol | GLY29 | Ligand | Hydrophobic | Amide-pi stacked | 4.93 | 88.94 |
| ALA17 | Alkyl | 4.79 | ||||
| C24 | 3.57 | |||||
| VAL30 | Ligand | 5.15 | ||||
| LEU2 | 5.21 | |||||
| C14 | 4.57 | |||||
| CYS28 | C19 | 4.35 | ||||
| CYS44 | 4.29 | |||||
| LEU2 | Ligand | 3.82 | ||||
| ILE9 | C24 | 4.82 | ||||
| LEU2 | C28 | 5.24 | ||||
| VAL3 | 4.48 | |||||
| C29 | 4.61 | |||||
| PHE5 | Ligand | Pi-alkyl | 5.18 | |||
| 5.39 | ||||||
| C19 | 5.11 | |||||
| C24 | 4.28 | |||||
| HIS6 | Ligand | 5.43 | ||||
| C24 | 4.96 | |||||
| C29 | 4.33 | |||||
| HIS47 | Ligand | 4.47 | ||||
| C14 | 4.72 | |||||
| TYR51 | 4.12 | |||||
| PHE98 | C19 | 5.09 | ||||
| VAL30 | Ligand | 4.31 | ||||
| LYS62 | 4.8 | |||||
| δ-tocotrienol | ASP48 | H36 | Hydrogen bond | Conventional hydrogen bond | 1.71 | 87.82 |
| GLY29 | Ligand | Pi-donor hydrogen bond | 2.93 | |||
| CYS28 | Other | Pi-sulfur | 5.93 | |||
| CYS44 | 4.86 | |||||
| HIS47 | Hydrophobic | Pi-pi T-shaped | 4.78 | |||
| ALA1 | C29 | Alkyl | 3.27 | |||
| ALA17 | Ligand | 4.90 | ||||
| 4.89 | ||||||
| LEU2 | C12 | 4.11 | ||||
| Ligand | 4.23 | |||||
| 4.79 | ||||||
| 4.36 | ||||||
| VAL3 | C24 | 4.77 | ||||
| LEU2 | C29 | 4.43 | ||||
| VAL3 | 4.34 | |||||
| PHE5 | Ligand | Pi-alkyl | 4.52 | |||
| HIS6 | C19 | 4.64 | ||||
| HIS47 | C12 | 5.20 | ||||
| PHE63 | Ligand | 4.95 | ||||
| C28 | 4.69 |
| Molecule | PreADMET | |||||||
|---|---|---|---|---|---|---|---|---|
| Absorption | Distribution | Absorption | Distribution | |||||
| %HIA | Caco-2 (nm/sec) | MDCK (nm/sec) | BPB% | BBB (Cbrain/Cblood) | GI absorption | BBB | P-gp | |
| Geranylgeraniol | 100 | 37.1 | 62.05 | 100 | 17.58 | High | No | No |
| α-tocotrienol | 97.91 | 29.13 | 21.78 | 100 | 19.21 | Low | No | Yes |
| β-tocotrienol | 97.9 | 27.94 | 24.31 | 100 | 19.01 | Low | No | Yes |
| γ-tocotrienol | 97.9 | 27.94 | 24.31 | 100 | 18.99 | Low | No | Yes |
| δ-tocotrienol | 97.89 | 26.83 | 27.42 | 100 | 18.83 | Low | No | Yes |
| Molecule | Toxicity Class | Predicted DL50 | Toxicity Type | Prediction | Probability |
|---|---|---|---|---|---|
| Geranylgeraniol | 5 | 5000 mg/kg | Hepatotoxicity | Inactive | 0.79 |
| Carcinogenicity | Inactive | 0.76 | |||
| Immunotoxicity | Inactive | 0.99 | |||
| Mutagenicity | Inactive | 0.97 | |||
| Cytotoxicity | Inactive | 0.85 | |||
| α-tocotrienol | 4 | 500 mg/kg | Hepatotoxicity | Inactive | 0.93 |
| Carcinogenicity | Inactive | 0.77 | |||
| Immunotoxicity | Inactive | 0.89 | |||
| Mutagenicity | Inactive | 0.92 | |||
| Cytotoxicity | Inactive | 0.87 | |||
| β-tocotrienol | 4 | 500 mg/kg | Hepatotoxicity | Inactive | 0.93 |
| Carcinogenicity | Inactive | 0.77 | |||
| Immunotoxicity | Inactive | 0.79 | |||
| Mutagenicity | Inactive | 0.92 | |||
| Cytotoxicity | Inactive | 0.87 | |||
| γ-tocotrienol | 4 | 500 mg/kg | Hepatotoxicity | Inactive | 0.93 |
| Carcinogenicity | Inactive | 0.77 | |||
| Immunotoxicity | Inactive | 0.61 | |||
| Mutagenicity | Inactive | 0.92 | |||
| Cytotoxicity | Inactive | 0.87 | |||
| δ-tocotrienol | 4 | 500 mg/kg | Hepatotoxicity | Inactive | 0.94 |
| Carcinogenicity | Inactive | 0.79 | |||
| Immunotoxicity | Inactive | 0.93 | |||
| Mutagenicity | Inactive | 0.91 | |||
| Cytotoxicity | Inactive | 0.86 |
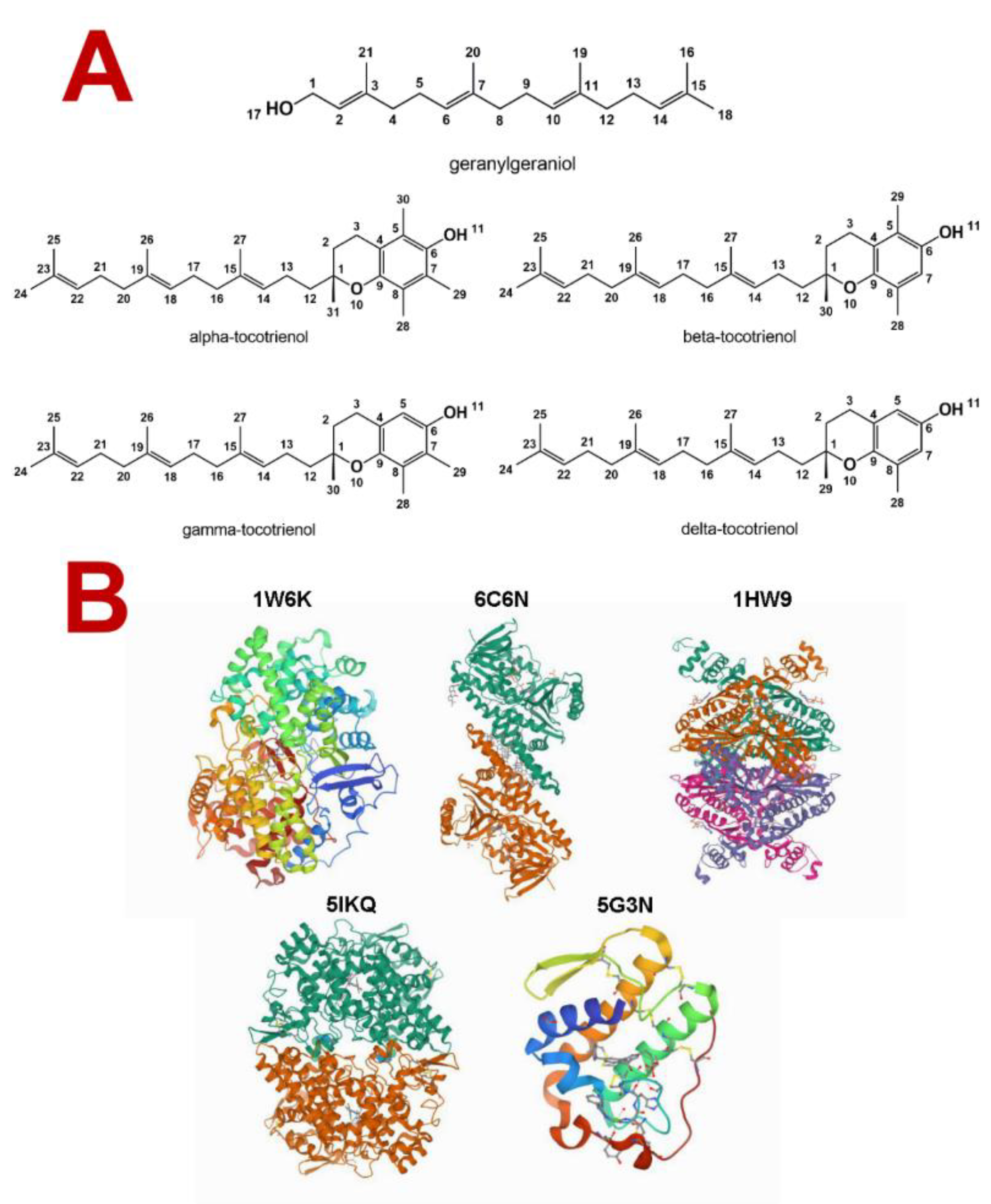
| Molecule | PDB ID | Resolution (Å) | RMSD (Å) | Docking Radius (Å) | x, y, z Coordinates |
|---|---|---|---|---|---|
| Lanosterol synthase (OSC) | 1W6K | 2.1 | 0.622 | 11.49 | 28.79, 69.02, 8.45 |
| Squalene epoxidase (SQLE) | 6C6N | 2.3 | 1.038 | 15.08 | −23.75, 92.76, 63.37 |
| HMG-CoA reductase (HMGR) | 1HW9 | 2.3 | 1.482 | 8.41 | 2.31, −8.29, −9.21 |
| Cyclooxygenase-2 (COX-2) | 5IKQ | 2.4 | 0.507 | 8.867 | 16.06, 43.11, 60.99 |
| Phospholipase A2 (sPLA2) | 5G3N | 1.8 | 0.507 | 9.132 | 7.48, 3.41, −0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batista, M.A.; de Lima Teixeira dos Santos, A.V.T.; do Nascimento, A.L.; Moreira, L.F.; Souza, I.R.S.; da Silva, H.R.; Pereira, A.C.M.; da Silva Hage-Melim, L.I.; Carvalho, J.C.T. Potential of the Compounds from Bixa orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols. Molecules 2022, 27, 1584. https://doi.org/10.3390/molecules27051584
Batista MA, de Lima Teixeira dos Santos AVT, do Nascimento AL, Moreira LF, Souza IRS, da Silva HR, Pereira ACM, da Silva Hage-Melim LI, Carvalho JCT. Potential of the Compounds from Bixa orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols. Molecules. 2022; 27(5):1584. https://doi.org/10.3390/molecules27051584
Chicago/Turabian StyleBatista, Mateus Alves, Abrahão Victor Tavares de Lima Teixeira dos Santos, Aline Lopes do Nascimento, Luiz Fernando Moreira, Indira Ramos Senna Souza, Heitor Ribeiro da Silva, Arlindo César Matias Pereira, Lorane Izabel da Silva Hage-Melim, and José Carlos Tavares Carvalho. 2022. "Potential of the Compounds from Bixa orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols" Molecules 27, no. 5: 1584. https://doi.org/10.3390/molecules27051584
APA StyleBatista, M. A., de Lima Teixeira dos Santos, A. V. T., do Nascimento, A. L., Moreira, L. F., Souza, I. R. S., da Silva, H. R., Pereira, A. C. M., da Silva Hage-Melim, L. I., & Carvalho, J. C. T. (2022). Potential of the Compounds from Bixa orellana Purified Annatto Oil and Its Granules (Chronic®) against Dyslipidemia and Inflammatory Diseases: In Silico Studies with Geranylgeraniol and Tocotrienols. Molecules, 27(5), 1584. https://doi.org/10.3390/molecules27051584