
Isolation, Identification and Pharmacological Effects of Mandragora autumnalis Fruit Flavonoids Fraction


Abstract
1. Introduction
2. Results
2.1. Chemistry
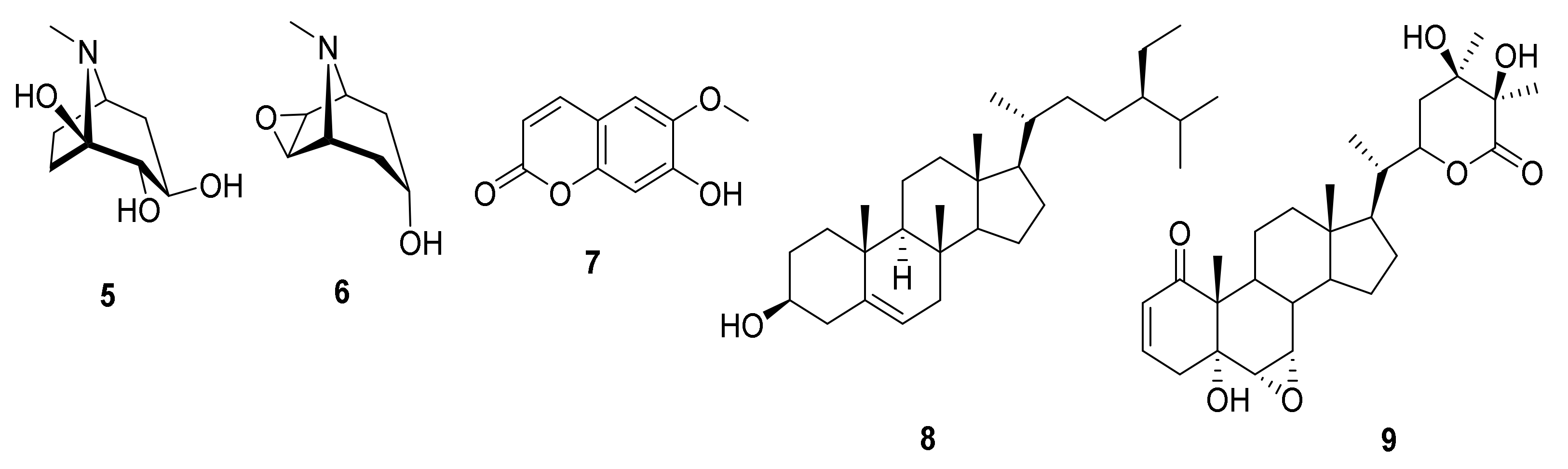
Identification of the Isolated Compounds
2.2. Biological Evaluations
2.2.1. Cytotoxicity
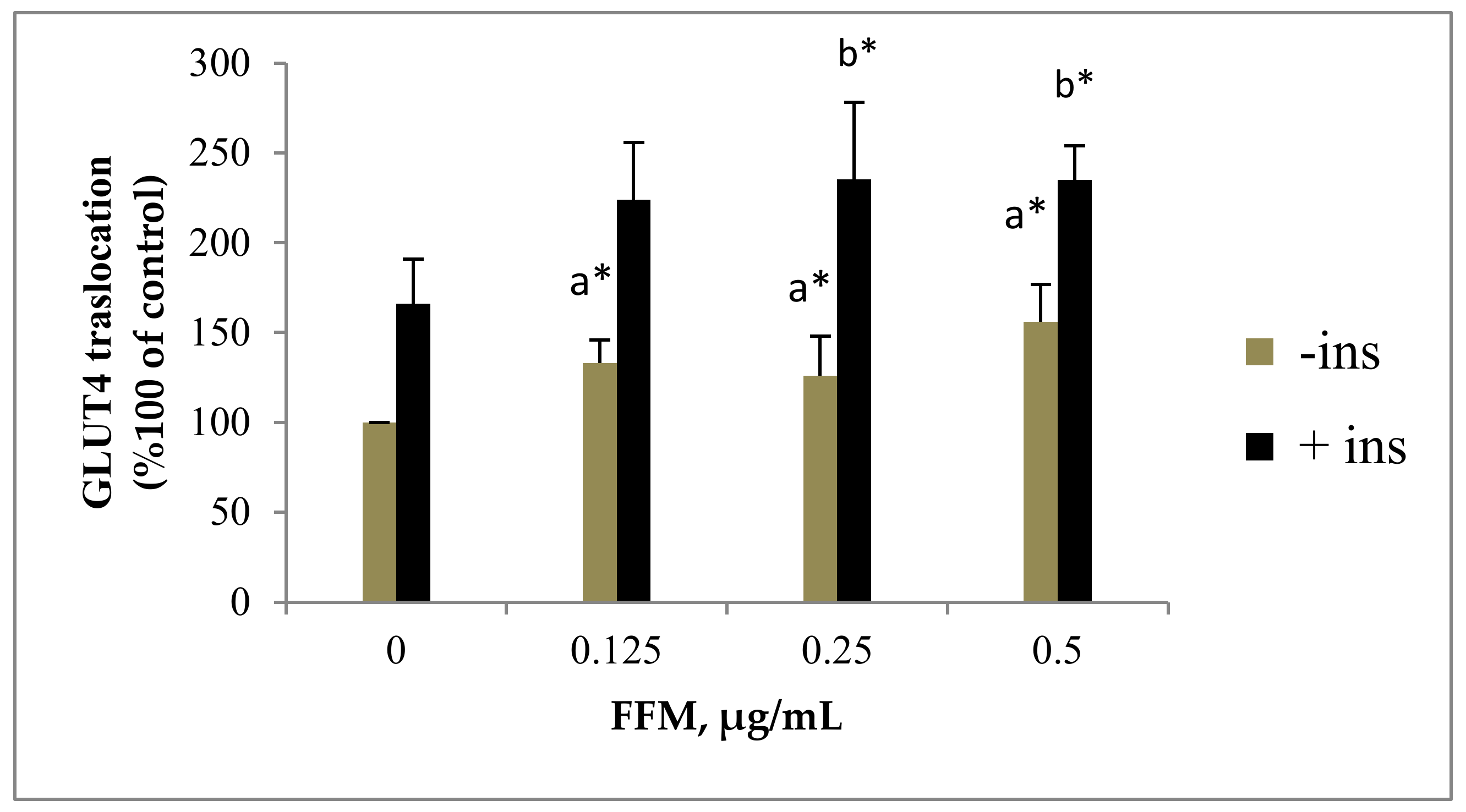
2.2.2. GLUT4 Translocation to the Plasma Membrane
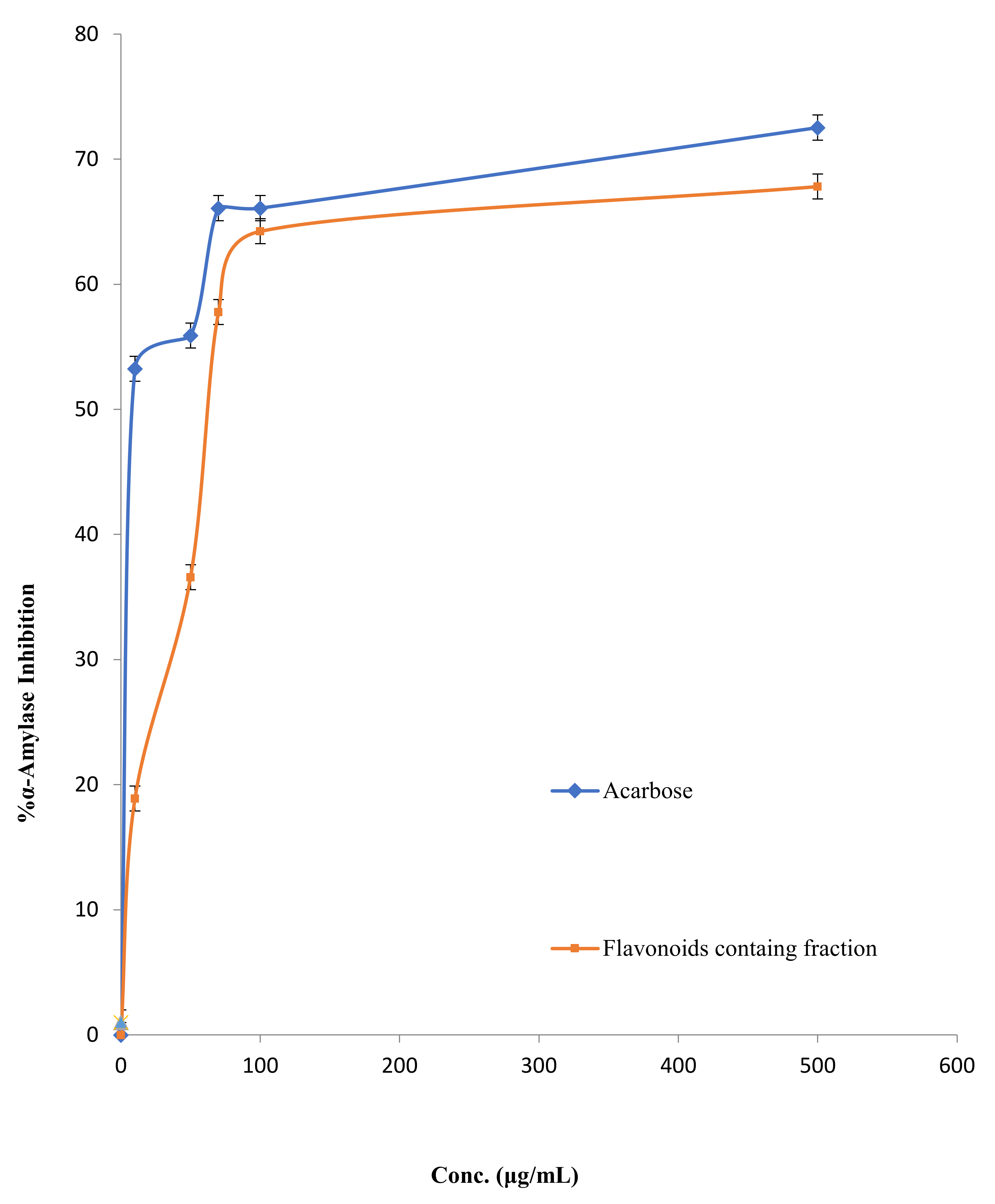
2.2.3. α-Amylase Inhibitory Potential
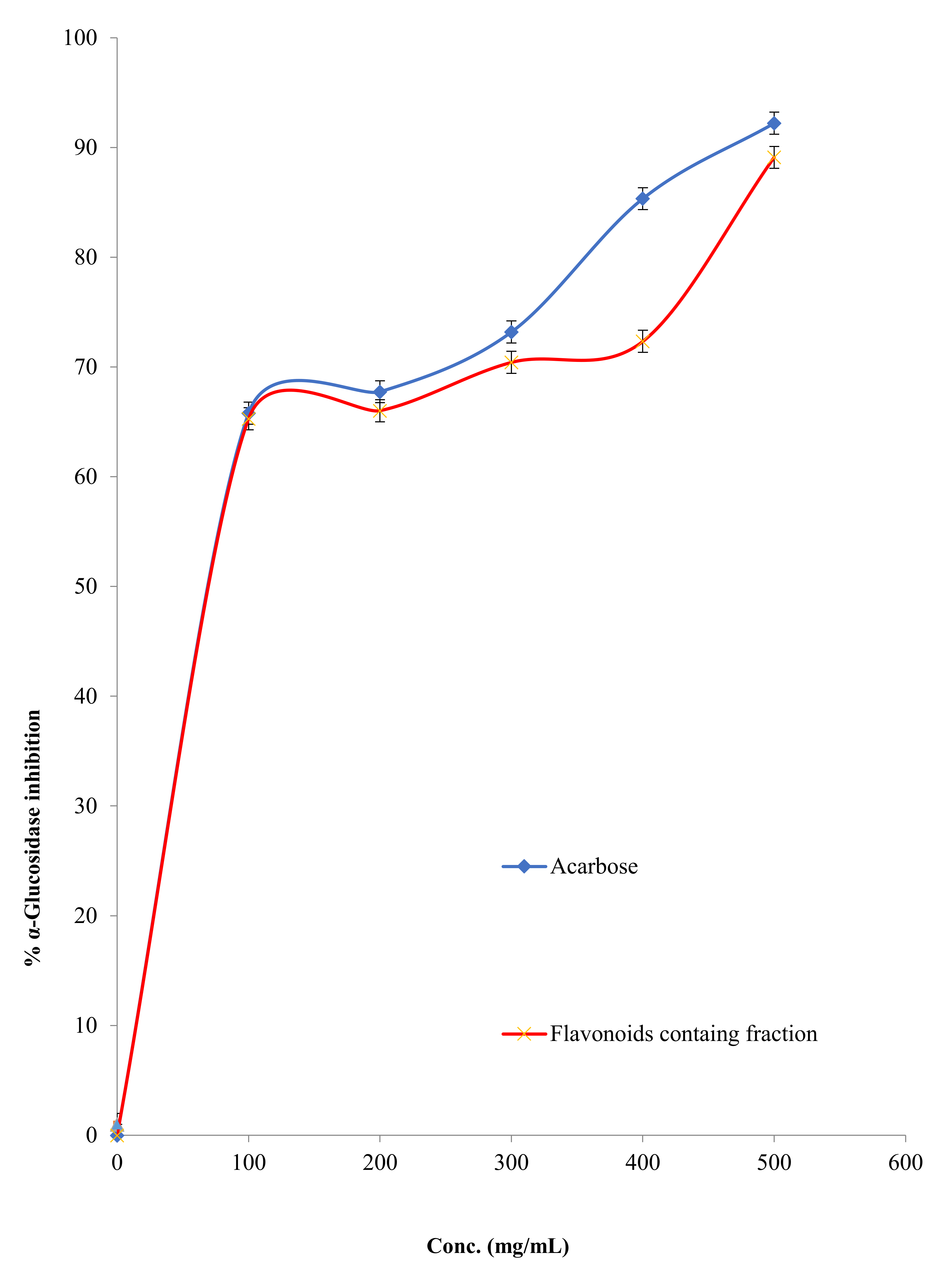
2.2.4. α-Glucosidase Inhibitory Activity
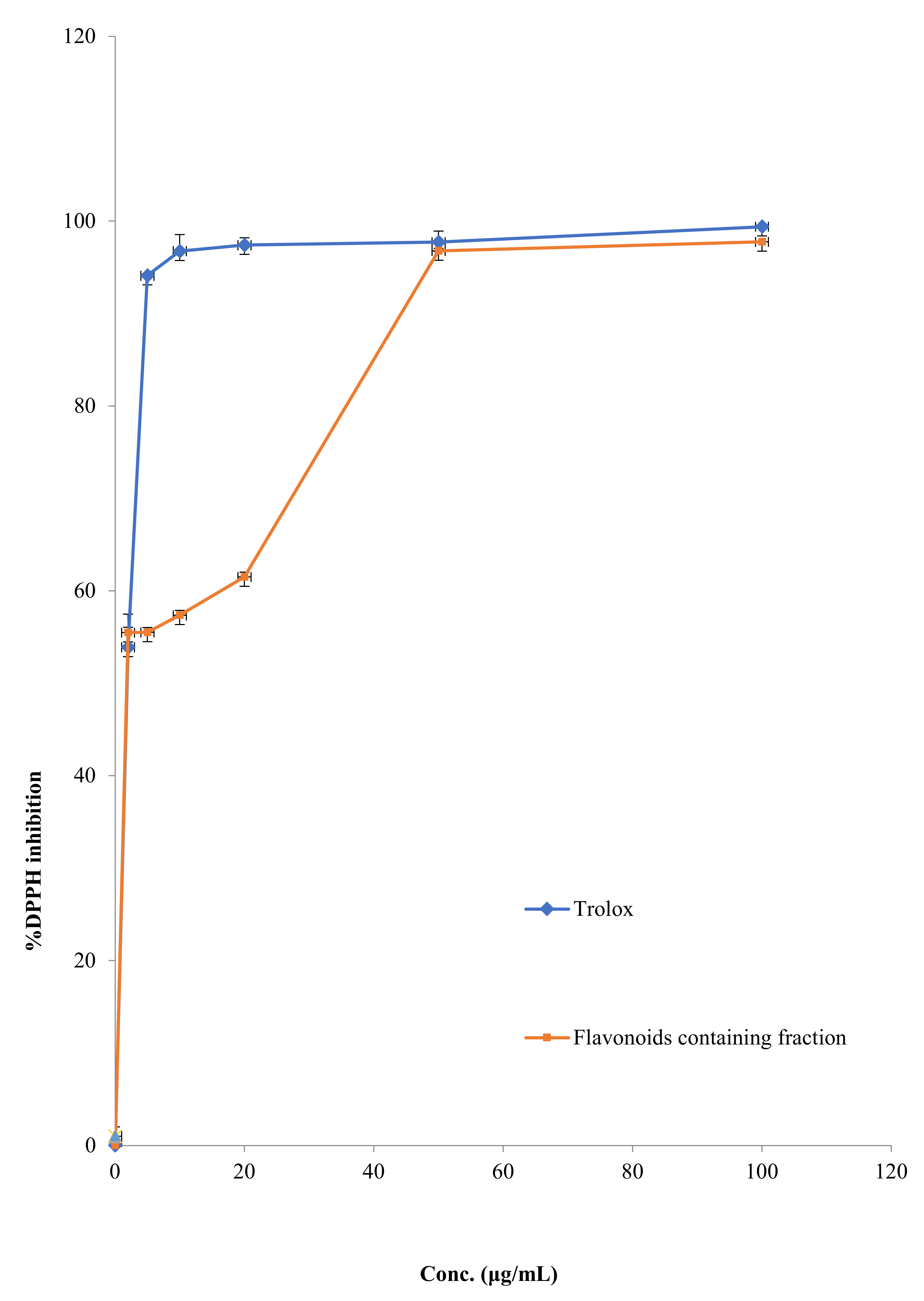
2.2.5. DPPH Scavenging Activity
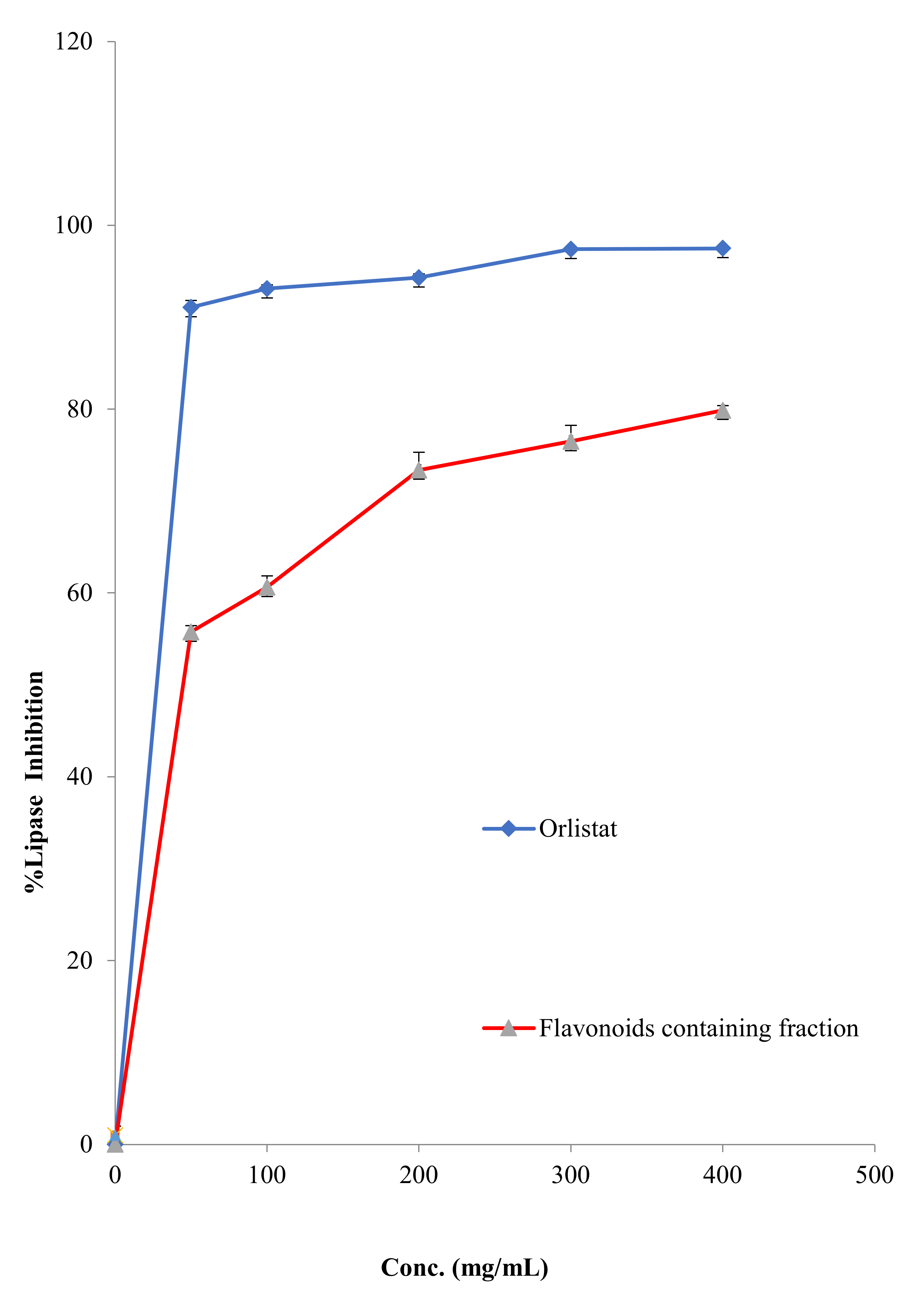
2.2.6. Antilipase Activity
2.2.7. Antimicrobial Capacity
3. Discussion
3.1. Chemical Characterization
3.2. Antidiabetic Effects
3.2.1. FFM Effects on GLUT4 Translocation
3.2.2. Carbohydrates Metabolic Enzymes
3.3. DPPH Scavenging Effect of FFM
3.4. Anti-Lipase Activity
3.5. Antimicrobial Activity
4. Materials and Methods
4.1. Chemicals, Reagents and Instruments
4.2. Plant Collection and Preparation
4.3. Extraction and Isolation of Compounds
4.4. Biological Method
4.4.1. Cell Growth and Treatment
4.4.2. Cytotoxicity
4.4.3. Determination of Surface GLUT4myc
4.4.4. α-Amylase Inhibitory Method
4.4.5. α-Glucosidase Inhibitory Activity
4.4.6. DPPH Scavenging Activity
4.4.7. Porcine Pancreatic Lipase Enzyme Inhibitory Assay
4.4.8. Antimicrobial Activity
4.5. Statistical Characterizations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Preedy, V.R.; Watson, R.R. Nuts and Seeds in Health and Disease Prevention; Academic Press: Cambridge, MI, USA, 2020. [Google Scholar]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Alamgir, A. Therapeutic Use of Medicinal Plants and Their Extract; Springer: New York, NY, USA, 2018. [Google Scholar]
- Azab, A. Solanaceae plants of Israel and Palestine-rich source of medicinally active natural products. Eur. Chem. Bull. 2020, 9, 199–261. [Google Scholar] [CrossRef]
- Alzweiri, M.; Al Sarhan, A.; Mansi, K.; Hudaib, M.; Aburjai, T. Ethnopharmacological survey of medicinal herbs in Jordan, the Northern Badia region. J. Ethnopharmacol. 2011, 137, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Mahmod, A.I.; Talib, W.H. Anticancer activity of Mandragora autumnalis: An in vitro and in vivo study. Pharmacia 2021, 4, 827–836. [Google Scholar] [CrossRef]
- Jodallah, N.B.E. Antioxidant and Antimicrobial Activity of Mandragora Autumnalis Bertol Extracts; An-Najah National University: Nablus, Palestine, 2013. [Google Scholar]
- Uysal, S.; Zengin, G.; Aktümsek, A. Antioxidant properties and enzyme inhibitory effects of extracts from Mandragora autumnalis and its fatty acid composition. Marmara Pharm. J. 2016, 20, 144–151. [Google Scholar] [CrossRef]
- Casini, E. Rethinking the Multifaceted Aspects of Mandrake in Ancient Egypt; Pisa University Press: Pisa, Italy, 2017. [Google Scholar]
- Al-Dinawari, B. Botany, Herbals and Healing in Islamic Science and Medicine; FSTC Ltd.: Manchester, UK, 2009. [Google Scholar]
- Hanuš, L.O.; Řezanka, T.; Spížek, J.; Dembitsky, V.M. Substances isolated from Mandragora species. Phytochemistry 2005, 66, 2408–2417. [Google Scholar] [CrossRef]
- Tuttolomondo, T.; Licata, M.; Leto, C.; Savo, V.; Bonsangue, G.; Gargano, M.L.; Venturella, G.; La Bella, S. Ethnobotanical investigation on wild medicinal plants in the Monti Sicani Regional Park (Sicily, Italy). J. Ethnopharmacol. 2014, 153, 568–586. [Google Scholar] [CrossRef]
- Jackson, B.P.; Berry, M.I. Mandragora-Taxonomy and Chemistry of the European Species; Academic Press: London, UK, 1979. [Google Scholar]
- Suleiman, R.K.; Zarga, M.A.; Sabri, S.S. New withanolides from Mandragora officinarum: First report of withanolides from the genus Mandragora. Fitoterapia 2010, 81, 864–868. [Google Scholar] [CrossRef]
- Suvan, J.E.; Finer, N.; D’Aiuto, F. Periodontal complications with obesity. Periodontol. 2000 2018, 78, 98–128. [Google Scholar] [CrossRef]
- Gashaw, M.; Berhane, M.; Bekele, S.; Kibru, G.; Teshager, L.; Yilma, Y.; Ahmed, Y.; Fentahun, N.; Assefa, H.; Wieser, A. Emergence of high drug resistant bacterial isolates from patients with health care associated infections at Jimma University medical center: A cross sectional study. Antimicrob. Resist. Infect. Control 2018, 7, 1–8. [Google Scholar] [CrossRef]
- Dadgostar, P. Antimicrobial resistance: Implications and costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef]
- Chanda, S.; Tiwari, R.K.; Kumar, A.; Singh, K. Nutraceuticals inspiring the current therapy for lifestyle diseases. Adv. Pharmacol. Sci. 2019, 2019, 6908716. [Google Scholar] [CrossRef]
- Lou, H.; Hu, L.; Lu, H.; Wei, T.; Chen, Q. Metabolic Engineering of Microbial Cell Factories for Biosynthesis of Flavonoids: A Review. Molecules 2021, 26, 4522. [Google Scholar] [CrossRef]
- Liu, H.; Mou, Y.; Zhao, J.; Wang, J.; Zhou, L.; Wang, M.; Wang, D.; Han, J.; Yu, Z.; Yang, F. Flavonoids from Halostachys caspica and their antimicrobial and antioxidant activities. Molecules 2010, 15, 7933–7945. [Google Scholar] [CrossRef]
- Monadi, T.; Azadbakht, M.; Ahmadi, A.; Chabra, A. A Comprehensive Review on the Ethnopharmacology, Phytochemistry, Pharmacology, and Toxicology of the Mandragora Genus; from Folk Medicine to Modern Medicine. Curr. Pharm. Des. 2021, 27, 3609–3637. [Google Scholar] [CrossRef]
- Wawer, I.; Zielinska, A. 13C CP/MAS NMR studies of flavonoids. Magn. Reson. Chem. 2001, 39, 374–380. [Google Scholar] [CrossRef]
- Lin, L.-J.; Huang, X.-B.; Lv, Z.-C. Isolation and identification of flavonoids components from Pteris vittata L. Springerplus 2016, 5, 1–3. [Google Scholar] [CrossRef]
- Shen, C.-C.; Chang, Y.-S.; Ho, L.-K. Nuclear-magnetic-resonance studies of 5, 7-dihydroxyflavonoids. Phytochemistry 1993, 34, 843–845. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Zhang, S.; Lu, W.; Liao, S.; Liu, X.; Shan, L.; Shen, X.; Jiang, H.; Zhang, W. Natural products as a gold mine for selective matrix metalloproteinases inhibitors. Bioorg. Med. Chem. 2012, 20, 4164–4171. [Google Scholar] [CrossRef]
- Sheliya, M.A.; Rayhana, B.; Ali, A.; Pillai, K.K.; Aeri, V.; Sharma, M.; Mir, S.R. Inhibition of α-glucosidase by new prenylated flavonoids from Euphorbia hirta L. herb. J. Ethnopharmacol. 2015, 176, 1–8. [Google Scholar] [CrossRef]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef]
- Echeverría, C.; Nualart, F.; Ferrada, L.; Smith, G.J.; Godoy, A.S. Hexose transporters in cancer: From multifunctionality to diagnosis and therapy. Trends Endocrinol. Metab. 2021, 32, 198–211. [Google Scholar] [CrossRef]
- AL-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef]
- Choi, H.-N.; Kang, M.-J.; Lee, S.-J.; Kim, J.-I. Ameliorative effect of myricetin on insulin resistance in mice fed a high-fat, high-sucrose diet. Nutr. Res. Pract. 2014, 8, 544–549. [Google Scholar] [CrossRef]
- Alkhalidy, H.; Moore, W.; Zhang, Y.; McMillan, R.; Wang, A.; Ali, M.; Suh, K.-S.; Zhen, W.; Cheng, Z.; Jia, Z. Small molecule kaempferol promotes insulin sensitivity and preserved pancreatic β-cell mass in middle-aged obese diabetic mice. J. Diabetes. Res. 2015, 2015, 532984. [Google Scholar] [CrossRef]
- Proença, C.; Freitas, M.; Ribeiro, D.; Tomé, S.M.; Oliveira, E.F.; Viegas, M.F.; Araújo, A.N.; Ramos, M.J.; Silva, A.M.; Fernandes, P.A. Evaluation of a flavonoids library for inhibition of pancreatic α-amylase towards a structure–activity relationship. J. Enzyme Inhib. Med. Chem. 2019, 34, 577–588. [Google Scholar] [CrossRef]
- Barber, E.; Houghton, M.J.; Williamson, G. Flavonoids as human intestinal α-glucosidase inhibitors. Foods 2021, 10, 1939. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-glucosidase and α-amylase by flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, C.; Zhang, B.; Huang, Q. The inhibitory effects of flavonoids on α-amylase and α-glucosidase. Crit. Rev. Food Sci. Nutr. 2020, 60, 695–708. [Google Scholar] [CrossRef]
- Rahman, M.M.; Islam, M.B.; Biswas, M.; Alam, A.K. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes 2015, 8, 621. [Google Scholar] [CrossRef]
- Khan, M.A.; Rahman, A.A.; Islam, S.; Khandokhar, P.; Parvin, S.; Islam, M.B.; Hossain, M.; Rashid, M.; Sadik, G.; Nasrin, S. A comparative study on the antioxidant activity of methanolic extracts from different parts of Morus alba L. (Moraceae). BMC Res. Notes 2013, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Sim, G.-S.; Lee, B.-C.; Cho, H.S.; Lee, J.W.; Kim, J.-H.; Lee, D.-H.; Kim, J.-H.; Pyo, H.-B.; Moon, D.C.; Oh, K.-W. Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharm. Res. 2007, 30, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Oussaada, S.M.; van Galen, K.A.; Cooiman, M.I.; Kleinendorst, L.; Hazebroek, E.J.; van Haelst, M.M.; ter Horst, K.W.; Serlie, M.J. The pathogenesis of obesity. Metabolism 2019, 92, 26–36. [Google Scholar] [CrossRef]
- Jaradat, N.; Zaid, A.N.; Hussein, F.; Zaqzouq, M.; Aljammal, H.; Ayesh, O. Anti-lipase potential of the organic and aqueous extracts of ten traditional edible and medicinal plants in palestine; a comparison study with orlistat. Medicines 2017, 4, 89. [Google Scholar] [CrossRef]
- Shimura, S.; Itoh, Y.; Yamashita, A.; Kitano, A.; Hatano, T.; Yoshida, T.; Okuda, T. Inhibitory effects of flavonoidson lipase. Nippon Shokuhin Kogyo Gakkaishi 1994, 41, 847–850. [Google Scholar] [CrossRef]
- Abou-El-Naga, I.; Said, D.; Gaafar, M.; Ahmed, S.; El-Deeb, S. A new scope for orlistat: Effect of approved anti-obesity drug against experimental microsporidiosis. Med. Mycol. 2019, 57, 181–195. [Google Scholar] [CrossRef]
- Obeidat, M. Antimicrobial activity of some medicinal plants against multidrug resistant skin pathogens. J. Med. Plant Res. 2011, 5, 3856–3860. [Google Scholar]
- Ayaz, F.A.; Hayırlıoglu-Ayaz, S.; Alpay-Karaoglu, S.; Grúz, J.; Valentová, K.; Ulrichová, J.; Strnad, M. Phenolic acid contents of kale (Brassica oleraceae L. var. acephala DC.) extracts and their antioxidant and antibacterial activities. Food Chem. 2008, 107, 19–25. [Google Scholar] [CrossRef]
- Rahman, M.A.A.; Moon, S.-S. Antimicrobial phenolic derivatives from Dendranthema zawadskii var. latilobum kitamura (Asteraceae). Arch. Pharm. Res. 2007, 30, 1374–1379. [Google Scholar] [CrossRef]
- Sarian, M.N.; Ahmed, Q.U.; Mat So’ad, S.Z.; Alhassan, A.M.; Murugesu, S.; Perumal, V.; Syed Mohamad, S.N.A.; Khatib, A.; Latip, J. Antioxidant and antidiabetic effects of flavonoids: A structure-activity relationship based study. Biomed. Res. Int. 2017, 2017, 8386065. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2015, 22, 132–149. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Kadan, S.; Sasson, Y.; Zaid, H. Portulaca oleracea L. extracts enhance GLUT4 translocation to the surface of muscle cells in insulin dependent and independent fashion. Adv. Med. Plant Res. 2019, 7, 85–90. [Google Scholar] [CrossRef]
- Zaid, H.; Talior-Volodarsky, I.; Antonescu, C.; Liu, Z.; Klip, A. GAPDH binds GLUT4 reciprocally to hexokinase-II and regulates glucose transport activity. Bioch. J. 2009, 419, 475–484. [Google Scholar] [CrossRef][Green Version]
- Ali, H.; Houghton, P.; Soumyanath, A. α-Amylase inhibitory activity of some Malaysian plants used to treat diabetes; with particular reference to Phyllanthus amarus. J. Ethnopharmacol. 2006, 107, 449–455. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kwon, C.-S.; Son, K.H. Inhibition of alpha-glucosidase and amylase by luteolin, a flavonoid. Biosci. Biotechnol. Biochem. 2000, 64, 2458–2461. [Google Scholar] [CrossRef]
- Jaradat, N.A.; Abualhasan, M. Comparison of phytoconstituents, total phenol contents and free radical scavenging capacities between four Arum species from Jerusalem and Bethlehem. Pharm. Sci. 2016, 22, 120–125. [Google Scholar] [CrossRef]
- Falahati, M.; Omidi Tabrizib, N.; Jahaniani, F. Anti dermatophyte activities of Eucalyptus camaldulensis in comparison with Griseofulvin. Iran. J. Pharmacol. Ther. 2005, 4, 80–100. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound 1 | Compound 2 | Compound 3 | Compound 4 | |||||
|---|---|---|---|---|---|---|---|---|
| C No | δC | δH | δC | δH | δC | δH | δC | δH |
| 1 | ||||||||
| 2 | 147.3 | 169.1 | 147.3 | 72.0 | 4.98 d, J = 11.2 Hz | |||
| 3 | 136.1 | 108.1 | 6.68 s | 136.3 | 83.5 | 4.50 d, J = 11.2, 6.2 Hz | ||
| 4 | 176.4 | 186.9 | 176.2 | 197.3 | ||||
| 4a | 103.5 | 108.9 | 103.4 | 101.0 | ||||
| 5 | 161.2 | 166.7 | 161.2 | 163.8 | ||||
| 6 | 98.7 | 6.19 d, J = 2.1 Hz | 103.9 | 6.19 d, J = 2.1 Hz | 98.6 | 6.18 d, J = 2.1 Hz | 95.4 | 5.86 d, J = 2.1 Hz |
| 7 | 164.4 | 169.3 | 164.3 | 167.3 | ||||
| 8 | 94.0 | 6.44 d, J = 2.1 Hz | 99.4 | 6.45 d, J = 2.1 Hz | 93.7 | 6.37 d, J = 2.1 Hz | 96.4 | 5.91 d, J = 2.1 Hz |
| 8a | 159.67 | 162.2 | 156.5 | 163.0 | ||||
| 1′ | 122.1 | 126.6 | 121.2 | 128.5 | ||||
| 2′ | 130.0 | 8.05 d, J = 8.9 Hz | 118.3 | 7.40 d, J = 2.2 Hz | 107.6 | 7.24 s | 115.8 | 6.88 d, J = 1.6 Hz |
| 3′ | 115.9 | 6.93 d, J = 8.9 Hz | 150.7 | 146.2 | 144.9 | |||
| 4′ | 156.6 | 154.9 | 136.3 | 145.4 | ||||
| 5′ | 115.9 | 6.93 d, J = 8.9 Hz | 120.9 | 6.89 d, J = 8.3 Hz | 146.2 | 115.6 | 6.75 m, 2 H overlap with H-6′ | |
| 6′ | 130.0 | 8.05 d, J = 8.9 Hz, 2 H | 124.1 | 7.43 dd, J = 8.3, 2.2 Hz | 107.6 | 7.24 s | 119.9 | 6.75 m, overlap with H-5′ |
| 3-OH | 10.80 | 10.79 | 5.77 d, J = 6.2 Hz | |||||
| 5-OH | 12.49 | 12.99 | 12.51 | 11.92 | ||||
| 7-OH | 10.12 | 10.85 | 9.36 | 10.85 | ||||
| 4′-OH | 9.42 | 9.95 | 8.82 | 9.05 | ||||
| 3′-OH | 9.43 | 9.23 | 9.00 | |||||
| 5′-OH | 9.23 | |||||||
| Microbial Strains | Fluconazole | Ampicillin | Ciprofloxacin | FFM |
|---|---|---|---|---|
| S. aureus | 0 | 3.12 ± 0.02 | 0.78 ± 0.01 | 1.5 ± 0.07 |
| E. faecium | 0 | 1.56 ± 0.01 | 0.78 ± 0.02 | 3.125 ± 0.03 |
| E. coli | 0 | 3.12 ± 0.23 | 1.56 ± 0.08 | 2.25 ± 0.07 |
| P. aeruginosa | 0 | 12.5 ± 0.13 | 3.12 ± 0.11 | 12.5 ± 0.98 |
| K. pneumoniae | 0 | 1 ± 0.02 | 0.13 ± 0.02 | 0.135 ± 0.01 |
| P. vulgaris | 0 | 18 ± 1.05 | 15 ± 0.35 | 22 ± 0.97 |
| MRSA | 0 | 60.5 ± 0.71 | 12.5 ± 0.54 | 25 ± 1.01 |
| E. floccosum | 0.78 ± 0.01 | 0 | 0 | 12.5 ± 0.88 |
| C. albicans | 1.56 ± 0.01 | 0 | 0 | 6.25 ± 0.48 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Maharik, N.; Jaradat, N.; Bassalat, N.; Hawash, M.; Zaid, H. Isolation, Identification and Pharmacological Effects of Mandragora autumnalis Fruit Flavonoids Fraction. Molecules 2022, 27, 1046. https://doi.org/10.3390/molecules27031046
Al-Maharik N, Jaradat N, Bassalat N, Hawash M, Zaid H. Isolation, Identification and Pharmacological Effects of Mandragora autumnalis Fruit Flavonoids Fraction. Molecules. 2022; 27(3):1046. https://doi.org/10.3390/molecules27031046
Chicago/Turabian StyleAl-Maharik, Nawaf, Nidal Jaradat, Najlaa Bassalat, Mohammed Hawash, and Hilal Zaid. 2022. "Isolation, Identification and Pharmacological Effects of Mandragora autumnalis Fruit Flavonoids Fraction" Molecules 27, no. 3: 1046. https://doi.org/10.3390/molecules27031046
APA StyleAl-Maharik, N., Jaradat, N., Bassalat, N., Hawash, M., & Zaid, H. (2022). Isolation, Identification and Pharmacological Effects of Mandragora autumnalis Fruit Flavonoids Fraction. Molecules, 27(3), 1046. https://doi.org/10.3390/molecules27031046