
Impact of Co-Delivery of EGCG and Tuna Oil within a Broccoli Matrix on Human Gut Microbiota, Phenolic Metabolites and Short Chain Fatty Acids In Vitro



Abstract
:1. Introduction
2. Results and Discussion
2.1. pH Value and Short-Chain Fatty Acids (SCFA) Changes
2.2. Transformation of EGCG during In Vitro Microbiota Fermentation
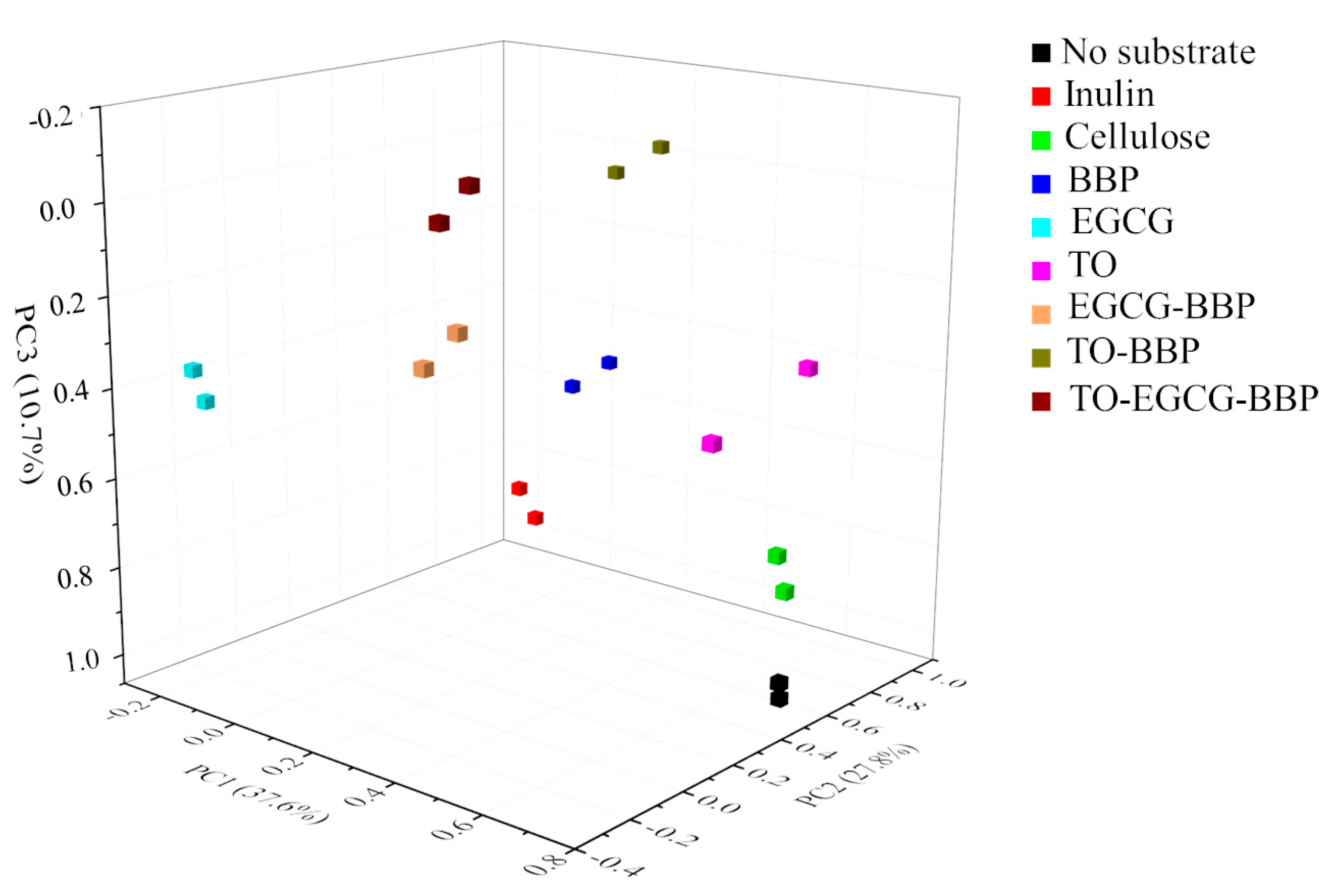
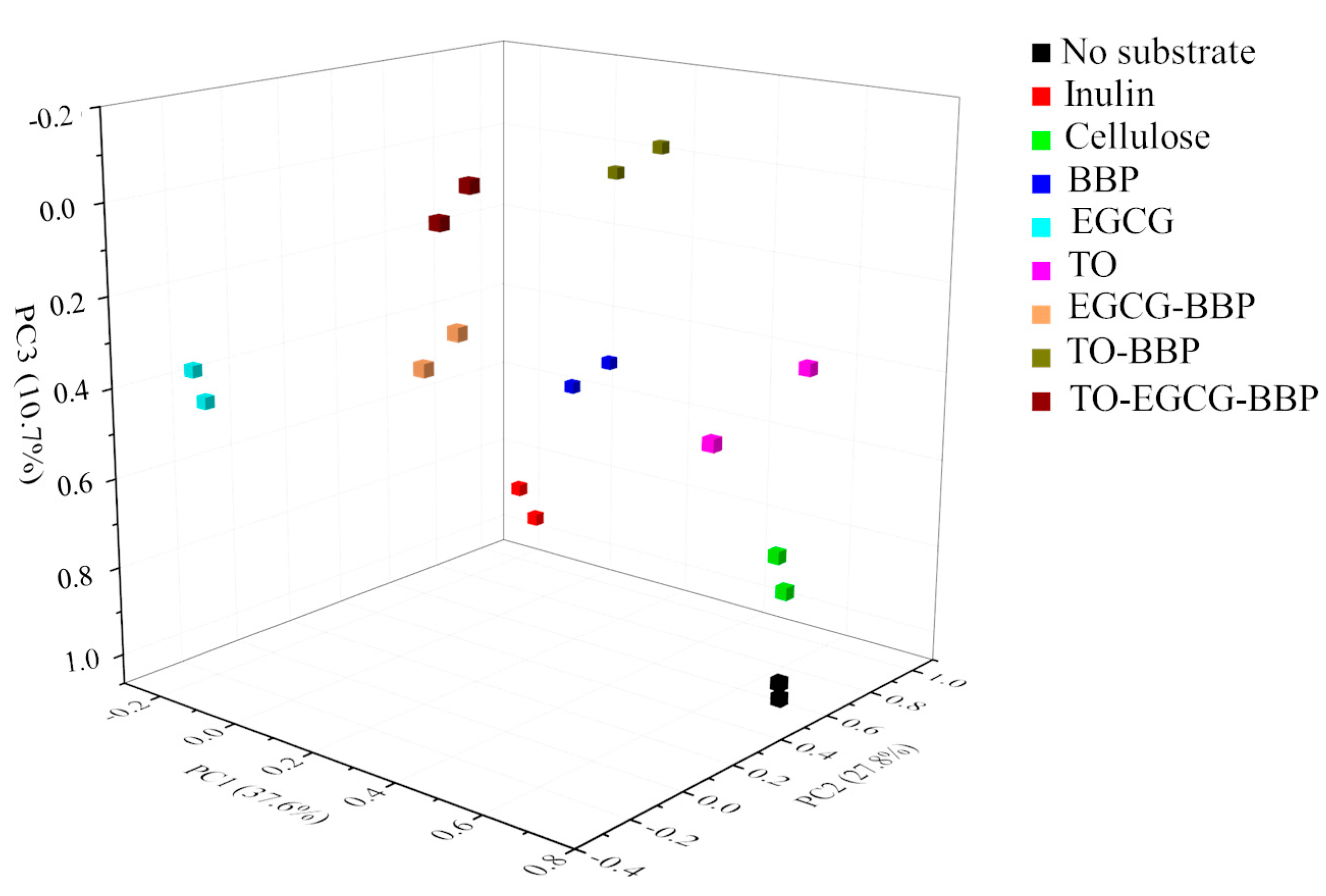
2.3. Microbiota Population Changes
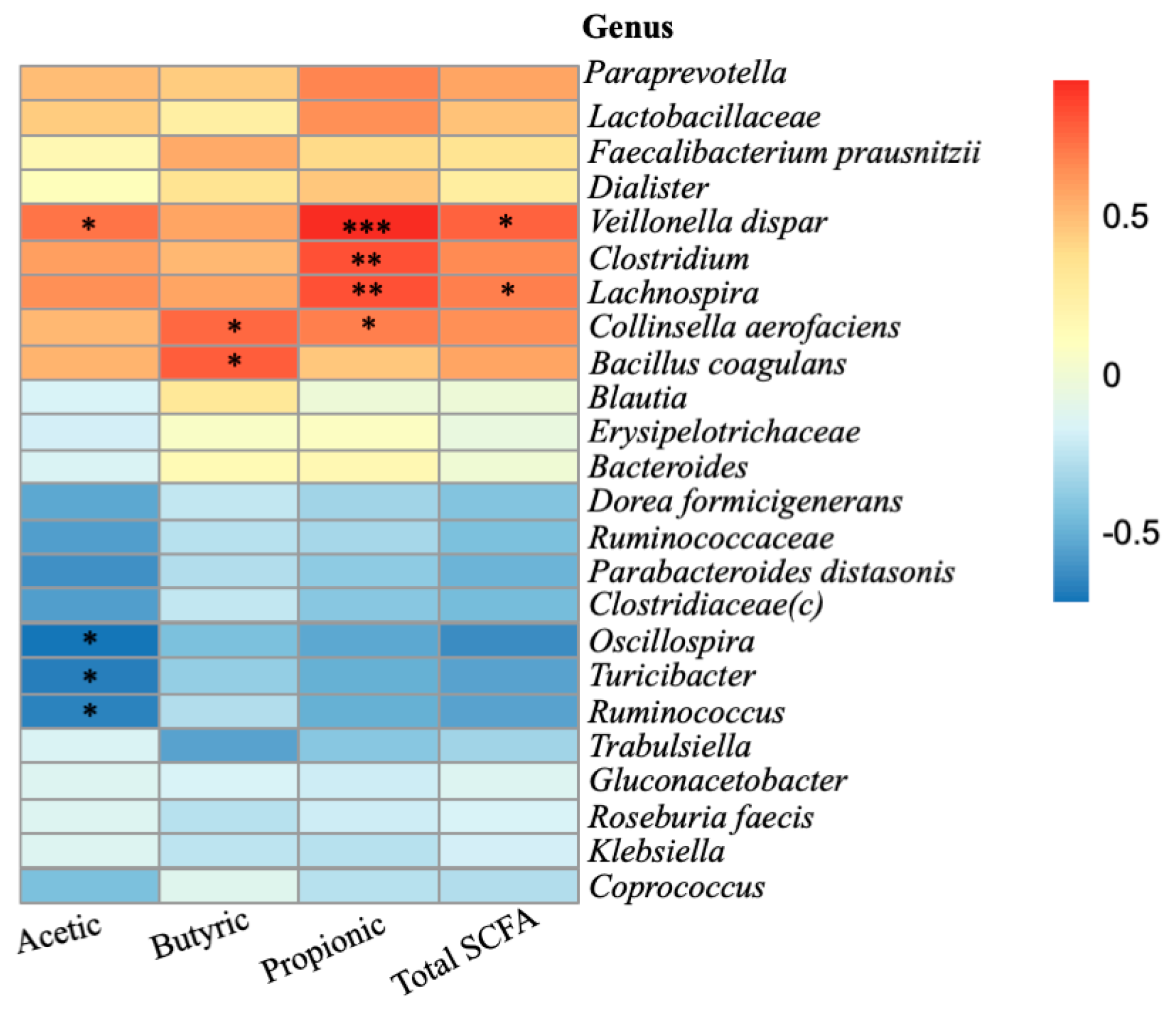
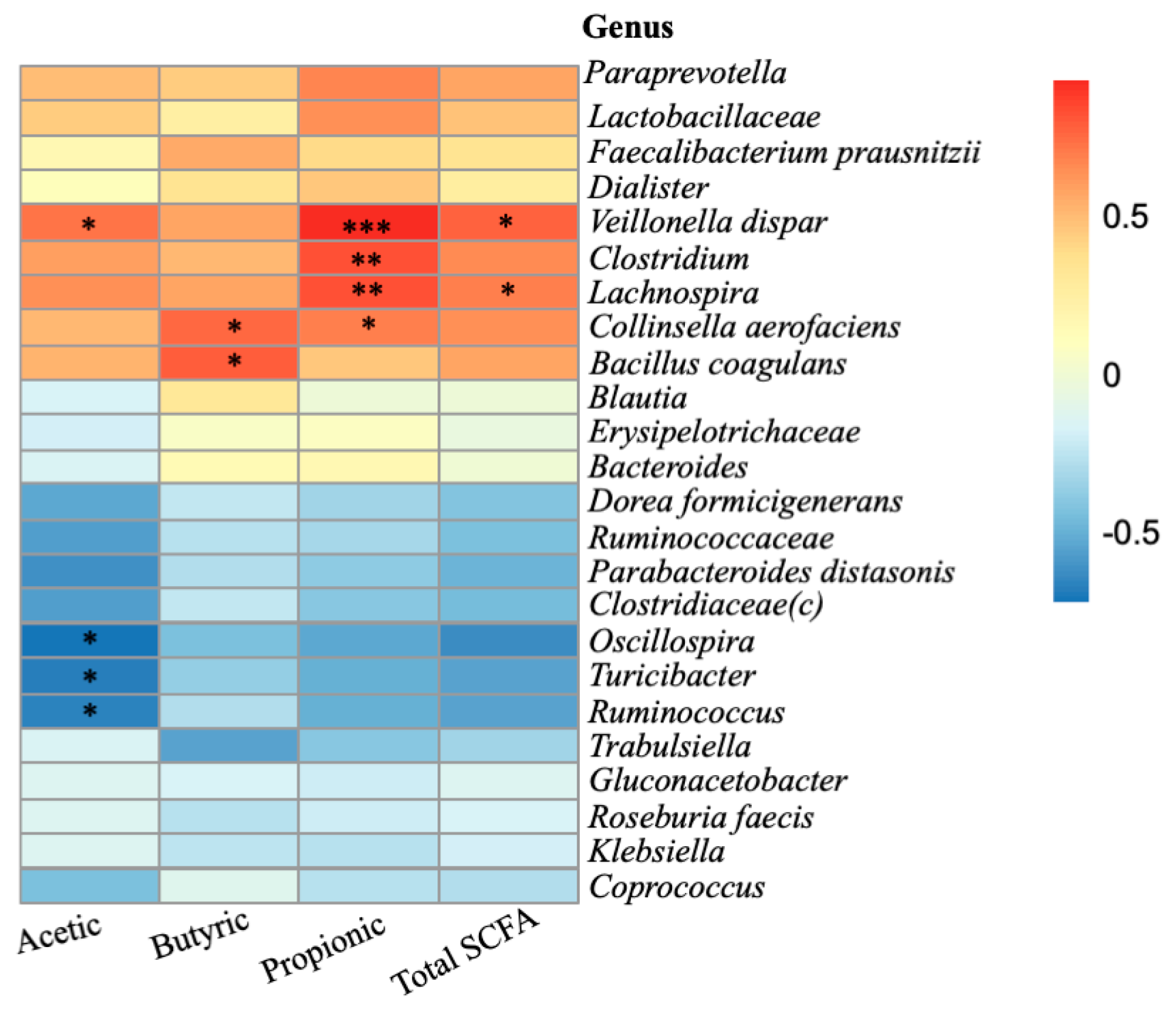
2.4. Relationship between Microbiota Composition and SCFA Formation
3. Materials and Methods
3.1. Material and Chemicals
3.2. Preparation of Formulations
3.3. In Vitro Gut Microbiota Fermentation
3.3.1. Preparation of the Fermentation Medium
3.3.2. Fresh Fecal Inoculum
3.3.3. Anaerobic Fermentation
3.4. Phenolic Compounds Analysis
3.5. Short Chain Fatty Acids (SCFA) Analysis
3.6. DNA Extraction and 16 S Gene Sequencing
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Paturi, G.; Mandimika, T.; Butts, C.A.; Zhu, S.; Roy, N.C.; McNabb, W.C.; Ansell, J. Influence of dietary blueberry and broccoli on cecal microbiota activity and colon morphology in mdr1a−/− mice, a model of inflammatory bowel diseases. Nutrition 2012, 28, 324–330. [Google Scholar] [CrossRef]
- Kaczmarek, J.L.; Liu, X.; Charron, C.S.; Novotny, J.A.; Jeffery, E.H.; Seifried, H.E.; Ross, S.A.; Miller, M.J.; Swanson, K.S.; Holscher, H.D. Broccoli consumption affects the human gastrointestinal microbiota. J. Nutr. Biochem. 2019, 63, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zandani, G.; Kaftori-Sandler, N.; Sela, N.; Nyska, A.; Madar, Z. Dietary broccoli improves markers associated with glucose and lipid metabolism through modulation of gut microbiota in mice. Nutrition 2021, 90, 111240. [Google Scholar] [CrossRef]
- Shi, M.; Ying, D.Y.; Ye, J.H.; Sanguansri, L.; Augustin, M.A. Broccoli byproducts for protection and co-delivery of EGCG and tuna oil. Food Chem. 2020, 326, 126963. [Google Scholar] [CrossRef]
- Unno, T.; Sakuma, M.; Mitsuhashi, S. Effect of dietary supplementation of (−)-epigallocatechin gallate on gut microbiota and biomarkers of colonic fermentation in rats. J. Nutr. Sci. Vitaminol. 2014, 60, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhu, X.; Sun, Y.; Hu, B.; Sun, Y.; Jabbar, S.; Zeng, X. Fermentation in vitro of EGCG, GCG and EGCG3” Me isolated from Oolong tea by human intestinal microbiota. Food Res. Int. 2013, 54, 1589–1595. [Google Scholar] [CrossRef]
- Liu, Z.; Bruins, M.E.; de Bruijn, W.J.; Vincken, J.P. A comparison of the phenolic composition of old and young tea leaves reveals a decrease in flavanols and phenolic acids and an increase in flavonols upon tea leaf maturation. J. Food Compos. Anal. 2020, 86, 103385. [Google Scholar] [CrossRef]
- Naito, Y.; Ushiroda, C.; Mizushima, K.; Inoue, R.; Yasukawa, Z.; Abe, A.; Takagi, T. Epigallocatechin-3-gallate (EGCG) attenuates non-alcoholic fatty liver disease via modulating the interaction between gut microbiota and bile acids. J. Clin. Biochem. Nutr. 2020, 67, 2–9. [Google Scholar] [CrossRef]
- Shi, M.; Ying, D.Y.; Hlaing, M.M.; Ye, J.H.; Sanguansri, L.; Augustin, M.A. Development of broccoli by-products as carriers for delivering EGCG. Food Chem. 2019, 301, 125301. [Google Scholar] [CrossRef]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of omega-3 fatty acids on the gut microbiota. Int. J. Mol. Sci. 2017, 18, 2645. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Hutkins, R.W.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastro. Hepat. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoensiddhi, S.; Conlon, M.A.; Vuaran, M.S.; Franco, C.M.; Zhang, W. Impact of extraction processes on prebiotic potential of the brown seaweed Ecklonia radiata by in vitro human gut bacteria fermentation. J. Funct. Foods 2016, 24, 221–230. [Google Scholar] [CrossRef]
- Ramos-Romero, S.; Hereu, M.; Molinar-Toribio, E.; Almajano, M.P.; Méndez, L.; Medina, I.; Taltavull, N.; Romeu, M.; Nogués, M.R.; Torres, J.L. Effects of the combination of ω-3 PUFAs and proanthocyanidins on the gut microbiota of healthy rats. Food Res. Int. 2017, 97, 364–371. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [Green Version]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, J.; Wei, Q.; Liu, Y.; Zhang, H.; Chen, X.; Xu, K. Epigallocatechin-3-O-gallate modulates the diversity of gut microbiota in ovariectomized rats. Food Sci. Nutr. 2020, 8, 1295–1302. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Bindels, L.B.; Porporato, P.; Dewulf, E.; Verrax, J.; Neyrinck, A.M.; Martin, J.; Scott, K.; Calderon, P.B.; Feron, O.; Muccioli, G. Gut microbiota-derived propionate reduces cancer cell proliferation in the liver. Br. J. Cancer 2012, 107, 1337. [Google Scholar] [CrossRef] [Green Version]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R. Butyrate, neuroepigenetics and the gut microbiome: Can a high fiber diet improve brain health? Neurosci. Lett. 2016, 625, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Bourquin, L.D.; Titgemeyer, E.C.; Fahey, G.C., Jr. Vegetable fiber fermentation by human fecal bacteria: Cell wall polysaccharide disappearance and short-chain fatty acid production during in vitro fermentation and water-holding capacity of unfermented residues. J. Nutr. 1993, 123, 860–869. [Google Scholar] [CrossRef]
- Sun, H.; Chen, Y.; Cheng, M.; Zhang, X.; Zheng, X.; Zhang, Z. The modulatory effect of polyphenols from green tea, oolong tea and black tea on human intestinal microbiota in vitro. J. Food Sci. Tech. 2018, 55, 399–407. [Google Scholar] [CrossRef]
- Takagaki, A.; Nanjo, F. Metabolism of (−)-epigallocatechin gallate by rat intestinal flora. J. Agric. Food Chem. 2009, 58, 1313–1321. [Google Scholar] [CrossRef]
- Chen, H.D.; Sang, S.M. Biotransformation of tea polyphenols by gut microbiota. J Funct. Foods 2014, 7, 26–42. [Google Scholar] [CrossRef]
- Roowi, S.; Stalmach, A.; Mullen, W.; Lean, M.E.J.; Edwards, C.A.; Crozier, A. Green Tea Flavan-3-ols: Colonic Degradation and Urinary Excretion of Catabolites by Humans. J. Agric. Food Chem. 2010, 58, 1296–1304. [Google Scholar] [CrossRef]
- van’t Slot, G.; Humpf, H.U. Degradation and metabolism of catechin, epigallocatechin-3-gallate (EGCG), and related compounds by the intestinal microbiota in the pig cecum model. J. Agric. Food Chem. 2009, 57, 8041–8048. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Pae, M.; Ren, Z.; Meydani, M.; Shang, F.; Smith, D.; Meydani, S.N.; Wu, D. Dietary supplementation with high dose of epigallocatechin-3-gallate promotes inflammatory response in mice. J. Nutr. Biochem. 2012, 23, 526–531. [Google Scholar] [CrossRef]
- Winter, S.E.; Winter, M.G.; Xavier, M.N.; Thiennimitr, P.; Poon, V.; Keestra, A.M.; Laughlin, R.C.; Gomez, G.; Wu, J.; Lawhon, S.D. Host-derived nitrate boosts growth of E. coli in the inflamed gut. Science 2013, 339, 708–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic characteristics of Bacillus coagulans and associated implications for human health and diseases. J. Funct. Foods 2020, 64, 103643. [Google Scholar] [CrossRef]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef]
- Santoru, M.L.; Piras, C.; Murgia, A.; Palmas, V.; Camboni, T.; Liggi, S.; Ibba, I.; Lai, M.A.; Orrù, S.; Blois, S. Cross sectional evaluation of the gut-microbiome metabolome axis in an Italian cohort of IBD patients. Sci. Rep. 2017, 7, 9523. [Google Scholar] [CrossRef]
- Sakanaka, S.; Juneja, L.R.; Taniguchi, M. Antimicrobial effects of green tea polyphenols on thermophilic spore-forming bacteria. J. Biosci. Bioeng. 2000, 90, 81–85. [Google Scholar] [CrossRef]
- Leal, J.M.; Suárez, L.V.; Jayabalan, R.; Oros, J.H.; Escalante-Aburto, A. A review on health benefits of kombucha nutritional compounds and metabolites. J. Food 2018, 16, 390–399. [Google Scholar]
- Yang, Z.W.; Ji, B.P.; Zhou, F.; Li, B.; Luo, Y.; Yang, L.; Li, T. Hypocholesterolaemic and antioxidant effects of kombucha tea in high-cholesterol fed mice. J. Sci. Food Agr. 2009, 89, 150–156. [Google Scholar] [CrossRef]
- Vina, I.; Semjonovs, P.; Linde, R.; Patetko, A. Glucuronic acid containing fermented functional beverages produced by natural yeasts and bacteria associations. Int. J. Res. Rev. Appl. 2013, 14, 17–25. [Google Scholar]
- Del Chierico, F.; Cardile, S.; Pietrobattista, A.; Liccardo, D.; Russo, A.; Candusso, M.; Basso, M.; Grimaldi, C.; Pansani, L.; Bernaschi, P. Liver Transplantation and Gut Microbiota Profiling in a Child Colonized by a Multi-Drug Resistant Klebsiella pneumoniae: A New Approach to Move from Antibiotic to “Eubiotic” Control of Microbial Resistance. Int. J. Mol. Sci. 2018, 19, 1280. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, D.; Chatterjee, T.K.; Dasgupta, A.; Lourduraja, J.; Dastidar, S.G. In vitro and in vivo antimicrobial action of tea: The commonest beverage of Asia. Bio. Pharm. Bull. 2005, 28, 2125–2127. [Google Scholar] [CrossRef] [Green Version]
- Yoda, Y.; Hu, Z.Q.; Zhao, W.-H.; Shimamura, T. Different susceptibilities of Staphylococcus and Gram-negative rods to epigallocatechin gallate. J. Infect. Chemother. 2004, 10, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Wakita, Y.; Shimomura, Y.; Kitada, Y.; Yamamoto, H.; Ohashi, Y.; Matsumoto, M. Taxonomic classification for microbiome analysis, which correlates well with the metabolite milieu of the gut. BMC Microbiol. 2018, 18, 188. [Google Scholar] [CrossRef] [Green Version]
- George, N.S.; Cheung, L.; Luthria, D.L.; Santin, M.; Dawson, H.D.; Bhagwat, A.A.; Smith, A.D. Pomegranate peel extract alters the microbiome in mice and dysbiosis caused by Citrobacter rodentium infection. Food Sci. Nutr. 2019, 7, 2565–2576. [Google Scholar] [CrossRef] [Green Version]
- McWhorter, A.; Haddock, R.; Nocon, F.A.; Steigerwalt, A.G.; Brenner, D.; Aleksić, S.; Bockemühl, J.; Farmer, J. Trabulsiella guamensis, a new genus and species of the family Enterobacteriaceae that resembles Salmonella subgroups 4 and 5. J. Clin. Microbiol. 1991, 29, 1480–1485. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, J.A.; Farmer, J., III. Trabulsiella. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: New York, NY, USA, 2015; pp. 1–12. [Google Scholar]
- Fukudo, S.; Kanazawa, M. Gene, environment, and brain-gut interactions in irritable bowel syndrome. J. Gastroen. Hepatol. 2011, 26, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Tuncil, Y.E.; Nakatsu, C.H.; Kazem, A.E.; Arioglu-Tuncil, S.; Reuhs, B.; Martens, E.C.; Hamaker, B.R. Delayed utilization of some fast-fermenting soluble dietary fibers by human gut microbiota when presented in a mixture. J. Funct. Foods 2017, 32, 347–357. [Google Scholar] [CrossRef]
- Salyers, A.A.; Leedle, J.A. Carbohydrate metabolism in the human colon. In Human Intestinal Microflora in Health and Disease; Academic Press: New York, NY, USA, 1983; pp. 129–146. [Google Scholar]
- Qin, P.; Zou, Y.; Dai, Y.; Luo, G.; Zhang, X.; Xiao, L. Characterization a Novel Butyric Acid-Producing Bacterium Collinsella aerofaciens Subsp. Shenzhenensis Subsp. Nov. Microorg. 2019, 7, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majeed, M.; Majeed, S.; Nagabhushanam, K.; Arumugam, S.; Beede, K.; Ali, F. Evaluation of the in vitro cholesterol-lowering activity of the probiotic strain Bacillus coagulans MTCC 5856. Int. Food Sci. Tech. 2019, 54, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Li, K.; Xiang, Y.; Zhou, W.; Gui, G.; Yang, H. The fecal microbiota composition of boar Duroc, Yorkshire, Landrace and Hampshire pigs. Asian-Australas. J. Anim. Sci. 2017, 30, 1456. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Tanwar, J.; Hameed, S.; Fatima, Z.; Manesar, G. Antimicrobial potential of epigallocatechin-3-gallate (EGCG): A green tea polyphenol. J. Biochem. Pharmacol. Res. 2014, 2, 167–174. [Google Scholar]
- Kuzuhara, T.; Sei, Y.; Yamaguchi, K.; Suganuma, M.; Fujiki, H. DNA and RNA as new binding targets of green tea catechins. J. Biol. Chem. 2016, 281, 17446–17456. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Substrates | pH at 24 h 2 | Total SCFA 3 (µmol/mL) | Acetic Acid Amount (µmol/mL) | Propionic Acid Amount (µmol/mL) | Butyric Acid Amount (µmol/mL) | ||||
|---|---|---|---|---|---|---|---|---|---|
| T = 0 h | T = 24 h | T = 0 h | T = 24 h | T = 0 h | T = 24 h | T = 0 h | T = 24 h | ||
| No substrate | 6.8 | 1.72 ± 0.21 a | 21.99 ± 2.07 d | 1.21 ± 0.13 a | 13.99 ± 1.42 f | 0.23 ± 0.03 a | 2.53 ± 0.21 d | 0.22 ± 0.03 b | 2.83 ± 0.21 e |
| Inulin | 4.2 | 2.21 ± 0.64 a | 42.80 ± 2.56 b | 1.54 ± 0.42 a | 26.41 ± 1.50 c | 0.31 ± 0.10 a | 6.37 ± 0.53 c | 0.30 ± 0.09 ab | 9.62 ± 0.49 a |
| Cellulose | 6.4 | 2.21 ± 1.22 a | 21.35 ± 1.47 d | 1.52 ± 0.79 a | 13.06 ± 0.98 f | 0.32 ± 0.22 a | 2.63 ± 0.16 d | 0.30 ± 0.19 ab | 3.16 ± 0.19 de |
| BBP | 5.2 | 2.52 ± 0.69 a | 49.16 ± 2.18 a | 1.45 ± 0.20 a | 31.78 ± 1.35 a | 0.40 ± 0.18 a | 11.27 ± 0.46 a | 0.43 ± 0.20 a | 5.87 ± 0.35 c |
| TO | 6.5 | 1.56 ± 0.25 b | 25.48 ± 0.87 c | 1.03 ± 0.14 a | 15.98 ± 0.46 e | 0.21 ± 0.04 a | 3.26 ± 0.20 d | 0.21 ± 0.04 b | 3.64 ± 0.10 d |
| EGCG | 6.5 | 1.40 ± 0.12 b | 4.48 ± 0.69 e | 1.01 ± 0.06 a | 3.26 ± 0.41 g | 0.18 ± 0.01 b | 0.59 ± 0.09 e | 0.17 ± 0.01 b | 0.41 ± 0.11 g |
| EGCG-BBP | 6.1 | 1.57 ± 0.06 b | 25.20 ± 1.63 c | 1.13 ± 0.04 a | 20.06 ± 0.64 d | 0.19 ± 0.00 b | 3.14 ± 0.67 d | 0.18 ± 0.00 b | 1.81 ± 0.25 f |
| TO-BBP | 5.4 | 1.78 ± 0.05 a | 48.31 ± 1.22 a | 1.33 ± 0.03 a | 29.75 ± 0.56 b | 0.21 ± 0.01 b | 9.89 ± 0.35 b | 0.19 ± 0.00 b | 8.38 ± 0.28 b |
| TO-EGCG-BBP | 6.1 | 1.56 ± 0.03 b | 22.71 ± 1.44 c,d | 1.15 ± 0.02 a | 18.49 ± 0.70 d | 0.18 ± 0.00 b | 2.62 ± 0.56 d | 0.17 ± 0.00 b | 1.50 ± 0.16 f |
| Sample | Initial Concentration of EGCG | After 24 h Fermentation | |||
|---|---|---|---|---|---|
| EGCG | Gallic Acid | EGC | GCG | ||
| EGCG | 15.00 ± 0.12 a (100%) | 7.83 ± 1.01 a (52.2%) | 0.17 ± 0.02 a | 0.37 ± 0.01 a | 1.31 ± 0.23 a |
| EGCG-BBP | 3.77 ± 0.04 b (100%) | 0.69 ± 0.05 b (18.3%) | 0.05 ± 0.01 b | NA | NA |
| TO-EGCG-BBP | 3.03 ± 0.02 c (100%) | 1.02 ± 0.14 b (33.7%) | 0.05 ± 0.01 b | 0.05 ± 0.01 b | 0.08 ± 0.00 b |
| No Substrate | Inulin | Cellulose | BBP | EGCG | TO | EGCG-BBP | TO-BBP | TO-EGCG-BBP | |
|---|---|---|---|---|---|---|---|---|---|
| Firmicutes | 75.3 ± 0.5 | 91.1 ± 1.2 | 74.9 ± 0.9 | 88.5 ± 4.2 | 33.1 ± 0.5 | 64.9 ± 9.0 | 13.5 ± 0.1 | 82.9 ± 3.3 | 13.7 ± 0.3 |
| Bacteroidetes | 14.9 ± 0.7 | 6.5 ± 0.8 | 13.7 ± 0.5 | 9.4 ± 3.9 | 8.3 ± 1.2 | 14.6 ± 0.6 | 3.3 ± 0.3 | 14.9 ± 3.1 | 6.1 ± 0.4 |
| Proteobacteria | 8.4 ± 0.9 | 1.4 ± 0.3 | 9.7 ± 0.3 | 0.9 ± 0.0 | 57.7 ± 1.6 | 19.6 ± 9.8 | 82.9 ± 0.3 | 1.2 ± 0.0 | 80.1 ± 0.2 |
| Others | 1.4 ± 0.3 | 1.1 ± 0.1 | 1.7 ± 0.1 | 1.2 ± 0.4 | 0.9 ± 0.1 | 1.0 ± 0.3 | 0.3 ± 0.1 | 1.0 ± 0.2 | 0.2 ± 0.0 |
| No Substrate | Inulin | Cellulose | BBP | EGCG | TO | EGCG-BBP | TO-BBP | TO-EGCG-BBP | |
|---|---|---|---|---|---|---|---|---|---|
| Coriobacteriaceae (Collinsella aerofaciens) | 0.45 ± 0.0007 | 0.90 ± 0.0000 | 0.75 ± 0.0007 | 0.95 ± 0.0035 | 0.25 ± 0.0007 | 0.40 ± 0.0014 | 0.15 ± 0.0007 | 0.70 ± 0.0014 | 0.05 ± 0.0007 |
| Bacteroidaceae (Bacteroides) | 10.30 ± 0.0042 | 5.35 ± 0.0064 | 10.00 ± 0.0028 | 7.70 ± 0.0339 | 6.20 ± 0.0113 | 10.70 ± 0.0042 | 2.80 ± 0.0014 | 12.60 ± 0.0311 | 5.25 ± 0.0035 |
| Porphyromonadaceae (Parabacteroides distasonis) | 3.50 ± 0.0042 | 0.80 ± 0.0014 | 2.45 ± 0.0021 | 0.80 ± 0.0014 | 0.65 ± 0.0007 | 2.90 ± 0.0014 | 0.20 ± 0.0000 | 0.95 ± 0.0007 | 0.25 ± 0.0007 |
| Paraprevotellaceae (Paraprevotella) | 0.35 ± 0.0021 | 0.15 ± 0.0007 | 0.55 ± 0.0007 | 0.75 ± 0.0021 | 0.70 ± 0.0000 | 0.20 ± 0.0000 | 0.15 ± 0.0007 | 1.05 ± 0.0007 | 0.30 ± 0.0000 |
| Bacillaceae (Bacillus coagulans) | 2.45 ± 0.0007 | 56.15 ± 0.0615 | 2.45 ± 0.0007 | 19.90 ± 0.0184 | 2.40 ± 0.0000 | 2.50 ± 0.0085 | 0.45 ± 0.0007 | 7.60 ± 0.0071 | 0.40 ± 0.0014 |
| Lactobacillaceae (Lactobacillus reuteri) | 1.30 ± 0.0028 | 1.45 ± 0.0049 | 1.55 ± 0.0007 | 9.80 ± 0.0113 | 1.95 ± 0.0064 | 1.95 ± 0.0120 | 0.20 ± 0.0000 | 1.00 ± 0.0042 | 0.15 ± 0.0007 |
| Turicibacteraceae (Turicibacter) | 0.95 ± 0.0007 | 0.45 ± 0.0007 | 1.30 ± 0.0014 | 0.50 ± 0.0000 | 0.90 ± 0.0014 | 0.95 ± 0.0007 | 0.30 ± 0.0000 | 0.35 ± 0.0007 | 0.40 ± 0.0000 |
| Clostridiaceae (Clostridium) | 0.90 ± 0.0170 | 1.30 ± 0.0042 | 3.40 ± 0.0028 | 14.10 ± 0.0325 | 0.70 ± 0.0014 | 4.05 ± 0.0035 | 0.35 ± 0.0007 | 17.45 ± 0.0049 | 0.45 ± 0.0007 |
| Clostridiaceae (c) | 4.80 ± 0.0007 | 0.65 ± 0.0007 | 1.80 ± 0.0014 | 0.55 ± 0.0007 | 0.70 ± 0.0028 | 1.40 ± 0.0014 | 0.20 ± 0.0000 | 0.50 ± 0.0000 | 0.25 ± 0.0007 |
| Lachnospiraceae (Blautia) | 2.15 ± 0.0028 | 8.30 ± 0.0057 | 8.65 ± 0.0092 | 3.95 ± 0.0007 | 1.65 ± 0.0007 | 6.35 ± 0.0049 | 0.90 ± 0.0014 | 4.25 ± 0.0049 | 0.75 ± 0.0007 |
| Lachnospiraceae (Coprococcus) | 4.95 ± 0.0028 | 1.15 ± 0.0035 | 2.35 ± 0.0021 | 1.20 ± 0.0014 | 1.95 ± 0.0007 | 1.55 ± 0.0007 | 0.90 ± 0.0014 | 1.55 ± 0.0007 | 1.10 ± 0.0000 |
| Lachnospiraceae (Dorea formicigenerans) | 7.70 ± 0.0042 | 2.75 ± 0.0049 | 12.75 ± 0.0021 | 3.40 ± 0.0028 | 1.50 ± 0.0014 | 10.05 ± 0.0106 | 0.70 ± 0.0014 | 4.15 ± 0.0021 | 0.75 ± 0.0007 |
| Lachnospiraceae (Lachnospira) | 2.00 ± 0.0007 | 0.50 ± 0.0014 | 0.90 ± 0.0000 | 4.50 ± 0.0028 | 0.50 ± 0.0000 | 0.95 ± 0.0007 | 0.35 ± 0.0007 | 7.75 ± 0.0318 | 0.45 ± 0.0007 |
| Lachnospiraceae (Roseburia faecis) | 13.00 ± 0.0007 | 0.05 ± 0.0007 | 0.70 ± 0.0000 | 0.30 ± 0.0014 | 1.90 ± 0.0014 | 0.50 ± 0.0000 | 0.80 ± 0.0014 | 1.00 ± 0.0042 | 1.15 ± 0.0007 |
| Ruminococcaceae | 0.95 ± 0.0085 | 0.95 ± 0.0007 | 4.65 ± 0.0021 | 1.80 ± 0.0014 | 2.05 ± 0.0007 | 3.90 ± 0.0014 | 0.70 ± 0.0014 | 2.30 ± 0.0014 | 0.80 ± 0.0014 |
| Ruminococcaceae (Fecalibacterium prausnitzii) | 0.45 ± 0.0035 | 5.55 ± 0.0049 | 6.05 ± 0.0021 | 5.10 ± 0.0057 | 7.25 ± 0.0021 | 5.20 ± 0.0099 | 1.85 ± 0.0007 | 8.40 ± 0.0071 | 1.75 ± 0.0007 |
| Ruminococcaceae (Oscillospira) | 4.70 ± 0.0042 | 0.40 ± 0.0014 | 3.55 ± 0.0007 | 0.45 ± 0.0007 | 0.90 ± 0.0000 | 3.50 ± 0.0028 | 0.35 ± 0.0007 | 0.55 ± 0.0007 | 0.35 ± 0.0007 |
| Ruminococcaceae (Ruminococcus) | 5.45 ± 0.0014 | 4.05 ± 0.0021 | 9.10 ± 0.0071 | 1.80 ± 0.0014 | 2.45 ± 0.0007 | 8.55 ± 0.0092 | 1.50 ± 0.0000 | 2.60 ± 0.0014 | 1.75 ± 0.0007 |
| Veillonellaceae (Dialister) | 4.10 ± 0.0014 | 0.85 ± 0.0007 | 1.30 ± 0.0014 | 1.35 ± 0.0035 | 1.00 ± 0.0000 | 1.30 ± 0.0028 | 0.70 ± 0.0028 | 1.50 ± 0.0014 | 0.30 ± 0.0000 |
| Veillonellaceae (Veillonella dispar) | 9.30 ± 0.0007 | 1.35 ± 0.0021 | 1.35 ± 0.0007 | 11.85 ± 0.0148 | 0.15 ± 0.0007 | 1.00 ± 0.0028 | NA | 10.55 ± 0.0134 | NA |
| Erysipelotrichaceae | 0.45 ± 0.0007 | 0.30 ± 0.0014 | 1.30 ± 0.0000 | 0.60 ± 0.0014 | 0.35 ± 0.0007 | 1.00 ± 0.0014 | 0.20 ± 0.0000 | 1.40 ± 0.0014 | 0.30 ± 0.0014 |
| Enterobacteriaceae (Gluconacetobacter) | 0.45 ± 0.0000 | NA | 0.10 ± 0.0000 | NA | 16.35 ± 0.0078 | 0.40 ± 0.0028 | 3.35 ± 0.0134 | NA | 2.65 ± 0.0106 |
| Enterobacteriaceae (Klebsiella) | 0.45 ± 0.0000 | 0.05 ± 0.0007 | 0.30 ± 0.0000 | NA | 39.55 ± 0.0064 | 1.50 ± 0.0085 | 14.35 ± 0.0559 | 0.05 ± 0.0007 | 12.95 ± 0.0530 |
| Enterobacteriaceae (Trabulsiella) | 0.45 ± 0.0099 | 0.85 ± 0.0035 | 8.20 ± 0.0028 | 0.40 ± 0.0000 | 0.80 ± 0.0014 | 16.30 ± 0.0877 | 64.95 ± 0.0658 | 0.65 ± 0.0007 | 64.30 ± 0.0622 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, M.; Watson, E.; Conlon, M.; Sanguansri, L.; Augustin, M.A. Impact of Co-Delivery of EGCG and Tuna Oil within a Broccoli Matrix on Human Gut Microbiota, Phenolic Metabolites and Short Chain Fatty Acids In Vitro. Molecules 2022, 27, 656. https://doi.org/10.3390/molecules27030656
Shi M, Watson E, Conlon M, Sanguansri L, Augustin MA. Impact of Co-Delivery of EGCG and Tuna Oil within a Broccoli Matrix on Human Gut Microbiota, Phenolic Metabolites and Short Chain Fatty Acids In Vitro. Molecules. 2022; 27(3):656. https://doi.org/10.3390/molecules27030656
Chicago/Turabian StyleShi, Meng, Emma Watson, Michael Conlon, Luz Sanguansri, and Mary Ann Augustin. 2022. "Impact of Co-Delivery of EGCG and Tuna Oil within a Broccoli Matrix on Human Gut Microbiota, Phenolic Metabolites and Short Chain Fatty Acids In Vitro" Molecules 27, no. 3: 656. https://doi.org/10.3390/molecules27030656
APA StyleShi, M., Watson, E., Conlon, M., Sanguansri, L., & Augustin, M. A. (2022). Impact of Co-Delivery of EGCG and Tuna Oil within a Broccoli Matrix on Human Gut Microbiota, Phenolic Metabolites and Short Chain Fatty Acids In Vitro. Molecules, 27(3), 656. https://doi.org/10.3390/molecules27030656